Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Gene nomenclature wikipedia , lookup
Genetic engineering wikipedia , lookup
Gene expression profiling wikipedia , lookup
Public health genomics wikipedia , lookup
Genome evolution wikipedia , lookup
Hardy–Weinberg principle wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Genetic drift wikipedia , lookup
Pharmacogenomics wikipedia , lookup
Population genetics wikipedia , lookup
Genome (book) wikipedia , lookup
Dominance (genetics) wikipedia , lookup
Human genetic variation wikipedia , lookup
Designer baby wikipedia , lookup
Koinophilia wikipedia , lookup
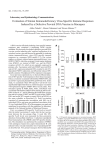
Behavior Genetics ( 2005) DOI: 10.1007/s10519-005-9017-8 Differential Functional Variability of Serotonin Transporter and Monoamine Oxidase A Genes in Macaque Species Displaying Contrasting Levels of Aggression-Related Behavior Jens R. Wendland,1,6 Klaus-Peter Lesch,1 Timothy K. Newman,2 Angelika Timme,3 Hélène Gachot-Neveu,4 Bernard Thierry,4 and Stephen J. Suomi5 Received 18 Jan. 2005—Final 17 June 2005 Functional allelic variation in the transcriptional control region of the serotonin transporter and monoamine oxidase A genes has been associated with anxiety- and aggression-related behavior in humans and, more recently, in nonhuman primates. Here, we have genotyped these polymorphic regions in seven species of the genus Macaca. Macaques exhibit exceptional inter-species variation in aggression-related social behavior as illustrated by recent studies showing overlapping patterns of aggression-based social organization grades and macaque phylogeny. We cloned and sequenced two new alleles of the serotonin transporter gene-linked polymorphic region in Barbary and Tibetan macaques. In addition, we observed that species displaying tolerant societies, with relaxed dominance and high levels of conciliatory tendency, were monomorphic for both the serotonin transporter gene and, with the exception of Tonkean macaques, the monoamine oxidase A gene. In contrast, those species known to exhibit intolerant, hierarchical and nepotistic societies were polymorphic at one or more of these loci. Rhesus (M. mulatta), the most intolerant and hierarchical species of macaques, showed the greatest degree of allelic variation in both genes. Additional investigation of a polymorphic repeat in exon III of the dopamine receptor D4 as well as a repeat/single nucleotide polymorphism in the 3¢untranslated region of the dopamine transporter which have both been implicated in the modulation of complex behavior failed to reveal a relationship between allelic variability and social organization grade. Taken together, these findings suggest that genetic variation of serotonergic neurotransmission may play an important role in determining inter-species differences in aggression related behavior in macaques. KEY WORDS: Allelic variation; behavioral genetics; evolution; nonhuman primates. 1 2 3 4 Clinical and Molecular Psychobiology, Department of Psychiatry and Psychotherapy, University of Würzburg, 97080 Würzburg, Germany. National Institute on Alcohol Abuse and Alcoholism, Laboratory of Clinical Studies-Primate Unit, Poolesville, MD 20837, USA. Institute of Human Biology and Anthropology, Freie Universität Berlin, 14195 Berlin, Germany. Centre d’Ecologie et Physiologie Energétiques, Centre National pour la Recherche Scientifique, Strasbourg, France. 5 6 National Institute on Child Health and Human Development, Laboratory of Comparative Ethology, Poolesville, MD 20837, USA. To whom correspondence should be addressed at Laboratory of Clinical Science, National Institute of Mental Health, 10 Center Dr.; Bldg. 10, Rm. 3D41, Bethesda, MD 20892-1264, USA. Tel.: +1-301-496 3421; Fax: +1-301-402 0188; e-mail: wendlandj@ mail.nih.gov 2005 Springer Science+Business Media, Inc. Wendland et al. INTRODUCTION In search of genetic variation influencing behavior, a significant role for neurotransmission, as well as evidence of functionality, are considered to be crucial (Ebstein et al., 2002). In humans, two genes of the serotonin (5-hydroxytryptamine, 5-HT) pathway meeting these criteria are the 5-HT transporter (5HTT, SLC6A4), which has been associated with anxiety- and depression-related traits (Caspi et al., 2003; Lesch et al., 1996); and monoamine oxidase A (MAOA), which is believed to influence aggressive behavior (Caspi et al., 2002; Deckert et al., 1999; see Munafo et al., 2003 and Sen et al., 2004 for reviews). Both the human 5-HTT and MAOA genes contain gene-linked polymorphic regions (LPR) located upstream of the coding sequence, termed 5-HTTLPR and MAOALPR, respectively. These polymorphisms have been shown in vitro to alter the transcriptional activity of 5-HTT and MAOA, and subsequently the function of the corresponding proteins (Deckert et al., 1999, Denney et al., 1999; Lesch, 2002, 1996; Sabol et al., 1998). In nonhuman primates, orthologous functional polymorphic regions have been identified in rhesus macaques (Macaca mulatta), designated as rhMAOALPR and rh5-HTTLPR, respectively (Bennett et al., 2002; Lesch et al., 1997; Newman et al., 2005). Several recent studies suggest that rh5HTTLPR genotypes