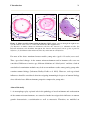
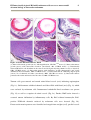
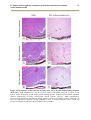
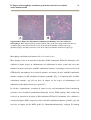
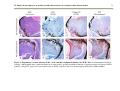
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
University of Veterinary Medicine Hannover The Role of Macrophages in Murine Intraocular Melanoma THESIS Submitted in partial fulfilment of the requirements for the degree Doctor of Veterinary Medicine Doctor medicinae veterinariae (Dr. med. vet.) by Marta M. Kilian Dramburg Hannover 2015 Academic supervision: Prof. C. Pfarrer Department of Anatomy University of Veterinary Medicine Hannover Prof. F. G. Holz Department of Ophthalmology University Hospital Bonn 1. Referee: Prof. C. Pfarrer / Prof. F. G. Holz 2. Referee: Prof. H.-J. Schuberth Day of oral examination: 26.11.2015 Contents List of Publications..................................................................................................................... List of Abbreviations.................................................................................................................. I Introduction.......................................................................................................................... 11 Human uveal melanoma....................................................................................................... 11 The tumour microenvironment and tumour associated macrophages.................................. 14 Tumour vascularisation ........................................................................................................ 17 Animal models ..................................................................................................................... 20 Aims of the study ................................................................................................................. 21 II Materials and Methods ...................................................................................................... 23 Animals ................................................................................................................................ 23 HCmel12 melanoma cell line............................................................................................... 24 Murine macrophage cell line................................................................................................ 25 Macrophage polarisation in vitro ......................................................................................... 25 Macrophage depletion .......................................................................................................... 27 Intraocular tumour cell injection .......................................................................................... 28 Histology and immunohistochemistry ................................................................................. 29 Flow cytometry analysis....................................................................................................... 31 Ex vivo imaging.................................................................................................................... 31 Statistical analysis ................................................................................................................ 32 Histology and immunohistochemistry of Hgf-Cdk4R24C mice eyes..................................... 32 III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumour biology of intraocular melanoma .......................................................................................... 33 Abstract ................................................................................................................................ 34 Introduction .......................................................................................................................... 35 Materials and Methods ......................................................................................................... 37 Animals ............................................................................................................................ 37 HCmel12 melanoma cell line........................................................................................... 38 Clinical procedures........................................................................................................... 39 Histology .......................................................................................................................... 39 Results .................................................................................................................................. 41 Histology of intraocular HCmel12 melanoma ................................................................. 41 Immunohistochemistry of intraocular HCmel12 melanoma............................................ 45 Metastases of HCmel12 melanoma.................................................................................. 46 Discussion ............................................................................................................................ 47 References ............................................................................................................................ 50 IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model ....................................................................................................................................... 53 Abstract ................................................................................................................................ 54 Introduction .......................................................................................................................... 56 Materials and Methods ......................................................................................................... 58 Animals ............................................................................................................................ 58 Mouse model of intraocular melanoma............................................................................ 59 Macrophage depletion (experiment 1) ............................................................................. 60 Macrophage polarisation (experiment 2) ......................................................................... 60 Histology and immunohistochemistry ............................................................................. 61 Flow cytometry analysis................................................................................................... 62 Statistical analysis ............................................................................................................ 63 Results .................................................................................................................................. 64 Macrophage depletion model (experiment 1) .................................................................. 64 Macrophage polarisation model (experiment 2) .............................................................. 67 Flow cytometry analysis................................................................................................... 72 Discussion ............................................................................................................................ 74 The role of TAM .............................................................................................................. 75 The impact of M2 macrophages....................................................................................... 76 The impact of age............................................................................................................. 78 Summary .......................................................................................................................... 79 Acknowledgements .............................................................................................................. 80 References ............................................................................................................................ 81 VI Origin of intratumoural vascular structures in a murine ocular tumour model........ 84 Abstract ................................................................................................................................ 85 Introduction .......................................................................................................................... 86 Materials and Methods ......................................................................................................... 88 Animals ............................................................................................................................ 88 Mouse model of intraocular melanoma............................................................................ 88 Ex vivo imaging................................................................................................................ 89 Histology and immunohistochemistry ............................................................................. 89 Statistical analysis ............................................................................................................ 90 Results .................................................................................................................................. 91 Ex vivo imaging................................................................................................................ 91 Histology and immunohistochemistry ............................................................................. 92 Discussion ............................................................................................................................ 95 Acknowledgements ............................................................................................................ 100 V Overexpression of Hepatocyte Growth Factor and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma103 Letter to the Editor ............................................................................................................. 104 References .......................................................................................................................... 108 VII Discussion....................................................................................................................... 109 Animal models ................................................................................................................... 109 Tumour associated macrophages in intraocular melanoma ............................................... 111 Impact of macrophage polarisation .................................................................................... 112 Impact of advanced age...................................................................................................... 114 Tumour associated macrophages and angiogenesis ........................................................... 116 The origin of tumour vessels.............................................................................................. 117 Outlook............................................................................................................................... 120 Summary ............................................................................................................................ 120 VIII References..................................................................................................................... 122 IX Summary.......................................................................................................................... 129 X Zusammenfassung ............................................................................................................ 132 XI Acknowledgements ......................................................................................................... 136 List of publications List of Publications Parts of this thesis are published or are in revision in following journals: Original Papers 1. Marta M. Kilian, Karin U. Loeffler, Christiane Pfarrer, Frank G. Holz, Christian Kurts, Martina C. Herwig Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumour biology of intraocular melanoma Current Eye Research, ISSN: 0271-3683 print / 1460-2202 online DOI: 10.3109/02713683.2015.1004721 2. Marta M. Kilian, Karin U. Loeffler, Christian Kurts, Tobias Hoeller, Christiane Pfarrer, Frank G. Holz, Martina C. Herwig Impact of macrophages on tumour growth characteristics in a murine ocular tumour model Experimental Eye Research manuscript submitted 3. Marta M. Kilian, Karin U. Löffler, Christiane Pfarrer, Tobias Höller, Frank G. Holz, Daniela Wenzel, Martina C. Herwig Origin of intratumoural vascular structures in a murine ocular tumour model Journal manuscript in preparation Letter to the Editor (Own data) 1. Marta M. Kilian, Karin U. Löffler, Frank G. Holz, Thomas Tüting, Martina C. Herwig Hgf-Cdk4 mice fail as a spontaneous ocular melanoma model but exhibit unusual histomorphologic ocular features Melanoma Research Accepted List of publications Contributions to Congresses 1. Marta M. Kilian, Karin U. Loeffler, Frank G. Holz, Christiane Pfarrer3, Christian Kurts, Martina C. Herwig Impact of macrophages and age on tumour growth characteristics in a murine ocular tumour model Poster at the 18th ECCO - 40th ESMO European Cancer Congress, Vienna, Austria Abstract Number 172 2. Marta M. Kilian, Karin U. Loeffler, Frank G. Holz, Christiane Pfarrer3, Christian Kurts, Martina C. Herwig The effects of polarised macrophages on melanoma growth characteristics in vitro and in vivo Poster at the ARVO 2015 Annual Meeting in Denver, Colorado, USA Kilian MM, et al. IOVS 2015,56:ARVO E-Abstract 5321 3. Marta M. Kilian, Karin U. Löffler, Frank G. Holz, Christiane Pfarrer, Tobias Höller, Christian Kurts, Martina C. Herwig Der Einfluss von Makrophagen auf das Tumorwachstum im Mausmodel für das intraokuläre Melanom Talk at the DOG 2014 in Leipzig, Germany; Abstract Number Fr06-05 4. Martina C. Herwig, Marta M. Kilian, Frank G. Holz, Daniela Wenzel, Karin U. Löffler Origin of Tumour-Associated Vessels and Vasculogenic Mimicry in an Intraocular Murine Melanoma Model Poster at the ARVO 2014 Annual Meeting in Orlando, Florida, USA Kilian MM, et al. IOVS 2014,55:ARVO E-Abstract 5066 5. Karin U. Löffler, Marta M. Kilian, Frank G. Holz, Thomas Tüting, Martina C. Herwig Morphologic and immunohistochemical features in the ageing Hgf-Cdk4 mouse eye Poster at the ARVO 2014 Annual Meeting in Orlando, Florida, USA Kilian MM, et al. IOVS 2014,55:ARVO E-Abstract 5071 6. Marta M. Kilian, Karin U. Loeffler, Hans E. Grossniklaus, Frank G. Holz, Christiane Pfarrer, Christian Kurts, Martina C. Herwig The effects of macrophages on tumour growth characteristics in a mouse-model of intraocular melanoma Poster at the ARVO 2014 Annual Meeting in Orlando, Florida, USA Kilian MM, et al. IOVS 2014,55:ARVO E-Abstract 5070 List of publications 7. Marta M. Kilian, Karin U. Löffler, Hans E. Grossniklaus, Frank G. Holz, Christian Kurts, Marina C. Herwig Evaluation einer neuen murinen Hautmelanomzelllinie als potentielles Mausmodell für das intraokuläre Melanom Poster at the DOG 2013 Annual Meeting in Berlin, Germany; Poster award winner Abstract Number A-572-0008-00718 8. Marta M. Kilian, Karin U. Loeffler, Hans E. Grossniklaus, Frank G. Holz, Christiane Pfarrer, Christian Kurts, Martina C. Herwig Comparison of Two Melanoma Cell Lines as Mouse-Models of Uveal Melanoma Poster at the ARVO 2013 Annual Meeting in Seattle, Washington, USA; Kilian MM, et al. IOVS 2013,54:ARVO E-Abstract 4203 List of Abbreviations Ab AEC BAP1 BC199 CDK4 CSC DMEM/F12 DT101 ECM ELISA FACS FCS GFP GNA11 GNAQ H&E HGF HMB45 HRP IFNγ IHC IL LPS MФ M1 M2 MVD NEAA NIH NK PAS PBS PEDF RAS RPE RPMI 1640 SCID TAM TEC TBS TGFβ TNFα UM VEGF VM Antibody 3-amino-9-ethylcarbazole BRCA1 associated protein-1 Melanoma antigen clone Cyclin-dependent kinase 4 Cancer stem cells Dulbeccos Modified Eagle Medium /F12, cell culture medium Melanoma antigen clone Extracellular matrix Enzyme-linked immunosorbent assay Fluorescence-activated cell sorting Fetal calve serum Green fluorescent protein Guanine nucleotide binding protein (G protein), alpha 11 Guanine nucleotide binding protein (G protein), q polypeptide Hematoxylin and eosin Hepatic growth factor Human melanoma black 45 antigen Horseradish peroxidase Interferon γ Immunohistochemistry Interleukin Lipopolysaccharide Macrophages Macrophages with a functional M1 polarisation Macrophages with a functional M2 polarisation Microvascular density Non-essential amino acid National Insitutes of Health Natural killer (cells) Periodic acid-Schiff Phosphate buffered saline Pigment epithelium-derived factor Rat sarcoma, ongcogene Retinal pigment epithelium Cell culture medium Roswell Park Memorial Institute Severe combined immunodeficiency Tumour associated macrophages Tumour endothelial cells Tris buffered saline Transforming growth factor β Tumour necrosis factor α Uveal melanoma Vascular endothelial growth factor Vasculogenic mimicry 11 I Introduction Human uveal melanoma Uveal melanoma is the most common primary intraocular tumour in the adult Caucasian population. It occurs usually in the elderly with an incidence of ~ 2-8 cases per million/year (Virgili, Gatta et al. 2007; Singh, Turell et al. 2011; Mallone, De Vries et al. 2012). Other ethnities exhibit a far lower incidence rate (Eagle 2013). Uveal melanoma arises from melanocytes of the choroid, the stroma of the ciliary body and/or the iris. It can develop from a pre-existing nevus or denovo. However, intraocular tumours may be of primary or of metastatic origin. Uveal metastases represent the most frequent ocular malignancy. They most commonly derive from breast (in female) or lung (in male) carcinoma but may also originate from prostate, skin, colon or other carcinomas (reviewed in (Arepalli, Kaliki et al. 2015)). Clinical characteristics of primary uveal melanoma are defined as tumour growth with a thickness of ≥ 3 mm, lipofuscin accumulation, subretinal fluid/serous retinal detachment and visual symptoms (Fig. 1) (Shields, Manalac et al. 2014). Figure 1. Cross section of an enucleated human eye with a primary uveal melanoma (left) and schematic drawing of an eye with uveal melanoma (right). 12 I Introduction Local treatment of the primary tumour is very promising and depends on tumour characteristics such as size and localisation (reviewed in (Singh and Singh 2012)). It ranges from enucleation of the globe to eye-saving strategies like local radiation therapy (brachytherapy), thermotherapy or proton beam therapy of the tumour. In about 40% of cases of uveal melanoma metastases develop within 10 years (Singh, Shields et al. 2001). A special characteristic of intraocular melanomas is their hematogenous pathway of metastasis due to the absence of lymphatic vessels in the eye. The metastases show a predisposition to the liver and are rarely detected in the lungs or other sites (Singh, Shields et al. 2001). At the time of initial diagnosis the ocular tumour often already has developed micrometastases which may remain dormant for decades (Bakalian, Marshall et al. 2008). However, once micrometastases undergo an angiogenic switch and start to grow, metastatic disease develops. Prognosis worsens dramatically upon metastatic disease due to lack of effective systemic treatment options (Kujala, Makitie et al. 2003). Many studies have focused on prognostic parameters of uveal melanoma to estimate the risk of metastasis and patient’s outcome. Such clinical and histopathological prognostic factors include (advanced) patient age, primary tumour size (diameter and prominence), cell type, ciliary body involvement, mean vascular density (MVD), extravascular matrix patterns/vasculogenic mimicry (VM) and infiltrating inflammatory cell type, in particular macrophages (Fig. 2) (Foss, Alexander et al. 1996; Folberg, Mehaffey et al. 1997; Shields, Kaliki et al. 2013; Kaliki, Shields et al. 2015). Therefore, histologic examination of enucleated eyes harbouring an uveal melanoma may provide prognostic information. In addition, molecular gene analysis of uveal melanoma biopsies and enucleation specimens became available recently, providing relevant prognostic information with regard to development of metastases and patient’s outcome. Such analysis can distinguish two I Introduction 13 prognostic molecular subsets of primary uveal melanoma: class 1 tumours with low metastatic risk and class 2 tumours with high metastatic risk (Onken, Worley et al. 2004). Chromosomal analysis and genetic profiling were found to be more reliable indicators for patient’s prognosis than histopathologic characteristics (Onken, Worley et al. 2012; Field and Harbour 2014). However, the histologic parameters reflect uveal melanoma growth characteristics and allow for their evaluation in animal models, which may deliver important insights into tumour biology, progression and metastasis. Figure 2: Prognostic histological features of uveal melanoma (A+B) An epitheloid cell type (A) has a worse prognosis than a spindle cell type (B, H&E); (C) The occurrence of vasculogenic mimicry and extravascular matrix patterns is associated with death from metastatic disease in uveal melanoma (PAS w/o hematoxylin); (D) A high mitotic activity is considered as a prognostic factor in many solid tumours including uveal melanoma (IHC Ser-10 antibody); (E) Tumour associated macrophages play an important role in the pathogenesis of uveal melanoma and their occurrence is associated with patients’ prognosis (IHC, CD68 [left] and CD68 + CD163 antibody[right]); (modified from (Damato, Eleuteri et al. 2011) and (Herwig, Bergstrom et al. 2013)). I Introduction 14 Several genetic anomalies (e.g. monosomy 3, GNAQ, GNA11, Bap1, class 2 gene expression profile) influencing tumour progression and metastasis have been identified (Sisley, Rennie et al. 1990; Prescher, Bornfeld et al. 1996; Onken, Worley et al. 2004; Ehlers, Worley et al. 2008). But unlike cutaneous or conjunctival melanoma, mutations in B-RAF, RAS or KIT genes occur rarely in uveal melanoma (Griewank, Westekemper et al. 2013). Characteristic mutations differ between uveal and cutaneous melanoma and even among tumours itself, accounting for different progression and metastatic behaviour (Mehnert and Kluger 2012). Metastatic dissemination is a complex multistep process with local invasion, intravasation, colonisation of distant tissues and adaptation to a foreign microenvironment at the secondary site. As metastatic potential is linked to many biological features of the primary tumour, basic research on the primary tumour microenvironment may offer new implications for strategies against metastasis and identify new therapeutic targets. The tumour microenvironment and tumour associated macrophages Each tumour has its own microenvironment. It is composed of neoplastic and non-neoplastic cells as well as the stroma and it involves their functional interactions through different cytokines, chemokines or growth factors. Such interactions may withhold tumour growth or create a synergistic interplay and enhance tumour progression as well as metastasis (Catalano, Turdo et al. 2013). Recent data showed that hypoxia is a crucial factor of the microenvironment. It can induce changes in gene expression of tumour cells and/or nonneoplastic cells and it thereby may influence apoptosis, invasion and angiogenesis via different mechanisms (Bronkhorst, Jehs et al. 2014). Hypoxia-induced secretion of chemotactic cytokines or growth factors recruits blood leucocytes into the tumour which in turn further orchestrate the microenvironment. Tumour associated inflammation is one of the I Introduction 15 hallmarks of cancer and includes the presence of infiltrating leucocytes. Main players of the inflammatory infiltrate in many tumours - like uveal melanoma - are tumour associated lymphocytes and macrophages (Sica and Mantovani 2012). To overcome hypoxia tumour cells and tumour associated macrophages (TAM) secrete pro-angiogenic factors in order to enhance angiogenesis and other strategies of tumour vascularisation (e.g. vasculogenic mimicry (Fig. 2C)). An inflammatory and pro-angiogenic microenvironment favours tumour growth and is thus correlated with a poor prognosis. Further, the direct crosstalk between melanoma cells and TAM may either enhance or reduce tumour growth (Hussein 2006). TAM are an important component of the leukocytic infiltrate in tumours and are believed to play a major role in malignant neoplasms (Fig. 3). In most solid tumours, including uveal melanoma, high numbers of TAM correlate statistically significant with poor prognosis (Allavena, Sica et al. 2008). In uveal melanoma, they are further associated with monosomy 3, several histopathologic features such as a high mean vascular density (MVD) or certain extravascular matrix patterns, and thus with an aggressive tumour phenotype (Folberg, Mehaffey et al. 1997; Makitie, Summanen et al. 2001; Hendrix, Seftor et al. 2003; Toivonen, Makitie et al. 2004; Maat, Ly et al. 2008). Different functional macrophage phenotypes (polarisation) exist, which are influenced by the present microenvironmental stimuli (Arnold, Henry et al. 2007). The TAM polarisation phenotype is determined by cytokines produced by cells in the tumour microenvironment (Arnold, Henry et al. 2007). It may range within a broad spectrum from M1 to M2 macrophages (Jager, Ly et al. 2011). M1-polarised macrophages exhibit pro-inflammatory, tumouricidal and anti-angiogenic qualities with the potential to suppress tumour growth and dissemination (Sica and Mantovani 2012). In nonprogressing tumours, TAM mainly resemble the M1 type and exhibit anti-tumour activity. In malignant and advanced tumours, TAM are rather polarised toward the M2 phenotype that I Introduction 16 instead favours tumour malignancy (Biswas and Mantovani 2010). Tumour driven macrophage polarisation tends to specialise TAM into the M2 phenotype with pro-angiogenic, immunosuppressive and tumour-favouring properties. TAM are a source and target for various cytokines and chemokines within the tumour microenvironment (Fig. 3). Figure 3: Role of tumour associated macrophages (TAM) within the tumour microenvironment. TAM are a source and target for various cytokines and chemokines which regulate tumour growth, progression and invasion; they further interact with other components like the stroma as well as with other players of the adaptive immunity (modified from (Mantovani, Sozzani et al. 2002)). M2-polarised TAM exhibit characteristic cytokine expression levels of IL-10high, IL-12low, TNFαlow, IFNγlow