Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
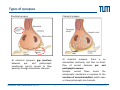
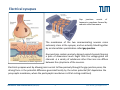
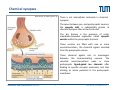
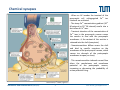
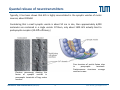
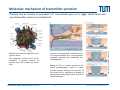
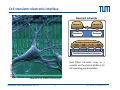
Synaptic transmission Purkinje cells in the cerebellum Synaptic endings cerebellar Purkinje cell The human brain contains at least 100 billion neurons, each with the ability to influence many other cells; highly efficient and sophisticated communication mechanisms are required → this is enabled by synapses, the functional contacts between neurons Study of activated synapse Biosensors and Bioelectronics (WS13/14) 68 Jose A. Garrido | [email protected] Synaptic transmission Biosensors and Bioelectronics (WS13/14) 69 Jose A. Garrido | [email protected] Types of synapses Chemical synapse Electrical synapse Synaptic cleft At chemical synapses, there is no intercellular continuity, and thus no direct flow of current between pre‐ and postsynaptic neurons. Synaptic current flows across the postsynaptic membrane in response to the secretion of neurotransmitters, which open or close postsynaptic ion channels. At electrical synapses, gap junctions between pre‐ and postsynaptic membranes permit current to flow passively through intercellular channels. Biosensors and Bioelectronics (WS13/14) 70 Jose A. Garrido | [email protected] Electrical synapses Gap junctions consist of hexameric complexes formed by subunits called connexons The membranes of the two communicating neurons come extremely close at the synapse, and are actually linked together by an intercellular specialization called gap junction. Neuroscience, 4th Edition, Figure 5.1a Gap junctions contain precisely aligned, paired channels forming a pore of dimensions much larger than the voltage‐gated ion channels → a variety of substances other than ions can diffuse between the cytoplasms of the neurons. Electrical synapses work by allowing ionic current to flow passively through the gap junction pores; the driving force is the potential difference generated locally by the action potential (AP depolarizes the presynaptic membrane, where the postsynaptic membrane is still at resting conditions) Biosensors and Bioelectronics (WS13/14) 71 Jose A. Garrido | [email protected] Electrical synapses Communication using gap junctions offers several advantages: ∙ current can flow in either direction, depending on which member of the couple pair is invaded by an action potential → bidirectional communication ∙ passive current flow across the junction is virtually instantaneous → extraordinarily fast transmission (without the typical delay of chemical synapses) Electrical synapses allow synchronization of electrical activity in hippocampal neurons. Neuron 1 and 2 connected through a electrical synapse. Generation of an AP in neuron 1 often result in a synchronized firing of an AP in neuron 2. Biosensors and Bioelectronics (WS13/14) Rapid transmission of signal at an electrical synapse of a crayfish. An AP in the presynaptic neuron causes postsynaptic neuron to be depolarized within a fraction of a millisecond. 72 Jose A. Garrido | [email protected] Chemical synapses Neuroscience, 4th Edition, Figure 5.1b There is not intercellular continuity in chemical synapses. The space between pre‐ and postsynaptic neurons, the synaptic cleft, is substantially greater at chemical synapses than at electrical ones The key feature is the presence of small, membrane‐bounded organelles called synaptic vesicles within the presynaptic terminal. These vesicles are filled with one or more neurotransmitters, the chemical signals secreted from the presynaptic neuron. These chemical agents acts as messengers between the communicating neurons. The secreted neurotransmitters open or close postsynaptic ligand‐gated ion channels after binding to specific receptor molecules, and thus eliciting an action potential in the postsynaptic membrane. Biosensors and Bioelectronics (WS13/14) 73 Jose A. Garrido | [email protected] Chemical synapses ‐ When an AP invades the terminal of the presynaptic cell, voltage‐gated Ca2+ ion channels are activated. ‐ The steep Ca2+ concentration gradient (10‐3 M external vs 10‐7 M internal) results into a fast influx of Ca2+ ions. ‐ Transient elevation of the concentration of Ca2+ ions in the presynaptic neuron causes the vesicles to fuse with the presynaptic membrane → the content of the vesicles is released into the cleft (exocytosis). ‐ Neurotransmitters diffuse across the cleft and bind to specific receptors on the membrane of the postsynaptic neuron, which causes ion channels of the postsynaptic membrane to open (or close). ‐ This neurotransmitter‐induced current flow alters the conductance and membrane potential of the postsynaptic neuron, increasing