interact with deleterious early rearing experience in captive rhesus macaques to influence orientation, attentional and affective capabilities (Champoux et al., 2002), sensitivity to ethanol (Barr et al., 2003a, 2004a) and limbic-hypothalamicpituitary-adrenal axis stress response (Barr et al., 2004b). With regard to aggressive behavior, a significant genotyperearing interaction in male rhesus macaques has recently been demonstrated for the rhMAOALPR (Newman et al., 2005). Geographically one of the most widely distributed species of nonhuman primates, rhesus macaques exhibit a degree of variation in aggression-related behavior that is unequalled in other primate species (Petit et al., 1997; Thierry, 1985; Thierry, 1990; de Waal and Luttrell, 1989). Thierry (2000) recently proposed a four-grade classification scheme for describing social organization across 16 species of the genus Macaca. This classification is based on the extent and asymmetry of aggression-related behavior within specific macaque species. Grade 1 species exhibit highly hierarchical and nepotistic societies as well as low levels of conciliatory behaviors, while grade 4 species can be considered as more tolerant, displaying relaxed dominance, open relationships and high levels of conciliatory behaviors. For example, the risk of a retaliatory attack from a subordinate is much higher in grade 4 species than in grade 1 after an initial attack by a dominant, but the retaliatory attack will likely be much less severe, and the probability of reconciliation much higher, in grade 4 than grade 1 macaques (Thierry, 2000). By mapping this distribution of social style grades onto phylogenetic trees provided by Purvis (1995) and Morales and Melnick (1998), a significant association with phylogeny was shown for 7 of the 16 traits used for the classification, including patterns of female rank acquisition and male-female dominance relationships (Thierry et al., 2000). This led to the conclusion that the structure of macaque social organization is influenced more by phylogeny than environment. Thus, it is reasonable to hypothesize that specific genetic polymorphisms contribute to the variability in species-level behavior. In order to provide a molecular basis for this hypothesis, we genotyped rh5-HTTLPR and rhMAOALPR in seven macaque species representing the spectrum of different social grades. In addition, we genotyped (1) a physiologically relevant repeat polymorphism in exon III of the dopamine receptor subtype 4 gene (DRD4) that has been associated with novelty seeking in humans (Benjamin et al., 1996; Ebstein et al., 1996), and (2) a functional repetitive element/single nucleotide polymorphism in the 3¢untranslated region of the dopamine transporter gene (DAT 3¢-UTR) which has been shown to be suggestive though not predictive of hyperactive behavior in rhesus macaques (Miller et al., 2001). MATERIALS AND METHODS Animals Macaca fascicularis. A total of 35 longtail macaques (19f, 16m) originated from two independent sources: (1) CIBA Vision, Strasbourg, France (12f, 13m, average pairwise relatedness <2%) and (2) the AAP Sanctuary for Exotic Animals, Almere, The Netherlands (7f, 4m, no data on kinship available). Macaca nemestrina. Twelve pigtail macaques (7f, 5m) were included in this study, two of which were unrelated females from the Centre de Primatologie, Strasbourg. The remainder originated from the above-mentioned AAP Sanctuary (no kinship data available) in Almere. Serotonin Transporter and MAOA Genes Variability in Macaques Macaca tonkeana. A total of 28 Tonkean macaques (10f, 18m) from the Centre de Primatologie at Strasbourg originated from four maternal lineages, which in turn emerged from a stock of seven breeders imported to France in 1972. Macaca sylvanus. The sample population of 87 Barbary macaques (60f, 27m) analyzed in this study was composed of four independent groups: (1) 13 males from the AAP Sanctuary in Almere; (2) 16 individuals (8f, 8m) from Naturzoo Rheine, Germany; (3) 46 females from Affenberg Salem, Germany; and (4) 12 individuals (6f, 6m, originally imported from Gibraltar) from Wildpark Daun, Germany. No data on kinship were available for these groups. Macaca thibetana and Macaca arctoides. Three Tibetan (Macaca thibetana, one female and male each that were unrelated and their son) and 2 unrelated stumptail macaques (Macaca arctoides, 1f, 1m) originated from the Centre de Primatologie, Strasbourg. Macaca mulatta. A total of 90 rhesus macaques (28f, 62m, no kinship data available) originated from the Deutsches Primatenzentrum, Göttingen, Germany (23f, 57m), and the AAP Sanctuary (5f, 5m). Both groups were independent of each other. These samples were genotyped only for the exon III polymorphism in the DRD4, as we and others have previously reported on rh5-HTTLPR (Bennett et al., 2002; Lesch et al., 1997), rhMAOALPR (Newman et al., 2005) and DAT 3¢-UTR (Miller et al., 2001). Polymerase Chain Reaction Genotyping and Data Analysis All protocols were approved by the relevant local animal care and use committees. After i.m. anesthetization with 15 mg/kg ketamine, peripheral blood was drawn and DNA extracted according to standard procedures. 