Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cardiovascular disease wikipedia , lookup
Heart failure wikipedia , lookup
Antihypertensive drug wikipedia , lookup
Cardiac surgery wikipedia , lookup
History of invasive and interventional cardiology wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Quantium Medical Cardiac Output wikipedia , lookup
Dextro-Transposition of the great arteries wikipedia , lookup
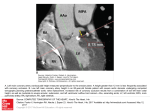
Am J Physiol Heart Circ Physiol 279: H2927–H2938, 2000. Effects of increased pressure inside or outside ventricles on total and regional myocardial blood flow G. S. ALDEA, H. MORI, W. K. HUSSEINI, R. E. AUSTIN†, AND J. I. E. HOFFMAN Cardiovascular Research Institute and the Department of Pediatrics, University of California, San Francisco, California 94143-0544 Received 24 January 2000; accepted in final form 19 July 2000 subendocardial flows; subepicardial flows; zero flow pressure, coronary venous pressure; pressure-flow curves or outside the heart have been associated with subendocardial ischemia and hemorrhage (10, 20). Investigators from Sendai, Japan, (28, 35) showed that increasing pressures outside the heart or in either ventricle in an isolated supported dog heart increased the zero flow pressure (Pzf), the coronary arterial perfusing pressure at which flow ceases. The experiments were done with maximally dilated vessels during episodes of postpacing arrest. The greatest effect was shown by increasing pericardial, or surrounding heart pressure (SHP); for increases of pericardial pressure from 0 to 15 and then to 30 mmHg, Pzf increased from 6 to 22 mmHg and then to 36 mmHg. Similar increases in right ventricular pressure (RVP) raised Pzf levels from 0 to 15 mmHg and then 30 mmHg, whereas similar increases in left ventricular pressure (LVP) increased Pzf from 6 to 12 mmHg and then to 15 mmHg. Such increases in Pzf PRESSURE ELEVATIONS INSIDE † Deceased 22 August 1998. Address for reprint requests and other correspondence: J. I. E. Hoffman, Box 0544, Univ. of California, San Francisco, CA 941430544 (E-mail: [email protected]). http://www.ajpheart.org would be expected to decrease driving pressures across the coronary circulation and so render the heart liable to global or regional ischemia. We wondered whether these different pressure changes would have different effects on flows in different layers of the left ventricular free wall, hypothesizing that the effects would be to decrease flows predominantly in the subendocardium for increases in LVP, predominantly in the subepicardium for increased pericardial pressures, and perhaps more evenly across the wall for increases in RVP. We also wondered whether the mechanism for the changes in Pzf involved the extramural venous waterfall that had been described previously by Scharf et al. (29) and Uhlig et al. (31) or whether it involved selective drop out of layers or smaller regions of the left ventricle. To examine these problems, we developed an isolated blood-perfused dog heart and repeated the experiments described above during cardioplegia, but with the addition of radioactive microspheres to measure regional flows. Because ischemia does not usually occur with adequate perfusion pressures, we concentrated on low perfusing and driving pressures. Furthermore, because we were becoming aware of the heterogeneity of myocardial blood flow, we examined not only average flow values in any myocardial layer, but in some experiments we examined the flows in several pieces of heart muscle within the myocardial layers. METHODS Preparation of the Heart The experimental protocol described was approved by the Committee on Animal Research of the University of California at San Francisco and was in compliance with the “Principles of Laboratory Animal Care” formulated by the National Society for Medical Research and the National Institutes of Health Guide for the Care and Use of Laboratory Animals [DHHS Publication No. (NIH) 80–23, Revised 1985, Office of Science and Health Reports, Bethesda, MD 20892]. We induced anesthesia in 40 healthy adult mongrel dogs weighing between 12.6 and 32.9 kg with 2 ml im Innovar followed by pentobarbital sodium (10–20 mg/kg iv) and mainThe costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 0363-6135/00 $5.00 Copyright © 2000 the American Physiological Society H2927 Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 Aldea, G. S., H. Mori, W. K. Husseini, R. E. Austin, and J. I. E. Hoffman. Effects of increased pressure inside or outside ventricles on total and regional myocardial blood flow. Am J Physiol Heart Circ Physiol 279: H2927–H2938, 2000.—Increasing pressures to 30 mmHg in right (RV) and left (LV) ventricles and surrounding heart (SH) in isolated, arrested, maximally vasodilated, blood-perfused dog hearts shifted pressure-flow (PF) curves rightward and increased zero flow pressure (Pzf) by an amount equal to the RV applied pressure, SH applied pressure, or two-thirds of the LV applied pressure. There were comparable increases in coronary venous pressures. Increasing LV or SH pressures decreased coronary blood flows, especially in the subendocardium. Decreases in driving pressure decreased flows in all layers, but even with driving pressure of 5 mmHg, a few subepicardial pieces had flow. We conclude with the following: 1) raising pressures inside or outside the heart shifts PF curves and raises Pzf by increasing coronary venous pressure; 2) the effects are most prominent in the subendocardial muscle layer; and 3) as driving pressures are decreased, there is a range of Pzf in the heart with the final Pzf recorded due to the last little piece of muscle to be perfused. H2928 VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW Fig. 1. Diagram of the preparation. Aorta (Ao); great cardiac vein (GCV); inferior vena cava (IVC); left atrium (LA); left main coronary artery (LMCA); left ventricle (LV); right atrium (RA); right ventricle (RV); superior vena cava (SVC); H2O heating for oxygenator; O2 to and from oxygenator. The dashed lines indicate intracardiac or intravascular tubing; all these tubes have only one end hole. Tubing not shown to scale. Drainage from RA to oxygenator is by gravity. was filtered through a 20-m PDF filter (Fenwal Electronics/ APD, Milford, MA) and infused with a roller pump (Masterflex R-7523, Cole-Parmer, Chicago, IL) through the Gregg cannula. For the rest of the experiment, blood was kept at 37°C. The ventricles were vented, and blood from the right atrium was returned to the oxygenator. In each dog, coronary perfusion was reestablished within 15 min after aortic cross clamping, and cardiac arrest was maintained for the rest of the experiment. The heart was then excised from the body by ligating and cutting the appropriate vessels. The right coronary artery was ligated. Large bore tubes were placed in each ventricle through the atria and connected to reservoirs that could be set at any desired pressures (Fig. 1). These tubes had bullet tips with a single draining hole. The positions of the tips of these tubes were verified by the response to fluid infusion. If the end of the tube was in the ventricle, then that ventricle was distended by the fluid. If in error, the end of the tube