and TGF-βhigh (Mantovani, Sozzani et al. 2002). A high M2/M1 ratio in uveal melanomas was shown to be related to worse prognosis and to histologic negative prognostic parameters (Herwig, Bergstrom et al. 2013). I Introduction 17 With increasing age, changes in the capability of the innate immunity induce a shift of the macrophage secretory profile towards a M2 profile (Gomez, Nomellini et al. 2008). Accordingly, macrophages from geriatric mice are particularly sensitive to signals that promote their M2-polarisation, which in turn may promote tumour development (Jackaman, Radley-Crabb et al. 2013). A M2-dominated tumour microenvironment comprises proangiogenic factors such as vascular endothelial growth factor (VEGF) and other growth factors, angiopoietins, certain interleukins and matrix metalloproteinases (Schmid and Varner 2010). Such a microenvironment is essential for tumour growth, tumour vascularisation and subsequently a basis for metastasis in uveal melanoma. Thus, TAM may promote a microenvironment which enhances angiogenesis, extracellular matrix remodelling and immune suppression (Mantovani, Sozzani et al. 2002). Tumour vascularisation Angiogenesis is a crucial point in the development of most solid tumours, including uveal melanoma. Solid tumours are dependent on constant blood supply to ensure tumour growth beyond diffusion limits and to satisfy the subsequent need for oxygen and nutrients. To address this demand tumours use different forms of vascularisation which represents one of the hallmarks of cancer (Hanahan and Weinberg 2011). One source of initial blood supply may be angiotropism and the incorporation of pre-existing vessels from surrounding structures (co-option) - in case of intraocular melanoma this includes structures such as the uvea. Second, the tumour may induce growth of new vessels by stimulating sprouting or intussusception from pre-existing host vessels (angiogenesis) or de novo formation of vessels by recruiting endothelial progenitor cells (vasculogenesis) (Fig. 4). I Introduction 18 Figure 4: Strategies of tumour vascularisation Co-option – colonisation of existing vessels; Angiogenesis – branching and colonisation of existing host vessels; vasculogenesis – formation of new vessels (de novo) from circulating endothelial progenitor cells (migrating from the bone marrow); vasculogenic mimicry - tumour cells lining channels of extracellular matrix forming vasculogenic networks (modified from (Burrell and Zadeh 2012)). Recently, a slowly increasing number of studies in different tumour entities reported vessel formation out of cancer cells (Ricci-Vitiani, Pallini et al. 2010; Wang, Chadalavada et al. 2010; Ohmura-Kakutani, Akiyama et al. 2014; Chroscinski, Sampey et al. 2015). Hypothetically, such tumour-derived blood vessel cells originated from multipotent cancer stem cells. However, their existence and function in tumour vascularisation remain subject to ongoing discussion. The essential role of sprouting angiogenesis in solid tumours and its potential therapeutic implications were first described by Folkman and coworkers (Folkman 1971). Many preclinical experiments indicated so far that solid tumours were profoundly angiogenesisdependant. However, the long-held belief that cancer patients can be effectively treated with I Introduction 19 anti-angiogenic therapy to prevent tumour progression and metastasis diminished after disappointing clinical results. Heterogeneity and complexity of tumours as well as alternative vascularisation strategies apart from angiogenesis may be a reason for that (Verheul, Voest et al. 2004). However, anti-angiogenic compounds such as bevacizumab (anti-VEGF antibody) still represent an adjuvant therapeutic option in patients with certain metastatic cancers such as breast carcinomas (Keating 2014). Alternative forms of primary tumour vascularisation in aggressive tumours may include mosaic vessels (tumour- and endothelial-lined vasculature) (Chang, di Tomaso et al. 2000), tumour-derived endothelial-like vessels, tumour-lined vascular channels (Timar and Toth 2000), and vasculogenic mimicry (Maniotis, Folberg et al. 1999). Vasculogenic mimicry (VM) - which was initially observed in uveal melanoma as well as in metastatic cutaneous melanoma (Maniotis, Folberg et al. 1999) - may represent an alternative perfusion to angiogenesis. VM is characterised by vasculogenic-like non-endothelial channels rich of PAS-positive extracellular matrix (ECM). It exhibits high expression levels of ECM related molecules (e.g. matrix-metalloproteinases, VE-cadherin, VEGF-A) and is lined by tumour cells (Fig. 4) (Seftor, Hess et al. 2012). VM is mainly found in aggressive and fast growing tumours in which tumour cell plasticity and the hypoxic microenvironment contribute to formation of this alternative perfusion pathway. Aggressive melanoma cells are potentially able to modify their surrounding microenvironment towards VM and thereby adapt to rapid tumour growth and overcome hypoxia (Hendrix, Seftor et al. 2003). The formation of microcirculatory patterns of VM, like back-to-back loops or parallel vessels with cross-linkings, are associated with death from metastatic disease in uveal as well as in cutaneous melanoma (Folberg, Mehaffey et al. 1997). I Introduction 20 Animal models Considering aforementioned characteristic features of uveal melanoma like hematogenous spread, different strategies of vascularisation, the inflammatory microenvironment and genetic attributes, a meaningful mouse model should offer similar properties. Since no spontaneous primary uveal melanomas in wild type mice have been described yet, all existing mouse models have restrictions or limitations. Different models have been developed including transgenic mice, intraocular inoculation with cutaneous melanoma cells into wild type mice, injection of human uveal melanoma cells into mice with severe combined immunodeficiency (SCID) or induction of uveal proliferations by chemicals, radiation or viruses (Dithmar, Albert et al. 2000). Pigmented intraocular tumours arising in transgenic mice were identified to be either of retinal pigment epithelium origin (Syed, Windle et al. 1998; Albert, Kumar et al. 2004) or as small uveal tumours which failed to metastasise to the liver (Kramer, Powell et al. 1998; Tolleson, Doss et al. 2005; Latendresse, Muskhelishvili et al. 2007). Injection of human uveal melanoma cells into immunodeficient mice also resulted in non-metastatic disease (Mueller, Maniotis et al. 2002) and thus does not allow to study influences and immunologic interactions between the tumour and the host. The best reasonable compromise and an established mouse model so far was created and established by injecting murine, cutaneous B16LS9 melanoma cells into the vitreous of C57Bl/6 mice, generating an ocular tumour with metastatic dissemination to the lungs and the liver (Fig. 5) (Diaz, Rusciano et al. 1999; Dithmar, Albert et al. 2000). 21 I Introduction C C C I I I I I L L M R R R A B C Figure 5: Mouse model of intraocular melanoma; H&E stained sections through the right mouse eye, R: retina, L: lens, I: iris, C: cornea, M: melanoma, 40x magnification. (A) Injection of murine cutaneous melanoma cells into the vitreous by a schematic needle; (B) Injected melanoma cells distribute throughout the vitreous after injection, arrow: point of previous injection; (C) Solid intraocular melanoma, nine days after tumour cell injection. For most of the above mentioned mouse models young mice (aged 8-12 weeks) were used. Thus, age-related changes in the tumour microenvironment and in immune cells were not considered. Differences in mice age, different definitions of “elderly mice” and thus a lack of standardised examination methods may lead to non-uniform results among study groups who examine tumour biology (Jackaman, Radley-Crabb et al. 2013). However, such age-related influences should be considered when investigating immunological aspects of tumour biology since old mice have different immune properties compared to young mice. Aims of the study 1. As macrophages play a pivotal role in the pathology of uveal melanoma and orchestration of the tumour microenvironment, we wanted to further investigate their influence on tumour growth characteristics, vascularisation as well as metastasis. Therefore, we modified an I Introduction 22 established B16 model in CX3CR1+/GFP mice (chapter III, Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumour biology of intraocular melanoma). 2. To determine the role of macrophages and their polarisation, we examined histopathological tumour characteristics of macrophage-depleted mice in comparison to untreated mice as well as of M1- and M2-conditioned tumours with regard to age (chapter IV, Impact of macrophages on tumour growth characteristics in a murine ocular tumour model). 3. Further, we intended to study the macrophage-dependent origin and composition of intratumoural vessels in transgenic flt-1/eGFP BAC mice (chapter V, Origin of intratumoural vascular structures in a murine ocular tumour model). 4. In permanent search of a mouse model of spontaneous uveal melanoma, we evaluated ocular findings of elderly Hgf-Cdk4(R24C) mice (chapter VI, Overexpression of hepatocyte growth factor and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma). 23 II Materials and Methods Animals Breeding and housing of C57Bl/6, CX3CR1+/GFP, flt-1/eGFP BAC and Hgf-Cdk4R24C mice mice was carried out and supervised according to the Association of Research in Vision and Ophthalmology statement for the Use of Animals in Ophthalmic and Vision Research. All experiments were approved by the Landesamt für Natur, Umwelt und. Verbraucherschutz NRW (Recklinghausen, Germany). In total, 143 mice of four different strains were used for the studies (Table 1). Of these, six C57Bl/6 mice and 58 CX3CR1+/GFP mice were examined at the age of 8-12 weeks (young), whereas three C57Bl/6 mice, 43 CX3CR1+/GFP mice and 15 flt1/eGFP BAC mice examined at the age of 8-12 months (old). Four C57Bl/6 mice and two CX3CR1+/GFP mice were kept as negative controls. The eyes of 12 Hgf-Cdk4R24C mice aged 11 months were examined. In CX3CR1+/GFP mice which are based on a C57Bl/6 strain, the transmembrane-receptor CX3CR1 for CX3C chemokine fractalkine had been replaced by green fluorescent protein (GFP) reporter gene. Hence, these mice exhibit an in vivo labelling of peripheral blood monocytes, macrophages, subsets of NK and dendritic cells, as well as of retinal microglia by fluorescence at 488nm (Jung, Aliberti et al. 2000). This innate fluorescent signal of monocytic cells may allow their direct visualisation by immunofluorescence microscopy. Homozygous male CX3CR1GFP/GFP mice for breeding were kindly provided by Prof. C. Kurts, Institute of Experimental Immunology, University of Bonn, Bonn, Germany. Female C57Bl/6 mice were crossbred with homozygous male CX3CR1GFP/GFP mice generating a heterozygous CX3CR1+/GFP mice generation. 24 II Materials and Methods Flt-1/eGFP BAC mice exhibit a persistent eGFP expression in small vessels which is driven by the flt-1 promoter (VEGF receptor 1, VEGFR-1) (Herz, Heinemann et al. 2012). Thus, all ocular vessels originating from the host (mouse) exhibit a constant eGFP expression. In Hgf-Cdk4R24C mice overexpression of hepatocyte growth factor (HGF) promotes cutaneous melanogenesis by activating RAS signal transduction pathways via its receptor c-MET (Landsberg, Gaffal et al. 2010). Additionally, the oncogenic germline mutation in the cyclindependent kinase 4 (CDK4 R24C) further contributes to melanoma development in HgfCdk4R24C mice because it functionally inactivates the tumour suppressor p16⁄INK4a, which is of critical importance for oncogene-induced senescence (Landsberg, Kohlmeyer et al. 2012). Table 1: Total animal numbers (all experiments), *=negative controls C57Bl/6 CX3CR1+/GFP flt-1/eGFP BAC Hgf-Cdk4R24C Young (8-12 weeks) Old (8-12 months) 6 (+4*) 58 (+2*) - 3 43 15 12 HCmel12 Melanoma Cell Line HCmel12 melanoma cells were generously provided by Prof. T. Tüting (Laboratory of Experimental Dermatology, University of Bonn, Bonn, Germany). This murine cutaneous melanoma cell line had been derived from an autochthonous cutaneous melanoma of an HgfCdk4 R24C mouse which spontaneously develops a spectrum of primary cutaneous melanomas (Landsberg, Gaffal et al. 2010). These cells exhibit particular characteristics that may qualify them as a cell line for a potent model of metastasising intraocular melanoma. HCmel12 II Materials and Methods 25 melanoma cells were obtained at cell culture passage seven. Cell lines at such an early passage number still exhibit “original” growth characteristics because mutations which may change their phenotype might have not established, yet. HCmel12 melanoma cells were cultured in RPMI 1640 + Hepes medium supplemented with 10% FCS, 1% Penicillin/Streptomycin, 1% NEAA, 1% Sodium-Pyruvate and 0.75% β-Mercaptoethanol. Cells were incubated at 37°C in a humidified atmosphere containing 5% CO2 and grown to approx. 70% confluence. HCmel12 melanoma cells were cryo-conserved at cell culture passage 8. After thawing, cells were cultured for another passage, trypsinised and washed in PBS buffer before aliquots suspended in PBS buffer were injected into the eye. Murine macrophage cell line A bone marrow derived and immortalised macrophage cell line from wild type C57Bl/6 mice was generated (Hornung, Bauernfeind et al. 2008) and kindly provided by Prof. Eicke Latz (Institute of Innate Immunity, University of Bonn, Bonn, Germany). The macrophage cell line was cultured in DMEM/F12 medium at 37°C in a humidified atmosphere containing 5% CO2, grown to max. 70% confluence and passaged every second to third day as described above. Cells were cryo-conserved at a density of approx. 1000 cells per µl. Macrophage polarisation and proliferation in vitro To investigate the impact of polarised macrophages on intraocular melanoma, HCmel12 melanoma cells were exposed in vitro to the supernatant of murine, polarised macrophages prior to intravitreal injection. Murine macrophages were cultured in medium without fetal calf serum (FCS) for two hours prior to in vitro polarisation to avoid any impact of cytokines or chemokines within the FCS. Macrophages were treated with specific cytokines and II Materials and Methods 26 lipopolysaccharide (LPS) to obtain different polarised phenotypes (according to a modified protocol by (Mosser and Zhang 2008)). IFNγ (200ng/ml) for M1-macrophages and Interleukin 4 (IL-4) (20ng/ml) for M2 macrophages were added to the serum-free medium in order to polarise macrophages. After 10 hours of incubation, LPS (100ng/ml) was supplemented for each macrophage type for a total incubation time of 24 hours. Polarisation status of unfixed macrophages was verified functionally by using a multiple cytokine ELISA covering the inflammatory cytokines IL-6, IL-10. IL-12, IL-23, TNFα and IFNγ (MultiAnalyte ELISArrayTM Kit, Mouse Mix-N-Match, Qiagen, Venlo, Netherlands). M1-specific expression of these cytokines comprises levels of IL-6high, IL-10low, IL-12high, IL-23high, TNFαhigh and IFNγhigh (M2-specific expression levels vice versa) (Mantovani, Sica et al. 2004). Phenotypical macrophage polarisation status of 4% PFA-fixed cells was examined by immunocytology with a double staining kit (MultiVision Polymer Detection System, Thermo Scientific, Waltham, MA, USA) using a F4/80- (for all macrophages) and CD163- (for M2 macrophages only; CD163, M-96, Santa Cruz Biotechnology, Santa Cruz, CA, USA) primary antibody. To examine the impact of a M1- or M2-conditioned tumour microenvironment, HCmel12 melanoma cells were treated with serum-free medium for two hours and afterwards exposed to the supernatant of M1- or M2-polarised macrophages for 20 hours, respectively (Figure 6). By this means, HCmel12 melanoma cells were incubated in a M1- or M2dominated microenvironment. These M1- or M2-conditioned HCmel12 melanoma cells were then injected intravitreally as described below. Cell proliferation rates of unpolarised and of M1- or M2-conditioned HCmel12 melanoma cells were compared in vitro using a BrdU proliferation assay (BrdU Cell Proliferation Assay, Milipore, Billerica, MA; USA). II Materials and Methods 27 Figure 6. Experimental design and workflow of in vitro polarisation of macrophages, its verification, conditioning of HCmel12 melanoma cells, and examination of the impact of polarised macrophages on melanoma growth. Macrophage depletion To study the impact of macrophages in intraocular melanoma in CX3CR1+/GFP and flt-1/eGFP BAC mice, macrophage-depleted mice were studied in comparison to untreated mice. Liposome-encapsulated dichloromethylene diphosphonate (Cl2MDP; Clodronate, ordered through clodronateliposomes.org) was used to systemically deplete blood monocytes and macrophages at a dosage of 0.1mg/g body weight (van Rooijen and van Kesteren-Hendrikx 2003). Mice received repeated intraperitoneal injections of Clodronate liposomes on days -6, 3, 0 (day of intravitreal tumour cell injection) and day 3. To evaluate macrophage depletion, liver sections of Clodronate-treated and untreated mice were stained immunohistochemically with a GFP-antibody (GFP Rabbit IgG, Life Technologies, Carlsbad, CA, USA; dilution 1:100) or F4/80-antibody (F4/80 Antibody, Cl:A3-1, AbD Serotec, Oxford, U.K.; dilution 1:100) as described below (Histology). II Materials and Methods 28 Intraocular tumour cell injection Experimental groups which received intraocular tumour cell injections consisted of 12 mice in the animal model experiment (chapter III), 95 mice in the depletion and polarisation experiment (chapter IV), and 15 mice in the vascularisation experiment (chapter V). General anaesthesia comprised Isofluran induction followed by intraperitoneal Ketamin/Xylazin injection. On day 0. in general anaesthesia, 1x105 HCmel12 melanoma cells suspended in 1µl PBS were injected intravitreally into the right eye of each mouse. An automatic microsyringe injector (UltraMicroPump III, World Precision Instruments, Sarasota, USA) with a 10µl microinjection syringe (701 ASRN, Hamilton, Reno, USA) equipped with a 34-gauge needle (RN Needle, Hamilton, Reno, USA) was used for injection. The needle was inserted approx. 1 mm posterior to the limbus through the sclera into the vitreous. Negative controls in the animal model experiment (chapter III) received 1µl of PBS, a sole needle stitch with no injecting agent (n=2) or no treatment at all (n=1). Intraocular injections were performed very carefully to avoid punctuation of the lens. Negative controls may show whether a noteworthy influx of immune cells was caused by the injection process of PBS or the stitch alone. Tumour bearing eyes of all mice were enucleated in general anaesthesia on day 9 and 12 CX3CR1+/GFP and C57Bl/6 mice were sacrificed on day 42 in the animal model experiment. Then tissue samples from lungs, liver and spleen, as well as the contralateral eye and mandibular lymph nodes were collected for further examination to verify metastatic routes and sites (chapter III). In all other experiments, mice were euthanised on day 9 with immediate enucleation of the tumour bearing eye (chapter IV+V). All but 20 eyes of CX3CR1+/GFP, all C57Bl/6 mice and 12 eyes of flt-1/eGFP BAC mice were processed for histology. 20 eyes of CX3CR1+/GFP mice were investigated by flow cytometry analysis and three eyes of flt-1/eGFP BAC mice were investigated by ex vivo imaging. II Materials and Methods 29 Histology and immunohistochemistry Specimens for histologic examination were submitted to 4% paraformaldehyde for 24 hours, dehydrated with alcohol in ascending order and cleared with xylene. After embedding in paraffin, 5 µm thick serial sections of the eye and five sections through the center of the other tissues were cut using a manual rotary microtome (Thermo Scientific Shandon Finesse 325 Microtome). They were routinely processed for light microscopic examination and stained with hematoxylin and eosin (H&E) or periodic acid-Schiff (PAS) without hematoxylin. H&E stains were investigated at 40-100x magnification to evaluate histologic characteristics of the tumour as well as strict intraocular tumour localisation versus tumour growth with extraocular extension. H&E stained sections of liver, lung, spleen, sentinel lymph nodes and the contralateral eye were scanned for (micro-) metastases at 100-400x magnification. PAS stained sections without hematoxylin were investigated for vasculogenic mimicry and extravascular matrix patterns using a polarised filter at 100-400x magnification. For immunohistochemical melanoma detection we used a melanoma marker (Anti-Melanoma antibody, HMB45+DT101+BC199, Abcam, Cambridge, UK; dilution 1:50). After deparaffinisation and rehydration, heat-mediated antigen retrieval was performed with 0.01M citrate buffer for 15 minutes. Washing steps were performed with TBS buffer (Tris buffer [TBS: 50mM, pH 7.6]). After blocking with hydrogen peroxide the sections were incubated with the primary antibody at 4°C overnight. The antigen-antibody-binding reaction was visualised with an antigen detection system using horseradish peroxidase (HRP) and 3-amino9-ethylcarbazole (AEC) as the chromogen (MaxHomo Mouse on Mouse Polymer HRP Detection Kit, Max Vision Biosciences Inc., Washington, USA). Macrophage labelling in the tumour bearing eye was achieved using a F4/80 primary antibody (Rat Anti-Mouse F4/80 Antibody, clone Cl:A3-1, AbD Serotec, Oxford, UK; dilution 1:100) II Materials and Methods 30 and additionally with a GFP-antibody in CX3CR1+/GFP mice (GFP Rabbit IgG Polyclonal Antibody Fraction, Life Technologies, Carlsbad, CA, USA; dilution 1:100). Tumour vasculature and mean vascular density (MVD) were determined in sections stained with a CD31-antibody (CD31, SZ31, Dianova, Hamburg, Germany, dilution 1:10). At 200x magnification, vessels were counted within a 0.25mm2 grid in 1-3 areas according to Foss et al. (Foss, Alexander et al. 1996). Staining of extracellular matrix (ECM) was additionally performed with a collagen IV antibody (Collagen IV Antibody, Novus Biologicals, Littleton, CO, USA dilution 1:100) and a laminin antibody (Laminin Antibody, Novus Biologicals, Littleton, CO, USA, dilution 1:100). In flt-1/eGFP BAC mice all smaller host vessels were additionally labelled by a GFP-antibody. Dual labelling in immunofluorescence microscopy of CD31 and GFP allows for determination of the origin of endothelial-lined vessels. Dual labeling of CD31 + Ki67 (Anti-Ki67, SP6, Abcam, Cambridge, UK, dilution 1:50) is supposed to reveal active angiogenesis in immunofluorescence microscopy. Tumour microvasculature was additionally labelled by Vascular Endothelial Cadherin (VECadherin (Phospho Tyr731) Antibody, Assaybiotech, Sunnywale, CA, USA; dilution 1:200) with antigen retrieval by pepsin (3 minutes), overnight incubation of the primary antibody at 4°C, blocking with hydrogen peroxide, a secondary antibody incubation for one hour at room temperature (Polyclonal Swine Anti-Rabbit Immunoglobulins, DakoCytomation, Glostrup, Denkmark; dilution 1:200), and streptavidin one hour at room temperature (Streptavidinbiotinylated horseradish peroxidase complex, GE Healthcare, Amersham, Buckinghamshire, U.K.; dilution 1:100). To distinguish the genuine ocular pigment from the immunoreaction we visualised the latter with the red chromogen AEC (15 minutes at room temperature) instead of a commonly used brown chromogen. II Materials and Methods 31 Flow cytometry analysis Ocular tumours of 20 CX3CR1+/GFP mice (young n=10 and old n=10) were dissected microsurgically, pooled, respectively, and digested in 0.5mg/ml collagenase and 100µg/mg DNAase I in RPMI medium for 40 minutes. The washed cells were filtered through a 100µm nylon mesh to obtain a single cell suspension. To evaluate macrophage infiltration within intraocular tumours, surface staining was performed with monoclonal antibodies against characteristic leucocyte and macrophage receptors which were conjugated to different fluorochromes: CD45 (Biolegend Inc., San Diego, CA, USA), CD11b (Biolegend), F4/80 (Biolegend), CD80 (Biolegend), CD163 (M-96, Santa Cruz Biotechnology, Santa Cruz, CA, USA + conjugation kit, Abcam, Cambridge, UK), and Gr-1 (Biolegend). Controls were run as single stains and unstained probes. Measurements were carried out on a flow cytometer (LSRFortessa, BD, Franklin Lakes, NJ, USA) and results were analysed by using FlowJo software (Tristar Inc. San Carlos, CA, USA). Ex vivo imaging In three flt-1/eGFP BAC mice, tumour-bearing and contralateral control eyes were enucleated immediately after euthanasia. Eyes were kept in warm buffer (PBS) and shortly after investigated using an Axio Zoom microscope (Carl Zeiss Microscopy GmbH, Jena, Germany). Specimens were examined for GFP-positive vessels in areas of extraocular tumour extension at x25 power magnification. Afterwards, the anterior parts of the eye were removed and intraocular tumour parts in the posterior compartment of the eye were examined for GFPpositive vessels. II Materials and Methods 32 Statistical analysis Statistical analysis was performed with IBM SPSS Statistics 22.0 (IBM Corp, Armonk, NY, USA). Evaluation of histological sections was performed by light microscopy by two independent investigators. Number of infiltrating macrophages as well as the presence and amount of collagen IV- and laminin-positive structures were analysed semi-quantitatively from 0 to +++ by two independent examiners and a p-value of (MCH, MMK). Tumour size was measured in mm2 at 40x magnification using ImageJ 1.45s (W Rasband, NIH, USA). Tumour size and MVD of different study groups were evaluated by an analysis of variance and a Post Hoc Test. Chi2 test and Pearson's correlation coefficient were used to determine the relationship between different study groups and tumour growth characteristics. Student’s t-test and ROC analysis was applied for interpretation of cytokine levels of in vitro polarised macrophages. A p≤0.05 was considered as statistically significant and significance level was indicated by using * for p ≤ 0.05. Histology and immunohistochemistry of Hgf-Cdk4R24C mice eyes 12 eyes from 11 month old Hgf-Cdk4R24C mice were investigated and were compared with three eyes from 10 months old C57Bl/6 mice. Step sections were stained with H&E and PAS, and sections in-between were investigated by immunohistochemical staining. Primary antibodies against Ki67 (proliferation marker), MART1 (melanocyte/melanoma marker), CKpan (epithelial cell differentiation) and F4/80 (macrophages) were used. Prior to immunohistochemistry, sections of these highly pigmented mice were bleached for better visualisation of the red chromogen