5-HTTLPR was amplified using oligonucleotide primers rhMUT 5¢-TCG ACT GGC GTT GCC GCT CTG AAT GC-3¢and rhINT 5¢-CAG GGG AGA TCC TGG GAG GGA-3¢under reaction conditions previously described (Lesch et al., 1997). MAOALPR was amplified using primers MAOAjrwF2 5¢-AGA AGG GCT GCG GGA AGC-3¢ and MAOA-jrwR 5¢-GTG CTC CAC TGG GAA CTG G-3¢under the following reaction conditions; 25– 50 ng DNA were mixed with reaction buffer containing 1.8 mM MgCl2, 4% DMSO, 0.2 mM dNTP each, 400 nM forward and reverse primer each and 1.25 U of Roche FastStart HighFidelity Taq polymerase in a final reaction volume of 25lL; amplifi- cation conditions were 2 min initial denaturaion at 95C followed by 35 cycles of 95¢C–65C–72C (30 s each) and a final elongation for 5 min at 72C. Amplification and restriction endonuclease digestion conditions followed the published protocols of Ebstein et al. (1996) for the DRD4 exon III polymorphism, and Miller et al. (2001) for DAT 3¢-UTR. Amplicons were separated by 3.5% NuSieve agarose gel electrophoresis and visualized by ethidium bromide (Sigma) staining. Genotypes were assigned based on estimated allele sizes established using sequenced controls from prior experiments. In the case of 5-HTTLPR, two new alleles were identified, cloned with the TA Cloning Dual Promoter Kit (Invitrogen) and sequenced by automated cycle sequencing employing the fluorescent dye termination method (ABI-Prism 310). Sequence alignment for 5-HTTLPR in Figure 1 was performed using human S, L, xL and xxL alleles (GenBank accession numbers X76753 and Y13147) and rhesus rhS, rhL and rhxL alleles (AF191557). In every species analyzed, observed genotype frequencies did not significantly deviate from Hardy– Weinberg equilibrium as determined by v2 test. Also, all allelic frequencies presented in this study did not significantly differ between sexes. RESULTS Serotonin Transporter Gene-linked Polymorphic Region We have previously reported on three different rh5-HTTLPR alleles in rhesus macaques, namely rhS, rhL and rhxL (Bennett et al., 2002; Lesch et al., 1997). In this study, we identified two additional alleles: one in Barbary macaques, which we designate msy (for Macaca sylvanus, GenBank accession number AY897212); the other one in Tibetan macaques, designated mti (AY897213). Sequence alignment revealed that the mti allele, which is 23 bp shorter than rhS, has most likely evolved from a deletion of three repetitive motifs in the second polymorphic locus (PL2) described previously (Fig. 1) (Lesch et al., 1997). The msy allele has almost exactly the same length as rhxL, but it displays an additional repetitive unit in PL2 and lacks the insertion further downstream of the rhxL allele (PL3, Fig. 1). All of the other macaque species investigated carried the rhL allele. In addition, all species analyzed, except for rhesus macaques, were monomorphic at this locus (Table I). Wendland et al. Fig. 1. Multiple sequence alignment of 5-HTTLPR alleles in humans and macaques. For alignment optimization and comprehensibility, hyphens were introduced and repetitive units separated by spaces. S, L, xl and xxL refers to human alleles described previously (GenBank acc. nos. X76753, Y13147); rhS, rhL and rhxL alleles were first observed in rhesus macaques (AF191557). Mti, Tibetan macaque allele (AY897213); msy, Barbary macaque allele (AY897212); PL, polymorphic locus. Note that variation in humans (PL1) and macaques (PL2 and 3) occurs at different, albeit close, genetic loci. Monoamine Oxidase A-linked Polymorphic Region Alleles in the promoter region of the MAOA gene had 5–7 repeats of the 18 bp repetitive sequence, designated rh5, rh6 and rh7, as described previously for rhesus macaques (Newman et al., 2005; Wendland et al., in press). Barbary macaques were monomorphic, carrying the rh6 allele, whereas stumptail and Tibetan macaques were monomorphic for the rh7 allele (Table I). The remaining macaque species were polymorphic: pigtail (rh6 and rh7), Tonkean (rh6 and rh7), longtail (rh6 and rh7) and rhesus macaques (rh5, rh6 and rh7, Table I). Thus, as with the 5-HTTLPR locus, variability of MAOALPR alleles was greatest in rhesus macaques. Dopamine Receptor D4 Exon III Polymorphism We observed only two DRD4 exon III alleles containing either 4 or 5 repeats. All Barbary macaques were monomorphic for the 4 repeat allele, while all Tibetan, pigtail and stumptail macaques were monomorphic for the 5 repeat allele. Rhesus, longtail and Tonkean macaques displayed polymorphism at the DRD4 locus with 4 and 5 repeat alleles, the frequencies of which are shown in Table I. Dopamine Transporter 3¢Untranslated Region Polymorphism All samples analyzed were monomorphic, exhibiting only the 12 repeat allele, with the exception of Tonkean and pigtail macaques (11 and 12 repeat alleles, Table I). The single nucleotide polymorphism (SNP) in the last repetitive unit recognized by the restriction enzyme Bst1107I varied only in pigtail macaques and in two Barbary macaques, both of which were heterozygous (Table I). SNP+ SNP) rh6 (47%) rh7 (53%) rh6 (26%) rh7 (74%) SNP+ (100%) 4 (19%) 5 (81%) 12 (100%) rhL (100%) rhL (100%) rhS (26%) rhL (74%) rhxL (0.2%) rh5 (35%) rh6 (25%) rh7 (40%) 4 (89%) 5 (11%) 12 (100%) 11 (8%) 12 (92%) SNP+ (63%) SNP) (37%) 5 (100%) 12 (5/7) 35 (16/19) 22–289b,c,d 2 M. nemestrina (Pigtail m.) 2 M. fascicularis (Longtail m.) 1 M. mulatta (Rhesus m.) SNP+ (99%) SNP) (1%) 12 (100%) 4 (100%) rh6 (100%) msy (100%) 87 (27/60) 3 M. sylvanus (Barbary m.) SNP+ 12 5 rh7 mti 3 (2/1) 3 M. thibetana (Tibetan m.) SNP+ 12 5 rh7 rhL 2 (1/1) 3 M. arctoides (Stumptail m.) 4 (25%) 5 (75%) 11 (18%) 12 (82%) SNP+ (100%) rh6 (34%) rh7 (66%) rhL (100%) 28 (18/10) 4 M. tonkeana (Tonkean m.) Seven different macaque species of all four social style grades as proposed by Thierry (2000) were genotyped. With the exception of Macaca tonkeana, macaque species exhibiting more tolerant societies (i.e. grades 3 and 4) carry only one kind of allele for 5-HTTLPR and MAOALPR each. In contrast, intolerant, hierarchical and nepotistic macaque species (grades 1 and 2) are polymorphic at one or more of these loci. a SNP +/) refers to the Bst1107I restriction enzyme polymorphism in the last repetitive unit of DAT 3¢-UTR. b Data for rh5-HTTLPR (n=289) were pooled from previous experiments (Bennett et al., 2002; Lesch et al., 1997). c rhMAOALPR data (n=217) are taken from Newman et al. (2005). d DAT 3¢-UTR data (n=22) from Miller et al. (2001); for DRD4 in rhesus macaques, n=90. Allelic frequencies were omitted for Tibetan and stumptail macaques due to small sample size and for DAT 3¢-UTR SNP in rhesus macaques due to missing details in cited work. DAT 3¢-UTRa DRD4 exon III MAOALPR Social organization category sensu Thierry Number of animals (male/female) 5-HTTLPR Macaque species Table I. Distribution of Alleles and Respective Frequencies in Polymorphic Genetic Loci Presumably Involved in Behavior Serotonin Transporter and MAOA Genes Variability in Macaques Wendland et al. DISCUSSION The present study has analyzed seven species of the genus Macaca for functional polymorphisms in genes of the serotonergic and dopaminergic pathway that are assumed to influence behavior. We have shown that genetic loci involved in the regulation of serotonergic neurotransmission, especially the 5-HTTLPR, vary not only in their sequence composition, but also in their allelic frequencies across macaque species. The presented data provide a molecular basis for future association studies of individual behavior with functional genetic variants in a novel inter-species approach within one genus of nonhuman primates. Macaques were investigated because of the behavioral classification developed by Thierry (2000) that is based on inter-species differences in aggression-related behavior, and also because of recent evidence for the involvement of rh5-HTTLPR (Champoux et al., 2002) and rhMAOALPR (Newman et al., 2005) in aggressive behavior, affective capacities, and emotion regulation in rhesus macaques. Moreover, it has been shown that Thierry’s social style grades cluster within species groups in a manner that is consistent with their phylogenetic relationships (Thierry et al., 2000). Our data show that macaque species (except Tonkean) exhibiting less aggressive, more tolerant behavior (grades 3 and 4) displayed no intraspecific variation in 5-HTTLPR and MAOA-LPR genotypes. In contrast, highly intolerant and hierarchical macaque species (grades 1 and 2) were polymorphic at one or both of these loci (Table I). The genetic loci analyzed in this study have been chosen, on the one hand, on the evidence of association with human personality traits as well as the presence of geneenvironment effects in captive rhesus macaques and, on the other hand, due to in vitro data showing an effect upon the molecular physiology of neurotransmission. The first gene variant to be associated with a human personality trait, in this case novelty seeking, was a polymorphism in the third cytoplasmatic loop of DRD4 (Benjamin et al., 1996; Ebstein et al., 1996). This polymorphism consists of 2–10 repetitive units of 48 bp each and is believed to influence second messenger signaling (Asghari et al., 1995; Kazmi et al., 2000; Lichter et al., 1993;Van Tol et al., 1992). Alleles with 4 or 7 repeats account for the vast majority of all alleles in humans (Lichter et al., 1993; Van Tol et al., 1992). In our study, macaques only carried alleles of the same size as human 4 and 5 repeat variants, and there was no apparent relationship between the degree of variability in this polymorphic locus and the social grade of macaque species (Table I). The second functional polymorphism in the dopaminergic pathway analyzed in this study was DAT 3¢-UTR. This variable number of tandem repeat (VNTR) polymorphism comprises 3–11 copies of a 40 bp motif in humans and has, among others, been associated with attention deficit hyperactivity disorder (Cook et al., 1995; Gill et al., 1997; Sano et al., 1993; Vandenbergh et al., 1992). Possibly as a cisacting regulatory determinant of mRNA half-life, this VNTR polymorphism has been shown in vitro to alter reporter gene expression (Conne et al., 2000; Fuke et al., 2001). Only one 12-repeat allele is known, to date, to exist in rhesus macaques, but a SNP in the last repetitive unit of VNTR that is recognized by the restriction enzyme Bst1107I was