was in the atrium, then the atrium and not the ventricle was distended; if that occurred the tube was repositioned. The reservoirs were also raised and then returned to control level to make sure that all blood returned to the reservoir and was not lost from the heart by leakage. The stump of the superior vena cava was tied around the cannula that entered the right ventricle. The stump of the inferior vena cava was tied around a catheter that drained blood from the right atrium and the coronary sinus by gravity to the oxygenator. Fluid-filled catheters for pressure measurement were placed in the left ventricle and the coronary sinus or great Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 tained anesthesia with 1% halothane in oxygen. We ventilated the dogs’ lungs through a cuffed endotracheal tube with a Harvard pump (Harvard Apparatus, S. Natick, MA). We kept arterial blood gases and pH within the normal range by adjusting inspired oxygen concentration, minute ventilation, or giving boluses of sodium bicarbonate. A heating pad was used to keep the dogs’ body temperature close to 37°C. Bilateral thoracotomies were performed and the sternum was divided. A modified Gregg cannula (4 mm, ID 3 mm) was introduced into the aorta through a left subclavian arteriotomy and secured in place with its tip just above the aortic valve. The dogs received 10,000 units iv of heparin sodium. They also received 5,000 units iv every 30 min throughout the experiment. A bolus of dipyridamole (2 mg iv) was also given. The heart was isolated by clamping the azygos vein, inferior and superior vena cava, pulmonary hilar vessels, brachiocephalic trunk, and descending aorta. The femoral arteries were cannulated with 24-Fr cannulas. As soon as the aorta was clamped, oxygenated St. Thomas’ Hospital cardioplegic solution (24) at 4°C was infused into the proximal aortic root at 60 mmHg pressure through the Gregg cannula. Asystole always occurred within 10 s of cross clamping. Each dog was exsanguinated through the femoral vessels, and the blood was collected for coronary perfusion. After reoxygenation with a Bio-2 membrane oxygenator (Bentley Laboratories, Irvine, CA) and the addition of potassium chloride (10 meq/l) and carbochromene hydrochloride (40 mg/kg, Hoechst-Roussel Pharmaceuticals, Somerville, NJ), blood VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW Regional Flow Determinations Regional myocardial flow was measured with radioactive microspheres 15 ⫾ 3 m in diameter (means ⫾ SD) labeled with one of nine radionuclides (157Gd, 57Co, 114In, 51Cr, 113 Sn, 85Sr, 95Nb, 54Mn, and 65Zn). Microsphere aggregation was prevented by suspending the microspheres in 1% Tween 80 in saline and then shaking the suspension vigorously for 5 min just before injection. One million microspheres were injected ⬃40 cm proximal to the Gregg cannula to allow adequate mixing. At the end of the experiment, surface vessels, fat, and the right ventricular free wall were removed from the heart, and the left ventricle was opened along the left anterior descending artery. The left ventricular free wall and septum were then unrolled, weighed, flattened under a weight, and placed in 4% Formalin for 5–7 days. Then it was reweighed and sectioned into four longitudinal segments: anterior, lateral, posterior, and septal; four vertical layers: basal, upper middle, lower middle, and apical; and three layers across the wall: subendocardial, midwall, and subepicardial. Each of the 48 pieces was weighed and then the radioactivity was counted in a 3-in. NaI (Tl) well counter, the counts were recorded on a 1,024 channel pulse-height analyzer (TM Analytic, Elk Grove, IL), and the contributions of each radionuclide were separated by a least-squares method (7). Flow (ml 䡠 min⫺1 䡠 100 g⫺1) was calculated as Qt ⫽ Q LMCA ⫻ 共C t /C total兲 Wt where Qt is flow in the tissue sample (ml 䡠 min⫺1 䡠 100 g⫺1); QLMCA is the flowmeter determined flow in the LMCA (ml/min); Ct is the count for a particular radionuclide in the tissue sample (counts/min); Ctotal is the total radioactivity of that radionuclide in the heart (counts/min); and Wt is the weight of the tissue sample (g). We weighed pieces of the left ventricles of two dogs after cardioplegia and four dogs that did not receive cardioplegia. The ventricle pieces were then dehydrated in a desiccator for 48 h until the dry weight was constant. Experimental Protocol Pressure-flow relations. All pressure-flow (PF) relations were determined during a steady-state period. With Pzf inside and around the heart, PF relations were determined at several coronary arterial pressures in descending order by lowering the reservoir pressure 1–2 mmHg every 2–4 min until steady flows were reached. Pzf levels were determined by lowering coronary arterial pressures until no flow was observed, and then the coronary arterial pressures were raised and lowered again to obtain a second Pzf measurement. Regional flows. In 31 hearts, regional flows were determined by nine radioactive microspheres at selected coronary arterial perfusion pressures during one of four preload conditions: zero in and around the heart, LVP 30, RVP 30, or SHP 30 mmHg. Coronary arterial perfusion pressures were changed in 5-mmHg steps, and recordings were made after pressures and flows had been stable for 5–10 min; these included pressures lowered until Pzf ceased. The reservoirs that controlled pressures in the ventricles were raised or lowered slowly to the desired pressures. Pressure around the heart was changed by admitting air into the box and recorded from a strain gauge in the box. At selected coronary arterial pressures, microspheres were injected to determine regional flows. Because we had only nine types of microsphere labels, various combinations of coronary arterial pressures with pressures in or around the heart were used in different hearts. In four hearts, comparisons were made in the same heart of different preloads (0, LVP ⫽ 30, and SHP ⫽ 30 mmHg) at three different low coronary arterial pressures. In other hearts, pressures were raised in either ventricle or around the heart in increments of 5 or 10 mmHg up to a maximum of 30 mmHg, and regional PF relations were determined at different perfusion pressures. Constant coronary arterial perfusing pressures and flows. Because the heart was quiescent and the vessels were maximally dilated, increases in preloads decreased flows, often markedly. For fixed resistances, the upstream and downstream pressures will vary with flows, so we determined what would happen to arterial and venous pressures with changes in preloads at constant pressures or flows. Constant coronary arterial perfusing pressures. In one heart we raised LVP in 5-mmHg increments from 0 to 30 mmHg while we kept coronary arterial perfusion pressure constant at each of the five pressures. In the same heart we repeated the measurements at four different coronary arterial perfusion pressures while raising surrounding heart pressure by 5-mmHg increments from 0 to 30 mmHg. In another heart we changed LVP and then RVP at four different coronary arterial perfusion pressures. In a third heart we changed RVP at a fixed coronary arterial perfusion pressure. Constant flows. In five hearts we changed the