AEC. 33 III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumour biology of intraocular melanoma Marta M. Kilian1, Karin U. Loeffler1, Christiane Pfarrer2, Frank G. Holz1, Christian Kurts3, Martina C. Herwig1 1 Department of Ophthalmology, University of Bonn, Bonn, Germany 2 Department of Anatomy, University of Veterinary Medicine Hannover, Hannover, Germany 3 Institute of Experimental Immunology, University of Bonn, Bonn, Germany Current Eye Research Informa Healthcare USA, Inc. ISSN: 0271-3683 print / 1460-2202 online DOI: 10.3109/02713683.2015.1004721 III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 34 Abstract Purpose: To establish a mouse model with histologic characteristics of uveal melanoma for investigation of intraocular tumour biology of melanoma. Methods: After injection of 1x105 of HCmel12 melanoma cells, a cutaneous melanoma cell line, into the vitreous of CX3CR1+/GFP or C57Bl/6 mice (n=12), tumour growth patterns, clinicopathological features, angiogenesis and metastatic behaviour were analysed by histology (H&E, PAS without hematoxylin) and immunohistochemistry (HMB45/MART-1Ab, F4/80-Ab, GFP-Ab, VE-cadherin-Ab). Results: HCmel12 cells formed intraocularly growing tumour masses which showed histologic features of intraocular melanoma such as angiotropism, intratumoural endotheliallined vasculature, vasculogenic mimicry including prognostic significant extravascular matrix patterns, and invasion by inflammatory cells, in particular macrophages. There was no difference in tumour growth characteristics between CX3CR1+/GFP and C57Bl/6 mice. Five out of ten mice proceeded to extrascleral tumour growth, and three of these developed metastases. Conclusions: Intraocularly injected HCmel12 cells developed tumour masses with histologic characteristics of aggressive melanoma similar to human uveal melanoma. Since hematogenous dissemination to the liver was not observed, intravitreally injected HCmel12 cells do not qualify as a model for metastasizing intraocular melanoma. However, since the eye represents a semi-closed compartment with access to constant blood supply, these intraocular tumours represent a model for studies of isolated parameters in general tumour biology of intraocular melanoma. III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 35 Introduction Uveal melanoma is the most common primary intraocular tumour in the adult Caucasian population. While local treatment of the primary tumour is very promising, the outcome worsens dramatically when metastatic disease appears (Kujala, Makitie et al. 2003). Uveal melanoma metastasises predominantly hematogenously to the liver and rarely to the lungs or other organs. Metastases develop in up to 40% of patients within 10 years of initial diagnosis (Singh, Shields et al. 2001). In order to sustain solid growth the tumour has to ensure sufficient blood supply. By releasing certain factors or chemokines and thereby influencing the microenvironment the tumour utilises angiogenesis and other sources like co-option of pre-existing vessels or formation of mosaic vessels (Hanahan and Weinberg 2011). Many growth characteristics concerning the tumour’s blood supply are related to metastasis and thus to prognosis. Microvascular density (MVD) has been found to be of prognostic value for uveal melanoma (Foss, Alexander et al. 1996; Makitie, Summanen et al. 1999). Intravascular ingrowth of uveal melanoma cells and extraocular growth are also contributing factors to metastasis formation (Ly, Odish et al. 2010). Metastasis is further associated with the primary tumour exhibiting certain patterned meshworks of fluid conducting non-endothelial channels described as extravascular matrix patterns or vasculogenic mimicry (VM) (Folberg, Mehaffey et al. 1997). VM is rich of PASpositive extracellular matrix (ECM) with high expression levels of ECM related molecules (e.g. matrix-metalloproteinases, VE-cadherin, VEGF-A) (Seftor, Hess et al. 2012). Apparently, tumour microcirculation consists of a combination of angiogenic vessels, mosaic vessels as well as ECM-rich VM meshworks (Hendrix, Seftor et al. 2003). Angiogenesis and the tumour microenvironment are further influenced by factors released by tumour infiltrating III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 36 leucocytes (Sica and Mantovani 2012), in particular lymphocytes and macrophages. Tumour growth favoring phenotypes of macrophages are preferably recruited into the tumour area (Bronkhorst and Jager 2012). Increased numbers of tumour associated macrophages (TAM) correlate - among other tumour characteristics - with high MVD and with patient’s death from metastatic disease (Toivonen, Makitie et al. 2004). In turn, the microenvironment has a major impact on primary tumour cells with regard to tumour progression and metastasis. Although each tumour may be genetically predetermined to enter lymphatic and/or blood vessels for dissemination (Nathanson 2003), successful metastases development is also affected by molecular interactions between circulating tumour cells and the pre-metastatic site (Mendoza and Khanna 2009). For human uveal melanoma certain receptors and factors accounting for liver-specific metastases have been determined, including c-met, a receptor for hepatocyte growth factor (HGF) /scatter factor (Bakalian, Marshall et al. 2008). Furthermore, several genetic anomalies (e.g. monosomy 3, class 2 gene expression profile, GNAQ, GNA11, Bap1) of uveal melanoma influencing tumour progression and metastasis have been identified (Sisley, Rennie et al. 1990; Onken, Worley et al. 2004; Ehlers, Worley et al. 2008). Nonetheless, characteristic mutations differ between uveal and cutaneous melanoma and even among tumours themselves (Mehnert and Kluger 2012). However, uveal and cutaneous melanomas do share fundamental characteristics in terms of tumour growth, inflammatory cell infiltration, microenvironmental stimuli and vascularisation. Considering the aforementioned characteristics of uveal melanoma like hematogenous spread, characteristic microcirculation, the inflammatory microenvironment and genetic attributes, a meaningful mouse model should offer similar characteristics. The aim of this study was to evaluate histopathologic growth and metastatic characteristics of intravitreally injected HCmel12 melanoma cells. Evaluation emphasised the applicability of III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 37 HCmel12 melanoma cells as a mouse model for intraocular melanoma in terms of tumour biology and metastasis. Materials and Methods Animals Breeding and housing of CX3CR1+/GFP mice and C57Bl/6 mice was carried out and supervised according to the Association of Research in Vision and Ophthalmology statement for the Use of Animals in Ophthalmic and Vision Research. Six CX3CR1+/GFP mice and six C57Bl/6 mice were examined at the age of 8-12 weeks, whereas two CX3CR1+/GFP mice and four C57Bl/6 mice of the same age were kept as negative controls (Table 1). In CX3CR1+/GFP mice which are based on a C57Bl/6 strain, the transmembrane-receptor CX3CR1 for CX3C chemokine fractalkine had been replaced by green fluorescent protein (GFP) reporter gene. Hence, these mice exhibit an in vivo labelling of peripheral blood monocytes, macrophages, subsets of NK and dendritic cells, as well as of retinal microglia by fluorescence at 488nm (Jung, Aliberti et al. 2000). This innate fluorescent signal of monocytic cells may allow their direct visualization by immunofluorescence microscopy. Homozygous male CX3CR1GFP/GFP mice for breeding were kindly provided by Prof. C. Kurts, Institute of Experimental Immunology, University of Bonn, Bonn, Germany. Female C57Bl/6 mice were crossbred with homozygous male CX3CR1GFP/GFP mice generating a heterozygous CX3CR1+/GFP mice generation. C57Bl/6 mice were also used as controls since the original III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 38 B16LS9 mouse model for metastasizing intraocular melanoma was established in this mouse strain (Diaz, Rusciano et al. 1999). HCmel12 melanoma cell line HCmel12 melanoma cells were generously provided by Prof. T. Tüting, Laboratory of Experimental Dermatology, University of Bonn, Bonn, Germany. This murine cutaneous melanoma cell line had been derived from an autochthonous cutaneous melanoma of an HgfCdk4 R24C mouse which spontaneously develops a spectrum of primary cutaneous melanomas (Landsberg, Gaffal et al. 2010). HCmel12 cells of an early cell culture passage exhibit particular characteristics like increased c-met expression, angiotropism and spontaneous metastatic properties that may qualify them as a cell line for a potent model of metastasizing intraocular melanoma. Furthermore, HCmel12 cells’ growth characteristics resemble those of human uveal melanoma regarding angiotropism and VM, as well as the attraction of a high number of macrophages (unpublished data). The HCmel12 melanoma cell line was cultured in RPMI 1640 + Hepes medium supplemented with 10% FCS, 1% Penicillin/Streptomycin, 1% NEAA, 1% Sodium-Pyruvate and 0.75% βMercaptoethanol. Cells were incubated at 37°C in a humidified atmosphere containing 5% CO2 and grown to approx. 70% confluence. HCmel12 melanoma cells were cryo-conserved at cell culture passage 8. After thawing, cells were cultured for another passage, trypsinised and washed in PBS buffer before aliquots suspended in PBS buffer were injected into the eye. III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 39 Clinical procedures The experimental group consisted of 12 mice; six mice were used as negative controls. General anaesthesia comprised Isofluran induction followed by intraperitoneal Ketamin/Xylazin injection. On day 0. in general anaesthesia, 1x105 HCmel12 melanoma cells suspended in 1µl PBS were injected intravitreally into the right eye of both mouse strains, CX3CR1+/GFP mice and C57Bl/6 mice. An automatic microsyringe injector (UltraMicroPump III, World Precision Instruments, Sarasota, USA) with a 10µl microinjection syringe (701 ASRN, Hamilton, Reno, USA) equipped with a 34-gauge needle (RN Needle, Hamilton, Reno, USA) was used for injection. The needle was inserted approx. 1 mm posteriour to the limbus through the sclera into the vitreous. Negative controls received 1µl of PBS (n=3, 1 CX3CR1+/GFP mouse, 2 C57Bl/6 mice), a sole needle stitch with no injecting agent (n=2, 1 CX3CR1+/GFP and 1 C57Bl/6 mouse) or no treatment at all (n=1, C57Bl/6 mouse). Negative controls may show whether a noteworthy influx of immune cells was caused by the injection process of PBS or the stitch alone. Tumour bearing eyes of CX3CR1+/GFP mice and C57Bl/6 mice were enucleated in general anaesthesia on day 9 and the mice were sacrificed on day 42. Then tissue samples from lungs, liver and spleen, as well as the contralateral eye and mandibular lymph nodes were collected for further examination to verify metastatic routes and sites. Histology Specimens were submitted to 4% paraformaldehyde for 24 hours, dehydrated with alcohol in ascending order and cleared with xylene. After embedding in paraffin, 5 µm thick serial sections of the eye and five sections through the center of the other tissues were cut using a manual rotary microtome (Thermo Scientific Shandon Finesse 325 Microtome). They were III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 40 routinely processed for light microscopic examination and stained with hematoxylin and eosin (H&E) or periodic acid-Schiff (PAS) without hematoxylin. H&E stains were investigated at 40-100x magnification to evaluate histologic characteristics of the tumour as well as strict intraocular tumour localization versus tumour growth with extraocular extension. Tumour size was measured at 40x magnification using ImageJ 1.45s. H&E stained sections of liver, lung, spleen, sentinel lymph nodes and the contralateral eye were scanned for metastases at 100-400x magnification. PAS stained sections without hematoxylin were investigated for VM and extravascular matrix patterns using a polarised filter at 100-400x magnification. For immunohistochemical melanoma detection we used a commercially available melanoma marker (Anti-Melanoma antibody, HMB45+DT101+BC199, Abcam, Cambridge, UK; dilution 1:50). After deparaffinization and rehydration, heat-mediated antigen retrieval was performed with 0.01M citrate buffer for 15 minutes. Washing steps were performed with TBS buffer (Tris buffer [TBS: 50mM, pH 7.6]). After blocking with hydrogen peroxide the sections were incubated with the primary antibody overnight at 4°C. The antigen-antibodybinding reaction was visualised with an antigen detection system using horseradish peroxidase (HRP) and 3-amino-9-ethylcarbazole (AEC) as the chromogen (MaxHomo Mouse on Mouse Polymer HRP Detection Kit, Max Vision Biosciences Inc., Washington, USA). Macrophage labelling was achieved using a F4/80 primary antibody (Rat Anti-Mouse F4/80 Antibody, clone Cl:A3-1, AbD Serotec, Oxford, UK; dilution 1:100) in both mouse strains and additionally with a GFP-antibody in CX3CR1+/GFP mice (GFP Rabbit IgG Polyclonal Antibody Fraction, Life Technologies, Carlsbad, CA, USA; dilution 1:100); tumour microvasculature was demonstrated by Vascular Endothelial Cadherin (VE-Cadherin (Phospho Tyr731) Antibody, Assaybiotech, Sunnywale, CA, USA; dilution 1:200) with antigen retrieval by pepsin (3 minutes), overnight incubation of the primary antibody at 4°C, III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 41 blocking with hydrogen peroxide, a secondary antibody incubation for one hour at room temperature (Polyclonal Swine Anti-Rabbit Immunoglobulins, DakoCytomation, Glostrup, Denkmark; dilution 1:200), streptavidin one hour at room temperature (Streptavidinbiotinylated horseradish peroxidase complex, GE Healthcare, Amersham, Buckinghamshire, U.K.; dilution 1:100) and visualization by AEC (15 minutes at room temperature). The analysis was performed by light microscopic evaluation by two independent investigators. Results We intended to evaluate intraocular growth and metastatic characteristics of HCmel12 cells which represent a new murine cutaneous cell line originating from an autochthonous melanoma in mice with a C57Bl/6 background. Therefore, six CX3CR1+/GFP mice and six C57Bl/6 mice received an intravitreal tumour cell injection at the age of 8-12 weeks. Enucleated eyes and potential organs for metastatic tumour localization were processed for histological examination. Histology of intraocular HCmel12 melanoma Of twelve mice two mice (1 CX3CR1+/GFP and 1 C57Bl/6 mouse) deceased early in the experiment and were excluded from statistics. The eyes of the remaining ten mice which had been injected with HCmel12 cells comprised five specimens with intraocular tumour growth III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 42 strictly confined to the eye (Fig. 1a), while the other five specimens exhibited additionally extraocular extension (Fig. 1b) (Table 1). Table 1: Experimental groups of animals Mouse 1 2 3 4 5 6 7 8 9 10 11 12 Genotype Tumour size[mm2] Extraocular Metastases C57 Bl/6 C57 Bl/6 C57 Bl/6 C57 Bl/6 C57 Bl/6 C57 Bl/6† CX3CR1+/GFP CX3CR1+/GFP CX3CR1+/GFP CX3CR1+/GFP CX3CR1+/GFP CX3CR1+/GFP† 0.1805 1.4929 0.3169 0.2049 0.142 No Yes No No Yes No Yes No No Yes 0.1353 0.0608 0.1179 0.6604 0.2727 Yes No Yes Yes No No No No Yes No †=deceased All mice received an intravitreal injection of 1x105 HCmel12 melanoma cells. Tumour bearing eyes were enucleated on day 9 and mice were euthanised on day 42. One mouse (C57Bl/6) deceased shortly after intravitreal injection allowing for histological evaluation of intraocular conditions immediately after tumour cell injection. In this mouse HCmel12 cells were distributed uniformly within the posteriour compartment of the eye. However, a few cells remained along the injection channel and outside intraocular structures (Fig. 1c). In ten mice with tumour bearing eyes H&E sections revealed tumour growth mostly around the optic nerve with weak pigmentation and occasional necrotic areas. Also, intraocular tumours exhibited invasive properties with destruction of surrounding ocular structures such as the retina and the choroid (n=7) (Fig. 1d+f). They eventually (n=5) proceeded to extraocular growth, most likely along the injection channel by following the weakest point along the sclera. III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 43 Figure 1: Ocular HCmel12 tumours, H&E (a) Intraocular tumour growth (arrow), 40x magnification, CX3CR1+/GFP mouse; (b) Intraocular tumour growth (arrow) with extraocular extension (arrowhead), 40x, C57Bl/6 mouse; (c) Intravitreally injected HCmel12 cells distribute in the vitreous and along the injecting channel on day of injection, 100x, C57Bl/6 mouse; (d) Intraocular tumour with destructive growth characteristics and evenly distributed microvasculature, 200x, C57Bl/6 mouse; (e) Intraocular tumour exhibiting angiotropism (arrow), blood channels and lakes (arrowhead), 400x, CX3CR1+/GFP mouse; (f) Intraocular tumour penetrates the retina and invades into the choroid, 200x, C57Bl/6 mouse. Tumour cells grew towards and around retinal blood vessels (n=8) indicating angiotropism (Fig. 1e). Solid tumours exhibited channels and lakes filled with blood (n=8) (Fig. 1e) which were enclosed by melanoma cells. Intratumoural endothelial lined vasculature was present (Fig. 1d) as well as co-option of retinal vessels (Fig. 2a). Further, H&E stains showed a scattered tumour infiltration by inflammatory cells. By PAS without hematoxylin, PASpositive ECM-rich channels enclosed by melanoma cells were detected (Fig. 2b). Extravascular matrix patterns were identified and ranged from straights (n=5), parallel vessels III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 44 (n=4), in some cases with interconnecting branches (n=2) to arcs (n=7), arcs with branches (n=7) and loops (n=4) (Fig. 2c). However, interconnecting loops creating networks were not observed. Figure 2: Ocular HCmel12 tumours, sections stained with PAS without hematoxylin viewed with a polarising microscope showing ECM-rich structures, CX3CR1+/GFP mouse (a) Co-option of retinal vessels (arrow), 200x magnification; (b+c) ECM-rich meshwork considered as vasculogenic mimicry with patterns like arcs with branches, parallels with interconnections or loops (arrows) 100-200x. Histologic characteristics of intraocular HCmel12 tumours showed no evident differences between CX3CR1+/GFP mice and C57Bl/6 mice. Also, measured intraocular tumour size did not differ statistically between CX3CR1+/GFP mice and C57Bl/6 mice (p=0.46) (Tab. 1). Tumour size was statistically not correlated with extraocular extension (p=0.078). In H&E sections, control mice injected with PBS or nothing, did not show any immunological reactions to the injection process. The removed contralateral eye without tumour was unremarkable in all investigated mice. III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 45 Immunohistochemistry of intraocular HCmel12 melanoma In CX3CR1+/GFP mice the innate GFP signal was too weak for allowing direct evaluation by immunofluorescence microscopy. However, labelling of macrophages using a F4/80- and a GFP-antibody revealed positively stained cells within the retina, most likely representing retinal microglia. Labelled cells were also found evenly distributed throughout intra- and extraocular parts of the tumour; in intraocular parts especially near the retina or the choroid (Fig. 3a). Some appeared as large cells with vacuoles or granula, others as cells with a small cell body and protruding dendrites (Fig. 3b). The microvasculature was positively stained by the VE-Cadherin antibody throughout the tumours (Fig. 3c) but did not correlate with the characteristic ECM-rich patterns as seen in PAS stains without hematoxylin. Figure 3 Immunohistochemical staining of ocular HCmel12 melanoma and remaining retina (asterisk), CX3CR1+/GFP mouse (a) GFP-antibody labelling of retinal microglia and infiltrating tumour associated macrophages, 100x magnification; (b) High power area indicated in box of Fig. 3a, 200x; (c) VE-cadherin-antibody labelling of microvasculature, 400x III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 46 Metastases of HCmel12 melanoma Distant micro- and macrometastases were diagnosed in H&E sections (Fig. 4a) as well as by an immunohistochemical melanoma marker (HMB45/MART-1) in lungs (Fig. 4b) in three out of ten examined mice. All of these mice exhibited previous extraocular tumour growth. No mice with tumours confined strictly to the eye showed any metastatic colonization in distant tissue. Statistically, the incidence of metastases was not correlated with tumour size (p=0.16) nor with the presence of extravascular matrix patterns (p=0.33). However, because of the small number of animals with metastases, this data must be interpreted carefully. Metastatic involvement of the lymphnodes could not be unequivocally detected due to the high cellularity of the lymph nodes and the unspecific immunohistochemical signal in this tissue. However, the immunohistochemical labelling in the lungs was confined to the metastases and interpreted as reliable. Metastases into the liver and spleen could not be detected. Figure 4: Pulmonary metastasis of HCmel12 melanoma (a) Lung metastasis, H&E, 100x magnification, C57Bl/6 mouse; (b) Immunohistochemical staining for HMB45/MART-1, 200x, CX3CR1+/GFP mouse III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 47 Discussion In order to study the pathogenesis of human uveal melanoma several mouse models have been developed including transgenic mice, intraocular inoculation with cutaneous melanoma cells into wildtype mice, injection of human uveal melanoma cells into SCID mice or induction of uveal proliferations by chemicals, radiation or viruses (Dithmar, Albert et al. 2000). Since spontaneous primary uveal melanoma has not been described in wildtype mice yet, existing mouse models are still subject to restrictions. Pigmented intraocular tumours arising in transgenic mice were identified to be either of retinal pigment epithelium origin (Syed, Windle et al. 1998; Albert, Kumar et al. 2004) or were small uveal tumours which failed to metastasise to the liver (Kramer, Powell et al. 1998; Tolleson, Doss et al. 2005; Latendresse, Muskhelishvili et al. 2007). Injection of human uveal melanoma cells into immunodeficient mice also resulted in lacking metastatic disease (Mueller, Maniotis et al. 2002) and does not allow the study of influences on tumour and host immunology. The best reasonable compromise, and an established mouse model thus far, was created by injecting murine, cutaneous B16LS9 melanoma cells into the vitreous of C57Bl/6 mice, generating an ocular tumour with metastatic dissemination to the lungs and the liver. According to Diaz and coworkers intraocular tumours were restricted to the eye at the time of enucleation, hence metastatic dissemination to the lungs and liver occurred hematogenously since the eye lacks lymphatic vessels (Diaz, Rusciano et al. 1999). Focussing further on hepatic metastases a mouse model with PEDF null mice has been generated (Lattier, Yang et al. 2013). Another group focussed on intraocular tumour growth of intraocularly injected B16 cells to investigate the distribution of extravascular matrix patterns in intraocular melanoma. VM was found along with endothelium-dependant vessels and with connection to the host’s circulation III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 48 (Chen, Zhang et al. 2009). These findings also show that the stroma compartment of the choroid is not mandatory for ECM and VM since the tumour cells were directly injected into the vitreous and not all tumours gained access to the murine choroid (Grossniklaus 1998). In our model, we investigated ocular tumours and mestastatic behaviour of HCmel12 melanoma cells, which represent a new murine cutaneous cell line with certain characteristics that may qualify them as a promising cell line for a metastasizing intraocular melanoma model. HCmel12 melanoma cells succeeded in solid intraocular tumour growth and showed properties of an aggressive melanoma, including metastatic spread to the lungs. There were no differences in tumour growth, size or metastatic characteristics between inoculated C57Bl/6 and CX3CR1+/GFP mice. Tumour size did not have an impact on extraocular extension and metastasis. However, as this statistical data is based on a small number of animals, further experiments are necessary to confirm this finding in a larger cohort. Interestingly - despite angiotropic growth, VM and an increased c-met expression of HCmel12 cells - none of the exclusively intraocular growing HCmel12 tumours succeeded in colonization of distant tissue. On the other hand, tumours with extraocular extension metastasised in 60% of cases preferably to the lungs, probably under participation of the conjunctival lymphatic system. This inability of forming hematogenously dispersed metastases may be due to failed vascular ingrowth of tumour cells or a lack of effective molecular interactions between trafficking primary tumour cells within the blood system and a potential pre-metastatic site like the liver. Thus, HCmel12 melanoma cells qualify very restrictedly as a meaningful mouse model for metastasizing uveal melanoma. However, focusing on intraocular tumour biology, HCmel12 melanoma cells show apparent similarities to human uveal melanoma. Intraocular solid tumours revealed aggressive growth and infiltration by inflammatory cells, particularly macrophages. Further, the tumour exhibited III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 49 angiotropic growth, angiogenesis and induced the formation of ECM-rich, PAS-positive channels considered as VM. Microcirculation of HCmel12 tumours revealed formation of prognostic significant ECM patterns which are known to be correlated with a class 2 gene expression profile of uveal melanoma (Onken, Worley et al. 2004; Onken, Lin et al. 2005). Isolated histopathologic factors like MVD, inflammatory cell infiltration, VM (including extravascular matrix patterns) are no longer crucial for evaluating prognosis in human uveal melanoma as gene expression profiling has been found to be a more meaningful indicator for patient’s prognosis (Onken, Worley et al. 2012). However, these histologic prognostic parameters are reflecting uveal melanoma growth characteristics and allow for their evaluation. Overall, intravitreally injected HCmel12 cells still represent murine melanomas of cutaneous origin. Even though they may share a number of genetic and histopathologic properties with human uveal melanoma, they still differ significantly in specific mutations (Hurst, Harbour et al. 2003). Animal models are rarely capable of imitating an entire human pathologic condition, but are able to reflect certain characteristics. Because the eye is lacking lymphatic drainage but comprises a constant blood supply, intraocular tumours exhibit a microenvironment of a virtual semi-closed system which can be used to study isolated parameters in tumour biology. As HCmel12 melanoma cells are preserved at a low cell culture passage, alterations in genetics and growth characteristics - which may be caused by ongoing cell culture procedures - are less probable. Intraocular HCmel12 growth resembles human intraocular melanoma in many ways. Thus, tumours of intravitreally injected HCmel12 melanoma cells qualify as a model for intraocular melanoma and allow studies on specific questions related to the tumour’s microcirculation, including VM patterns. III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 50 With regard to intraocular melanoma, this study shows that intraocularly injected HCmel12 cells are a potent model for intraocular tumour biology allowing for analysis of tumour angiogenesis, extravascular matrix patterns and microenvironmental influences of inflammatory cells like macrophages. References 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. Kujala, E., T. Makitie, and T. Kivela, Very long-term prognosis of patients with malignant uveal melanoma. Invest Ophthalmol Vis Sci, 2003. 