suggestive (though not predictive) of hyperactive behavior in a sample of 22 adult male rhesus macaques (Miller et al., 2001). In longtail macaques, alleles with 11 and 12 repeats have recently been described, exhibiting significantly different activities on reporter gene assays (InoueMurayama et al., 2002). We observed an allele of 11 repeats in Tonkean (allelic frequency 18%) and pigtail macaques (in 2 out of 24 alleles), but not in longtail macaques (Table I). Regarding the Bst1107I polymorphism, only Barbary and pigtail macaques were polymorphic, while all other analyzed species lacked the restriction site. As in the case of DRD4, we found no relationship between macaque social style grade and within-species variability of the DAT 3¢-UTR VNTR or SNP. In serotonergic neurotransmission, the 5-HTT plays a key role in controlling intrasynaptic availability of 5-HT (reviewed in Murphy et al., 2004). The human 5-HTT was the second gene for which a positive association with personality traits was shown (Lesch et al., 1996). The variation consists of a 43 bp insertion/deletion in the transcriptional control region which affects reporter gene expression in vitro (Lesch et al., 1996). In numerous follow-up studies, the initial observation of association with anxietyand depression-related traits was replicated in several but not all studies (for review see, e.g., Lesch, 2002; Sen et al., 2004). Recent data from a prospectivelongitudinal birth cohort study (Caspi et al., 2003) have shown that individuals carrying one or two copies of the S allele are more likely to exhibit diagnosable depression after stressful life events than L/L genotype individuals, supporting a role as Serotonin Transporter and MAOA Genes Variability in Macaques susceptibility gene. To date, at least 14 different alleles have been discovered in humans, 6 in great apes (Hominidae), 5 in Müller’s gibbons (Hylobates muelleri) and 3 in rhesus macaques (Bennett et al., 2002; Inoue-Murayama et al., 2000; Lesch et al., 1997; Nakamura et al. 2000). While the functional implications for most of these alleles remain unclear, a significant difference in reporter gene expression was demonstrated for the human S and L alleles as well as rhesus S and L alleles (Bennett et al., 2002; Lesch et al., 1996). In humans this functional dichotomy was subsequently confirmed by different approaches at both the pre- and postsynaptic level (David et al. 2005, reviewed in Lesch, 2002). Here, we observed two novel alleles in Barbary and Tibetan macaques, both of which displayed sequence differences from rhS and rhL in the PL2 described previously (Fig. 1, Lesch et al., 1997). Interestingly, the msy allele carries the highest number of repeats in PL2 of all macaques. Bearing in mind that macaques represent a monophyletic lineage (Delson, 1980) and that Barbary macaques were the first species to diverge from this clade (Morales and Melnick, 1998), the msy allele thus comes closest of all currently available sequences to the ‘‘progenitor’’ 5-HTTLPR allele postulated previously (Lesch et al., 1997). Moreover, our data suggest that, with the exception of the very rare rhxL allele, macaque 5-HTTLPR alleles could have evolved from an msy-like ancestral allele by deletion of repetitive units in the PL2 (Fig. 1). The usefulness of the nonhuman primate model in the study of behavioral genetics is generally attributed to the ability to control and manipulate the animals’ environment (Barr et al., 2003b). Here, however, we have used a complementary approach by comparing species within the genus Macaca displaying different patterns of aggression-based social organization. The findings presented in this study support the importance of the central serotonergic system in behavioral genetics. Numerous association studies have looked at correlations between genetic polymorphisms and traits in considerably diverse species including, in particular, humans, nonhuman primates and rodents. By contrast, our study represents a novel attempt towards elucidating the genetic basis of social behavior by comparing well-established functional polymorphisms in genes of the serotonergic or dopaminergic pathway in different species within one genus. Macaque species are ideally suited to this purpose since they exhibit broad interspecific variation in patterns of aggression, dominance and temperament (Aureli et al., 1997; Petit et al., 1997; Thierry, 1985; Thierry, 1990; Thierry, 2000; de Waal and Luttrell, 1989). Moreover, it was shown for this genus that social organization and behavior have changed little during several hundred thousand years (Thierry et al., 2000). Ecological and climatic conditions are likely to have varied considerably during such a long period of time, and if one assumed that macaque behavior and social organization were solely influenced by environment, one should also expect changes in behavior as an expression of adjustment to changing peristasis. Instead, however, the influence of phylogeny was proposed by several studies (Chan, 1996; Matsumura, 1999; Thierry, 2000). In line with this hypothesis, Thierry and coworkers demonstrated significant association of his classification of social