Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
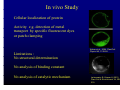
Isolation Purification and Characterisation Isolation, of Proteins from Plants Seema Mishra, Advanced Course on Bioinorganic Chemistry & Biophysics of Plants, summer semester 2012 In vivo Studyy Cellular localization of protein Activity e.g. detection of metal transport by specific fluorescent dyes or p patch clamping p g Limitations : No structural determination Kobae et al., 2004, PlantCell Physiol 45, 1749-58 No analysis of binding constant No analysis of catalytic mechanism Leitenmaier B, Küpper H (2011) Plant Cell & Environment 34, 208219 I) Protein Isolation: Sources (expression) of plant proteins Native expression (i (i.e. e in intact organs or in tissue culture) Advantages: - Correct folding - Correct C t post-translational tt l ti l processing i ((e.g. iinsertion ti off active ti centre, t phosphorylation, h h l ti glycosylation, cleavage, ...) - No cloning needed Disadvantages - Usuallyy lower yyield - Often many similar proteins in the same organisms difficulties with purification root freezing in liquid nitrogen and hydroponic plant cultivation in Küpper lab I) Protein Isolation: Sources (expression) of plant proteins Heterologous expression (e.g. in bacteria, yeast or insect cells) Advantages: - Usually higher yield - Specific expression of one protein in large amounts and possibly with specific tag (e g His-tag) (e.g. His tag) facilitates purification Disadvantages - Often problems with folding, in particular in the case of large proteins and integral membrane proteins p cofactors ((e.g. g iron-sulfur clusters)) is difficult or impossible, p - Insertion of complicated other post-translational modifications may be missing or different compared to the native protein possibly wrong conclusions about functions/mechanisms I) Protein Isolation: Methods for isolation Grinding in liquid nitrogen Used for hard tissues and cells like roots, stems, but also for hard-walled cells like some algae and cyanobacteria Low L ttemperature t protects t t the th proteins t i during d i grinding i di Time consuming (manual grinding) or requiring suitable machinery (expensive solution: lab mill, several thousand €, cheap solution: wheat mill, about 200 €) Liquid nitrogen cooled wheat mill in Küpper lab I) Protein Isolation: Methods for isolation Ultrasound homogenisation Used for soft tissues like some leaves, and a post-treatment after grinding Does not require freezing and thus may avoid artefacts of freezing, but may cause artefacts t f t by b heating h ti off the th sample l Requires expensive (usually several thousand €) machinery I) Protein Isolation: Methods for isolation French Press Used for individual cells (plant cell culture, algae or bacteria) without or with soft walls Does not require freezing and thus may avoid artefacts of freezing Requires R i very expensive i ((usually ll many th thousand d €) machinery hi From: www.diversified-equipment.com From: www.pegasusscientific.com I) Protein Isolation: Methods for isolation Lysis buffer Used only for bacteria or animal cells May cause degradation No N machinery hi needed d d From: bio-ggs.blogspot.com/2009/11/ggs-live-western... II) Protein Purification: Overview of Principles Separation by expression site Selective use of tissues or organelles Separation of soluble from membrane proteins by centrifugation Separation by size Ultrafiltration Ult filt ti Size exclusion chromatography Preparative native gel electrophoresis Separation by charge g chromatography g p y Ion exchange Isoelectric focussing (as chromatography, in solution or in gel electrophoresis) Separation by specific binding sites Metal affinity chromatography using natural or artificial metal binding sites Immuno-Chromatography Immuno Chromatography using immobilised antibodies Magnetic separation using magnetically tagged antibodies II) Protein Purification: Separation by size in native gels Principle: Small proteins are less retained by the fibers of the gel than large proteins, so that small proteins migrate faster foto of a green gel (Küpper et al., 2003, Funct Plant Biol) From: www.columbia.edu/.../c2005/lectures/lec6_09.html III) Protein Identification: Overview of Principles Size determination Size exclusion chromatography or SDS PAGE Comparison with expected size of protein (known e e.g. g