44(11): p. 4651-9. Singh, A.D., C.L. Shields, and J.A. Shields, Prognostic factors in uveal melanoma. Melanoma Res, 2001. 11(3): p. 255-63. Hanahan, D. and R.A. Weinberg, Hallmarks of cancer: the next generation. Cell, 2011. 144(5): p. 646-74. Foss, A.J., et al., Microvessel count predicts survival in uveal melanoma. Cancer Res, 1996. 56(13): p. 2900-3. Makitie, T., et al., Microvascular density in predicting survival of patients with choroidal and ciliary body melanoma. Invest Ophthalmol Vis Sci, 1999. 40(11): p. 2471-80. Ly, L.V., et al., Intravascular presence of tumor cells as prognostic parameter in uveal melanoma: a 35-year survey. Invest Ophthalmol Vis Sci, 2010. 51(2): p. 65865. Folberg, R., et al., The microcirculation of choroidal and ciliary body melanomas. Eye (Lond), 1997. 11 ( Pt 2): p. 227-38. Seftor, R.E., et al., Tumor cell vasculogenic mimicry: from controversy to therapeutic promise. Am J Pathol, 2012. 181(4): p. 1115-25. Hendrix, M.J., et al., Vasculogenic mimicry and tumour-cell plasticity: lessons from melanoma. Nat Rev Cancer, 2003. 3(6): p. 411-21. Sica, A. and A. Mantovani, Macrophage plasticity and polarization: in vivo veritas. J Clin Invest, 2012. 122(3): p. 787-95. Bronkhorst, I.H. and M.J. Jager, Uveal Melanoma: The Inflammatory Microenvironment. J Innate Immun, 2012. Toivonen, P., et al., Microcirculation and tumor-infiltrating macrophages in choroidal and ciliary body melanoma and corresponding metastases. Invest Ophthalmol Vis Sci, 2004. 45(1): p. 1-6. Nathanson, S.D., Insights into the mechanisms of lymph node metastasis. Cancer, 2003. 98(2): p. 413-23. III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 51 Mendoza, M. and C. Khanna, Revisiting the seed and soil in cancer metastasis. Int J Biochem Cell Biol, 2009. 41(7): p. 1452-62. Bakalian, S., et al., Molecular pathways mediating liver metastasis in patients with uveal melanoma. Clin Cancer Res, 2008. 14(4): p. 951-6. Sisley, K., et al., Cytogenetic findings in six posteriour uveal melanomas: involvement of chromosomes 3, 6, and 8. Genes Chromosomes Cancer, 1990. 2(3): p. 205-9. Onken, M.D., et al., Gene expression profiling in uveal melanoma reveals two molecular classes and predicts metastatic death. Cancer Res, 2004. 64(20): p. 7205-9. Ehlers, J.P., et al., Integrative genomic analysis of aneuploidy in uveal melanoma. Clin Cancer Res, 2008. 14(1): p. 115-22. Mehnert, J.M. and H.M. Kluger, Driver mutations in melanoma: lessons learned from bench-to-bedside studies. Curr Oncol Rep, 2012. 14(5): p. 449-57. Jung, S., et al., Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol Cell Biol, 2000. 20(11): p. 4106-14. Diaz, C.E., et al., B16LS9 melanoma cells spread to the liver from the murine ocular posteriour compartment (PC). Curr Eye Res, 1999. 18(2): p. 125-9. Landsberg, J., et al., Autochthonous primary and metastatic melanomas in Hgf-Cdk4 R24C mice evade T-cell-mediated immune surveillance. Pigment Cell Melanoma Res, 2010. 23(5): p. 649-60. Dithmar, S., D.M. Albert, and H.E. Grossniklaus, Animal models of uveal melanoma. Melanoma Res, 2000. 10(3): p. 195-211. Syed, N.A., et al., Transgenic mice with pigmented intraocular tumors: tissue of origin and treatment. Invest Ophthalmol Vis Sci, 1998. 39(13): p. 2800-5. Albert, D.M., et al., Effectiveness of 1alpha-hydroxyvitamin D2 in inhibiting tumor growth in a murine transgenic pigmented ocular tumor model. Arch Ophthalmol, 2004. 122(9): p. 1365-9. Kramer, T.R., et al., Pigmented uveal tumours in a transgenic mouse model. Br J Ophthalmol, 1998. 82(8): p. 953-60. Latendresse, J.R., et al., Two cases of uveal amelanotic melanoma in transgenic TyrHRAS+ Ink4a/Arf heterozygous mice. Toxicol Pathol, 2007. 35(6): p. 827-32. Tolleson, W.H., et al., Spontaneous uveal amelanotic melanoma in transgenic TyrRAS+ Ink4a/Arf-/- mice. Arch Ophthalmol, 2005. 123(8): p. 1088-94. Mueller, A.J., et al., An orthotopic model for human uveal melanoma in SCID mice. Microvasc Res, 2002. 64(2): p. 207-13. Lattier, J.M., et al., Host pigment epithelium-derived factor (PEDF) prevents progression of liver metastasis in a mouse model of uveal melanoma. Clin Exp Metastasis, 2013. Chen, L., et al., A pilot study of vasculogenic mimicry immunohistochemical expression in intraocular melanoma model. Oncol Rep, 2009. 21(4): p. 989-94. Grossniklaus, H.E., Tumor vascularity and hematogenous metastasis in experimental murine intraocular melanoma. Trans Am Ophthalmol Soc, 1998. 96: p. 721-52. Onken, M.D., et al., Association between microarray gene expression signature and extravascular matrix patterns in primary uveal melanomas. Am J Ophthalmol, 2005. 140(4): p. 748-9. III Intravitreally injected HCmel12 melanoma cells serve as a mouse model of tumor biology of intraocular melanoma 34. 35. 52 Onken, M.D., et al., Collaborative ocular oncology group report number 1: prospective validation of a multi-gene prognostic assay in uveal melanoma. Ophthalmology, 2012. 119(8): p. 1596-603. Hurst, E.A., J.W. Harbour, and L.A. Cornelius, Ocular melanoma: a review and the relationship to cutaneous melanoma. Arch Dermatol, 2003. 139(8): p. 1067-73. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 53 IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model Marta M. Kilian1, Karin U. Loeffler1, Christian Kurts2, T. Hoeller3 Christiane Pfarrer4, Frank G. Holz1, Martina C. Herwig1 1 Department of Ophthalmology, University of Bonn, Bonn Germany 2 Institute of Experimental Immunology, University of Bonn, Bonn, Germany 3 Institute for Medical Biometry, Informatics and Epidemiology, University of Bonn, Bonn, Germany 4 Department of Anatomy, University of Veterinary Medicine Hannover, Hannover, Germany Manuscript in preparation IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 54 Abstract Purpose: Tumour associated macrophages (TAM), extravascular matrix patterns, and advanced patients’ age are associated with a poor prognosis in e.g. uveal melanoma. We concentrated on specific mechanisms of TAM under the influence of age to study the relation between the tumour microenvironment and tumour growth characteristics. Methods: Ninety five CX3CR1+/GFP mice (young 8-12 weeks, old 10-12 months) received an intravitreal injection of 1x105 HCmel12 melanoma cells. Subgroups were either systemically macrophage-depleted by Clodronate liposomes (n=23) or received melanoma cells, which were pre-incubated with the supernatant of M1- or M2-polarised macrophages (n=26). Eyes were processed histologically/immunohistochemically (n=75), or for flow cytometry (n=20) to analyse tumour size, mean vascular density (MVD), extravascular matrix patterns, extracellular matrix (ECM) and the presence/polarisation of TAM. Results: Extravascular matrix patterns were more frequently present in untreated old compared to young mice (p=0.024) as well as in untreated mice of both age groups compared to macrophage-depleted mice (p=0.014). Independent from age, M2-conditioned tumours showed more TAM (p=0.001), increased ECM levels (p=0.024) and a higher MVD (p=0.02) than M1-conditioned tumours. Flow cytometry showed a larger proportion of M2 macrophages in old than in young mice. Conclusions: The results indicate that old mice represent a more suitable tumour model compared with young mice since the histologic tumour pattern better resembles human tumours. TAM appear to be responsible for a more aggressive tumour phenotype. Tumour favouring and pro-angiogenic effects can directly be attributed to a M2-dominated microenvironment rather than to age-dependent factors alone. However, an aged IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 55 immunoprofile with an increased number of M2 macrophages may provide a tumourfavouring basis. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 56 Introduction Histologic tumour characteristics such as angiogenesis, vascular-like structures which are rich of PAS positive extracellular matrix (ECM), organised into fluid conducting, but nonendothelial channels and termed extravascular matrix patterns or vasculogenic mimicry as wells as inflammatory cells, in particular tumour associated macrophages (TAM) can be found in various tumours, such as uveal melanoma, cutaneous melanoma, breast cancer and others (Hendrix, Seftor et al. 2003; Mantovani, Marchesi et al. 2007; Allavena, Sica et al. 2008). In UM, outcome is associated with these characteristics and also correlates with patients’ age, tumour size, and extraocular extension (Foss, Alexander et al. 1996; Folberg, Mehaffey et al. 1997; Shields, Kaliki et al. 2013). Histopathologic factors are no longer crucial for prognosis, since chromosomal analysis and genetic profiling were found to be reliable indicators for patients’ outcome (Onken, Worley et al. 2012; Field and Harbour 2014). However, those histologic parameters reflect tumour growth characteristics and allow for their evaluation in animal models. The tumour microenvironment is characterised by a synergistic interplay between the tumour cells, the stroma and ECM, different inflammatory cell types, and soluble factors such as cytokines. Interactions between the tumour and the microenvironment may orchestrate tumour progression and metastasis (Catalano, Turdo et al. 2013) and a more detailed understanding of these interactions might help to interfere or even prevent these events. TAM are a major component of the inflammatory infiltrate in solid tumours. High numbers of TAM and their specific microenvironment correlate statistically with poor prognosis in different tumours (Allavena, Sica et al. 2008). In uveal melanoma, they are further associated with monosomy 3 and several histopathologic factors such as high mean vascular density (MVD) IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 57 and extravascular matrix patterns/ VM, and thus with a prognostically aggressive tumour phenotype (Folberg, Mehaffey et al. 1997; Makitie, Summanen et al. 2001; Hendrix, Seftor et al. 2003; Toivonen, Makitie et al. 2004; Maat, Ly et al. 2008). TAM functional polarisation may range within a broad spectrum from pro-inflammatory, tumouricidal M1 macrophages to anti-inflammatory, tumour-favoring M2 macrophages (Jager, Ly et al. 2011). Cytokines produced by tumour cells and other cells in the tumour microenvironment determine whether TAM become M1 or M2 (Arnold, Henry et al. 2007). In response to the tumour microenvironment TAM tend to specialise predominantly into the M2-phenotype with proangiogenic and tumour-favoring properties (Mantovani, Sozzani et al. 2002). Also, a high M2/M1 ratio in UM was shown to be related to worse prognosis and to histologic negative prognostic parameters (Herwig, Bergstrom et al. 2013). As the microenvironment and the capability of innate immunity changes with increasing age macrophages shift their secretory profile towards a M2-profile (Gomez, Nomellini et al. 2008). Macrophages from geriatric mice are also particularly sensitive to signals that promote their M2-polarisation, which in turn may promote tumour development (Jackaman, Radley-Crabb et al. 2013). Thus, the tumour microenvironment and advanced age have an influence on TAM polarisation. As macrophages play a pivotal role in the pathogenesis of several tumours and orchestrate the tumour microenvironment, we investigated their influence on tumour biology in a murine ocular tumour model. Herein, the tumour exhibits histologic features such as angiotropism, angiogenesis, extravascular matrix patterns/ VM and inflammatory cell infiltration which allow to study the intraocular tumour microenvironment. The latter can be further influenced by systemic drug application or intraocular injections (Kilian, Loeffler et al. 2015). In our study, we focussed on histopathologic factors with regard to tumour size and different forms of vascularisation. The general role of macrophages was examined in mice depleted of IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 58 macrophages in comparison to untreated mice. In addition, in order to address the impact of M1- or M2-polarised macrophages, we investigated intraocular tumours of M1- or M2conditioned melanoma cells. To study the influence of age on tumour growth patterns and macrophage invasion, the experiments were conducted both in young and in old mice. Materials and Methods Animals Breeding, housing and investigations of CX3CR1+/GFP mice were carried out and supervised according to accepted standards of the Policy on Humane Care and Use of Laboratory Animals of the National Institutes of Health (NIH), Bethesda, MD, USA. In CX3CR1+/GFP mice the transmembrane-receptor CX3 CR1 for CX3C chemokine fractalkine is replaced by green fluorescent protein (GFP) reporter gene. Hence, these mice exhibit GFP expression in peripheral blood monocytes, subsets of NK, dendritic cells and retinal microglia as well as macrophages, in which we were particularly interested (Jung, Aliberti et al. 2000). We studied intraocular tumours of 95 CX3CR1+/GFP mice in two different age groups. Young mice were examined at the age of 8-12 weeks; old mice were 10-12 months of age. In the first experiment, we investigated the age-dependent influence of TAM in 49 CX3CR1+/GFP mice. Macrophages were depleted systemically by repeated intraperitoneal injection of Clodronate liposomes in young (n=13) and old (n=10) mice and compared to untreated mice (young n=15, old n=11) (Tab. 1). In the second experiment, the impact of a micromilieu dominated IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 59 by M1 or M2 macrophages on tumour growth patterns was investigated in another cohort of 26 CX3CR1+/GFP mice (young M1: n=7, M2: n=7; old M1: n=6, M2: n=6) (Tab. 1). Tumours of 10 young and 10 old CX3CR1+/GFP mice were examined in regard to number of inflammatory cells, macrophages and their expression of the M2 marker CD163 by flow cytometry analysis (Tab. 1). Table 1: Number of animals for macrophage (MФ) depletion and polarisation experiment as well as flow cytometry analysis Experiment Young mice (8-12 weeks) Old mice (10-12 months) MФ depletion MФ polarisation FACS MФ Presence MФ Depletion M1conditioned tumours M2conditioned tumours n= 15 n= 13 n= 7 n= 7 n= 10 n= 11 n= 10 n= 6 n= 6 n= 10 Mouse model of intraocular melanoma According to a previously established mouse model of metastasizing ocular melanoma (Diaz, Rusciano et al. 1999), we created a modified mouse model of intraocular melanoma to study tumour biology (Kilian, Loeffler et al. 2015). In our model, 1x105 HCmel12 cells (a murine cutaneous melanoma cell line) were injected intravitreally into the right eye of each mouse in all study groups on day 0 under general anaesthesia. Mice were sacrificed on day 9 post intravitreal injection, the right eye was enucleated and processed for histologic or immunohistochemical examination or for flow cytometry analysis. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 60 Macrophage depletion (experiment 1) For the first experiment, macrophage-depleted mice were studied in comparison to untreated mice. Liposome-encapsulated dichloromethylene diphosphonate (Cl2MDP; Clodronate, ordered through clodronateliposomes.org) was used to systemically deplete blood monocytes and macrophages at a dosage of 0.1mg/g body weight (van Rooijen and van KesterenHendrikx 2003). Mice received repeated intraperitoneal injections of Clodronate liposomes on days -6, -3, 0 (day of intravitreal tumour cell injection) and day 3. To evaluate macrophage depletion, liver sections of Clodronate-treated and untreated mice were stained immunohistochemically with a GFP-antibody (GFP Rabbit IgG, Life Technologies, Carlsbad, CA, USA; dilution 1:100) or F4/80-antibody (F4/80 Antibody, Cl:A3-1, AbD Serotec, Oxford, U.K.; dilution 1:100). Accordingly, HCmel12 melanoma cells were cultured in vitro in standard medium or in supernatant of unpolarized macrophages for 20 hours. Then, cell proliferation rates were compared by a BrdU cell proliferation assay (BrdU Cell Proliferation Assay, Milipore, Billerica, MA; USA). Macrophage polarisation (experiment 2) For the second experiment, HCmel12 melanoma cells were exposed in vitro to the supernatant of polarised macrophages prior to intravitreal injection. Murine macrophages were cultured in medium without fetal calf serum (FCS) for two hours prior to in vitro polarisation to avoid any impact of cytokines or chemokines within the FCS. Macrophages were treated with different cytokines and lipopolysaccharide (LPS) to obtain different polarised phenotypes according to a modified protocol (Mosser and Zhang 2008). IFNγ (200ng/ml) for M1macrophages and Interleukin 4 (IL-4) (20ng/ml) for M2 macrophages were added to the serum-free medium in order to polarise macrophages. After 10 hours of incubation, LPS IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 61 (100ng/ml) was supplemented for each macrophage type for a total incubation time of 24 hours. Polarisation status of unfixed macrophages was verified by using a multiple cytokine ELISA comprising the inflammatory cytokines IL-6, IL-10. IL-12, IL-23, TNFα and IFNγ (Multi-Analyte ELISArrayTM Kit, Mouse Mix-N-Match, Qiagen, Venlo, Netherlands). M1specific expression of inflammatory cytokines comprises characteristic levels of IL-6high, IL10low, IL-12high, IL-23high, TNFαhigh and IFNγhigh (M2-specific expression levels vice versa) (Mantovani, Sica et al. 2004). Phenotypical macrophage polarisation status of 4% PFA-fixed cells was examined by immunocytology with a double staining kit (MultiVision Polymer Detection System, Thermo Scientific, Waltham, MA, USA) using a F4/80- (for all macrophages) and CD163- (for M2 macrophages only; CD163, M-96, Santa Cruz Biotechnology, Santa Cruz, CA, USA) primary antibody. To examine the impact of a M1- or M2-conditioned tumour microenvironment, HCmel12 melanoma cells were treated with serum-free medium for two hours and afterwards exposed to the supernatant of M1- or M2polarised macrophages for 20 hours, respectively. These M1- or M2-conditioned HCmel12 melanoma cells were then injected intravitreally as described above. Cell proliferation rates and growth characteristics of these M1- or M2-conditioned HCmel12 melanoma cells were then compared in vitro using a BrdU proliferation assay and in vivo after intravitreal injection. Histology and immunohistochemistry Processing of histologic specimens of both experiments was carried out as described before (Kilian, Loeffler et al. 2015). Briefly, in hematoxylin-eosin (H&E) stained paraffin sections, tumour size was measured in mm² at 40x magnification using ImageJ 1.45s (W Rasband, National Institute of Health (NIH), USA); vascular characteristics like angiotropism, cooption of retinal vessels and intratumoural angiogenesis were evaluated at 200x IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 62 magnification. Sections stained with periodic acid-Schiff reaction (PAS) without hematoxylin were investigated for analysis of ECM-rich channels/vasculogenic mimicry along with their prognostic significant extravascular matrix patterns. For determination of the MVD, sections were stained with a CD31-antibody (CD31, SZ31, Dianova, Hamburg, Germany, dilution 1:10). At 200x magnification, vessels were counted within a 0.25mm2 grid in 1-3 areas according to Foss et al. (Foss, Alexander et al. 1996). Staining of ECM was additionally performed with a collagen IV antibody (Collagen IV Antibody, Novus Biologicals, Littleton, CO, USA dilution 1:100) and a laminin antibody (Laminin Antibody, Novus Biologicals, Littleton, CO, USA, dilution 1:100). To distinguish the genuine ocular pigment from the immunoreaction we visualised the latter with the red chromogen AEC. Macrophage infiltration of the eye and of ocular tumours was verified by a GFP-antibody. Number of infiltrating macrophages as well as the presence and amount of collagen IV- and lamininpositive structures were analysed semi-quantitatively from 0 to +++ by two independent examiners (MCH, MMK). Flow cytometry analysis Ocular tumours of 10 young and 10 old mice were dissected microsurgically, pooled, respectively, and digested in 0.5mg/ml collagenase and 100µg/mg DNAase I in RPMI medium for 40 minutes. The washed cells were filtered through a 100µm nylon mesh. Surface staining was performed with monoclonal antibodies for CD45 (Biolegend Inc., San Diego, CA, USA), CD11b (Biolegend), F4/80 (Biolegend), CD80 (Biolegend), CD163 (M-96, Santa Cruz Biotechnology, Santa Cruz, CA, USA + conjugation kit, Abcam, Cambridge, UK), and Gr-1 (Biolegend). Controls were run as single stains and unstained probes. Measurements IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 63 were carried out on a flow cytometer (LSRFortessa, BD, Franklin Lakes, NJ, USA) and results were analysed by using FlowJo software (Tristar Inc. San Carlos, CA, USA). Statistical analysis Statistical analysis was performed with IBM SPSS Statistics 22.0 (IBM Corp, Armonk, NY, USA). Tumour size in mm2 and MVD of different study groups were evaluated by an analysis of variance and a Post Hoc Test. Chi2 test and Pearson's correlation coefficient were used to determine the relationship between different study groups and tumour growth characteristics. Student’s t-test and ROC analysis was applied for interpretation of cytokine levels of in vitro polarised macrophages. Comparison of in vitro melanoma cell proliferation between differently treated cells was verified by student’s t-test. A p≤0.05 was considered as statistically significant and significance level was indicated by using * for p ≤ 0.05. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 64 Results Macrophage depletion model (experiment 1) All 49 CX3CR1+/GFP mice that received an intravitreal HCmel12 tumour cell injection developed an intraocular tumour. Inflammatory cell infiltration was examined on H&E stained sections as well as immunohistochemically. Intraocular tumours of untreated mice exhibited high macrophage recruitment at an even distribution throughout the tumour and within the retina, often associated with vascular structures. Apart from two very small tumours, all tumours exhibited angiotropism, co-option of retinal vessels and formed own blood conducting structures. Immunohistochemical staining for CD31 indicated endotheliumlined vasculature in the tumours. MVD revealed no significant differences between the study groups of different age and macrophage status (p=0.542). In addition to endothelium-lined vessels, specific extravascular matrix patterns were present in all tumours, including areas with PAS-positive straights, parallels with and without cross-links, arcs with and without branching, and loops. Since Folberg et al. showed PAS-positive parallels with cross-links, loops and networks to be of prognostic value in choroidal melanoma (Folberg, Mehaffey et al. 1997), we focused on these three extravascular matrix patterns (see below). Intraperitoneal injection of Clodronate liposomes led to systemic macrophage depletion which could be verified in immunohistochemical stains of the liver: while untreated mice exhibited frequent F4/80-posititive staining of Kupffer cells, Clodronate liposome-treated mice of both age groups lacked positively stained cells (Supplementary Fig. S1). IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 65 Supplementary Figure S1: Experiment 1, systemic macrophage status confirmed by immunohistochemical staining for F4/80 (red), x200 magnification. (A) Liver section of untreated mouse; (B) Liver section of macrophage-depleted mouse. Intraocular tumours of untreated old mice exhibited more frequently extravascular matrix patterns than tumours of untreated young mice (p=0.024) when analysed in PAS stained sections without hematoxylin (Fig. 1). In tumours of untreated mice of both ages these patterns were significantly more frequent than in systemically macrophage-depleted mice of both age groups (p=0.014). When subdivided into young and old mouse eyes, this effect was much more pronounced in old mice (pyoung=0.34, pold=0.012). IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 66 Figure 1: Experiment 1, ocular tumours of young (A-B), old (C-D) and old macrophage-depleted (E-F) mice, x100 magnification; (A,C,E) Sections stained with H&E giving an overview of the tumour with solid tumour growth, angiotropism and angiogenesis; (B,D,F) sections stained with PAS without hematoxylin showing extravascular matrix pattern (arrows); x100 magnification. Prognostically significant extravascular matrix pattern were more frequently found in untreated old mice in comparison to untreated young (p=0.024) as well as between untreated mice of both age groups in comparison with macrophage-depleted mice (p=0.014). IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 67 Collagen IV staining labelled endothelium dependent vessels, extravascular matrix patterns as well as other ECM-rich structures in the tumour. However, these collagen IV-positive patterns did not exactly represent the patterns seen with PAS w/o hematoxylin. There was no significant difference in collagen IV expression between the four study groups (p=0.213). However, aged mice showed a tendency of having more collagen IV positive structures compared to the young (p=0.074). Intraocular tumour size [in mm2] was statistically not correlated with macrophage status or age (p=0.529). The presence of prognostic significant extravascular matrix patterns was also not correlated with tumour size (p=0.611). However, in vitro melanoma cell proliferation was increased significantly in cells which were incubated in macrophage supernatant in comparison to cells cultured in standard medium (p=0.005) (Fig. 2A). Macrophage polarisation model (experiment 2) Macrophage polarisation in vitro Macrophage polarisation was functionally verified and confirmed by a multiple cytokine ELISA as described in methods. Levels of IL-12 and IL-23 were significantly higher in M1than in M2-polarised macrophages (pIL-12=0.043, pIL-23=0.012) and IL-10 levels were significantly higher in M2 macrophages (pIL-10=0.008) (Supplementary Fig. S2). Levels of IL6, TNFα and IFNγ levels were exceedingly high and could not be used for statistical analysis. IFNγ levels were not evaluated for M1 macrophages due to their initial polarisation with this cytokine. However, IFNγ level within the M2-polarised culture was evidently low, in agreement with reported M2 characteristics. Further, immunocytological dual labelling with the M2 marker CD163 and the murine macrophage marker F4/80 revealed increased numbers of CD163 labelled cells in the M2-polarised macrophage cell culture compared to the M1- IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 68 polarised cell culture (ratio CD163:F4/80 in M1-polarised culture 4:10; in M2-polarised culture 7:10) (Supplementary Fig. S3). The combination of the functional characterization by the multiple cytokine ELISA and phenotypical staining suggests a reliable in vitro polarisation of macrophages. Supplementary Figure S2: Experiment 2, multiple cytokine ELISA of macrophage polarisation in vitro; M0: unpolarised macrophages (MФ), M1: MФ polarised with IFNγ+LPS, M2: MФ polarised with IL-4+LPS, MM: HCmel12 melanoma cells, MM+M0 Co: HCmel12 melanoma cells cultured in M0 supernatant, Neg: negative; characteristic cytokine levels for M1- and M2-polarised MФ with significant differences in IL-10. IL-12, IL-23 expression, IL-6 and TNFα levels were exceeding measurable dimension for M1- and M2-polarised MФ, IFNγ was also exceeding measurable dimension for M1 MФ, but was evidently low for M2 MФ. HCmel12 melanoma cells expressed no reasonable levels of investigated cytokines, however, after co-incubation with M0 supernatant elevated levels of IL-6 and TNFα were measured. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 69 Supplementary Figure S3: Experiment 2, immunocytochemistry of in vitro polarised macrophages. Ratio M2 macrophage marker CD163 (blue) / pan-macrophage marker F4/80 (red). (A) Macrophages after M1 polarisation (ratio 0.42 with predominantly round morphology); (B) Macrophages after M2 polarisation (ratio 0.74 with predominantly dendritic morphology). Macrophage conditioned melanoma cells in vivo and in vitro Mice having received an intravitreal injection of M2-conditioned HCmel12 melanoma cells exhibited a higher degree of inflammatory cell infiltration in ocular (retina and uvea) and tumoural structures than mice with M1-conditioned tumours. Accordingly, increased levels of GFP-labelled macrophages were found in tumours and retinas of mice with M2-conditioned tumours compared to M1-conditioned tumours (p=0.001) (Fig. 3). Comparing M1- and M2conditioned tumours, age did not have an impact on the degree of inflammatory cell infiltration of the tumour bearing eye (p=0.747). In all mice, angiotropism, co-option of retinal vessels and intratumoural blood conducting structures were identified immunohistochemically and by H&E staining. Mice which had received an intravitreal injection of M2-conditioned HCmel12 melanoma cells exhibited a statistically higher MVD compared to mice with M1-conditioned tumours (p=0.02). Age did not have an impact on the MVD (p=0.132). Immunohistochemically, collagen IV-staining IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 70 was detected in all tumours. The staining reaction was found predominantly at tumour borders and mostly sparing the central area of tumours (Fig. 3). Collagen IV-positive structures within the tumour were increased in M2-conditioned tumours compared to M1-conditioned tumours (p=0.024) but were not age dependent (p=0.84). In PAS sections without hematoxylin, extravascular matrix patterns were not unequivocally evaluable and were thus not statistically evaluated. Tumour size was not correlated with previous M1- or M2-conditioning of HCmel12 cells, nor with age (p=0.179). ). However, in vitro cell proliferation was significantly increased in melanoma cells which were previously incubated in M2-polarised supernatant in comparison to cells cultured in supernatant of M1-polarised macrophages (p=0.005) (Fig. 2B). Figure 2: BrdU cell proliferation assay of HCmel12 melanoma cells (MM) which were cultured in standard medium (standard), supernatant of unpolarized macrophages (M0) or of polarized macrophages (M1, M2). Melanoma cells cultured in unpolarized macrophage supernatant (M0) showed a significantly higher proliferation rate (p=0.005) than cells cultured in standard medium (standard) (A). Melanoma cells cultured in supernatant of M2-polarized macrophages (M2) exhibited a significantly higher proliferation rate compared to cells cultured in supernatant of M1-polarized macrophages (M1) (p=0.005). IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 71 Figure 3: Experiment 2, ocular tumours of M1- (A-D) and M2-conditioned tumour cells (E-H), H&E and immunohistochemical staining, x100 magnification; significant differences between M1- and M2-conditioned tumours regarding microvessel density (MVD, p=0.02), collagen IV expression (extracellular matrix (ECM), p=0.024) and level of macrophage infiltration (GFP, p=0.001). IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 72 Flow cytometry analysis Pooled ocular tumours of young (n=10) and old (n=10) CX3CR1+/GFP mice were investigated for infiltrating leucocytes (CD45). The fraction of CX3CR1 GFP positive and F4/80 positive macrophages was subclassified into bone marrow derived infiltrating macrophages (F4/80 positive, GFPhigh, Gr-1high) and resident macrophages (microglia) (F4/80 positive, GFPhigh, Gr1low) as well as M1 macrophages (CD80high, CD163low) and M2 macrophages (CD80intermediate, CD163high). In pooled intraocular tumours 7.44% of cells in young and 7.1% of cells in old mice were CD45 positive leucocytes. The majority of these (70.43% in young, 72.97% in old mice) were CD11b positive phagocytes of which 79.13% (in young) and 74% (in old) were found to be GFP positive cells (CX3CR1). Almost all GFP positive cells were macrophages (F4/80 positive). Old mice showed a slight majority of CD163 positive cells (M2 marker) when compared to young mice. However, noteworthy differences were found between bone marrow recruited macrophages (Gr-1 positive) infiltrating the tumour and resident macrophages (Gr-1 negative) with regard to age. Young mice exhibited increased levels of Gr-1 positive macrophages when compared to the old (young 73.27%, old 62%, p=0.004) (Supplementary Fig. S4). Monocytes which are recruited into the tumour via the blood circulation migrate into the tumour as unpolarised phenotypes and proliferate into macrophages. Depending on the existing tumour microenvironment, they are polarised into tumouricidal M1 or tumour-favoring M2 macrophages. In our study, bone marrow derived and infiltrating macrophages (Gr-1 positive) exhibited more frequently M2 markers (CD163) in old mice than in their younger counterparts (p=0.08) (Fig. 4). This suggests a M2dominated tumour microenvironment in old mice (Tab. 2). IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model CD45 Young mice Old mice 7.44 7.10 70.43 72.97 79.13 74.00 35.77 33.90 98.67 97.70 30.20 31.90 73.27 62.00 99.83 99.50 32.47 37.73 + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + CD45 CD11b CD45 CD11b GFP CD45 CD11b GFP F4/80 CD45 CD11b GFP F4/80 CD80 high CD45 CD11b GFP F4/80 CD163 CD45 CD11b GFP F4/80 Gr-1 high CD45 CD11b GFP F4/80 Gr-1 CD80 high CD45 CD11b GFP F4/80 Gr-1 CD163 high 73 Supplementary Figure S4: Flow cytometry analysis of surface markers of tumour associated inflammatory cells [in%], in particular macrophages, in young (n=10) and old (n=10) mice. 45,00 + Percentage of CD45 CD11b GFP Gr-1 cells M2 Marker CD163 + + 40,00 35,00 + 37,73 30,00 32,47 25,00 20,00 Young mice Old mice Figure 4: Flow cytometry analysis of tumour associated inflammatory macrophages (CD45+CD11b+GFP+Gr-1+) in intraocular melanoma regarding their expression of the M2 marker CD163. In old mice, these bone marrow derived macrophages exhibited more frequently a M2 phenotype with elevated levels of CD163high macrophages than their younger counterparts (p=0.08). 74 IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model Table 2: Overview of tumour characteristics comparing untreated young and untreated old mice. Young mice Old mice Measurement p-value 0.68 ± 0.048 0.88 ± 0.035 Vessel/mm2 0.138 46.66 % 90.9 % Presence yes/no 0.024 1.65 2.26 φ score (0-3) 0.074 Intratumoural CD45+ cells 7.44 % 7.1 % Percentage 0.11 Inflammatory Gr-1+ cells 73.27 % 62.0 % Percentage 0.025 CD163+ M2 macrophages 32.47 % 37.73 % Percentage 0.087 MVD (mean) Extravascular matrix pattern Collagen IV Tumours in old mice have a more aggressive phenotype and better resemble human malignancies in terms of higher MVD, presence of prognostically significant extravascular matrix pattern, amount of collagen IV, intratumoural leucocytes, bone marrow derived and tumour infiltrating macrophages and M2 polarised macrophages. Discussion Certain tumour characteristics which are part of the tumour microenvironment appear to represent general features of tumour biology. Mouse models allow to study such features, including immunologic aspects or different forms of vascularisation (Chen, Zhang et al. 2009; Kilian, Loeffler et al. 2015). In our mouse model, cutaneous melanoma cells are injected into the mouse eye. Resulting tumours exhibit histologic features such as angiotropism, VM/ extravascular matrix patterns, and inflammatory cell infiltration. Since the eye represents a IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 75 semi-closed system with access to blood vessels (but no lymphatic vessels), the intraocularly injected tumour cells may grow inside an immune privileged compartment towards a tumour mass and develop a microenvironment which can be iatrogenically manipulated by systemic or intraocular injections (Kilian, Loeffler et al. 2015). Thus, our model provides the opportunity to address hypotheses on general tumour biology including vascularisation and matrix remodelling. As particularly macrophages play a pivotal role in various tumours including UM, we used our established mouse model to specifically study their influence on intraocular growth characteristics. The role of TAM To determine the impact of macrophages (regardless of their polarisation) we compared intraocular melanoma growth characteristics in vivo in systemically macrophage- depleted mice in comparison to untreated mice. Depletion of macrophages can result in tumour growth inhibition and reduced angiogenesis as shown in an athymic mouse model of cutaneous melanoma (Gazzaniga, Bravo et al. 2007). Another animal study of an intraocular melanoma model also showed a tumour promoting role of M2 macrophages for tumour outgrowth in old mice (Ly, Baghat et al. 2010). In our model, tumour progression in terms of tumour size did not correlate with macrophage status or age. However, in vitro results showed increased cell proliferation rates in melanoma cells which were grown in an environment (supernatant) shaped by unpolarised macrophages. In vivo, advanced age and the presence of macrophages were individually associated with an increased occurrence of prognostically significant extravascular matrix patterns and thus, with an aggressive phenotype. Melanoma cells are able to actively remodel the ECM in order to arrange stromal patterns such as vascular networks (Daniels, Boldt et al. 1996). Remodelling of the microcirculation to form such IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 76 vascular networks appears a precondition for metastasis in UM und thereby a determinant for patient’s outcome. Besides tumour cells themselves, macrophages are also potentially able to enhance extravascular matrix pattern formation, tumour remodelling and angiogenesis by producing specific inflammatory cytokines or enzymes (Seftor, Seftor et al. 2001). The effect of an increased frequency of prognostically significant extravascular matrix patterns in aged and untreated mice might be directly related to the crosstalk between melanoma cells and TAM. The impact of M2 macrophages A tumour may take advantage of an inflammatory microenvironment that is dominated by M2 macrophages. In many tumours including uveal melanoma infiltrating TAM are indeed of a M2 polarisation phenotype and are statistically correlated with tumour progression (Mantovani, Sozzani et al. 2002; Gordon and Martinez 2010; Mantovani and Sica 2010). Our results from the polarisation experiment display tumour characteristics under iatrogenically induced extreme tumour microenvironments of M1 or M2 dominance. They do not model actual conditions in intraocular tumours but demonstrate in which ways polarised macrophages are capable of influencing tumour characteristics (and thus prognosis). In vitro as well as in vivo, our results point to far more aggressive tumour growth patterns in M2conditioned tumours in terms of tumour cell proliferation rates and histologic growth characteristics. Infiltration with leucocytes was much more pronounced in M2- than in M1conditioned tumours whereby increased numbers of lymphocytes and macrophages were found throughout the tumour but also throughout adjacent ocular structures, particularly in vicinity to vessels. In uveal melanoma, high lymphocytic infiltration of the tumour is often associated with an increased number of M2 macrophages which in turn is correlated with IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 77 monosomy 3 and thus with a poor prognosis (Bronkhorst, Vu et al. 2012). Furthermore, in our M2-conditioned tumours collagen IV-positive matrix patterns were more frequently present, indicating an increased tissue remodelling compared to M1-conditioned tumours. Collagen IV positive ECM is correlated with tumour size and/ or a worse prognosis in different tumour entities (Krecicki, Zalesska-Krecicka et al. 2001; Ioachim, Charchanti et al. 2002; Pierard, Pierard-Franchimont et al. 2012). Changes in the composition and content of ECM influence tumour and stromal properties and may promote metastatic spread (Gilkes, Semenza et al.). Further, stromal cells are important contributors to the tumour’s microenvironment as released mediators such as metalloproteinases may promote tissue remodelling and cell migration. In cutaneous melanoma, among other factors, collagen IV was found to influence tumour cell mobility and thus may contribute to tumour progression and may possibly be a prognostic indicator (Pierard, Pierard-Franchimont et al. 2012). A synergistic interplay between the host and the tumour aiming for increased tissue remodelling and angiogenesis is crucial for the tumour’s circulation and progression (Catalano, Turdo et al. 2013). In line with this, co-option of host vessels and increased collagen IV rates were found predominantly adjacent to the retina (i.e. at the edge of the tumour) in our model. As these effects were pronounced in tumours with M2-conditioned melanoma cells, such a remodelling effect might be attributed to a microenvironment dominated by M2 macrophages. In addition, MVD was also significantly increased in M2-conditioned tumours. MVD is a histopathological indicator associated with tumour microvessels that has independent prognostic significance in uveal melanoma and other tumour entities. It is further correlated with largest basal diameter and abundance of TAM and thus with a worse prognosis (Makitie, Summanen et al. 1999; Makitie, Summanen et al. 2001). Enhanced angiogenesis in terms of an increased MVD in M2-conditioned tumours, again, points to a more aggressive phenotype, IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 78 which supports the paradigm of pro-angiogenic, tumour-favoring effects of M2-polarised macrophages. In our study, advanced age did not have an impact on growth characteristics of the conditioned ocular tumours while M2-conditioned tumours exhibited a worse phenotype than M1-conditioned tumours. This suggests that the impact on poor prognosis can be attributed more directly to an increased M2 influence rather than to other age-dependent factors alone. The impact of age Since advanced age was associated with an increased occurrence of M2 macrophages and more frequent extravascular matrix patterns, we concluded that old mice represent a better tumour model than young mice. Thus, for questions related to the tumour microenvironment old mice represent a better animal model than their younger counterparts and should be used in studies. This is in accordance with the situation in human tumours where age and extravascular matrix patterns are associated with a worse prognosis. Apparently, advanced age has an impact on the recruitment and functional phenotype of macrophages (Gomez, Nomellini et al. 2008). In our model, bone marrow derived TAM (Gr-1 positive) exhibited increased M2 phenotypical characteristics in old mice when compared to the young, as shown by flow cytometry analysis. This effect was slightly beneath statistical significance (p=0.08). However, since macrophage polarisation is a continuum and other studies are in accordance with our observation, the tumour microenvironment in old mice appears to be M2-dominated. This is in line with the fact that macrophages from geriatric mice are particularly sensitive to signals that promote their M2 polarisation (Jackaman, Radley-Crabb et al. 2013). Further, in an anterior chamber melanoma model tumour progression and outgrowth in old mice depended on the presence of macrophages which had a IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 79 predisposition for the M2 phenotype (Ly, Baghat et al. 2010). This suggests a M2-driven progression and thus an influence on the patient’s prognosis. However, in our polarisation experiment we could not document a significant influence of advanced age on histopathologic growth factors when compared to the young. This might be attributed to the dominant effect of the iatrogenically manipulated M2 microenvironment so that the age’s impact diminished. Overall, as old mice exhibited a M2-dominated tumour microenvironment an aged immunoprofile may provide a basis for more aggressive tumours while a progressing aggressive tumour phenotype is depending on infiltrating macrophages and their M2 phenotype. Summary Old mice exhibited more aggressive tumour phenotypes and thereby mimicking human intraocular melanomas more appropriately. Pro-angiogenic and tumour promoting factors such as extravascular matrix patterns, stroma remodelling and MVD could be directly attributed to the existence of TAM in vivo, in particular to M2 macrophages. In vitro results confirmed increased tumour malignancy in M2-dominated conditions. Since old mice also exhibited increased numbers of M2 macrophages in the tumour, an aged immunoprofile provides a basis for tumour growth with M2 macrophages being directly involved into a more aggressive phenotype. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 80 Acknowledgements We gratefully acknowledge research funding from BONFOR (University of Bonn) and the “Verein der Rheinisch-Westfälischen Augenärzte (RWA)”. We have no financial disclosures to report. We would like to thank Hans E. Grossniklaus (Emory University, Atlanta, GA, USA) for scientific guidance, Thomas Tüting (Laboratory of Experimental Dermatology, University of Bonn, Bonn, Germany) for kindly providing the HCmel12 murine melanoma cell line and Eicke Latz (Institute of Innate Immunity, University of Bonn, Bonn, Germany) for kindly providing the murine macrophages for in vitro experiments. Further we are thankful to André Tittel (Institute of Experimental Immunology, University of Bonn, Bonn, Germany) and Tobias Bald (Laboratory for Experimental Dermatology, University of Bonn, Bonn, Germany) for their assistance in flow cytometry and management of the melanoma cell line, respectively. Especially, we want to thank Parand Widmar and Claudine Strack (Department of Ophthalmology, University of Bonn, Bonn, Germany) for their assistance in histology. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 81 References 1. Mantovani A, Marchesi F, Porta C, Sica A, Allavena P. Inflammation and cancer: breast cancer as a prototype. Breast 2007;16 Suppl 2:S27-33. 2. Hendrix MJ, Seftor EA, Hess AR, Seftor RE. Vasculogenic mimicry and tumor-cell plasticity: lessons from melanoma. Nat Rev Cancer 2003;3:411-21. 3. Allavena P, Sica A, Solinas G, Porta C, Mantovani A. The inflammatory microenvironment in tumor progression: the role of tumor-associated macrophages. Crit Rev Oncol Hematol 2008;66:1-9. 4. Shields CL, Kaliki S, Furuta M, Fulco E, Alarcon C, Shields JA. American Joint Committee on Cancer classification of posteriour uveal melanoma (tumor size category) predicts prognosis in 7731 patients. Ophthalmology 2013;120:2066-71. 5. Folberg R, Mehaffey M, Gardner LM, Meyer M, Rummelt V, Pe'er J. The microcirculation of choroidal and ciliary body melanomas. Eye (Lond) 1997;11 ( Pt 2):22738. 6. Foss AJ, Alexander RA, Jefferies LW, Hungerford JL, Harris AL, Lightman S. Microvessel count predicts survival in uveal melanoma. Cancer Res 1996;56:2900-3. 7. Onken MD, Worley LA, Char DH, Augsburger JJ, Correa ZM, Nudleman E, Aaberg TM, Jr., Altaweel MM, Bardenstein DS, Finger PT, Gallie BL, Harocopos GJ, et al. Collaborative ocular oncology group report number 1: prospective validation of a multi-gene prognostic assay in uveal melanoma. Ophthalmology 2012;119:1596-603. 8. Field MG, Harbour JW. Recent developments in prognostic and predictive testing in uveal melanoma. Curr Opin Ophthalmol 2014;25:234-9. 9. Catalano V, Turdo A, Di Franco S, Dieli F, Todaro M, Stassi G. Tumor and its microenvironment: a synergistic interplay. Semin Cancer Biol 2013;23:522-32. 10. Makitie T, Summanen P, Tarkkanen A, Kivela T. Tumor-infiltrating macrophages (CD68(+) cells) and prognosis in malignant uveal melanoma. Invest Ophthalmol Vis Sci 2001;42:1414-21. 11. Toivonen P, Makitie T, Kujala E, Kivela T. Microcirculation and tumor-infiltrating macrophages in choroidal and ciliary body melanoma and corresponding metastases. Invest Ophthalmol Vis Sci 2004;45:1-6. 12. Maat W, Ly LV, Jordanova ES, de Wolff-Rouendaal D, Schalij-Delfos NE, Jager MJ. Monosomy of chromosome 3 and an inflammatory phenotype occur together in uveal melanoma. Invest Ophthalmol Vis Sci 2008;49:505-10. 13. Jager MJ, Ly LV, El Filali M, Madigan MC. Macrophages in uveal melanoma and in experimental ocular tumor models: Friends or foes? Prog Retin Eye Res 2011;30:129-46. 14. Arnold L, Henry A, Poron F, Baba-Amer Y, van Rooijen N, Plonquet A, Gherardi RK, Chazaud B. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J Exp Med 2007;204:1057-69. 15. Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol 2002;23:549-55. 16. Herwig MC, Bergstrom C, Wells JR, Holler T, Grossniklaus HE. M2/M1 ratio of tumor associated macrophages and PPAR-gamma expression in uveal melanomas with class 1 and class 2 molecular profiles. Exp Eye Res 2013;107:52-8. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 82 17. Gomez CR, Nomellini V, Faunce DE, Kovacs EJ. Innate immunity and aging. Exp Gerontol 2008;43:718-28. 18. Jackaman C, Radley-Crabb HG, Soffe Z, Shavlakadze T, Grounds MD, Nelson DJ. Targeting macrophages rescues age-related immune deficiencies in C57BL/6J geriatric mice. Aging Cell 2013;12:345-57. 19. Kilian MM, Loeffler KU, Pfarrer C, Holz FG, Kurts C, Herwig MC. Intravitreally Injected HCmel12 Melanoma Cells Serve as a Mouse Model of Tumor Biology of Intraocular Melanoma. Curr Eye Res 2015:1-8. 20. Jung S, Aliberti J, Graemmel P, Sunshine MJ, Kreutzberg GW, Sher A, Littman DR. Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol Cell Biol 2000;20:4106-14. 21. Diaz CE, Rusciano D, Dithmar S, Grossniklaus HE. B16LS9 melanoma cells spread to the liver from the murine ocular posteriour compartment (PC). Curr Eye Res 1999;18:125-9. 22. van Rooijen N, van Kesteren-Hendrikx E. "In vivo" depletion of macrophages by liposome-mediated "suicide". Methods Enzymol 2003;373:3-16. 23. Mosser DM, Zhang X. Activation of murine macrophages. Curr Protoc Immunol 2008;Chapter 14:Unit 14 2. 24. Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 2004;25:677-86. 25. Chen L, Zhang S, Li X, Sun B, Zhao X, Zhang D, Zhao S. A pilot study of vasculogenic mimicry immunohistochemical expression in intraocular melanoma model. Oncol Rep 2009;21:989-94. 26. Gazzaniga S, Bravo AI, Guglielmotti A, van Rooijen N, Maschi F, Vecchi A, Mantovani A, Mordoh J, Wainstok R. Targeting tumor-associated macrophages and inhibition of MCP-1 reduce angiogenesis and tumor growth in a human melanoma xenograft. J Invest Dermatol 2007;127:2031-41. 27. Ly LV, Baghat A, Versluis M, Jordanova ES, Luyten GP, van Rooijen N, van Hall T, van der Velden PA, Jager MJ. In aged mice, outgrowth of intraocular melanoma depends on proangiogenic M2-type macrophages. J Immunol 2010;185:3481-8. 28. Daniels KJ, Boldt HC, Martin JA, Gardner LM, Meyer M, Folberg R. Expression of type VI collagen in uveal melanoma: its role in pattern formation and tumor progression. Lab Invest 1996;75:55-66. 29. Seftor RE, Seftor EA, Koshikawa N, Meltzer PS, Gardner LM, Bilban M, StetlerStevenson WG, Quaranta V, Hendrix MJ. Cooperative interactions of laminin 5 gamma2 chain, matrix metalloproteinase-2, and membrane type-1-matrix/metalloproteinase are required for mimicry of embryonic vasculogenesis by aggressive melanoma. Cancer Res 2001;61:6322-7. 30. Mantovani A, Sica A. Macrophages, innate immunity and cancer: balance, tolerance, and diversity. Curr Opin Immunol 2010;22:231-7. 31. Gordon S, Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity 2010;32:593-604. 32. Bronkhorst IH, Vu TH, Jordanova ES, Luyten GP, Burg SH, Jager MJ. Different subsets of tumor-infiltrating lymphocytes correlate with macrophage influx and monosomy 3 in uveal melanoma. Invest Ophthalmol Vis Sci 2012;53:5370-8. IV Impact of macrophages on tumour growth characteristics in a murine ocular tumour model 83 33. Ioachim E, Charchanti A, Briasoulis E, Karavasilis V, Tsanou H, Arvanitis DL, Agnantis NJ, Pavlidis N. Immunohistochemical expression of extracellular matrix components tenascin, fibronectin, collagen type IV and laminin in breast cancer: their prognostic value and role in tumor invasion and progression. Eur J Cancer 2002;38:2362-70. 