aggression in macaques based on two independently derived phylogenies (Thierry et al., 2000). In a similar approach investigating rodents of the genus Microtus known to display strong species differences in social structure, a significant effect of a polymorphic microsatellite upstream of the avpr1a gene upon in vitro and in vivo gene expression and social engagement and bonding behavior was recently demonstrated (Hammock and Young, 2005). Our findings reveal that functional polymorphisms in the serotonergic pathway vary both within and between macaque species: tolerant, less hierarchical and nepotistic macaque species (grades 3 and 4) were monomorphic at rh5-HTTLPR and rhMAOALPR (Barbary, Tibetan and stumptail macaques) or displayed only two alleles for rh5-HTTLPR (Tonkean macaques). In contrast, highly asymmetrically aggressive and hierarchical species (grades 1 and 2) were polymorphic in the serotonergic pathway, and the most intolerant species (rhesus macaques, grade 1) also displayed the largest number of alleles at both loci. In summary, we feel it is justified at this point to hypothesize that rh5-HTTLPR and rhMAOALPR gene variants affect key elements of macaque social behavior, in particular the exceptional level of interspecies variation in aggression-related behavior in macaques. Certainly, despite its intrigue, our conclusion bears several caveats. Firstly, although all individuals in each species except for Tibetan and stumptail macaques originated from at least two independent sources (4 sources in the case of monomorphic Barbary macaques), we cannot rule out the possibility of having missed alleles through founder effects in the colonies from which our samples were derived. Secondly, the relative paucity of rh5-HTTLPR and rhMAOALPR variation in grade 3 and 4 species Wendland et al. could be due to statistical chance; for example, it has been calculated that one would need a minimum sample size of n=51 to detect all alleles with frequencies of ‡10% at the 95% confidence level (Gregorius, 1980). Finally, it would clearly not be justified to deduce social or behavioral variability from genetic variability at the population level as long as data on both behavior and genotype on individual subjects are lacking. Nevertheless, our results provide a first molecular basis to create falsifiable hypotheses for the study of specific aspects of social behavior, such as aggression, in a behaviorally exceptionally diverse and well-characterized genus of non-human primates. Therefore, to corroborate or disprove our hypothesis of an involvement of rh5-HTTLPR and rhMAOALPR underlying variability in social aggression, analyses of larger sample populations derived from unrelated individuals, as well as investigations of more macaque species of all grades are needed. Ideally, one would have behavioral and genotype data on individuals representing macaques of all species to perform association studies. Owing to 5HT system’s tremendous complexity with more than a dozen receptors and serotonergic nerve endings widely distributed in the CNS, we need an integrative model for serotoninergic neurotransmission spanning from seemingly simple molecular variations such as 5-HTTLPR and MAOALPR and synapse homeostasis to a highly complex phenomenon such as interindividual behavior in order to better understand the way in which genes might affect primate behavioral traits. DISCLAIMER In compliance with NIH/NIMH ethics guidelines, we hereby state that the views expressed do not necessarily represent those of NIH or NIMH or the federal government of the USA. ACKNOWLEDGMENTS The authors thank Gabriela Ortega, Nicole Steigerwald and Mukta Kundu for excellent technical assistance in blood and DNA sample processing, genotyping, cloning and data collection. We are greatly indebted to G. Flügge, Deutsches Primatenzentrum, Göttingen, Germany; I. den Hartog and E. van Lavieren, AAP Sanctuary for Exotic Animals, Almere, The Netherlands; A. Johann, Naturzoo Rheine, Germany; G. de Tuerkheim, E. Merz and W. Angst, Affenberg Salem, Germany; S. Bost and D. Reichle, Wildpark Daun, Germany; and CIBA Vision, Strasbourg, France, for providing blood samples of different macaque species. Supported by the Division of Intramural Research, National Institute of Child Health & Human Development, National Institutes of Health and the European Commission (NEWMOOD LSHM-CT-2003– 503474) and the Deutsche Forschungsgemeinschaft (SFB 581, KFO 125/1–1). REFERENCES Asghari, V., Sanyal, S., Buchwaldt, S., Paterson, A., Jovanovic, V., and Van Tol, H. H. (1995). Modulation of intracellular cyclic AMP levels by different human dopamine D4 receptor variants. J. Neurochem. 65:1157–1165. Aureli, F., Das, M., and Veenema, H. C. (1997). Differential kinship effect on reconciliation in three species of macaques (Macaca fascicularis, M. fuscata, and M. sylvanus). J. Comp. Psychol. 111:91–99. Barr, C. S., Newman, T. K., Becker, M. L., Champoux, M., Lesch, K. P., Suomi, S. J., Goldman, D., and Higley, J. D. (2003a). Serotonin transporter gene variation is associated with alcohol sensitivity in rhesus macaques exposed to early-life stress. Alcohol. Clin. Exp. Res. 27:812–817. Barr, C. S., Newman, T. K., Becker, M. L., Parker, C. C., Champoux, M., Lesch, K. P., Goldman, D., Suomi, S. J., and Higley, J. D. (2003b). The utility of the non-human primate; model for studying gene by environment interactions in behavioral research. Genes Brain Behav. 