preparation to keep flows constant at three or four rates: 25, 50, 75, and 100 ml/min. In three other hearts we kept flows constant at one low rate (28.5–33 ml/min) and at one high rate (80–87 ml/min). In two hearts we varied RVP, in four hearts we varied the surrounding heart pressure, and in five hearts we varied the LVP; all of these pressures were changed in 5-mmHg increments from 0 to 30 mmHg. For any intervention, the other two pressures were kept at zero. In three of these hearts, two different pressures were varied one at a Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 cardiac vein. The Gregg cannula was then advanced just beyond the orifice of the left main coronary artery (LMCA) and secured in place with a silk suture. Adenosine infusion at a rate of 20 g 䡠 kg body wt⫺1 䡠 min⫺1 was begun. After the first few experiments, to ensure maximal vasodilatation, we added 40 mg/kg chromonar hydrochloride to the reservoir and then doubled the rate of adenosine infusion until flow did not increase further. All subsequent experiments were done with this combined vasodilator protocol. LMCA pressure was measured at the tip of the Gregg cannula via an internally placed stainless steel tube. All pressure lines were connected to Statham D23 Db pressure transducers (Spectra-Med, Critical Care Division, Oxnard, CA), and zero level was set at the coronary sinus ostium. LMCA flow was measured by an in-line electromagnetic flow transducer (Howell Instruments, Camarillo, CA) attached to a Narcomatic flowmeter. The transducer was placed just proximal to the Gregg cannula, and Pzf was checked frequently by transient occlusion of the circuit. To avoid perfusion disturbances when checking Pzf, the flow transducer was in one arm of a bypass; during flow recording, the other arm of the bypass was occluded, and during Pzf testing, the bypass was opened and the tube clamped proximal to the flow transducer. The flowmeter was calibrated by timed collection of blood in a measuring cylinder. Once the perfusion had been established, the isolated perfused heart was transferred to an airtight Plexiglas box. It was suspended from the lid of the box with an aortic cross clamp, and the various tubes were brought out through ports. We were careful to make sure that the box remained airtight. Figure 1 shows the preparation. H2929 H2930 VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW time. Two other hearts, in which changes in the left heart and surrounding pressure were studied, had abnormally high coronary vascular resistances and were excluded from further analysis. Ventricular volumes. In two hearts, we measured left ventricular volume during changes of LVP, SHP, or both simultaneously at the end of the experiment. The left ventricular volume was measured by adding saline from a reservoir through the left ventricular tube. Volume changes due to altering pressures in or around the heart were then measured from the height of fluid in the cannula. Coronary vascular resistances. These were calculated by dividing the flow rate (ml/min) into the pressure difference (mmHg) between coronary arterial pressure and pressure in the coronary sinus or great cardiac vein. This difference is also called the driving pressure. Regression analysis was done with Statview version 4.5 and SuperANOVA version 1.11. Statistical significance was set at 0.05. We used linear and quadratic regression analyses and analysis of covariance. RESULTS Global PF Relations Figure 2 shows typical PF relations in the arrested heart with maximally dilated vessels when pressures are zero in and around the ventricles and when pressures are raised to 30 mmHg in the right ventricle (Fig. 2, top) and around the heart or in the left ventricle (Fig. 2, bottom). When pressures were raised, the PF curves showed parallel shifts to the right, the change being always more marked when pressure was raised outside the heart. Intermediate increases in pressure had intermediate effects. The effect of raising these pressures on Pzf is shown in Table 1; duplicate determinations of Pzf agreed closely. A 30-mmHg increase in pressure around the heart increased Pzf from a mean of 8.6 to 41.3 mmHg, the same increase in the left ventricle increased Pzf from a mean of 8.6 to 23.8 mmHg, and the same increase in the right ventricle increased Pzf from a mean of 8.6 to 32.5 mmHg. Pzf varied in the control states, that is, when no external pressures were applied, and the differences appeared to be related to the different flow rates. Venous Pressures Pressures in the great coronary vein or coronary sinus behaved identically and will be referred to hereafter as coronary venous pressures. The effect of increasing pressures in or around the heart on coronary venous pressures is complex. On the one hand, venous pressures increased, but in addition coronary arterial pressures increased, coronary flows decreased, or both of these occurred simultaneously, and the final consequences for venous pressures depend on interactions between all these events. To clarify the effects, we conducted experiments with both controlled coronary arterial pressures and controlled flows. Fig. 2. Top: representative pressure-flow (PF) curves made with 0 pressures in and around the heart (0/0/0) and with a pressure of 30 mmHg in the RV (0/30/0). Bottom: representative PF curves made with 0 pressures in and around the heart (0/0/0), a pressure of 30 mmHg around the heart (0/0/30), and a pressure of 30 mmHg in the LV (30/0/0). Increases of pressure above 0 produce an almost parallel shift to the right. L, LV pressure; R, RV pressure; S, surrounding heart pressure. Controlled Coronary Arterial Pressure We studied controlled coronary pressure in three hearts (Fig. 3). As expected, the higher coronary arterial pressures were associated with the higher flows. With Pzf in and around the heart, coronary venous pressures were 5–10 mmHg higher at the highest than the lowest coronary arterial pressures (and flows). As pressures in or around the heart were raised in 5-mmHg increments, flows decreased at all coronary arterial pressures, the rate of decrease being greatest for the highest flows. When RVP was increased in 5-mmHg increments, there was initially no or little increase in coronary sinus pressure in one of two hearts. At higher RVP, the increase in coronary sinus pressure became greater until in both hearts it increased with a slope of 1 to reach the same pressure as that in the right ventricle. Increments in surrounding heart pressure tended to be accompanied by similar increments in pressure in the great cardiac vein. When LVP was raised from 0 to 5 mmHg, coronary venous pressure decreased slightly; after that it increased steadily up to LVP of 30 mmHg. The rate of increase was similar at different coronary arterial pressures, but the maximal pressure Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 Analysis of Results VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW Table 1. Pzf under different conditions Statistic Control Plv30 Prv30 Psh30 n Maximum 75% Mean Median 25% Minimum 18 18.0 9.0 8.6 8.0 7.5 1.0 12 38.5 29.8 24.4 23.8 16.5 14.0 2 35.0 35.0 32.5 32.5 30.0 30.0 9 50.0 49.0 41.3 41.0 37.5 26.0 Values are means ⫾ SE; n ⫽ number of