34. Pierard GE, Pierard-Franchimont C, Delvenne P. Malignant melanoma and its stromal nonimmune microecosystem. J Oncol 2012;2012:584219. 35. Krecicki T, Zalesska-Krecicka M, Jelen M, Szkudlarek T, Horobiowska M. Expression of type IV collagen and matrix metalloproteinase-2 (type IV collagenase) in relation to nodal status in laryngeal cancer. Clin Otolaryngol Allied Sci 2001;26:469-72. 36. Gilkes DM, Semenza GL, Wirtz D. Hypoxia and the extracellular matrix: drivers of tumor metastasis. Nat Rev Cancer;14:430-9. 37. Makitie T, Summanen P, Tarkkanen A, Kivela T. Microvascular density in predicting survival of patients with choroidal and ciliary body melanoma. Invest Ophthalmol Vis Sci 1999;40:2471-80. V Origin of intratumoral vascular structures in a murine ocular tumour model 84 V Origin of intratumoural vascular structures in a murine ocular tumour model Marta M. Kilian1, Karin U. Loeffler1, T. Hoeller2, Frank G. Holz1, Daniela Wenzel3, Martina C. Herwig1 1 Department of Ophthalmology, University of Bonn, Bonn Germany 2 Institute for Medical Biometry, Informatics and Epidemiology, University of Bonn, Bonn, Germany 3 Institute of Physiology I, University of Bonn, Bonn, Germany V Origin of intratumoral vascular structures in a murine ocular tumour model 85 Abstract Purpose: Angiogenesis represents the main pathway of tumour vascularisation. In our study, we investigated characteristics of tumour vascularisation with respect to the origin of tumour vessels and the impact of tumour associated macrophages (TAM) in a murine model of intraocular melanoma. Materials and Methods: 15 flt-1/eGFP BAC mice which express GFP in all smaller vessels, received an intravitreal melanoma cell injection. Ocular tumours were examined by ex vivo imaging (n=3) or by histology (H&E, PAS without hematoxylin) and immunohistochemistry (GFP, CD31, collagen IV, laminin, Ki67) (n=12). One group of mice was macrophagedepleted by systemic Clodronate injections prior to tumour cell inoculation (n=7). Results: Dual labelling of GFP (murine host-vessels) and CD31 (all vessels) revealed that almost all tumour vessels co-expressed GFP and CD31; very few vessels expressed only CD31. Active angiogenesis (dual labelling CD31+Ki67) was found predominantly at tumour borders adjacent to the retina and was more pronounced in untreated mice when compared to macrophage-depleted mice. Conclusion: Rapidly growing tumours are dependent on a highly pro-angiogenic microenvironment in which they are able to adopt host vessels and initiate angiogenic sprouting from incorporated vessels. TAM exhibited pro-angiogenic properties which underlines their important role in tumour progression and circulation. V Origin of intratumoral vascular structures in a murine ocular tumour model 86 Introduction Angiogenesis is a crucial point in the development of most solid tumours, including melanomas. A specific feature of uveal melanomas is their hematogenous pathway of metastasis due to the absence of lymphatic vessels in the eye. This underlines the importance of angiotropism and angiogenesis for metastatic dissemination and represents a potential target for therapeutic intervention. For enduring growth, solid tumours are dependent on constant blood supply and use different forms of vascularisation (Hanahan and Weinberg 2011). One source of initial blood supply may be angiotropic growth and the incorporation of pre-existing vessels from surrounding structures. Secondly, the tumour may induce growth of new vessels by stimulating sprouting from pre-existing host vessels, known as angiogenesis. Further, alternative forms of tumour vascularisation in aggressive tumours have been proposed such as mosaic vessels (tumourand endothelial-lined vasculature) (Chang, di Tomaso et al. 2000), vasculogenesis (recruitment of endothelial progenitor cells) (Asahara, Murohara et al. 1997), tumour-lined vascular channels (Timar and Toth 2000), and vasculogenic mimicry (Maniotis, Folberg et al. 1999; Hillen and Griffioen 2007). The latter does not represent true vessels but is composed of matrix meshworks of fluid conducting channels, which are lined by tumour cells instead of endothelial cells and which share anastomoses with “true” blood vessels (Chen, Zhang et al. 2009; Seftor, Hess et al. 2012). However, angiogenesis still represents the main pathway of perfusion in most primary tumours and is initiated by the “angiogenic switch”. It depends on a pro-angiogenic tumour microenvironment and is regulated by pro- and anti-angiogenic factors, which are produced by both malignant cells and the host cells recruited to the tumour site including tumour V Origin of intratumoral vascular structures in a murine ocular tumour model 87 infiltrating macrophages (reviewed in (Burrell and Zadeh 2012)). A pro-angiogenic microenvironment comprises several growth factors such as VEGF, as well as angiopoietins, interleukins, matrix metalloproteinases and others. Tumour associated macrophages (TAM) orchestrate the tumour microenvironment in response to secreted factors from tumour cells and other players of the micromilieu in uveal melanoma. TAM tend to specialise into a M2 phenotype with pro-angiogenic and tumour promoting properties (Mantovani, Sozzani et al. 2002). Subsequently, M2-polarised TAM are able to directly enhance tumour angiogenesis and vasculogenic mimicry as shown in our intraocular melanoma model (unpublished data, under review). In uveal melanoma and in other ocular and non-ocular tumours an increased angiogenesis along with a high mean vascular density (MVD) as well as vasculogenic mimicry were associated with an increased infiltration with TAM and with patient’s outcome (Makitie, Summanen et al. 1999; Makitie, Summanen et al. 2001). In order to better understand different strategies of intraocular tumour perfusion we studied tumour vascularisation in our intraocular melanoma model. We investigated vascular patterns and the origin of intratumoural endothelial-lined vasculature in flt-1/eGFP BAC mice which express GFP in all smaller blood vessels (Herz, Heinemann et al. 2012). Intratumoural GFPexpressing vessels might be visualised by in vivo and ex vivo imaging at different time stages. Discrimination between angiogenic and vasculogenic as well as between host- and tumourderived vessels can be performed by immunohistochemical dual-labelling. As macrophages play a pivotal role in tumour angiogenesis, we investigated their influence on different forms of vascularisation strategies and on tumour circulation patterns, in particular on active angiogenesis and on vasculogenic mimicry. V Origin of intratumoral vascular structures in a murine ocular tumour model 88 Materials and Methods Animals Breeding and housing of flt-1/eGFP BAC mice was carried out and supervised according to the Association of Research in Vision and Ophthalmology statement for the Use of Animals in Ophthalmic and Vision Research. These transgenic mice exhibit a persistent eGFP expression in small vessels which is driven by the flt-1 promoter (VEGF receptor 1, VEGFR1) (Herz, Heinemann et al. 2012). Thus, all ocular vessels originating from the host (mouse) exhibit a constant eGFP expression. 15 flt-1/eGFP BAC mice were used for the study. Three mice were investigated at the age of eight months for ex vivo imaging, 12 mice were investigated at the age of 10 months for histology and immunohistochemistry, as intraocular melanomas in such old mice better resemble human tumours with regard to histologic features of the tumour microenvironment and vascularisation. Mouse model of intraocular melanoma As previously described, 1x105 of murine cutaneous HCmel12 melanoma cells were intravitreally injected into the right eye of each mouse of all study groups at day 0 in general anaesthesia (Kilian, Loeffler et al. 2015). Mice were sacrificed at day 10 post intravitreal injection, right eyes were enucleated and processed for histologic or immunohistochemical examination. Macrophage-depleted mice (n=7) were studied in comparison to untreated mice (n=5). Liposome-encapsulated dichloromethylene diphosphonate (Cl2MDP; clodronate, ordered through clodronateliposomes.org) was used to systemically deplete blood monocytes and V Origin of intratumoral vascular structures in a murine ocular tumour model 89 macrophages at a dosage of 0.1mg/g body weight (van Rooijen and van Kesteren-Hendrikx 2003). Seven mice received repeated intraperitoneal injections of Clodronate liposomes at days -6, -3, 0 and 3 and 6 of study. To evaluate actual macrophage depletion, liver sections of Clodronate-treated and untreated mice were stained immunohistochemically with a F4/80 antibody (F4/80 Antibody, Cl:A3-1, AbD Serotec, Oxford, U.K.; dilution 1:100). Ex vivo imaging In three mice, tumour-bearing and contralateral control eyes were enucleated immediately after euthanasia. Eyes were kept in PBS and shortly after investigated using an Axio Zoom microscope (Carl Zeiss Microscopy GmbH, Jena, Germany). Specimens were examined for GFP-positive vessels in areas of extraocular tumour extension and after removing the anteriour parts of the eye for GFP-positive vessels in the intraocular tumour. Histology and immunohistochemistry Processing of histologic specimens of 12 mice was carried out as described before (Kilian, Loeffler et al. 2015). Briefly, in hematoxylin eosin (H&E) stains, tumour size was measured in mm² at 40x magnification using ImageJ 1.45s (W Rasband, NIH, USA); vascular characteristics like angiotropism, co-option of retinal vessels and intratumoural endothelial vessels were evaluated at 200x magnification. PAS (periodic acid-Schiff reaction) stains without hematoxylin were investigated for ECM-rich channels/VM along with their prognostic significant extravascular matrix patterns (parallels with cross-linkings, loops, networks). For analysis of the MVD, sections were stained with a GFP- and CD31-antibody (GFP Rabbit IgG, Life Technologies, Carlsbad, CA, USA; dilution 1:100; CD31, SZ31, Dianova, Hamburg, Germany, dilution 1:10). Under x200 power, counts of positive staining V Origin of intratumoral vascular structures in a murine ocular tumour model 90 vessels inside a 0.25mm2 graticule were performed in 1-3 areas according to introduced standards (Foss, Alexander et al. 1996). Staining of ECM was performed with a collagen IV antibody (Collagen IV Antibody, Novus Biologicals, Littleton, CO, USA dilution 1:100) and a laminin antibody (Laminin Antibody, Novus Biologicals, Littleton, CO, USA, dilution 1:100). Dual labelling of CD31 + GFP, CD31 + Collagen IV, and CD31 + Ki67 (Anti-Ki67, SP6, Abcam, Cambridge, UK, dilution 1:50) was performed and investigated by immunofluorescence microscopy. Dual labelling of CD31 and GFP allows for determination of the origin of endothelial-lined vessels. Vessels, which are positive for CD31 and simultaneously for GFP are of host-origin (mouse). On the other hand, exclusively CD31positive vessels are derived from other origins e.g. vasculogenesis of from the tumour itself. Dual labelling for CD31 and the proliferation marker Ki67 is supposed to reveal active angiogenesis. Statistical analysis Statistical analysis was performed with IBM SPSS Statistics 22.0 (IBM Corp, Armonk, NY, USA). Tumour sizes in mm2 and MVD of different study groups were evaluated by an analysis of variance and a Post Hoc Test. Chi2 test and Pearson's correlation coefficient were used to determine the relationship between different study groups and tumour growth characteristics. A p≤0.05 was considered as statistically significant and significance level was indicated by using * for p ≤ 0.05. V Origin of intratumoral vascular structures in a murine ocular tumour model 91 Results Ex vivo imaging At investigation by the Axio Zoom microscope at x25 magnification, networks of GFPexpressing small vessels were identified on the ocular surface and in the anterior chamber. In detail, a compact network was found along the limbus which was drained by a major vein. Fine vessels were observed within the pigmented iris as well as in adherent orbital structures such as the conjunctiva and muscle parts (Fig.1A). Two of three investigated mouse eyes exhibited extraocular tumour extension which presented as dark masses near the ocular limbus (Fig.1B). The location of the extraocular tumour apparently represented the site of previous intravitreal injection as the tumour probably took the path of least resistance for outgrowth. GFP-positive vessels were surrounding the extraocular tumour basis but no obvious vessels were found on the tumour surface at this early stage of tumour growth (day 10). Figure 1.: Eyes of flt-1/eGFP BAC mice viewed by Axio Zoom microscope (x25 magnification) P: pupil I: iris V: vein, T: extraocular tumour part (A) Eye of control mouse; (B) Eye of tumour inoculated mouse with extraocular tumour extension V Origin of intratumoral vascular structures in a murine ocular tumour model 92 Histology and immunohistochemistry All mice which received an intravitreal HCmel12 tumour cell injection developed solid intraocular tumours, which gained access to the host’s blood circulation. In H&E stains, these tumours presented with angiotropic growth, co-option of retinal or choroidal vessels, networks of endothelial vessels as well as of vasculogenic mimicry. Dual-labelling of CD31 and GFP revealed that the majority of tumour vessels were positive for both markers and were thus of mouse origin (Fig.2A+B). Endothelial vessels labelled exclusively by the CD31-antibody were very sparse and represented only small vessels with a narrow lumen. A vessel sequence representing mosaic vessels (CD31+GFP+ vessels in linear arrangement with CD31+GFP- vessels) was not unequivocally observed in any of the tumours. A B Figure 2: Vascular structures in intraocular tumours ((A) x100 and (B) x200 magnification) (A+B) Dual labelling for host vessels’ GFP (green) and CD31 (red) revealed most intratumoural vessels being positive for both markers and thus of angiogenic host origin. Only few vessels were exclusively CD31 positive (arrows). V Origin of intratumoral vascular structures in a murine ocular tumour model 93 Active proliferation of endothelial vessels was regarded as a positive nuclear Ki67 staining in a CD31 positive endothelial cell (cytoplasm). Such a dual-labelling was found predominantly at tumour margins which were adjacent to host structures. This suggests active sprouting of blood vessels from surrounding ocular structures (Fig.3A+B). Intraperitoneal injection of Clodronate liposomes led to systemic macrophage depletion as examined in immunohistochemical stains of the liver. While untreated mice exhibited frequent F4/80-posititive staining of Kupffer cells throughout the liver, Clodronate liposometreated mice were lacking positively staining cells. In untreated mice 39.1% of endothelial vessels showed proliferative activity compared to 30.9% of proliferating vessels in macrophage-depleted mice (p=0.114). A B Figure 3: Active angiogenesis in intraocular tumours ( (A)x100 and (B) x200 magnification) (A+B) Dual labelling for Ki67 (green) and CD31 (red) revealed active angiogenesis (arrows) which was predominantly found at tumour margins adjacent to host structures. Collagen IV expression revealed no statistical significant difference between the two study groups (p=0.364). However, staining for collagen IV suggested a slightly pronounced expression in untreated mice compared to macrophage-depleted mice. Collagen IV-positive V Origin of intratumoral vascular structures in a murine ocular tumour model 94 structures formed arcing and looping patterns, exhibited a patchy distribution and they were mainly found in close vicinity to endothelial vessels. Such tumour areas with increased tissue remodelling were also positive for laminin and were located predominantly at tumour margins (Fig.4A). Intraocular tumour size did not differ statistically between both study groups (mean size: untreated mice 0.7mm2, macrophage-depleted mice 0.73mm2, p=0.752). MVD did not differ statistically between untreated and macrophage-depleted mice (p=0.513). In PAS stains without hematoxylin, ECM-rich structures of untreated mice exhibited more frequently prognostic extravascular matrix patterns. However, this effect was statistically not significant (p=0.197) in contrast to another experiment with an analogous tumour model. This might be attributed to the minor number of animals in this study. A B Figure 4: Extracellular matrix (ECM) in intraocular tumours ((A) x100 and (B) x200 magnification) (A+B) Dual labelling for the ECM component collagen IV (green) and CD31 (red) showed abundant ECM structures adjacent to endothelial vessels and of arcing and looping patterns. V Origin of intratumoral vascular structures in a murine ocular tumour model 95 Discussion In many tumours different strategies of vascularisation and correlation to tumour progression have been well described (reviewed in (Hillen and Griffioen 2007)). Generally, angiogenesis is thought to be the main pathway of tumour vascularisation. The essential role of angiogenesis in solid tumours and its potential therapeutic implications were first described by Folkman and coworkers (Folkman 1971). However, the long-held belief that cancer patients can be effectively treated with anti-angiogenic therapy diminished after disappointing clinical results. Heterogeneity and complexity of tumours and of their vascular architecture may be a reason for that (Verheul, Voest et al. 2004). Co-option of host vessels, mosaic vessels, vasculogenesis and VM might be of more importance than previously thought. In uveal, cutaneous and oral melanoma, numerous studies have focused on angiogenesis and other mechanisms by which melanomas ensure their vascularisation. Besides sprouting angiogenesis, also patterns of co-option of host vessels, mosaic vessels, tumour-lined vasculature, and networks of vessel-like structures not lined by endothelial cells (vasculogenic mimicry) have been described (reviewed in (Pastushenko, Vermeulen et al. 2014), (Hendrix, Seftor et al. 2002; Lee, Nagai et al. 2002; Zhang, Zhang et al. 2006). These studies used PAS and immunohistochemical staining of endothelial vessels, extracellular matrix (ECM) and other structures (e.g. CD31, collagen IV) to visualise and investigate the characteristics of melanoma vascularisation. In our study, we were able to further address the origin of tumour vessels in an intraocular melanoma model with immunocompetent mice. V Origin of intratumoral vascular structures in a murine ocular tumour model 96 Origin of tumour vessels Almost all tumour vessels in flt-1/eGFP BAC mice exhibited a co-expression of GFP and CD31 and were thus of angiogenic host origin. Apparently, the tumour adopted host vessels including their capacity for angiogenesis and benefited from a pro-angiogenic microenvironment. Thus, angiogenesis represented the main vascularisation strategy of the tumour. However, a minor number of intratumoural vessels were found to be negative for GFP and solely marked by the CD31 antibody and must, subsequently, be of other origin than sprouting angiogenesis from host vessels. These vessels might have risen from two other potential origins: from bone marrow derived endothelial progenitor cells in terms of vasculogenesis or from tumour-derived endothelial vessels. Circulating endothelial precursors, shed from the vessel wall or mobilised from the bone marrow, can also contribute to tumour vascularisation (vasculogenesis) (Rafii 2000). Such endothelial precursor cells may line intratumoural vessels and express characteristic markers (CD31, CD34, VEGFR-2) along with the stem cell marker CD133 (Timmermans, Plum et al. 2009). To identify such vasculogenic vessels formed by endothelial progenitor cells we (intend to) perform dual labelling of CD31 and CD133. Shall we be able to confirm that these vessels have originated from vasculogenesis by being built of endothelial progenitor cells (CD31+CD133+GFP-), this would proof heterogeneity of tumour vascularisation in our model. Further, flt-1/eGFP BAC mice would represent a potent model to distinguish between vasculogenesis (CD31+CD133+GFP- vessels) and angiogenesis (CD31+CD133+GFP+ vessels). This might offer to study these different forms of vascularisation in several tumour entities and other pathologies like injury or inflammatory processes. On the other hand, if CD31+GFP- vessels in flt-1/eGFP BAC mice reveal not to have originated from vasculogenesis (CD133-), another hypothesis of their origin might be a V Origin of intratumoral vascular structures in a murine ocular tumour model 97 derivation from cancer stem cells. This hypothesis might further be supported by dual labelling of CD31 and a melanoma marker (e.g. HMB45/MART-1). Two studies in glioblastoma found that blood vessel cells within the tumours contained genetic markers characteristic of cancer cells (with stem cell like properties), suggesting that these blood vessels were of tumour origin (Ricci-Vitiani, Pallini et al. 2010; Wang, Chadalavada et al. 2010). Another group describes „tumour endothelial cells (TEC)“ as intratumoural vessels of tumour origin with highly angiogenic phenotypes and higher proliferative and migratory abilities than „normal endothelial cells“ (=host-derived, angiogenic endothelial cells) (Ohmura-Kakutani, Akiyama et al. 2014). However, these conclusions were based on in vitro studies and their transferability into in vivo conditions remains unanswered. In an ischemic, nude mouse model with inoculated human aggressive melanoma cells, however, newly formed vasculature within the tumour showed endothelial cells of human and of mouse origin in a linear arrangement. Thus, besides angiogenic murine vessels, human melanoma cells may acquire endothelial-like morphology and are incorporated in capillaries, and thus, may actively contribute to neovascularisation (Mihic-Probst, Ikenberg et al. 2012). Our ongoing studies may reveal the origin of CD31+GFP- intratumoural vessels in flt-1/eGFP BAC mice and either offer a new model for distinguishing vasculogenesis from angiogenesis or contribute to recent discussions regarding tumour-derived endothelial vessels. This would represent the first orthotopic animal model of such tumour-derived endothelial cells in melanoma. Intratumoural vascularisation and TAM In our study, angiogenesis represented the primary mechanism of tumour vascularisation. However, other vascularisation strategies were also observed, like co-option of host vessels V Origin of intratumoral vascular structures in a murine ocular tumour model 98 and VM. Studies in murine and human cutaneous melanoma revealed that vascularisation strategies such as VM and mosaic vessels were not uniformly distributed throughout the tumour but appeared in patches. Blood supply was sustained simultaneously by VM and endothelial lined vasculature (Zhang, Zhang et al. 2006). In our model, true mosaic vessels which are lined by host endothelial cells (CD31+GFP+) with a transient change to tumour endothelial cells (HMB45/MART-1+CD31+GFP-) were not yet unequivocally observed. However, in accordance with the study by Zhang (vide supra), we observed a patchy distribution of VM and collagen IV positive ECM structures. This underlines how the intraocular tumour ensures constant blood supply by diverse strategies in our model. In order to effectively inhibit tumour vascularisation – as a potential therapeutic interventionall different strategies of vascularisation should be addressed. One point of action might be manipulation of the pro-angiogenic tumour microenvironment which in turn is mainly orchestrated by TAM (in case of ocular melanoma and many other tumours). Manipulating TAM and thus altering the tumour microenvironment might be a promising approach to control tumour progression and vascularisation (Bronkhorst and Jager 2013). In our study, the impact of macrophages on different tumour vascularisation strategies was determined to a minor extend. We examined intraocular tumours of macrophage-depleted mice in comparison to untreated mice. Dual labelling of CD31 and Ki67 (proliferation marker) revealed active angiogenesis which was predominantly found at tumour margins located near host structures. This is in accordance with a study which found high numbers of proliferating endothelial cells at tumour margins of poor differentiated (and thus aggressive) tumours (Raluca, Cimpean et al. 2015). In these regions the tumour might have orchestrated the microenvironment towards pro-angiogenic conditions in order to induce sprouting angiogenesis from surrounding host structures. Studies investigating MVD and the proportion V Origin of intratumoral vascular structures in a murine ocular tumour model 99 of active angiogenesis in cutaneous melanoma and other tumour entities showed that the number of proliferating endothelial cells (CD31+/CD34+/Nestin+ and Ki67+) but not MVD was correlated with tumour stage (Baeten, Castermans et al. 2006; Hillen, van de Winkel et al. 2006; Kruger, Stefansson et al. 2013). Thus, the proportion of proliferating endothelial cells might be also an indicator for tumour progression and possibly also for patient’s outcome. In our study, in untreated mice 39.1% of endothelial vessels showed proliferative activity compared to 30.9% of proliferating vessels in macrophage depleted mice (p=0.114). As tumours in untreated mice showed a tendency of an increased rate of active angiogenesis in comparison to macrophage-depleted mice, TAM might have directly shaped the tumour microenvironment by secreting pro-angiogenic factors – probably in response to a crosstalk with melanoma cells due to hypoxic conditions. TAM are generally able to stimulate proliferation and migration of endothelial cells (reviewed in (Lamagna, Aurrand-Lions et al. 2006)) and may thereby influence the rate of active angiogenesis in aggressive tumours. This points to another pathway by which TAM directly affect prognostically significant histopathologic tumour characteristics. Further, untreated mice more frequently showed prognostically significant extravascular matrix patterns than macrophage-depleted mice. This underlines the pro-angiogenic properties of TAM and their important role in tumour progression and vascularisation. In another study, we were able to better retrace the role of macrophages in intraocular tumour biology by using an increased number of animals (unpublished, under review). Thus, understanding the role of TAM in tumour vascularisation, controlling or re-polarising TAM into tumouricidal phenotypes might offer new strategies to manipulate the tumour microenvironment and thus control tumour progression (Catalano, Turdo et al. 2013). V Origin of intratumoral vascular structures in a murine ocular tumour model 100 Summary Intraocular tumours exhibited different forms of vascularisation including angiogenesis and vasculogenic mimicry and potentionally also vasculogenesis or tumour-derived endothelial vessels. Angiogenesis represented the main vascularisation strategy, whereas only very few vessels derived from other origins. Active angiogenesis was found throughout the tumour but mainly at tumour borders and was pronounced in untreated mice when compared to macrophage-depleted mice. TAM had an impact on the frequency of extravascular matrix pattern and on active angiogenesis. Targeting the tumour microenvironment which is orchestrated by pro-angiogenic TAM might offer new therapeutic targets to control tumour vascularisation, progression and metastasis. Acknowledgements We gratefully acknowledge research funding from BONFOR (University of Bonn) and the “Verein der Rheinisch-Westfälischen Augenärzte (RWA)”. We would like to thank Thomas Tüting (Laboratory of Experimental Dermatology, University of Bonn, Bonn, Germany) for kindly providing the HCmel12 murine melanoma cell line and Katia Herz (Institute of Physiology I, University of Bonn, Bonn, Germany) for the assistance ex vivo imaging. Especially, we want to thank Parand Widmar and Claudine Strack (Department of Ophthalmology, University of Bonn, Bonn, Germany) for their assistance in histology. V Origin of intratumoral vascular structures in a murine ocular tumour model 101 References 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011;144(5):646-74. Chang YS, di Tomaso E, McDonald DM, Jones R, Jain RK, Munn LL. Mosaic blood vessels in tumors: frequency of cancer cells in contact with flowing blood. Proc Natl Acad Sci U S A 2000;97(26):14608-13. Timar J, Toth J. Tumor sinuses- vascular channels. Pathol Oncol Res 2000;6(2):83-6. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997;275(5302):964-7. Maniotis AJ, Folberg R, Hess A, Seftor EA, Gardner LM, Pe'er J, et al. Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol 1999;155(3):739-52. Hillen F, Griffioen AW. Tumour vascularization: sprouting angiogenesis and beyond. Cancer Metastasis Rev 2007;26(3-4):489-502. Seftor RE, Hess AR, Seftor EA, Kirschmann DA, Hardy KM, Margaryan NV, et al. Tumor cell vasculogenic mimicry: from controversy to therapeutic promise. Am J Pathol 2012;181(4):1115-25. Chen L, Zhang S, Li X, Sun B, Zhao X, Zhang D, et al. A pilot study of vasculogenic mimicry immunohistochemical expression in intraocular melanoma model. Oncol Rep 2009;21(4):989-94. Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol 2002;23(11):549-55. Makitie T, Summanen P, Tarkkanen A, Kivela T. Microvascular density in predicting survival of patients with choroidal and ciliary body melanoma. Invest Ophthalmol Vis Sci 1999;40(11):2471-80. Makitie T, Summanen P, Tarkkanen A, Kivela T. Tumor-infiltrating macrophages (CD68(+) cells) and prognosis in malignant uveal melanoma. Invest Ophthalmol Vis Sci 2001;42(7):1414-21. Herz K, Heinemann JC, Hesse M, Ottersbach A, Geisen C, Fuegemann CJ, et al. Live monitoring of small vessels during development and disease using the flt-1 promoter element. Basic Res Cardiol 2012;107(2):257. Kilian MM, Loeffler KU, Pfarrer C, Holz FG, Kurts C, Herwig MC. Intravitreally Injected HCmel12 Melanoma Cells Serve as a Mouse Model of Tumor Biology of Intraocular Melanoma. Curr Eye Res 2015:1-8. van Rooijen N, van Kesteren-Hendrikx E. "In vivo" depletion of macrophages by liposome-mediated "suicide". Methods Enzymol 2003;373:3-16. Foss AJ, Alexander RA, Jefferies LW, Hungerford JL, Harris AL, Lightman S. Microvessel count predicts survival in uveal melanoma. Cancer Res 1996;56(13):2900-3. Pastushenko I, Vermeulen PB, Van den Eynden GG, Rutten A, Carapeto FJ, Dirix LY, et al. Mechanisms of tumour vascularization in cutaneous malignant melanoma: clinical implications. Br J Dermatol 2014;171(2):220-33. V Origin of intratumoral vascular structures in a murine ocular tumour model 17. 18. 19. 20. 21. 22. 23. 24. 102 Lee YJ, Nagai N, Siar CH, Nakano K, Nagatsuka H, Tsujigiwa H, et al. Angioarchitecture of primary oral malignant melanomas. J Histochem Cytochem 2002;50(11):1555-62. Zhang S, Zhang D, Wang Y, Zhao W, Guo H, Zhao X, et al. Morphologic research of microcirculation patterns in human and animal melanoma. Med Oncol 2006;23(3):403-9. Hendrix MJ, Seftor RE, Seftor EA, Gruman LM, Lee LM, Nickoloff BJ, et al. Transendothelial function of human metastatic melanoma cells: role of the microenvironment in cell-fate determination. Cancer Res 2002;62(3):665-8. Mihic-Probst D, Ikenberg K, Tinguely M, Schraml P, Behnke S, Seifert B, et al. Tumor cell plasticity and angiogenesis in human melanomas. PLoS One 2012;7(3):e33571. Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med 1971;285(21):1182-6. Verheul HM, Voest EE, Schlingemann RO. Are tumours angiogenesis-dependent? J Pathol 2004;202(1):5-13. Bronkhorst IH, Jager MJ. Inflammation in uveal melanoma. Eye (Lond) 2013;27(2):217-23. Catalano V, Turdo A, Di Franco S, Dieli F, Todaro M, Stassi G. Tumor and its microenvironment: a synergistic interplay. Semin Cancer Biol 2013;23(6 Pt B):52232. 103 VI Overexpression of Hepatocyte Growth Factor and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma Marta M. Kilian1, Martina C. Herwig1, Frank G. Holz1, Thomas Tüting2, Karin U. Loeffler1 1 Department of Ophthalmology, University of Bonn, Bonn, Germany 2 Experimental Dermatology, University of Bonn, Bonn, Germany Letter to the Editor Accepted at Melanoma Research VI Overexpression of HGF and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma 104 Editor, Searching for an animal model of spontaneous uveal melanoma, we evaluated the ocular findings in Hgf-Cdk4R24C mice, an established mouse model of spontaneous skin melanoma. In humans and in Hgf-Cdk4R24C mice overexpression of hepatocyte growth factor (HGF) may promote cutaneous melanogenesis by activating RAS signal transduction pathways via its receptor c-MET (Landsberg, Gaffal et al. 2010). In uveal melanoma, high expression of cMET correlates with an aggressive tumour phenotype (Mallikarjuna, Pushparaj et al. 2007). Additionally, the oncogenic germline mutation in the cyclin-dependent kinase 4 (CDK4 R24C) further contributes to melanoma development in Hgf-Cdk4R24C mice and human skin because it functionally inactivates the tumour suppressor p16⁄INK4a, which is of critical importance for oncogene-induced senescence (Landsberg, Kohlmeyer et al. 2012). This study should also further clarify the role and impact of HGF and its receptor tyrosine kinase c-MET as well as cyclin-dependent kinase 4 (CDK4) on ocular melanocytes in comparison with dermal melanocytes. The majority of these highly pigmented mice spontaneously develops a spectrum of nevi and primary melanomas without prior carcinogen treatment within the first year of life (Landsberg, Gaffal et al. 2010). These melanomas are predominantly spreading to regional lymph nodes and the lungs (Tormo, Ferrer et al. 2006). As Hgf-Cdk4R24C mice represent an established model of spontaneous cutaneous melanoma, we questioned whether this mouse strain concurrently shows prolifferations and tumourigenesis in ocular melanocytic structures. 12 eyes from 11 month old Hgf-Cdk4R24C mice (C57Bl/6 background) were investigated and were compared with three eyes from 10 months old C57Bl/6 mice. Step sections were stained with H&E and PAS, and sections in-between were labelled with antibodies to Ki67 VI Overexpression of HGF and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma 105 (proliferation marker), MART1 (melanocyte/melanoma marker), CKpan (epithelial cell differentiation) and F4/80 (macrophages). Prior to immunohistochemistry (IHC), sections were bleached for better visualisation of the red chromogen AEC (3-amino-9-ethylcarbazole). At the age of 11 months all Hgf-Cdk4R24C mice exhibited apparent dermal melanocytic proliferations, partly exceeding 2 mm and were - according to (Landsberg, Gaffal et al. 2010) - therefore considered as melanomas. By gross examination of the eyes, ocular alterations were not observed (Figure 1A+B). No significant increase in epithelial pigmentation of either corneal or conjunctival epithelium was noted (Figure 1C+D). Proliferation of uveal melanocytes or cells of the retinal pigment epithelium (RPE) was also not observed and the choroid appeared even somewhat less pigmented with some amelanotic cellular areas compared to C57Bl/6 mice (n=3) (Figure 1G+H). IHC so far showed no significant difference in labelling between Hgf-Cdk4R24C and C57Bl/6 mouse eyes. However, histologically, the most marked difference between Hgf-Cdk4R24C and wildtype mice was the incidental finding of heavily pigmented cells in the anterior corneal stroma (and conjunctiva to a lesser degree) and along the chamber angle (Figure 1C-F). This was associated with inflammation, vascularisation and a marked proliferation of these pigmented cells in three of twelve eyes (Figure 1C). Histological and immunohistochemical staining was unable to further characterise the cellular corneal infiltrate. In Hgf-Cdk4R24C mice of 11 months of age, despite of apparent dermal melanocytic proliferations progressing toward cutaneous melanomas, no major difference in pigmentation or cellular components was observed in the ocular epithelium or in the posterior segment. Thus, Hgf-Cdk4R24C mice up to this age do not qualify as a model for spontaneous conjunctival or uveal melanoma. A delayed melanomagenesis at an even more advanced age may not completely be ruled out. Apparently, genetic alterations regarding Hgf and Cdk4 and VI Overexpression of HGF and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma 106 the subsequent deregulation of the c-MET mediated RAS signal transduction pathways as well as the p16/Ink4a-dependent cell cycle regulation apparently lead to cutaneous primary and metastatic melanomas. However, uveal or conjunctival cell cycles do not seem to be profoundly affected. To date, several characteristic genetic anomalies for tumour progression and metastasis of cutaneous as well as uveal melanoma have been identified (van den Bosch, Kilic et al. 2010). But unlike in cutaneous melanoma, B-Raf, Ras or Kit mutations occur rarely in uveal melanoma and characteristic mutations differ between uveal and cutaneous melanoma (van den Bosch, Kilic et al. 2010). Although mutations similar to skin melanoma such as B-RAF V600 have been found in conjunctival melanomas (Lake, Jmor et al. 2011), our findings underline genetic differences also between conjunctival and skin melanomas. However, since remarkable changes regarding pigmentation were noted in the cornea and chamber angle in these animals, further investigation of the anterior segment might allow studying mechanisms of corneal neovascularisation and possibly also glaucoma. VI Overexpression of HGF and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma 107 Figure 1: Hgf-Cdk4R24C (A,C,E,G) and C57Bl/6 (B,D,F,H) mouse eyes, H&E; arrows indicating corneal intrastromal pigment. (A+B) Overview of mouse eyes; (C+D) Cornea: (C) cornea with a deeply pigmented cellular lesion (stars) within the anterior stroma and diffuse neovascularisation (arrowheads) and (D) cornea without evidence of corneal pigment; (E+F) Chamber angle: (E) chamber angle with increased pigmentation of the trabecular meshwork and cornea with intrastromal pigment and (F) unremarkable chamber angle; (G+H) Unremarkable choroid and retina of the HgfCdk4R24C mouse and C57Bl/6 mouse. VI Overexpression of HGF and an oncogenic CDK4 variant in mice alters corneal stroma morphology but does not lead to spontaneous ocular melanoma 108 References 1. 2. 3. 4. 5. 6. Landsberg J, Gaffal E, Cron M, Kohlmeyer J, Renn M, Tuting T: Autochthonous primary and metastatic melanomas in Hgf-Cdk4 R24C mice evade T-cell-mediated immune surveillance. Pigment Cell Melanoma Res 2010;23:649-660. Mallikarjuna K, Pushparaj V, Biswas J, Krishnakumar S: Expression of epidermal growth factor receptor, ezrin, hepatocyte growth factor, and c-Met in uveal melanoma: an immunohistochemical study. Curr Eye Res 2007;32:281-290. Landsberg J, Kohlmeyer J, Renn M, Bald T, Rogava M, Cron M, Fatho M, Lennerz V, Wolfel T, Holzel M, Tuting T: Melanomas resist T-cell therapy through inflammationinduced reversible dedifferentiation. Nature 2012;490:412-416. Tormo D, Ferrer A, Gaffal E, Wenzel J, Basner-Tschakarjan E, Steitz J, Heukamp LC, Gutgemann I, Buettner R, Malumbres M, Barbacid M, Merlino G, Tuting T: Rapid growth of invasive metastatic melanoma in carcinogen-treated hepatocyte growth factor/scatter factor-transgenic mice carrying an oncogenic CDK4 mutation. Am J Pathol 2006;169:665-672. van den Bosch T, Kilic E, Paridaens D, de Klein A: Genetics of uveal melanoma and cutaneous melanoma: two of a kind? Dermatol Res Pract 2010;2010:360136. Lake SL, Jmor F, Dopierala J, Taktak AF, Coupland SE, Damato BE: Multiplex ligation-dependent probe amplification of conjunctival melanoma reveals common BRAF V600E gene mutation and gene copy number changes. Invest Ophthalmol Vis Sci 2011;52:5598-5604. 109 VII Discussion Animal models Experimental animal models are valuable tools for a better understanding of the pathophysiology of human disease and for identifying novel therapeutic targets. Based on genetic alterations found in human tumours, multiple models have been generated in genetically modified or drug-induced animals. As no mouse model of spontaneous and metastasising uveal melanoma has been described yet, there is an unmet need for such a model for basic and pre-clinical research. Many groups examined ocular findings of transgenic mice which mostly represented models of spontaneous skin melanoma or intended to induce intraocular melanomas with radiation or other compounds (Dithmar, Albert et al. 2000). Although several studies described ocular melanocytic proliferations, most pigmented intraocular tumours however, revealed to originate from the RPE or failed to metastasise (Kramer, Powell et al. 1998; Syed, Windle et al. 1998; Albert, Kumar et al. 2004; Tolleson, Doss et al. 2005; Latendresse, Muskhelishvili et al. 2007). Recently, Schiffner and co-workers showed in another transgenic mouse breed - with spontaneous skin melanoma - that pigmented choroidal proliferations mimicked spontaneous uveal melanoma (Schiffner, Braunger et al. 2014). However, applicability as a model for studying intraocular melanomas remains questionable. In constant search for a potential mouse model, we evaluated ocular findings in Hgf-Cdk4R24C mice. These represent a mouse model of spontaneous and metastasising skin melanoma and their eyes have not yet been examined for ocular melanomas. (Landsberg, Gaffal et al. 2010). Apart from incidental findings of unusual heavily pigmented cells in the cornea and conjunctiva, however, no evidence for melanocytic VII Discussion 110 proliferations in the uvea was found (Kilian, Loeffler et al. 2015, in print). These negative findings underline genetic differences between uveal, conjunctival and cutaneous melanoma which result in different pathways of tumourigenesis, pathology and potential therapeutic targets of these tumours (van den Bosch, Kilic et al. 2010) (chapter VI). To date, all existing mouse models of uveal melanoma exhibit limitations and one need to carefully select a model which best qualifies for the specific research questions. To investigate intraocular tumour biology the best compromise appears to inject intravitreally cutaneous melanoma cells in order to induce an intraocular melanoma. Despite genetic differences between cutaneous and uveal melanoma, such a model allows investigations on intraocular tumour characteristics and immunology in an immunocompetent animal. In our study on the role of tumour associated macrophages (TAM) in intraocular melanoma we modified an established murine model. Following the intravitreal injection of murine cutaneous HCmel12 melanoma cells into the eyes of CX3CR1+/GFP mice, intraocular tumours showed solid tumour growth, angiotropism, angiogenesis, vasculogenic mimicry with extravascular matrix patterns as well as inflammatory cell infiltration including macrophages. In addition, metastases were observed in regional lymph nodes and lungs. However, metastasis occurred only in mice which previously exhibited extraocular tumour extension. Thereby, these tumours might have gained access to the conjunctival lymphatics and metastasised via the lymphatic system. Haematogenously dispersed metastases of tumours strictly confined to the eye were not observed. Thus, this mouse model does not qualify as a model for metastasising intraocular melanoma. However, although being of cutaneous origin intraocular tumours of HCmel12 melanoma cells showed apparent similarities to human uveal melanoma such as solid tumour growth, angiogenesis, macrophage infiltration and other (vide supra). These characteristics appear to represent general features of tumour biology since they VII Discussion 111 can be observed in many solid tumour entities. Thus, tumours of intravitreally injected HCmel12 melanoma cells qualify as a model for intraocular melanoma and allow studies on questions related to the tumour’s microenvironment, immunologic aspects as well as angiogenesis and other vascularisation strategies (chapter III). Tumour associated macrophages in intraocular melanoma The eye represents a semi-closed system with access to blood vessels but no lymphatic vessels. Thereby, the intraocularly injected tumour cells may grow inside an immune privileged compartment and may develop a microenvironment which can be manipulated iatrogenically by systemic or intraocular injections. In uveal melanoma and other tumours, TAM represent a major player within the tumour microenvironment and may contribute to tumour progression (Allavena, Sica et al. 2008; Mantovani and Sica 2010). As we were interested in the role of macrophages in intraocular melanoma, we applied our described intraocular melanoma model to mice with systemically depleted macrophages in comparison to untreated mice of two different ages (young 8-12 weeks; old 10-12 months). By repeated intraperitoneal injections of liposome-encapsulated clodronate macrophages were effectively depleted. Systemic depletion of macrophages can result in tumour growth inhibition and reduced angiogenesis in cutaneous melanoma (Gazzaniga, Bravo et al. 2007). In our animal model, TAM did not have a statistical significant impact on tumour size. However, in vitro results showed increased cell proliferation rates in melanoma cells which were grown in an environment (supernatant) shaped by unpolarised macrophages (chapter IV). In vivo, advanced age and the presence of macrophages were individually associated with an increased occurrence of prognostically significant extravascular matrix patterns (untreated young vs. untreated old mice: p=0.024; untreated vs. macrophage depleted mice: p=0.014; VII Discussion 112 chapter IV). This patterned microcirculation characteristic of vasculogenic mimicry was first described in uveal melanoma (Folberg, Pe'er et al. 1992; Folberg, Rummelt et al. 1993; Maniotis, Folberg et al. 1999; Folberg, Hendrix et al. 2000). These structures are part of the extracellular matrix (ECM), share anastomoses with “true” blood vessels and may contribute to perfusion in aggressive tumours (Maniotis, Folberg et al. 1999; Chen, Zhang et al. 2009; Seftor, Hess et al. 2012). Aggressive melanomas which need to gain access to oxygen and nutrients are able to actively remodel the ECM in order to arrange stromal patterns such as vascular networks (Daniels, Boldt et al. 1996). Besides tumour cells themselves, TAM are also potentially able to enhance tissue remodelling and extravascular matrix pattern formation (Seftor, Seftor et al. 2001). Thus, the frequency of prognostically significant extravascular matrix pattern in aged and untreated mice might be directly related to the crosstalk between melanoma cells and TAM. Impact of macrophage polarisation TAM obtain their functional polarisation phenotype in response to stimuli present in the tumour microenvironment. Polarised macrophages differ in terms of receptor expression, effector function as well as cytokine and chemokine production (Mantovani, Sozzani et al. 2002). In many malignant tumours - including uveal melanoma - infiltrating TAM exhibit predominantly a M2 polarisation phenotype (Mantovani, Sozzani et al. 2002; Pierard, PierardFranchimont et al. 2012). Presence and a higher number of such a M2 phenotype are associated with tumour favouring properties and are statistically correlated to poor prognosis (Herwig, Bergstrom et al. 2013). Our results from the polarisation experiment (chapter IV) display tumour characteristics under iatrogenically induced tumour microenvironments of extreme M1- or M2 dominance. This VII Discussion 113 experimental design does not model actual conditions in intraocular tumours but demonstrates in which ways polarised macrophages may influence tumour characteristics (and thus prognosis). In vitro as well as in vivo, our results indicate a far more aggressive tumour phenotype in M2-conditioned tumours. Intraocular tumours of HCmel12 melanoma cells which were previously incubated in M2-dominated conditions - showed an increased infiltration of inflammatory cells (lymphocytes and macrophages) compared to M1conditioned tumours (p=0.001) (chapter IV). In uveal melanoma, a high infiltration with lymphocytes and M2 macrophages is statistically correlated with negative prognostic factors such as monosomy 3 and thus with a poor prognosis (Bronkhorst, Vu et al. 2012). Generally, M2-polarised TAM are regarded as a paradigm for cancer promoting inflammation. They may orchestrate tumour growth as well as matrix deposition and stroma remodelling in order to construct a metastatic niche (Sica 2010). Accordingly, our M2-conditioned tumours exhibited higher levels of ECM collagen IV positive structures which resulted in an increased tissue remodelling compared to M1-conditioned tumours (p=0.024). This effect of increased levels of collagen IV positive structures was also suggestively observed in untreated mice when compared to systemically macrophage depleted mice (in CX3CR1+/GFP mice chapter IV, in flt1/eGFP BAC mice chapter V), but did not reach statistical significance. In cutaneous melanoma, collagen IV was found to influence tumour cell mobility and thus contributes to tumour progression and may possibly represent a prognostic indicator (Pierard, PierardFranchimont et al. 2012). In our model with M2-conditioned tumours, we were able to directly address increased tissue remodelling rates to a M2-dominated microenvironment. In addition, MVD was also significantly increased in M2-conditioned tumours compared to M1conditioned tumours (p=0.01). In uveal melanoma and other solid tumours, MVD represents a well-known histopathological prognostic indicator and is further statistically correlated with VII Discussion 114 tumour size, number of infiltrating TAM and finally with metastasis (Folberg, Mehaffey et al. 1997; Makitie, Summanen et al. 1999). However, results from the depletion experiment in CX3CR1+/GFP mice (chapter IV) and flt-1/eGFP BAC mice (chapter V) did not show significant differences in MVD between macrophage-depleted and untreated mice. This might be explained by either an insufficient majority of M2 macrophages over M1 macrophages in untreated mice or by a short time (nine days) of tumour growth in our model. Thus, a M2associated pro-angiogenic influence on the MVD could not emerge. However, in iatrogenically M2-conditioned tumours such a pro-angiogenic effect on the MVD could be demonstrated (polarisation experiment chapter IV). This underlines functionally statistical data of previously reported correlations between the level of TAM infiltration and histopathological prognostic factors such as extravascular matrix patterns or MVD. Thus in our model, pro-angiogenic and other tumour-promoting factors could be attributed directly and functionally to a tumour microenvironment dominated by M2 macrophages. Impact of advanced age Advanced age is associated with defects in both the innate and the adaptive arm of the immune system. The complex process of immunosenescence affects numbers, function, and early stages of immune cell activation (Gomez, Boehmer et al. 2005; Gomez, Nomellini et al. 2008). A dysfunctional innate immune system represents a contributing factor for patients’ outcomes after injury, inflammation or neoplastic disease in the elderly. Aging may have different effects on macrophages (as a part of the innate immune system) depending on the type of tissue (Stout and Suttles 2005; Gomez, Nomellini et al. 2008). In tumours of aged mice, TAM exhibit more often a M2 polarisation, since macrophages from geriatric mice are particularly sensitive to M2 phenotype promoting signals (Jackaman, Radley-Crabb et al. VII Discussion 115 2013). To address and evaluate age-dependent variations and changes in tumour growth characteristics and macrophage abundance/polarisation we conducted our studies in two different age groups (young 8-12 weeks; old 10-12 months). We identified differences in histopathologic features as well as in macrophage polarisation (chapter IV). In our model, TAM of old mice showed increased expression levels of a M2 marker (CD163), as shown in flow cytometry analysis. Particularly, tumour infiltrating Gr-1 positive macrophages - which are derived from the bone marrow, enter the tumour as circulating monocytes and then proliferate and polarise into macrophages - showed an increased rate of the M2 marker when compared to their younger counterparts (p=0.08). This effect was slightly below statistical significance. However, as macrophage polarisation is a continuum and other studies are in accordance with our observation, the tumour microenvironment in old mice appears to be M2dominated (generally as well as in our experimental design). Such a M2-dominated tumour microenvironment features a pro-angiogenic basis for the tumour to induce angiogenesis or other vascularisation strategies. In physiologic and neoplastic conditions, angiogenesis is altered and delayed in aging due to different impairments in pathways regarding matrix composition, growth factor expression and inflammatory response (Yamaura and Matsuzawa 1980; Cohen 1994; Sadoun and Reed 2003). In our experiments, we could not retrace an impact of age on the MVD comparing young and old animals (p=0.542). However, since mice were