2:336–340. Barr, C. S., Newman, T. K., Lindell, S., Becker, M. L., Shannon, C., Champoux, M., Suomi, S. J., and Higley, J. D. (2004a). Early Experience and Sex Interact to Influence LimbicHypothalamic-Pituitary-Adrenal-Axis Function After Acute Alcohol Administration in Rhesus Macaques (Macaca mulatta). Alcohol. Clin. Exp. Res. 28:1114–1119. Barr, C. S., Newman, T. K., and Shannon, C. et al. (2004b). Rearing condition and rh5-HTTLPR interact to influence limbic-hypothalamic-pituitary-adrenal axis response to stress in infant macaques. Biol. Psychiat. 55:733–738. Benjamin, J., Li, L., Patterson, C., Greenberg, B. D., Murphy, D. L., and Hamer, D. H. (1996). Population and familial association between the D4 dopamine receptor gene and measures of Novelty Seeking. Nat. Genet. 12:81–84. Bennett, A. J., Lesch, K. P., Heils, A., Long, J. C., Lorenz, J. G., Shoaf, S. E., Champoux, M., Suomi, S. J., Linnoila, M. V., and Higley, J. D. (2002). Early experience and serotonin transporter gene variation interact to influence primate CNS function. Mol. Psychiat. 7:118–122. Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Martin, J., Craig, I. W., Taylor, A., and Poulton, R. (2002). Role of genotype in the cycle of violence in maltreated children. Science 297:851– 854. Caspi, A., Sugden, K., and Moffitt, T. E. et al. (2003). Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301:386–389. Champoux, M., Bennett, A., Shannon, C., Higley, J. D., Lesch, K. P., and Suomi, S. J. (2002). Serotonin transporter gene polymorphism, differential early rearing, and behavior in rhesus monkey neonates. Mol. Psychiat. 7:1058–1063. Chan, L. K. W. (1996). Phylogenetic interpretations of primate socioecology: with special reference to social and ecological Serotonin Transporter and MAOA Genes Variability in Macaques diversity in Macaca. In E. Martins (ed.), Phylogenies and the Comparative Method in Animal Behavior. New York: Oxford University Press, pp. 324–360. Conne, B., Stutz, A., and Vassalli, J. D. (2000). The 3¢untranslated region of messenger RNA: a molecular ’hotspot’ for pathology? Nat. Med. 6:637–641. Cook, E. H. Jr., Stein, M. A., Krasowski, M. D., Cox, N. J., Olkon, D. M., Kieffer, J. E., and Leventhal, B. L. (1995). Association of attention-deficit disorder and the dopamine transporter gene. Am. J. Hum. Genet. 56:993–998. David, S. P., Murthy, N. V., Rabiner, E. A., Munafo, M. R., Johnstone, E. C., Jacob, R., Walton, R. T., and Grasby, P. M. (2005). A functional genetic variation of the serotonin (5-HT) transporter affects 5-HT1A receptor binding in humans. J. Neurosci. 25:2586–2590. Deckert, J., Catalano, M., and Syagailo, Y. V. et al. (1999). Excess of high activity monoamine oxidase A gene promoter alleles in female patients with panic disorder. Hum. Mol. Genet. 8:621– 624. Delson, E. (1980). Fossil macaques, phyletic relationships and a scenario of deployment. In D. G. Lindburg (ed.), The Macaques: Studies in Ecology, Behavior, and Evolution. New York: Van Nostrand Reinhold, pp. 10–30. Denney, R. M., Koch, H., and Craig, I. W. (1999). Association between monoamine oxidase A activity in human male skin fibroblasts and the genotype of the MAO promoter-associated variable number tandem repeat. Hum. Genet. 105:541–551. Ebstein, R. P., Benjamin, J., and Belmaker, R. H. (2002). Behavioral Genetics, Genomics, and Personality. In R. Plomin, J. C. DeFries, I. W. Craig and P. McGuffin (eds.), Behavioral Genetics in the Postgenomic Era. Washington, DC: American Psychological Association, pp. 365–388. Ebstein, R. P., Novick, O., Umansky, R., Priel, B., Osher, Y., Blaine, D., Bennett, E. R., Nemanov, L., Katz, M., and Belmaker, R. H. (1996). Dopamine D4 receptor (D4DR) exon III polymorphism associated with the human personality trait of Novelty Seeking. Nat. Genet. 12:78–80. Fuke, S., Suo, S., Takahashi, N., Koike, H., Sasagawa, N., and Ishiura, S. (2001). The VNTR polymorphism of the human dopamine transporter (DAT1) gene affects gene expression. Pharmacogenomics J. 1:152–156. Gill, M., Daly, G., Heron, S., Hawi, Z., and Fitzgerald, M. (1997). Confirmation of association between attention deficit hyperactivity disorder and a dopamine transporter polymorphism. Mol. Psychiat. 2:311–313. Gregorius, H.