dogs. Plv30, left ventricular pressure of 30 mmHg; Prv30, right ventricular pressure of 30 mmHg; Psh30, surrounding heart pressure of 30 mmHg; and Pzf, zero flow pressure. Controlled Flow Rates Changing RVP. In two hearts, RVP was increased from 0 to 30 mmHg at each of four different constant flow rates between 25 and 100 ml/min (Fig. 4, left). When RVP was 0, the coronary venous pressures were 8 and 13 mmHg at a flow rate of 25 ml/min and increased to 11 and 32 mmHg, respectively, at a flow rate of 100 ml/min. When RVP was increased from 5 to 15 mmHg, the pressure in the great cardiac vein rose ⬍4 mmHg, and then at higher RVP the venous pressure of the venous pressure-imposed pressure relation increased at a slope near 1 in six of eight studies; the RVP at which venous pressure increased seemed to be greater at the higher flow rates. By contrast, all of the coronary arterial pressures increased almost linearly as imposed pressures increased, with slopes that were independent of flow rates. All of the coronary arterial slopes were ⬍1, significantly so in six of eight. The pressure differences from the coronary artery to the great cardiac vein were relatively constant, with a tendency to be highest at higher RVP. Changes in coronary vascular resistance showed similar patterns and, as expected, decreased with increasing flow rates. Changing surrounding heart pressure. In four hearts, as surrounding pressures were increased from 0 to 30 mmHg at different flow rates, there were parallel increases in venous pressures with no or minimal interaction between flow rates and pressures (Fig. 4, center). Six of the slopes of the venous pressureimposed pressure relations or their differences were linear, and three slopes had a significant but minimal Fig. 3. Coronary inflow flow rates (top) and pressure in the coronary sinus or great cardiac vein (bottom) with perfusion pressures fixed at different levels, as denoted by the symbols. Numbers indicate the individual hearts. In each panel, flows and pressures are recorded at 5-mmHg increments in RV pressure (Prv), surrounding heart pressure (Psh), or LV pressure (Plv). Increases of pressure in or around the heart decrease coronary flows and increase coronary venous pressures. When pressures are increased to 30 mmHg around the heart or in the RV, the venous pressures consistently reach 30 mmHg. When pressure in the LV is increased to 30 mmHg, the venous pressure consistently reaches only 20 mmHg. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 achieved in the coronary sinus was never greater than ⬃60% of that in the left ventricle, even at the highest flows and coronary arterial pressures. H2931 H2932 VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW quadratic component. None of the nine individual slopes was significantly different from 1. Coronary arterial pressures rose in parallel at each flow rate as SHP were increased at constant flow rates, and there was no interaction between flow and slope. Quadratic components were small, usually positive, and significant in only three of nine studies. Three of the nine slopes of the arterial-imposed pressure relations were slightly but significantly ⬎1. Pressure differences between the coronary artery and vein were almost constant at all surrounding heart pressures for any given flow rate. Coronary vascular resistances tended to increase slightly but not significantly at the higher surrounding heart pressures. Changing LVP. In six studies in five hearts, as LVP was increased from 0 to 30 mmHg, cardiac vein pressures increased roughly linearly, with average venous pressure-imposed pressure slopes well below 1 (Fig. 4, right) Coronary arterial pressures increased roughly linearly with increasing LVP; the slopes of the coronary arterial-imposed pressure relations were all significantly below 1. The pressure differences from the coronary artery to vein tended to remain constant. In the control state, the resistance was lowest at the highest flows and approximately doubled at the lowest flows. The steady-state vascular resistances at each flow rate remained constant or increased slightly as pressures were increased in the left ventricle. Ventricular volumes. Changing LVP in one heart from 10 to 30 mmHg increased left ventricular volume from 20 to 65 ml. The SHP was increased incrementally in that heart and one other to 25 and 30 mmHg when LVP was high (28 and 30 mmHg, respec- tively); there was an incremental decrease in ventricular volume to near control values (Fig. 5). Regional PF Relations In four hearts we examined the effects of raising LVP or SHP on flow to the left ventricular free wall (Fig. 6A). Driving pressure (coronary arterial minus venous pressure) was set at 30, 20, and 10 mmHg. Without any pressure in or around the heart, total coronary flow (measured in ml 䡠 min⫺1 䡠 g⫺1) decreased from 1.5 to 0.85 to 0.4 as driving pressure decreased. When LVP was raised to 30 mmHg, the flows at each driving pressure were ⬃40–50% of control flows, and almost identical Fig. 5. LV volume as Plv was increased in 5-mmHg increments (dotted line) or as SHP was increased in 5-mmHg increments after the ventricle had been distended by pressures of 30 mmHg (F) or 28 mmHg (䊐). Note the large changes in volume caused by these imposed pressures. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 Fig. 4. Representative examples of changes in coronary arterial and venous pressures and their differences at various fixed perfusion rates, as indicated by the symbols in the key, at 5-mmHg increments in pressure in the RV (left), around the heart (middle), and in the LV (right). Top shows the coronary arterial (inflow) pressures with solid symbols connected by solid lines, and the coronary venous (outflow) pressures with open symbols connected by the dashed lines. Bottom shows the arteriovenous pressure differences at each flow rate. Numbers indicate the individual hearts. Increases in pressure in or around the ventricles increase both coronary arterial and venous pressures in approximately linear fashion to similar extents, so that the arteriovenous differences remain almost constant. VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW H2933 Fig. 6. A: coronary flow at driving pressures of 10, 20, or 30 mmHg in control state (solid bars), when Plv was 30 mmHg (open bars), and when Psh was 30 mmHg (hatched bars). B: inner-toouter flow ratio per gram at driving pressures of 10, 20, or 30 mmHg in control state (solid bars), when LVP was 30 mmHg (open bars), and when SHP was 30 mmHg (hatched bars). n ⫽ 4 for each measurement. Vertical lines represent standard deviations. Increasing pressures in the LV or around the heart decreased coronary flows and inner-to-outer flow ratios per gram to similar extents. each layer. The heterogeneity of flow in these hearts when pressures in and around the heart were zero has already been reported (1). In another heart, at driving pressures of 30, 20, and 10 mmHg, we examined local flows per minute per gram when all pressures in and around the heart were zero, and again when LVP was raised to 30 mmHg. As expected, flows were heterogeneous in all layers. At 30 mmHg driving pressure, mean flows were higher in the subendocardium than the subepicardium