from reference or cDNA) Western W t Blotting Bl tti Binding to specific primary antibody, detected via labelled or enzymatically active secondary antibody Biochemical assays in native gels de t cat o of o enzymes e y es by their t e characteristic c a acte st c act activity ty Identification Identification of metalloproteins by their metal content Mass Spectrometry Fragmentation of the protein, identification of fragment sizes, and subsequent comparison i tto a library lib off kknown ffragmentation t ti patterns tt N-terminal Seqencing (Edman degradation) Sequential chemical removal of individual amino acids from the N-terminus III) Protein Identification by assays in native gels Example p of a metalloprotein p identification in native g gels: Scanning g of the g gels with laser ablation inductively coupled mass spectrometry (LA-ICP-MS) for obtaining information about metal binding to individual bands of a gel from: Polatajko A, Azzolini M, et al (2007) JAnalAtomSpect 22, 878-887 III) Protein Identification by assays in native gels Example of a biochemical assay in native gels: identification of Cd-Carboanhydrase by its metal content combined with an in-gel carboanhydrase assay f from: Lane L TW, TW Morel M l FMM (2000) PNAS97, PNAS97 4627 4627-4631 4631 III) Protein Identification: Mass Spectrometry Principle 1) protein band is cut of out gel 2) protein is digested into peptides 3) mass spectrometry 4) comparison of the fragment sizes with a database 5) assignment of likely sequences to fragments 6) comparison of the fragment sequences with a database 7) result: list of proteins from the database that have a similar fragmentation pattern from: Gingras AC et al al, 2004 2004, J Physiol 563 563, 11 11-21 21 III) Protein Identification: Mass Spectrometry 3) mass spectrometry 4) comparison of the fragment sizes with a database from: http://www.med.monash.edu.au/biochem/facilities/proteomics/maldi.html http://www med monash edu au/biochem/facilities/proteomics/maldi html III) Protein Identification: Mass Spectrometry 3a) further fragmentation of one of the fragments from the digest in the mass spectrometer, then again mass spectrometry (MSMS) p of the fragment g sizes with a database 4a)) comparison from: http://www.med.monash.edu.au/biochem/facilities/proteomics/maldi.html III) Protein Identification: Mass Spectrometry 5) assignment of likely sequences to fragments 6) comparison of the fragment sequences with a database 7) result: list of proteins from the database that have a similar fragmentation pattern from: http://www.med.monash.edu.au/biochem/facilities/proteomics/maldi.html III) Protein Identification: Mass Spectrometry from: Gingras AC et al, 2004, J Physiol 563, 11-21 Limitations and artefacts contamination of gel bands with other proteins identification of the contamination (very common: keratin from skin!) instead of the protein of interest not all proteins have the same detection efficiency (problems e.g. if many cysteines present)) even small contaminations sometimes lead to wrong p g identifications not all proteins are in the databases database may show results that have a similar fragmentation pattern, but are otherwise unrelated III) Protein Identification: N-terminal sequencing Principle identification of the amino acid by y reversed phase HPLC and comparison with HPLC of standard mixture from: www.ufrgs.br/depbiot/blaber/section4/section4.htm III) Protein Identification: N-terminal sequencing Sample preparation 1) run acrylamide gel (denaturating or native) 2) blot onto PVDF (NOT nitrocellulose) membrane using glycine glycine-free free buffer 3) stain the membrane with ponceau red (CBB and other stains also work) 4) submit for sequencing Limitations and artefacts problems with contaminations in eukaryotic proteins, the N-terminus is often blocked (e.g. by methylation), which equ ed co complicated p cated de de-blocking b oc g p procedures. ocedu es Also so non-polymerised o po y e sed ac acrylamide y a de required remains in the gel can cause blocking, so let the gel polymerise over night Cys residues cannot be detected, and also glycosylated redidues may appear as blank spaces in the sequence IV) Protein Characterisation: Overview of Principles Size determination Charge determination Analysis of cofactors Analysis of the 3-dimensional Structure Activity tests IV) Protein Characterisation: Size determination Comparison with predicted size by native and denaturating gel electrophoresis and by size exclusion chromatography can show native oligomerisation, post-translational modification but also artefactual degradation/aggregation 200 KDa 116 KDa 97 KDa 66KDa 45KDa 31 KDa SDS gel and Western blot of TcHMA4 size shows post-translational p processing as cDNA sequence predicts 128kDa (Parameswaran, Leitenmaier et al., 2007, BBRC) IV) Protein Characterisation: Charge determination Isoelectric focussing (IEF) can reveal the isoelectric point where the net charge of the protein is zero, i.e. balance between protonation of carboxyl groups and deprotonation of amino groups is achieved pH alkaline region neutral t l region i acidic region from: commons.wikimedia.org IV) Protein Characterisation: Analysis of cofactors Identification and reactivity: UV/VIS Absorption and fluorescence Spectroscopy Metal content (about 30% of all proteins are metalloproteins!): AAS, ICP-MS/OES, EDX/PIXE/XRF UV/VIS spectrum of TcHMA4 Cadmium g causes binding ligand-metalcharge-transfer (LMCT) bands indicating cysteine ligation (Leitenmaier and Küpper unpublished) Küpper, IV) Protein Characterisation: Analysis of cofactors Metal ligands: EXAFS, EPR, heteronuclear NMR provide information about ligand types and their spatial arrangement around the active site (each of these techniques has different strenghts) EXAFS spectrum of Zn-TcHMA4 mixed S/N g , multiple p ligands, scattering shows histidine contribution (Leitenmaier and Küpper, unpublished) IV) Protein Characterisation: Analysis of the 3D Structure Circular Dichroism (CD) Spectroscopy From: www.proteinchemist.com/cd/cdspec.html information about proportions of secondary structure types in a protein particularly useful when X-ray crystallography and NMR are not applicable IV) Protein Characterisation: Analysis of the 3D Structure X-ray crystallography E Example: l iin th the nickel-binding i k l bi di transcription t i ti factor f t NikR, NikR th the mechanism h i was concluded l d d from the X-ray structure From: Schreiter ER et al., 2006, PNAS103, 13676-81 Ni binding causes RHH domains to rotate about the flexible interdomain linkers to orient their antiparallel -strands toward the same face of the repressor, allowing each to occupy the DNA major groove of an operator palindrome half half-site site Binding of Ni creates a surface of the MBD suitable for interacting with DNA by stabilization of a helix and a loop IV) Protein Characterisation: Activity tests Titration of an enzyme with its substrate(s) reveals binding constants and possible substrate b t t inhibition i hibiti Activation of a human Cu-ATPase Hung et al. Biochem.J. 2007, 401 TcHMA4 Fraction3 051108 350 Both are P1B-Type ATPases showing the same activation pattern by p y „„their“ metal after reconstitution into artificial lipid vesicles Zn Z Cd 300 turnover rate[1/s] 250 200 Activity of TcHMA4 150 (purified by Leitenmaier and Kü Küpper) ) 100 50 0 0,01 0,1 1 conc. metal[µM] 10 IV) Protein Characterisation: Activity tests Two-dimensional data, e.g. substrate concentration and temperature reveal insights into interactions between factors that are decisive for enzyme activity, e.g. temperature dependence of substrate binding and influence of the substrate on the thermostability of the enzyme Activity of TcHMA4 (Leitenmaier et al (2011) Biochimica et Biophysica Acta (Biomembranes) 1808, 2591-2599) Example: Flowchart of expression, isolation, purification and characterisation of the Cd/Zn-ATPase TcHMA4 Grow plants expressing TcHMA4 hydroponically, harvest and freeze roots Grind the frozen roots with isolation buffer in liquid q N2, thaw Centrifuge, g discard the supernatant p ((soluble pproteins)) and resuspend p the pellet (membrane proteins) with solubilisation buffer Centrifuge for removing insoluble residue, collect solubilised protein Immobilized Metal Affinity Chromatography on Ni-IDA column Identify: Quantify: In SDS Western-Blotting, gel via fluorescent f Edman Sequ. dye Metal binding: AAS/ICP, S/ C EXAFS, S UV/VIS Activity tests ( catalytical properties) The slides can be downloaded from the workgroup homepage www.uni-konstanz.de ni konstan de Department of Biology Biolog Workgroups Workgro ps Küpper lab, lab or directlyy http://www.uni-konstanz.de/FuF/Bio/kuepper/Homepage/AG_Kuepper_Homepage.html