-R. (1980). The probability of losing an allele when diploid genotypes are sampled. Biometrics 36:643–652. Hammock, E. A. D., and Young, L. J. (2005). Microsatellite instability generates diversity in brain and sociobehavioral traits. Science 308:1630–1634. Inoue-Murayama, M., Adachi, S., Mishima, N., Mitani, H., Takenaka, O., Terao, K., Hayasaka, I., Ito, S., and Murayama, Y. (2002). Variation of variable number of tandem repeat sequences in the 3¢-untranslated region of primate dopamine transporter genes that affects reporter gene expression. Neurosci. Lett. 334:206–210. Inoue-Murayama, M., Niimi, Y., Takenaka, O., Okada, K., Matsuzaki, I., Ito, S. i., and Murayama, Y. (2000). Allelic variation of the serotonin transporter gene polymorphic region in apes. Primates 41:267–273. Kazmi, M. A., Snyder, L. A., Cypess, A. M., Graber, S. G., and Sakmar, T. P. (2000). Selective reconstitution of human D4 dopamine receptor variants with Gi alpha subtypes. Biochemistry 39:3734–3744. Lesch, K. P. (2002). Neuroticism and Serotonin: A Developmental Genetic Perspective. In R. D. JC Plomin, J. C. DeFries, I. W. Craig and P. McGuffin (eds.), Behavioral Genetics in the Postgenomic Era. Washington, DC: American Psychological Association, pp. 389–424. Lesch, K. P., Bengel, D., Heils, A., Sabol, S. Z., Greenberg, B. D., Petri, S., Benjamin, J., Muller, C. R., Hamer, D. H., and Murphy, D. L. (1996). Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274:1527–1531. Lesch, K. P., Meyer, J., and Glatz, K. et al. (1997). The 5-HT transporter gene-linked polymorphic region (5-HTTLPR) in evolutionary perspective: alternative biallelic variation in rhesus monkeys. Rapid communication. J. Neural. Transm. 104:1259–1266. Lichter, J. B., Barr, C. L., Kennedy, J. L., Van Tol, H. H., Kidd, K. K., and Livak, K. J. (1993). A hypervariable segment in the human dopamine receptor D4 (DRD4) gene. Hum. Mol. Genet. 2:767–773. Matsumura, S. (1999). The evolution of ‘‘egalitarian’’ and ‘‘despotic’’ social systems among macaques. Primates 40:23–31. Miller, G. M., De La Garza, R. D. 2nd, Novak, M. A., and Madras, B. K. (2001). Single nucleotide polymorphisms distinguish multiple dopamine transporter alleles in primates: implications for association with attention deficit hyperactivity disorder and other neuropsychiatric disorders. Mol. Psychiat. 6:50–58 Morales, J. C., and Melnick, D. J. (1998). Phylogenetic relationships of the macaques (Cercopithecidae: Macaca), as revealed by high resolution restriction site mapping of mitochondrial ribosomal genes. J. Hum. Evol. 34:1–23. Munafo, M. R., Clark, T. G., Moore, L. R., Payne, E., Walton, R., and Flint, J. (2003). Genetic polymorphisms and personality in healthy adults: a systematic review and meta-analysis. Mol. Psychiat. 8:471–484. Murphy, D. L., Lerner, A., Rudnick, G., and Lesch, K. P. (2004). Serotonin transporter: gene, genetic disorders, and pharmacogenetics. Mol. Interv. 4:109–123. Nakamura, M., Ueno, S., Sano, A., and Tanabe, H. (2000). The human serotonin transporter gene linked polymorphism (5-HTTLPR) shows ten novel allelic variants. Mol. Psychiat. 5:32–38. Newman, T. K., Syagailo, Y., Barr, C. S., Wendland, J. R., Champoux, M., Graessle, M., Suomi, S. J., Higley, J. D., and Lesch, K. P. (2005). MAOA Gene Promoter Variation and Rearing Experience Influences Aggressive Behavior in Rhesus Monkeys. Biol. Psychiat. 57:167–172. Petit, O., Abegg, C., and Thierry, B. (1997). A comparative study of aggression and conciliation in three cercopithecine monkeys (Macaca fuscata, Macaca nigra, Papio papio). Behaviour 134:415–431. Purvis, A. (1995). A composite estimate of primate phylogeny. Philos. Trans. R. Soc. Lond. B Biol. Sci. 348:405–421. Sabol, S. Z., Hu, S., and Hamer, D. (1998). A functional polymorphism in the monoamine oxidase A gene promoter. Hum. Genet. 103:273–279. Sano, A., Kondoh, K., Kakimoto, Y., and Kondo, I. (1993). A 40nucleotide repeat polymorphism in the human dopamine transporter gene. Hum. Genet. 91:405–406. Sen, S., Burmeister, M., and Ghosh, D. (2004). Meta-analysis of the association between a serotonin transporter promoter polymorphism (5-HTTLPR) and anxiety-related personality traits. Am. J. Med. Genet. B Neuropsychiatr. Genet. 127:85–89. Thierry, B. (1985). Social development in three species of macaque (Macaca mulatta, M. fascicularis, M. tonkeana): a preliminary report on the first ten weeks of life. Behav. Process. 11:89–95. Thierry, B. (1990). Feedback loop between kinship and dominance: The macaque model. J. Theor. Biol. 145:511–521. Thierry, B. (2000). Covariation of conflict management patterns across macaque species. In F. Aureli and F. B. M. de Waal (eds.), Natural Conflict Resolution. Berkeley: University of California Press. Wendland et al. Thierry, B., Iwaniuk, A. N., and Pellis, S. M. (2000). The influence of phylogeny on the social behaviour of macaques (Primates: Cercopithecidae, genus Macaca). Ethology 106:713–728. Van Tol, H. H., Wu, C. M., Guan, H. C., Ohara, K., Bunzow, J. R., Civelli, O., Kennedy, J., Seeman, P., Niznik, H. B., and Jovanovic, V. (1992). Multiple dopamine D4 receptor variants in the human population. Nature 358:149–152. Vandenbergh, D. J., Persico, A. M., Hawkins, A. L., Griffin, C. A., Li, X., Jabs, E. W., and Uhl, G. R. (1992). Human dopamine transporter gene (DAT1) maps to chromosome 5p15.3 and displays a VNTR. Genomics 14:1104–1106. de Waal, F. B. M., and Luttrell, L. M. (1989). Toward a comparative socioecology of the genus Macaca: Different dominance styles in rhesus and stumptailed macaques. Am. J. Primatol. 19:83–109. Wendland, J. R., Hampe, M., Newman, T. K., Syagailo, Y., Meyer, J., Schempp, W., Timme, A., Suomi, S. J., and Lesch, K. P. (in press) Structural variation of the monoamine oxidase A gene promoter polymorphism in nonhuman primates. Genes Brain Behav. doi: 10.1111/j.1601-183X.2005.00130.x Edited by Jeanne Wehner