when LVP was zero (Fig. 8) and decreased to similar low flow rates as driving pressure was reduced to 10 mmHg. When LVP was raised to 30 mmHg at a driving pressure of 30 mmHg, flows decreased more in the subendocardium than in the subepicardium. The effect of raising LVP was almost equivalent to decreasing the driving pressure by ⬎10 mmHg. At a driving pressure of 20 mmHg, not only were the flow rates lower in both layers, but also the degree of heterogeneity was reduced. All pieces of muscle, however, had detectable flows. Only at a driving pressure of 10 mmHg was flow indistinguishable from zero in all the subendocardial pieces and in most but not all of the subepicardial pieces. The dry weight of the left ventricle was unaffected by cardioplegia. DISCUSSION Fig. 7. Representative examples of flow (ml 䡠 min⫺1 䡠 100 g⫺1) in subepicardium (epi) and subendocardium (endo) for 10-mmHg increments in LV pressure (left) or around the heart (right). For both imposed pressures, changes were more marked in the subendocardium. We have confirmed the findings of others (28, 35) that changes of pressure in or around the arrested heart with maximally dilated vessels shift the PF curves to the right and increase Pzf. The relationship of the changes in Pzf to the changes in applied pressure was also confirmed. We also confirmed the relative decrease in subendocardial blood flow when, in hearts with maximally dilated vessels, left ventricular diastolic pressure was increased (17). The new findings in our study are as follows. First, increases in pressure in the ventricles or around the heart are accompanied by increases in coronary venous pressure. The increases Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 values were found when surrounding pressure was raised to 30 mmHg. When in the same hearts the flows were examined in the three layers of the left ventricular free wall, with no pressures loading the heart, the inner:outer flow ratio per gram was just below 1.5 at driving pressures of 30 and 20 mmHg and 1.0 at a driving pressure of 10 mmHg (Fig. 6B). When LVP was raised to 30 mmHg, the inner-to-outer ratio decreased to 1, 0.6, and 0.5, respectively. When surrounding heart pressure was increased to 30 mmHg, the innerto-outer ratios were ⬃0.8, 0.7, and 0.6, respectively. These changes in regional flows were due mainly to changes in subendocardial flows. If driving pressure was held at 20 mmHg and LVP or SHP was raised to 10, 20, and 30 mmHg, subepicardial flow changed little, but subendocardial flow decreased profoundly (Fig. 7). We also examined the heterogeneity of flow in the six hearts in which flows were measured in 16 pieces in H2934 VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW in venous pressure matched the increases in pressure applied around the heart or in the right ventricle. The increase in venous pressure when pressure was raised in the left ventricle was about two-thirds of the applied pressure. Second, Pzf is not uniform across the heart wall or in any given layer of the left ventricular wall. It tends to be higher in the subendocardial than the subepicardial muscle layer. Third, increased pressures both inside the left ventricle and outside the heart cause a decrease in myocardial blood flow that is most marked in the subendocardium. Fourth, there were greater differences between the arterial than the venous pressures as flows changed. These differences probably reflected the greater resistance to flow in the microvascular bed than in the larger extramural veins. Critique of Methods We found this technique difficult. Several experiments had to be discarded because of leaks in the box or the heart or angulation of a cannula. Occasionally, the cardioplegia failed, and the heart began to beat. If this happened, we gave more cardioplegic solution and repeated the experiment. The data from two hearts were discarded because the coronary vascular resistances were abnormally high; we found no specific reasons for these abnormalities. Even in the acceptable experiments, there were large differences in coronary vascular resistance that could not be explained entirely by differences in left ventricular mass. One possibility is that the length of the muscle fibers was altered by prior distention (strain softening) (18) so that measurements made without prior distention could differ from those made after the heart had been distended. Nevertheless, the pattern of response from heart to heart was consistent despite the changes in basal vascular resistance. The arrested heart is not an ideal model for a beating heart in several respects. We doubt whether the effects of raising pressures around the heart would be much affected by beating. On the other hand, ventricular contraction might well affect coronary venous drainage and certainly increases coronary venous pressure because coronary venous flow is largely systolic (2, 23). Nevertheless, in at least one study (28), the effect of a raised left ventricular diastolic pressure in the beating heart was quantitatively and qualitatively similar to that reported in the present study. We note too that these hearts had maximally dilated vessels, so that vascular tone and metabolic vascular regulatory effects did not vary as they would in most states in the beating heart. On the other hand, loss of coronary flow reserve in a beating heart will produce loss of vascular tone and metabolic regulation (12), so that our findings might be applicable to such regions. Despite its disadvantages, working on a heart arrested by cardioplegia has the advantage of prolonged diastolic arrest without ischemia. This allowed us to make careful repeated measurements and to attempt to separate confounding mechanisms. Effects on Venous Pressures Increasing pressures around the heart above atmospheric compresses the extramural coronary veins that will then act as vascular waterfalls (21, 29, 31) and regulate venous pressures at the level of the pressure outside them. The fact that coronary venous pressures rose linearly and to the same extent with pressures around the heart explains the increase in Pzf, the Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 Fig. 8. Column graph shows the numbers of pieces (vertical y-axis) with each flow rate/gram/minute (x-axis) for LV pressures of 0 or 30 mmHg and driving pressures across the coronary circulation of 30, 20, and 10 mmHg; these pressures are shown on the z- axis. Thus 30/20 indicates that driving pressure was 20 mmHg and LV pressure was 30 mmHg. Data taken from 1 heart. For discussion, see text. A: subepicardial layer. B: subendocardial layer. VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW compresses them between the myocardium and the connective tissue that surrounds the vessels. The initial negligible response of venous pressure up to LVP of 10 mmHg is probably related to the normal vascular waterfall pressure in those veins, but after this pressure they become collapsible vessels with a vascular waterfall dependent on the surrounding pressures. The fact that at higher ventricular pressures the venous pressure is about one- to two-thirds of that in the ventricle may be due to dissipation of some of the LVP across the wall of the ventricle, especially at the