or decreasing the probability of action potential firing. Neuroscience, 4th Edition, Figure 5.3 Biosensors and Bioelectronics (WS13/14) 74 Jose A. Garrido | [email protected] The discovery of neurotransmitter release / 1926, Otto Loewi ‐ Two frogs’ hearts isolated and perfused; the perfusate flowing through the stimulated heart is collected and transferred to the second heart. ‐ Electrical stimulation of the vagus nerve slows the heartbeat of the stimulated heart → the second heart, even without stimulation, reduces its beating frequency Neuroscience, 4th Edition, Figure 5.4 ‐ The released substance was referred to as “vagus substance”; later on was shown to be acetylcholine (ACh) Biosensors and Bioelectronics (WS13/14) 75 Jose A. Garrido | [email protected] Neurotransmitters Criteria that define a neurotransmitter 1. The molecule must be present within the presynaptic neuron 2. The substance must be released in response to presynaptic depolarization and the release must be Ca2+ dependent 3. Specific receptors for the substance must be present on the postsynaptic cell Neuroscience, 4th Edition, Figure 6.1 Biosensors and Bioelectronics (WS13/14) 76 Jose A. Garrido | [email protected] Postsynaptic receptors families Ligand‐gated ion channels (ionotropic): combine transmitter binding and channel functions into a single molecular entity. They are typically made up of 4 to 5 protein subunits, each of which contribute to the pore of the ion channel Neuroscience, 4th Edition, Figure 5.23 G‐protein‐coupled receptors (metabotropic): ion movement results from one or more metabolic steps. Do not have ion channels as part of their structure. Affect channels by the activation of G‐proteins, which dissociate from the receptor and interact directly with the channel or bind to other effector proteins, such as enzymes, that make intracellular messengers →open/close ion channels Biosensors and Bioelectronics (WS13/14) 77 Jose A. Garrido | [email protected] Quantal release of neurotransmitters A study case: ACh release at neuromuscular junctions “Quantal components of the end‐plate potential”, del Castillo J & Katz B, J Physiol (1954) 124, 560–573. The experiment ∙ The axon of the motor neuron innervating the muscle fiber is stimulated with an extracellular electrode ∙ An intracellular microelectrode is inserted into the postsynaptic muscle cell to record its electrical responses ∙ An action potential in the presynaptic motor neuron can elicit a transient depolarization of the postsynaptic muscle fiber: this is called an end plate potential (EPP) End plate of a neuromuscular junction ∙ EPP are typically above the threshold, and can elicit a postsynaptic action potential, which causes the muscle fiber to contract. Acetylcholine ∙ There exist a pronounced delay between the stimulation of the presynaptic motor neuron and the postsynaptic action potential, which is typical of all chemical synapses. Biosensors and Bioelectronics (WS13/14) 78 Jose A. Garrido | [email protected] Quantal release of neurotransmitters A study case: ACh release at neuromuscular junctions “Quantal components of the end‐plate potential”, del Castillo J & Katz B, J Physiol (1954) 124, 560–573. ∙ In the absence of stimulation of the presynaptic neuron, spontaneous changes in the muscle cell membrane are observed. ∙ These potential changes have the same shape than EPPs, but their magnitude is much smaller, ~ 0.5mV (50 mV for EPPs) ∙ EPPs and these small events are sensitive to pharmacological agents that block postsynaptic ACh receptors. → the spontaneous events are called miniature end plate potential (MEPPs) low [Ca2+] bath ∙ If the neuromuscular junction is bathed in a low [Ca2+] solution, neurotransmitter secretion is strongly reduced → reduction of the EPP magnitude below the threshold for AP. ∙ In this situation, very small EPP are recorded, with a magnitude varying from trial to trial, and of similar shape of the spontaneous MEPP. Biosensors and Bioelectronics (WS13/14) 79 Jose A. Garrido | [email protected] Quantal release of neurotransmitters A study case: ACh release at neuromuscular junctions “Quantal components of the end‐plate potential”, del Castillo J & Katz B, J Physiol (1954) 124, 560–573. No EPP in response to stimulation Quantized distribution of EPP amplitudes evoked in a low Ca2+ solution Prediction of statistical model Peaks of EPP amplitudes tend to occur in integer multiples (“quanta”) of the mean amplitude of MEPPS. The number of such quanta contained in a given EPP varied randomly with time. “Quantal” fluctuations in the amplitude of EPPs indicate that EPPs are made up of individual units, each equivalent to a MEPP response occurs in units about the size of single MEPPs distribution of spontaneous MEPP amplitudes Biosensors and Bioelectronics (WS13/14) 80 Jose A. Garrido | [email protected] Quantal release of neurotransmitters A Poisson distribution The number of quanta that occurred for a given EPP as a result of nerve stimulation can be described by a Poisson distribution (del Castillo & Katz, 1954) P ( n, ) n n! e where n=0, 1, 2, 3, … μ corresponds to the mean value or the expected value of the number of occurrences obtained from a large population of the same event The Poisson distribution gives the probability for the number of occurrences (n=0,1,2,…) of a given event taking place within a certain time or space ‐ the events occur independently of each other ‐ the probability for each single event is small It is assumed that each EPP is composed of the sum of a number n of units (“quanta”), with mean amplitude and standard deviation identical to the mean amplitude and standard deviation of the MEPP m mean amplitude of EPP number of units that on average enter on a single EPP as a mean amplitude of MEPP result of nerve stimulation if the number of units that enter in a EPP follows a Poisson distribution, the mean value m should correspond to the mean value of the distribution → m=μ Biosensors and Bioelectronics (WS13/14) 81 Jose A. Garrido | [email protected] Quantal release of neurotransmitters A Poisson distribution In the case of a Poisson distribution, the mean value can be obtained by determining the probability that the event does not occur P (0, ) e number of absent responses number of nerve impulses number of absent responses number of nerve impulses ln Thus, in the case of a Poisson distribution number of absent responses mean amplitude of EPP ln mean amplitude of MEPP number of nerve impulses slope 1 mean amplitude of EPP mean amplitude of MEPP Biosensors and Bioelectronics (WS13/14) 82 Jose A. Garrido | [email protected] Quantal release of neurotransmitters Typically, it has been shown that ACh is highly concentrated in the synaptic vesicles of motor neurons, about 100mM. Considering that a small synaptic vesicle is about 50 nm in size, then approximately 6,000 molecules are contained in a single vesicle. Of them, only about 1500 ACh actually bind to postsynaptic receptors (10‐20% efficiency) Fine structure of vesicle fusion sites in presynaptic terminals. Proteinaceous structures arrange vesicles in rows Electron microscopy showing the fusion of synaptic vesicles in presynaptic terminals of frog motor neurons Biosensors and Bioelectronics (WS13/14) 83 Jose A. Garrido | [email protected] Role of Ca2+ The voltage clamp detects current flowing across the presynaptic membrane when the membrane potential is depolarized. Pharmacological agents that block currents flowing through Na+ and K+ channels reveal a remaining inward Ca2+ current, which triggers transmitter secretion → EPP Upon treatment with cadmium (a Ca2+ channel blocker) eliminates both the presynaptic Ca2+ current and the postsynaptic response . Neuroscience, 4th Edition, Figure 5.10 Biosensors and Bioelectronics (WS13/14) 84 Jose A. Garrido | [email protected] Molecular mechanism of transmitter secretion Model of the molecular organization of a synaptic vesicle. The cytoplasmic surface of the vesicle membrane is densely covered by proteins, only 70% of which are shown here Biosensors and Bioelectronics (WS13/14) Neuroscience, 4th Edition, Figure 5.14 Precisely how an increase in presynaptic Ca2+ concentration goes on to trigger vesicle fusion and neurotransmitter release is not understood Proteins in the presynaptic membrane and in the vesicle membrane form macromolecular complexes bringing both membranes into close apposition. Binding of Ca2+ to receptor proteins in the vesicle (synaptotagmin) seems to induce chemical changes (allowing the insertion of this protein into a membrane & binding to other proteins) that produce the final fusion of these membranes . 85 Jose A. Garrido | [email protected] Postsynaptic membrane permeability changes The binding of neurotransmitters (e.g. ACh) to postsynaptic receptors opens ion channels in the postsynaptic membrane Outside‐out patch clamp measurement of single ACh receptor current Individual ACh receptors only open briefly The opening of a large number of channels is synchronized by the brief duration of ACh secretion from presynaptic terminals. End plate current (EPC): macroscopic current resulting from the summed opening of many ion channels The EPC is normally inwards, it causes the postsynaptic membrane potential to depolarize. This depolarization change is the EPP, which might elicit an AP. Biosensors and Bioelectronics (WS13/14) 86 Jose A. Garrido | [email protected] Cell‐transistor electronic interface Neuronal networks neuron 1 neuron 2 Stimulator Receptor Extended neuronal network Distributed stimulator/receptors P. Fromherz Field Effect Transistor array as a versatile and functional platform for cell recording and stimulation Neurons on CMOS transistors Biosensors and Bioelectronics (WS13/14) 87 Jose A. Garrido | [email protected]