subepicardium. The effects of raising LVP on venous pressures were similar to those on Pzf. Other investigators have found similar effects on Pzf when preload was increased. Ellis and Klocke (17) found that, after abolishing coronary autoregulation, raising left atrial pressure from 6 to 20 mmHg increased Pzf from 12 to 19 mmHg. The same group later used a capacitance-free method to assess Pzf in long diastoles in a vasodilated beating heart; increasing preload from 6 to 10 mmHg to 31–35 mmHg increased Pzf from 14 to 23 mmHg (6). Duncker et al. (15) observed that the higher Pzf in dogs with left ventricular hypertrophy (24 vs. 12 mmHg in controls) was not only associated with a higher left ventricular end-diastolic pressure (14 vs. 6 mmHg) but also noted that increasing left ventricular diastolic pressure from 6 to 16 mmHg in the control dogs by infusing blood increased Pzf in them from 14 to 23 mmHg. Finally, the same group showed that the higher Pzf in dogs with chronic left ventricular hypertrophy (26 vs. 13 mmHg in controls) was associated with a greater left ventricular diastolic pressure (22 mmHg in hypertrophy vs. 9 mmHg in controls) (16). With exercise, Pzf went from 26 to 41 mmHg in dogs with hypertrophy, and in controls Pzf went from 13 to 24 mmHg. At the same time, left ventricular enddiastolic pressure rose with exercise from 22 to 39 mmHg with hypertrophy and from 9 to 14 mmHg in controls. In both the studies with left ventricular hypertrophy, the increments of Pzf equaled or exceeded the increments in left ventricular diastolic pressure; whether this is a specific feature of hypertrophy is not known. Thus the relationship of increasing left ventricular diastolic pressure to Pzf appears to be similar in beating and nonbeating hearts. Whether the relationship is achieved by an increased coronary venous pressure in beating hearts remains to be determined. Some caution is needed in interpreting these results as due entirely to the interaction between left ventricular distension and coronary venous pressures. Duncker et al. (14) observed in exercising dogs with maximal coronary vasodilatation that, although left ventricular end-diastolic pressure increased from 5.5 to 10.6 mmHg, measured Pzf increased from 12.6 to 23.3 mmHg, that is, by more than the increase in end-diastolic pressure. When the exercise-mediated increments in heart rate, contractility, and ␣-adrenergic vasoconstriction were prevented, the increases in Pzf and end-diastolic pressure were then equivalent. Whether the effects of heart rate, contractility, and Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 parallel shift of the PF curves, the decrease in coronary flows at fixed perfusing pressures, and the increases in coronary arterial pressures at fixed coronary flows. Increasing RVP should have no effect on coronary venous pressures until the RVP exceeds the normal vascular waterfall pressures of ⬃10–12 mmHg (31). After that pressure, the venous pressure and the RVP increased at the same rate with a slope of 1 in two studies with fixed flows and one study with fixed perfusion pressures. At the highest RVP of 30 mmHg, coronary venous pressures were equal to or close to 30 mmHg. Pzf also increased to a similar extent at the highest RVP. How does an increased RVP affect coronary venous pressures? Stretching the right ventricle will stretch the veins on its surface and even impede direct septal venous drainage into the right ventricle but is unlikely to have direct mechanical effects on veins over the left ventricle. These veins constitute most of the left coronary arterial venous drainage, and they and some right ventricular venous drainage return blood via the coronary sinus to the right atrium. In the experiments done by Mafuyama et al. (28, 35), the tube inserted into the right ventricle had multiple holes in it and was tied into the right atrium at its entry. Therefore, changes in right ventricular and atrial pressures were identical, and the shift of Pzf was probably due to increases in coronary venous pressures due to the raised right atrial pressures once venous waterfall pressures had been exceeded. We did not think that this mechanism explained our results, because in all our experiments the right atrium was undistended because of the gravity drainage. Had right atrial pressures increased, distension of the right atrium would have been obvious. Furthermore, the maintenance of different steady RVP from the reservoir required the reservoir-right ventricular system to be a closed system. Had there been significant tricuspid incompetence, blood would have leaked out into the right atrium and the oxygenator, thereby depleting the reservoir and preventing the RVP from remaining steady. These changes were not seen. Therefore, we considered other possible mechanisms for the coronary venous pressure changes that followed an increase in RVP. Direct pressure on or obstruction of the ostium and the first part of the coronary sinus seems unlikely because there is no anatomic connection between the coronary sinus and the right ventricle. There is no doubt that there would have been massive septal shift, but whether this could have changed coronary pressures is uncertain. We are left without a clear explanation of the rise in coronary venous pressure unless it was due to an unappreciated and unexplained rise in right atrial pressure. Increasing LVP to 5 or 10 mmHg had no or little effect on coronary venous pressures or even decreased them slightly; at higher LVP the coronary venous pressures rose roughly linearly but achieved only ⬃33–84% of the pressures in the ventricle. We believe that the mechanism of the rise in coronary venous pressure is related to the distention of the left ventricle that stretches the veins on the surface of the heart and H2935 H2936 VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW Effects on Arterial Pressures Coronary arterial pressures needed to maintain a constant flow increased linearly or almost linearly with all changes of pressures in or around the heart. When pressures were increased in the right ventricle, coronary arterial pressures rose substantially even when venous pressures had initially risen very little. The initial rise in coronary arterial pressure was therefore not due to the change in venous pressure but probably reflects an increase in septal vascular resistance as the right ventricular volume and pressure increased. When pressures were increased in the left ventricle, a similar finding was noted in some dogs, and perhaps in these the increased coronary arterial pressure was due to compression of the left ventricular subendocardium. Slopes of Coronary Arterial Pressure Imposed pressure relations were above 1 and were greater than the slopes of the venous pressures when pressures were changed around the heart. Increasing pressures around the heart decreases left ventricular volume and probably increases coronary vascular resistance, so that coronary arterial pressures were affected both by the raised venous pressures and the increased vascular resistance. For increased RVP, the coronary arterial slopes were below 1, even when at the higher pressures the venous slopes were 1. This probably reflects the effects of venous distention in distending microvessels and reducing coronary vascular resistance without a counterbalancing increase in resistance in the left ventricular free wall. For increased LVP, the coronary arterial slopes were well below 1 and usually greater than the venous pressure slopes. We wonder whether this reflects a balance between lowering vascular resistance by venous and ventricular distention (remembering that the empty ventricle is abnormally small) and raising vascular resistance by compression of the wall, particularly the subendocardium. Effects on Regional Flows As expected in a maximally vasodilated coronary vascular bed, subendocardial flows were higher than subepicardial flows until driving pressures were very low. Increasing LVP at a fixed coronary arterial pressure had little effect on subepicardial flows but reduced subendocardial flows in proportion to the increase in LVP. We interpret this to mean that the increased LVP or the accompanying distention of the left ventricle compressed or stretched (or both) subendocardial more than subepicardial vessels. Inasmuch as high LVP increased the volume of the left ventricle considerably, the stretching of the subepicardial vessels had little effect on the flow through them; this speaks for radial compression of the subendocardial vessels as the main reason for the decreased flows in the inner layer, or else for a marked difference in the length of the subepicardial and subendocardial vessels in response to increase in ventricular volume. We were surprised to find similar effects after increases of surrounding heart pressure; we had hypothesized that inwardly directed radial compression would have had its greatest effect on subepicardial vessels. The greater effect on subendocardial vessels might have been due to the considerable decrease in left ventricular volume caused by an increase in pressure around the heart. Such a decrease in volume would severely distort subendocardial vessels and increase the resistance to flow through them. Utley et al. (33) reached a similar conclusion. Four studies by others of experimental pericardial tamponade are consistent with our results in showing a decreased inner-toouter flow ratio during tamponade (13, 25, 30, 36). In one of these studies, tamponade was shown to cause diffuse, punctate subendocardial hemorrhages (36). An additional study demonstrated decreased systolic coronary flow during tamponade (22); this is compatible with decreased subendocardial contractility due to ischemia. Our data support these studies and are noteworthy in that, in our studies, these changes occurred despite a maintained driving pressure, which does not usually occur during pericardial tamponade. The heterogeneity of flow in pieces within a layer of the left ventricle agrees with other studies (3, 5, 37), including those done with an arrested heart (1). The greater flow in the subendocardium than the subepicardium at low driving pressures has been noted before in our laboratory (34) and is almost certainly due to the greater conductance in subendocardial vessels in the arrested heart (11). Raising LVP decreased flows more in the subendocardium than in the subepicardium, as discussed above. At a driving pressure of 10 mmHg and no pressure in the left ventricle, all pieces of muscle in all layers had flow, albeit small, so that Pzf levels had to be below 10 mmHg, as we found. With a high LVP, however, all the subendocardial pieces had flows close to zero and not distinguishable from it by the microsphere method; Austin et al. (4) concluded that the confidence limits for the measurement of flows by the microsphere method were ⫾0.10 ml/min. However, two Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 ␣-adrenergic stimulation on Pzf are mediated through coronary venous pressure is unknown. Although the heart was arrested and the vessels were maximally dilated, at zero pressures in and around the heart vascular resistances were lowest at the highest flow rates and about twice as high at the lowest flow rates. This indicates that the increased flows and accompanying higher perfusing pressures dilated the resistance vessels passively, as previously described (19). Interestingly enough, the arteriovenous pressure difference and therefore the coronary vascular resistance tended to be constant or even to increase at any given flow as pressures were raised in or around the heart, even though the increases in coronary arterial and venous pressures would be expected to cause resistance to decrease (19). This finding suggests that the dilating effect of raised vascular pressures was opposed by increased vascular resistances, probably mainly subendocardial, related to compression of the left ventricular wall or, for RVP, the septum. VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW This work was supported in part by National Heart, Lung, and Blood Institute (NHLBI) Program Project Grant HL-25847. G. S. Aldea received NHLBI Training Grant HL-07192, H. Mori was supported by the American Heart Association, California Affiliate, and R. H. Austin was a Stanley J. Sarnoff Fellow. REFERENCES 1. Aldea GS, Austin RE, Flynn AE, Coggins DL, Husseini W, and Hoffman JIE. Heterogeneous delivery of cardioplegic solution in the absence of coronary artery disease. J Thorac Cardiovasc Surg 99: 345–353, 1990. 2. Armour JA and Klassen GA. Pressure and flow in epicardial coronary veins of the dog heart: response to positive inotropism. Can J Physiol Pharmacol 62: 38–48, 1984. 3. Austin, RE Jr, Aldea GS, Coggins DL, Flynn AE, and Hoffman JIE. Profound spatial heterogeneity of coronary reserve. Discordance between patterns of resting and maximal myocardial blood flow. Circ Res 67: 319–331, 1990. 4. Austin, RE Jr, Hauck WW, Aldea GS, Flynn AE, Coggins DL, and Hoffman JIE. Quantitating error in blood flow measurements with radioactive microsphere. Am J Physiol Heart Circ Physiol 257: H280–H288, 1989. 5. Austin, RE Jr, Smedira NG, Squiers TM, and Hoffman JIE. Influence of cardiac contraction and coronary vasomotor tone on regional myocardial blood flow. Am J Physiol Heart Circ Physiol 266: H2542–H2553, 1994. 6. Aversano T, Klocke FJ, Mates RE, and Canty JM Jr. Preload-induced alterations in capacitance-free diastolic pressureflow relationships. Am J Physiol Heart Circ Physiol 246: H410– H417, 1984. 7. Baer RW, Payne BD, Verrier ED, Vlahakes GJ, Molodowitch D, Uhlig PN, and Hoffman JIE. Increased number of myocardial blood flow measurements with radionuclide-labeled microspheres. Am J Physiol Heart Circ Physiol 246: H418–H434, 1984. 8. Bellamy RF. Comments on interpretation and physiological significance of diastolic coronary artery pressure-flow relationships in the canine coronary bed. Circ Res 57: 485–486, 1985. 9. Bellamy RF. Diastolic coronary pressure-flow relations in the dog. Circ Res 43: 92–101, 1978. 10. Buckberg GD, Fixler DE, Archie JP, and Hoffman JIE. Experimental subendocardial ischemia in dogs with normal coronary arteries. Circ Res 30: 67–81, 1972. 11. Chilian WM. Microvascular pressures and resistances in the left ventricular subepicardium and subendocardium. Circ Res 69: 561–570, 1991. 12. Coggins DL, Flynn AE, Austin RE Jr, Aldea GS, Muehrcke D, Goto M, and Hoffman JIE. Nonuniform loss of regional flow reserve during myocardial ischemia in dogs. Circ Res 67: 253– 264, 1990. 13. Cohen MV. Myocardial blood flow in dogs with coronary occlusion: effects of isoproterenol. Am Heart J 109: 1032–1038, 1985. 14. Duncker DJ, Van Zon NS, Crampton M, Herrlinger S, Homans DC, and Bache RJ. Coronary pressure-flow relationship and exercise: contributions of heart rate, contractility, and ␣1-adrenergic tone. Am J Physiol Heart Circ Physiol 266: H795– H810, 1994. 15. Duncker DJ, Zhang J, and Bache RJ. Coronary pressureflow relation in left ventricular hypertrophy. Importance of changes in back pressure versus changes in minimum resistance. Circ Res 72: 579–587, 1993. 16. Duncker DJ, Zhang J, Pavek TJ, Crampton MJ, and Bache RJ. Effect of exercise on coronary pressure-flow relationship in hypertrophied left ventricle. Am J Physiol Heart Circ Physiol 269: H271–H281, 1995. 17. Ellis A and Klocke FJ. Effects of preload on the transmural distribution of perfusion and pressure-flow relationships in the canine coronary vascular bed. Circ Res 46: 68–77, 1980. 18. Emery JL, Omens JH, and McCulloch AD. Biaxial mechanics of the passively stretched left ventricle. Am J Physiol Heart Circ Physiol 272: H2289–H2305, 1997. 19. Hanley FL, Messina LM, Grattan MT, and Hoffman JIE. The effect of coronary inflow pressure on coronary vascular resistance in the isolated dog heart. Circ Res 54: 760–772, 1984. 20. Hoffman JIE. Transmural myocardial perfusion. Prog Cardiovasc Dis 29: 429–464, 1987. 21. Hoffman JIE and Spaan JAE. Pressure-flow relations in coronary circulation. Physiol Rev 70: 331–390, 1990. 22. Jarmakani JMM, McHale PA, and Greenfield JC Jr. The effect of cardiac tamponade on coronary hemodynamics in the awake dog. Cardiovasc Res 9: 112–117, 1975. 23. Klassen GA and Armour JA. Canine coronary venous pressures: responses to positive inotropism and vasodilation. Can J Physiol Pharmacol 61: 213–221, 1983. 24. Ledingham SJM, Braimbridge MV, and Hearse DJ. Improved myocardial protection by oxygenation of St. Thomas’ Hospital cardioplegic solutions: studies in the rat. J Thorac Cardiovasc Surg 95: 103–111, 1988. 25. Martins JB, Manuel WJ, Marcus ML, and Kerber RE. Comparative effects of catecholamines in cardiac tamponade: experimental and clinical studies. Am J Cardiol 46: 59–66, 1980. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 pieces of the subepicardium had flows above this threshold. Hence, in this heart, zero flow at the main coronary arterial level would not have been detected at a driving pressure of 10 mmHg and a LVP of 30 mmHg. These results suggest that there is probably a distribution of Pzf across the left ventricular wall and that the concept of Pzf applies to only a small portion of the heart rather than being a function of global flow (see Bellamy, Refs. 8 and 9). Rouleau and White (26) raised coronary sinus pressure to 18–28 mmHg at fixed inflow pressures and observed that flow was decreased in the subepicardium but not in the subendocardium. They concluded that this finding indicated that the high venous pressures exceeded intramyocardial pressures only in the subepicardium and that an intramyocardial vascular waterfall with a high surrounding pressure regulated subendocardial flow. A more direct demonstration of this was provided by Satoh et al. (27), who found that as LVP was raised in the arrested heart, Pzf increased and was similar to subepicardial intramyocardial pressure that was lower than that in the subendocardium. Moreover, there are differences of Pzf even within a layer. It is likely that the curvature observed frequently at the lower end of the PF curve (21) is in part due to sequential drop out of small regions within a layer as well as between layers. These differing regional effects suggest that Pzf cannot be a function only of extracardiac venous waterfalls but is related also to intramyocardial pressure or different regional microvascular time constants (21). Our study has some important clinical implications. Clinically, a dilated heart with an elevated left ventricular end-diastolic pressure has been associated with a vicious cycle of further dilatation and failure. Our studies suggest that elevation of transmural pressure produces a disproportionate reduction in subendocardial blood flow, even if coronary arterial pressures are maintained, and could lead to subendocardial ischemia. This may be one of the reasons for the subendocardial ischemia reported in congestive cardiomyopathy (32). Furthermore, if cardioplegia is done in a patient with a high left ventricular diastolic pressure, there may be inadequate delivery of cardioplegic solution and thus inadequate myocardial protection in the subendocardium, even at normal perfusing pressures. H2937 H2938 VENTRICULAR TRANSMURAL PRESSURE AND CORONARY BLOOD FLOW 32. Unverferth DV, Magorien RD, Lewis RP, and Leier CV. The role of subendocardial ischemia in perpetuating myocardial failure in patients with nonischemic congestive cardiomyopathy. Am Heart J 105: 176–179, 1983. 33. Utley JR, Michalsky GB, Mobin-Uddin K, and Bryant LR. Subendocardial vascular distortion at small ventricular volumes. J Surg Res 17: 114–24, 1974. 34. Verrier ED, Baer RW, Hickey RF, Vlahakes GJ, and Hoffman JIE. Transmural pressure-flow relations during diastole in the canine left ventricle (Abstract). Circulation 3: 62, 1980. 35. Watanabe J, Maruyama Y, Satoh S, Keitoku M, and Takishima T. Effects of the pericardium on the diastolic left coronary pressure-flow relationship in the isolated dog heart. Circulation 75: 670–675, 1987. 36. Wechsler AS, Auerbach BJ, Graham TC, and Sabiston DC. Distribution of intramyocardial blood flow during pericardial tamponade. J Thorac Cardiovasc Surg 68: 847–856, 1974. 37. Yipintsoi T, Dobbs WA Jr, Scanlon PD, Knopp TJ, and Bassingthwaighte JB. Regional distribution of diffusible tracers and carbonized microspheres in the left ventricle of isolated dog hearts. Circ Res 33: 573–587, 1973. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.1 on May 15, 2017 26. Rouleau JR and White M. Effects of coronary sinus pressure elevation on coronary blood flow distribution in dogs with normal preload. Can J Physiol Pharmacol 63: 787–797, 1985. 27. Satoh S, Maruyama Y, Watanabe J, Keitoku M, Hangai K, and Takishima T. Comparison of zero-flow pressure (Pf ⫽ 0) in diastolic coronary pressure-flow relationship with diastolic pressures in outer (IMPo) and inner (IMPi) myocardial layers. Circulation 76: 489, 1987. 28. Satoh S, Watanabe J, Keitoku M, Itoh N, Maruyama Y, and Takishima T. Influences of pressure surrounding the heart and intracardiac pressure on the diastolic coronary pressure-flow relation in excised canine hearts. Circ Res 63: 788–797, 1988. 29. Scharf SM, Bromberger-Barnea B, and Permutt S. Distribution of coronary venous flow. J Appl Physiol 30: 657–662, 1971. 30. Taichman GC, Byrne P, Forester GV, and Keon WJ. Regional myocardial blood flow during pericardial tamponade. Can J Physiol Pharmacol 62: 539–543, 1983. 31. Uhlig PN, Baer RW, Vlahakes GJ, Hanley FL, Messina LM, and Hoffman JIE. Arterial and venous coronary pressure-flow relations in anesthetized dogs. Evidence for a vascular waterfall in epicardial coronary veins. Circ Res 55: 238–248, 1984.