terminated at day nine post intraocular tumour cell injection, differences in MVD might have not reached statistical significance in this early tumour stages (vide supra). Alternative vascularisation strategies - such as vasculogenic mimicry - gain importance for tumours in aged mice (and presumably also in humans). In our macrophage-depletion and macrophage-polarisation studies, advanced age was individually associated with more frequent extravascular matrix patterns (p=0.024) and a tendency of increased collagen IV VII Discussion 116 positive ECM levels (p=0.074). Further, an increased appearance of M2 macrophages was found in aged mice in comparison to young mice (flow cytometry analysis, p=0.08, chapter IV). This is in accordance with the situation in human uveal and cutaneous melanoma where age, extravascular matrix patterns, increased collagen IV levels and an increased amount of M2 macrophages are correlated statistically with a worse prognosis (Folberg, Mehaffey et al. 1997; Makitie, Summanen et al. 2001; Pierard, Pierard-Franchimont et al. 2012). In our study, these tumour characteristics in old mice are associated with a more aggressive tumour phenotype and better resemble human tumours. However, most described mouse models of tumour biology are still conducted in young mice. A study design comprising old mice is costly in terms of time and expenses. However, considering our results, we concluded that old mice represent a better tumour model than young mice and should be used for studies on the tumour microenvironment, vascularisation or tumour associated macrophages. Tumour associated macrophages and angiogenesis To investigate the role of macrophages in tumour angiogenesis we applied the same experimental design (macrophage-depleted mice in comparison to untreated mice) in flt1/eGFP BAC mice, which exhibit GFP expression in all murine smaller vessels. Dual labelling of CD31 (endothelial vessel marker) and Ki67 (proliferation marker) revealed active angiogenesis which was predominantly found at tumour margins located near host structures. This finding is in accordance with a study which found high numbers of proliferating endothelial cells at tumour margins of poorly differentiated (and thus aggressive) tumours (Raluca, Cimpean et al. 2015). In these regions the tumour might have orchestrated the microenvironment towards pro-angiogenic conditions in order to induce angiogenesis from surrounding host structures. Studies investigating MVD and the proportion of active VII Discussion 117 angiogenesis in cutaneous melanoma and other tumour entities showed that the number of proliferating endothelial cells (CD31+/CD34+/Nestin+ and Ki67+) but not MVD was correlated with tumour stage (Baeten, Castermans et al. 2006; Hillen, van de Winkel et al. 2006; Kruger, Stefansson et al. 2013). Thus, the proportion of proliferating endothelial cells might be also an indicator for tumour progression and possibly also for the patient’s outcome. In our study, in untreated mice 39.1% of endothelial vessels showed proliferation activity compared to 30.9% of proliferating vessels in macrophage-depleted mice (p=0.114). As tumours in untreated mice showed a tendency of an increased rate of active angiogenesis in comparison to macrophage-depleted mice, TAM might have directly shaped the tumour microenvironment by secreting pro-angiogenic factors – probably in response to a crosstalk with melanoma cells due to hypoxic conditions. TAM are in general able to stimulate proliferation and migration of endothelial cells (reviewed in (Lamagna, Aurrand-Lions et al. 2006)) and may thereby influence the rate of active angiogenesis in aggressive tumours. This indicates another pathway by which TAM may directly affect histopathologic tumour characteristics. The origin of tumour vessels When tumours grow beyond the size which can be supplied by diffusion from surrounding structures, they are forced to angiotropic growth and/or to attract blood vessels from the surrounding stroma and/or to initiate other vascularisation strategies. This process is regulated by a variety of pro- and anti-angiogenic factors, and it is an indispensable prerequisite for further growth of the tumour (Carmeliet and Jain 2000). In uveal, cutaneous and oral melanoma, numerous studies have focused on angiogenesis and other mechanisms of tumour vascularisation. Besides sprouting angiogenesis, also patterns of mosaic vessels, tumour-lined vasculature, co-option of host vessels and networks of vessel-like structures not lined by VII Discussion 118 endothelial cells (vasculogenic mimicry) have been described (reviewed in (Pastushenko, Vermeulen et al. 2014), (Hendrix, Seftor et al. 2002; Lee, Nagai et al. 2002; Zhang, Zhang et al. 2006). These studies used PAS and immunohistochemical staining of endothelial vessels, ECM and other structures (e.g. CD31, collagen IV) to visualise and investigate the characteristics of melanoma vascularisation. In our study, we further addressed the origin of tumour vessels by using the flt-1/eGFP BAC mouse with endogenous endothelial GFP expression. Almost all intratumoural vessels exhibited a co-expression of CD31 and GFP and were thus of host origin (chapter V). Apparently, rapidly growing tumours are dependent on a highly pro-angiogenic microenvironment in which they are able to adopt host vessels and initiate angiogenic sprouting from incorporated vessels as a main vascularisation strategy. This is in accordance with the general doctrine that angiogenesis represents the main pathway of tumour vascularisation. However, a minor number of intratumoural vessels in the described flt-1/eGFP BAC mouse model were found to be solely marked by the CD31 antibody and must, subsequently, be of other origin than angiogenesis. These vessels might have originated from two other vascularisation strategies: vasculogenesis (endothelial precursor cells) or tumour-derived vessels. To distinguish its origin further investigation of their surface markers are needed. Circulating endothelial precursors - shed from the vessel wall or mobilised from the bone marrow - can also contribute to tumour vascularisation in terms of vasculogenesis (Rafii 2000). Such endothelial precursor cells express characteristic vessel markers (e.g. CD31, CD34, VEGFR-2) along with the stem cell marker CD133 (Timmermans, Plum et al. 2009). To identify such vessels formed by endothelial progenitor cells we (intend to) perform dual staining of CD31 and CD133. Should we be able to demonstrate that these vessels originated from vasculogenesis by being built of endothelial progenitor cells (CD31+CD133+GFP-), this would explain heterogeneity of VII Discussion 119 tumour vascularisation in our model. Further, flt-1/eGFP BAC mice would represent a potent model to distinguish between vasculogenesis (CD31+CD133+GFP- vessels) and angiogenesis (CD31+CD133+GFP+ vessels). This might offer a new model to study these two different forms of vascularisation in further pathologies like injury, inflammatory processes or other tumour entities. On the other hand, if CD31+GFP- vessels in flt-1/eGFP BAC mice reveal not to have originated from vasculogenesis (CD133-), another hypothesis of their origin might be a derivation from cancer stem cells. This hypothesis might further be supported by dual labelling of CD31 and a melanoma marker (e.g. HMB45/MART-1). Two studies in glioblastoma showed that intratumoural blood vessels expressed genetic markers of cancer cells (with stem cell like properties). This suggests that these blood vessels were of tumour origin (Ricci-Vitiani, Pallini et al. 2010; Wang, Chadalavada et al. 2010). Another group describes „tumour endothelial cells (TEC)“ as intratumoural vessels of tumour origin with highly angiogenic phenotypes and higher proliferative and migratory abilities than „normal endothelial cells“ (=host-derived, angiogenic endothelial cells) (Ohmura-Kakutani, Akiyama et al. 2014). However, these conclusions were based on in vitro studies and their translation into in vivo conditions remains unknown. However, in an ischemic nude mouse model with inoculated human aggressive melanoma cells, newly formed vasculature within the tumour showed endothelial cells of human and of mouse origin in a linear arrangement. Thus, besides angiogenic murine vessels, human melanoma cells may have acquired endothelial-like morphology and were incorporated in capillaries. Along these lines, the tumour may actively contribute to neovascularisation (Mihic-Probst, Ikenberg et al. 2012). Our ongoing studies may reveal the origin of CD31+GFP- intratumoural vessels in flt-1/eGFP BAC mice and either offer a new model for distinguishing vasculogenesis from angiogenesis VII Discussion 120 or contribute to recent discussions regarding tumour-derived endothelial vessels. This would represent the first orthotopic animal model of such tumour-derived endothelial cells in melanoma. Outlook Manipulating the tumour microenvironment may offer an approach for novel therapeutic strategies. As TAM represent major orchestrators of the tumour microenvironment, targeting TAM and limiting their pro-angiogenic and tumour-favouring properties may be promising. This could be achieved by blocking the polarisation cascade into an M2 phenotype by e.g. a Stat6 inhibitor (Binnemars- Postma et al. 2015, Poster at the ECC 2015) or by repolarising TAM from the M2 into a tumouricidal M1 phenotype (e.g. by zoledronic acid) and thereby moving the balance toward a tumouricidal immune function. However, when considering future systemic therapies, which alter macrophage properties, other important macrophage functions in resistance against infections or wound healing need to be respected. Further (animal) studies on pathways of secreted cytokines or chemokines are needed which promote such a pro-angiogenic and tumour-favouring microenvironment. Altering these pathways might potentially enable us to help control tumour progression and metastasis. As there are several promising therapeutic options for the primary intraocular tumour, such novel therapeutic strategies will be of interest regarding the prevention of metastasis of uveal melanoma and improving patients’ outcome. Summary No spontaneous melanoma mouse model has been described yet, thus, all murine models are subject to certain limitation. To investigate tumour biology and the role of macrophages in VII Discussion 121 intraocular melanoma, we modified an established mouse model by injecting murine cutaneous HCmel12 melanoma cells into the vitreous of mouse eyes. Intraocular tumours exhibited characteristic histopathologic features similar to human uveal melanoma, which were influenced by advanced age of mice and by the presence of tumour associated macrophages (TAM). The frequency of prognostically significant extravascular matrix patterns and level of active angiogenesis were directly correlated with the presence of TAM. Mean vascular density, collagen IV-positive extracellular matrix and the amount of infiltrating immune cells were significantly increased in tumours with a M2-dominated microenvironment. Old mice exhibited histopathologic and immunologic features, which better resembled human tumours. Thus, we suggest to use mice of advanced age in future studies on the tumour microenvironment. Angiogenesis represented the main vascularisation strategy, however, the origin of few CD31+GFP- vessels – originated from either vasculogenesis or tumour-derived endothelial vessels - remains subject to ongoing investigations. Taken together, an aged immunoprofile provides a basis for tumour growth with M2 macrophages being directly involved into a more aggressive tumour phenotype. 122 VIII References Albert, D. M., A. Kumar, et al. (2004). "Effectiveness of 1alpha-hydroxyvitamin D2 in inhibiting tumor growth in a murine transgenic pigmented ocular tumor model." Arch Ophthalmol 122(9): 1365-9. Allavena, P., A. Sica, et al. (2008). "The Yin-Yang of tumor-associated macrophages in neoplastic progression and immune surveillance." Immunol Rev 222: 155-61. Allavena, P., A. Sica, et al. (2008). "The inflammatory micro-environment in tumor progression: the role of tumor-associated macrophages." Crit Rev Oncol Hematol 66(1): 1-9. Arepalli, S., S. Kaliki, et al. (2015). "Choroidal metastases: origin, features, and therapy." Indian J Ophthalmol 63(2): 122-7. Arnold, L., A. Henry, et al. (2007). "Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis." J Exp Med 204(5): 1057-69. Asahara, T., T. Murohara, et al. (1997). "Isolation of putative progenitor endothelial cells for angiogenesis." Science 275(5302): 964-7. Baeten, C. I., K. Castermans, et al. (2006). "Proliferating endothelial cells and leukocyte infiltration as prognostic markers in colorectal cancer." Clin Gastroenterol Hepatol 4(11): 1351-7. Bakalian, S., J. C. Marshall, et al. (2008). "Molecular pathways mediating liver metastasis in patients with uveal melanoma." Clin Cancer Res 14(4): 951-6. Biswas, S. K. and A. Mantovani (2010). "Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm." Nat Immunol 11(10): 889-96. Bronkhorst, I. H. and M. J. Jager (2012). "Uveal Melanoma: The Inflammatory Microenvironment." J Innate Immun. Bronkhorst, I. H. and M. J. Jager (2013). "Inflammation in uveal melanoma." Eye (Lond) 27(2): 217-23. Bronkhorst, I. H., T. M. Jehs, et al. (2014). "Effect of hypoxic stress on migration and characteristics of monocytes in uveal melanoma." JAMA Ophthalmol 132(5): 614-21. Bronkhorst, I. H., T. H. Vu, et al. (2012). "Different subsets of tumor-infiltrating lymphocytes correlate with macrophage influx and monosomy 3 in uveal melanoma." Invest Ophthalmol Vis Sci 53(9): 5370-8. Burrell, K. and G. Zadeh (2012). Molecular Mechanisms of Tumor Angiogenesis. Carmeliet, P. and R. K. Jain (2000). "Angiogenesis in cancer and other diseases." Nature 407(6801): 249-57. Catalano, V., A. Turdo, et al. (2013). "Tumor and its microenvironment: a synergistic interplay." Semin Cancer Biol 23(6 Pt B): 522-32. Chang, Y. S., E. di Tomaso, et al. (2000). "Mosaic blood vessels in tumors: frequency of cancer cells in contact with flowing blood." Proc Natl Acad Sci U S A 97(26): 1460813. Chen, L., S. Zhang, et al. (2009). "A pilot study of vasculogenic mimicry immunohistochemical expression in intraocular melanoma model." Oncol Rep 21(4): 989-94. VIII References 123 Chroscinski, D., D. Sampey, et al. (2015). "Registered report: tumour vascularization via endothelial differentiation of glioblastoma stem-like cells." Elife 4. Cohen, H. J. (1994). "Biology of aging as related to cancer." Cancer 74(7 Suppl): 2092-100. Damato, B., A. Eleuteri, et al. (2011). "Estimating prognosis for survival after treatment of choroidal melanoma." Prog Retin Eye Res 30(5): 285-95. Daniels, K. J., H. C. Boldt, et al. (1996). "Expression of type VI collagen in uveal melanoma: its role in pattern formation and tumor progression." Lab Invest 75(1): 55-66. Diaz, C. E., D. Rusciano, et al. (1999). "B16LS9 melanoma cells spread to the liver from the murine ocular posterior compartment (PC)." Curr Eye Res 18(2): 125-9. Dithmar, S., D. M. Albert, et al. (2000). "Animal models of uveal melanoma." Melanoma Res 10(3): 195-211. Eagle, R. C., Jr. (2013). "The pathology of ocular cancer." Eye (Lond) 27(2): 128-36. Ehlers, J. P., L. Worley, et al. (2008). "Integrative genomic analysis of aneuploidy in uveal melanoma." Clin Cancer Res 14(1): 115-22. Field, M. G. and J. W. Harbour (2014). "Recent developments in prognostic and predictive testing in uveal melanoma." Curr Opin Ophthalmol 25(3): 234-9. Folberg, R., M. J. Hendrix, et al. (2000). "Vasculogenic mimicry and tumor angiogenesis." Am J Pathol 156(2): 361-81. Folberg, R., M. Mehaffey, et al. (1997). "The microcirculation of choroidal and ciliary body melanomas." Eye (Lond) 11 ( Pt 2): 227-38. Folberg, R., J. Pe'er, et al. (1992). "The morphologic characteristics of tumor blood vessels as a marker of tumor progression in primary human uveal melanoma: a matched casecontrol study." Hum Pathol 23(11): 1298-305. Folberg, R., V. Rummelt, et al. (1993). "The prognostic value of tumor blood vessel morphology in primary uveal melanoma." Ophthalmology 100(9): 1389-98. Folkman, J. (1971). "Tumor angiogenesis: therapeutic implications." N Engl J Med 285(21): 1182-6. Foss, A. J., R. A. Alexander, et al. (1996). "Microvessel count predicts survival in uveal melanoma." Cancer Res 56(13): 2900-3. Gazzaniga, S., A. I. Bravo, et al. (2007). "Targeting tumor-associated macrophages and inhibition of MCP-1 reduce angiogenesis and tumor growth in a human melanoma xenograft." J Invest Dermatol 127(8): 2031-41. Gilkes, D. M., G. L. Semenza, et al. "Hypoxia and the extracellular matrix: drivers of tumour metastasis." Nat Rev Cancer 14(6): 430-9. Gomez, C. R., E. D. Boehmer, et al. (2005). "The aging innate immune system." Curr Opin Immunol 17(5): 457-62. Gomez, C. R., V. Nomellini, et al. (2008). "Innate immunity and aging." Exp Gerontol 43(8): 718-28. Gordon, S. and F. O. Martinez (2010). "Alternative activation of macrophages: mechanism and functions." Immunity 32(5): 593-604. Griewank, K. G., H. Westekemper, et al. (2013). "Conjunctival melanomas harbor BRAF and NRAS mutations and copy number changes similar to cutaneous and mucosal melanomas." Clin Cancer Res 19(12): 3143-52. Grossniklaus, H. E. (1998). "Tumor vascularity and hematogenous metastasis in experimental murine intraocular melanoma." Trans Am Ophthalmol Soc 96: 721-52. Hanahan, D. and R. A. Weinberg (2011). "Hallmarks of cancer: the next generation." Cell 144(5): 646-74. VIII References 124 Hendrix, M. J., E. A. Seftor, et al. (2003). "Vasculogenic mimicry and tumour-cell plasticity: lessons from melanoma." Nat Rev Cancer 3(6): 411-21. Hendrix, M. J., R. E. Seftor, et al. (2002). "Transendothelial function of human metastatic melanoma cells: role of the microenvironment in cell-fate determination." Cancer Res 62(3): 665-8. Herwig, M. C., C. Bergstrom, et al. (2013). "M2/M1 ratio of tumor associated macrophages and PPAR-gamma expression in uveal melanomas with class 1 and class 2 molecular profiles." Exp Eye Res 107: 52-8. Herz, K., J. C. Heinemann, et al. (2012). "Live monitoring of small vessels during development and disease using the flt-1 promoter element." Basic Res Cardiol 107(2): 257. Hillen, F. and A. W. Griffioen (2007). "Tumour vascularization: sprouting angiogenesis and beyond." Cancer Metastasis Rev 26(3-4): 489-502. Hillen, F., A. van de Winkel, et al. (2006). "Proliferating endothelial cells, but not microvessel density, are a prognostic parameter in human cutaneous melanoma." Melanoma Res 16(5): 453-7. Hornung, V., F. Bauernfeind, et al. (2008). "Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization." Nat Immunol 9(8): 84756. Hurst, E. A., J. W. Harbour, et al. (2003). "Ocular melanoma: a review and the relationship to cutaneous melanoma." Arch Dermatol 139(8): 1067-73. Hussein, M. R. (2006). "Tumour-associated macrophages and melanoma tumourigenesis: integrating the complexity." Int J Exp Pathol 87(3): 163-76. Ioachim, E., A. Charchanti, et al. (2002). "Immunohistochemical expression of extracellular matrix components tenascin, fibronectin, collagen type IV and laminin in breast cancer: their prognostic value and role in tumour invasion and progression." Eur J Cancer 38(18): 2362-70. Jackaman, C., H. G. Radley-Crabb, et al. (2013). "Targeting macrophages rescues age-related immune deficiencies in C57BL/6J geriatric mice." Aging Cell 12(3): 345-57. Jager, M. J., L. V. Ly, et al. (2011). "Macrophages in uveal melanoma and in experimental ocular tumor models: Friends or foes?" Prog Retin Eye Res 30(2): 129-46. Jung, S., J. Aliberti, et al. (2000). "Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion." Mol Cell Biol 20(11): 4106-14. Kaliki, S., C. L. Shields, et al. (2015). "Uveal melanoma: estimating prognosis." Indian J Ophthalmol 63(2): 93-102. Keating, G. M. (2014). "Bevacizumab: a review of its use in advanced cancer." Drugs 74(16): 1891-925. Kilian, M. M., K. U. Loeffler, et al. (2015). "Intravitreally Injected HCmel12 Melanoma Cells Serve as a Mouse Model of Tumor Biology of Intraocular Melanoma." Curr Eye Res: 1-8. Kramer, T. R., M. B. Powell, et al. (1998). "Pigmented uveal tumours in a transgenic mouse model." Br J Ophthalmol 82(8): 953-60. Krecicki, T., M. Zalesska-Krecicka, et al. (2001). "Expression of type IV collagen and matrix metalloproteinase-2 (type IV collagenase) in relation to nodal status in laryngeal cancer." Clin Otolaryngol Allied Sci 26(6): 469-72. VIII References 125 Kruger, K., I. M. Stefansson, et al. (2013). "Microvessel proliferation by co-expression of endothelial nestin and Ki-67 is associated with a basal-like phenotype and aggressive features in breast cancer." Breast 22(3): 282-8. Kujala, E., T. Makitie, et al. (2003). "Very long-term prognosis of patients with malignant uveal melanoma." Invest Ophthalmol Vis Sci 44(11): 4651-9. Lake, S. L., F. Jmor, et al. (2011). "Multiplex ligation-dependent probe amplification of conjunctival melanoma reveals common BRAF V600E gene mutation and gene copy number changes." Invest Ophthalmol Vis Sci 52(8): 5598-604. Lamagna, C., M. Aurrand-Lions, et al. (2006). "Dual role of macrophages in tumor growth and angiogenesis." J Leukoc Biol 80(4): 705-13. Landsberg, J., E. Gaffal, et al. (2010). "Autochthonous primary and metastatic melanomas in Hgf-Cdk4 R24C mice evade T-cell-mediated immune surveillance." Pigment Cell Melanoma Res 23(5): 649-60. Landsberg, J., J. Kohlmeyer, et al. (2012). "Melanomas resist T-cell therapy through inflammation-induced reversible dedifferentiation." Nature 490(7420): 412-6. Latendresse, J. R., L. Muskhelishvili, et al. (2007). "Two cases of uveal amelanotic melanoma in transgenic Tyr-HRAS+ Ink4a/Arf heterozygous mice." Toxicol Pathol 35(6): 82732. Lattier, J. M., H. Yang, et al. (2013). "Host pigment epithelium-derived factor (PEDF) prevents progression of liver metastasis in a mouse model of uveal melanoma." Clin Exp Metastasis. Lee, Y. J., N. Nagai, et al. (2002). "Angioarchitecture of primary oral malignant melanomas." J Histochem Cytochem 50(11): 1555-62. Ly, L. V., A. Baghat, et al. (2010). "In aged mice, outgrowth of intraocular melanoma depends on proangiogenic M2-type macrophages." J Immunol 185(6): 3481-8. Ly, L. V., O. F. Odish, et al. (2010). "Intravascular presence of tumor cells as prognostic parameter in uveal melanoma: a 35-year survey." Invest Ophthalmol Vis Sci 51(2): 658-65. Maat, W., L. V. Ly, et al. (2008). "Monosomy of chromosome 3 and an inflammatory phenotype occur together in uveal melanoma." Invest Ophthalmol Vis Sci 49(2): 50510. Makitie, T., P. Summanen, et al. (1999). "Microvascular density in predicting survival of patients with choroidal and ciliary body melanoma." Invest Ophthalmol Vis Sci 40(11): 2471-80. Makitie, T., P. Summanen, et al. (2001). "Tumor-infiltrating macrophages (CD68(+) cells) and prognosis in malignant uveal melanoma." Invest Ophthalmol Vis Sci 42(7): 141421. Mallikarjuna, K., V. Pushparaj, et al. (2007). "Expression of epidermal growth factor receptor, ezrin, hepatocyte growth factor, and c-Met in uveal melanoma: an immunohistochemical study." Curr Eye Res 32(3): 281-90. Mallone, S., E. De Vries, et al. (2012). "Descriptive epidemiology of malignant mucosal and uveal melanomas and adnexal skin carcinomas in Europe." Eur J Cancer 48(8): 116775. Maniotis, A. J., R. Folberg, et al. (1999). "Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry." Am J Pathol 155(3): 739-52. Mantovani, A., F. Marchesi, et al. (2007). "Inflammation and cancer: breast cancer as a prototype." Breast 16 Suppl 2: S27-33. VIII References 126 Mantovani, A. and A. Sica (2010). "Macrophages, innate immunity and cancer: balance, tolerance, and diversity." Curr Opin Immunol 22(2): 231-7. Mantovani, A., A. Sica, et al. (2004). "The chemokine system in diverse forms of macrophage activation and polarization." Trends Immunol 25(12): 677-86. Mantovani, A., S. Sozzani, et al. (2002). "Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes." Trends Immunol 23(11): 549-55. Mehnert, J. M. and H. M. Kluger (2012). "Driver mutations in melanoma: lessons learned from bench-to-bedside studies." Curr Oncol Rep 14(5): 449-57. Mendoza, M. and C. Khanna (2009). "Revisiting the seed and soil in cancer metastasis." Int J Biochem Cell Biol 41(7): 1452-62. Mihic-Probst, D., K. Ikenberg, et al. (2012). "Tumor cell plasticity and angiogenesis in human melanomas." PLoS One 7(3): e33571. Mosser, D. M. and X. Zhang (2008). "Activation of murine macrophages." Curr Protoc Immunol Chapter 14: Unit 14 2. Mueller, A. J., A. J. Maniotis, et al. (2002). "An orthotopic model for human uveal melanoma in SCID mice." Microvasc Res 64(2): 207-13. Nathanson, S. D. (2003). "Insights into the mechanisms of lymph node metastasis." Cancer 98(2): 413-23. Ohmura-Kakutani, H., K. Akiyama, et al. (2014). "Identification of tumor endothelial cells with high aldehyde dehydrogenase activity and a highly angiogenic phenotype." PLoS One 9(12): e113910. Onken, M. D., A. Y. Lin, et al. (2005). "Association between microarray gene expression signature and extravascular matrix patterns in primary uveal melanomas." Am J Ophthalmol 140(4): 748-9. Onken, M. D., L. A. Worley, et al. (2012). "Collaborative ocular oncology group report number 1: prospective validation of a multi-gene prognostic assay in uveal melanoma." Ophthalmology 119(8): 1596-603. Onken, M. D., L. A. Worley, et al. (2004). "Gene expression profiling in uveal melanoma reveals two molecular classes and predicts metastatic death." Cancer Res 64(20): 7205-9. Pastushenko, I., P. B. Vermeulen, et al. (2014). "Mechanisms of tumour vascularization in cutaneous malignant melanoma: clinical implications." Br J Dermatol 171(2): 220-33. Pierard, G. E., C. Pierard-Franchimont, et al. (2012). "Malignant melanoma and its stromal nonimmune microecosystem." J Oncol 2012: 584219. Prescher, G., N. Bornfeld, et al. (1996). "Prognostic implications of monosomy 3 in uveal melanoma." Lancet 347(9010): 1222-5. Rafii, S. (2000). "Circulating endothelial precursors: mystery, reality, and promise." J Clin Invest 105(1): 17-9. Raluca, B. A., A. M. Cimpean, et al. (2015). "Endothelial Cell Proliferation and Vascular Endothelial Growth Factor Expression in Primary Colorectal Cancer and Corresponding Liver Metastases." Asian Pac J Cancer Prev 16(11): 4549-53. Ricci-Vitiani, L., R. Pallini, et al. (2010). "Tumour vascularization via endothelial differentiation of glioblastoma stem-like cells." Nature 468(7325): 824-8. Sadoun, E. and M. J. Reed (2003). "Impaired angiogenesis in aging is associated with alterations in vessel density, matrix composition, inflammatory response, and growth factor expression." J Histochem Cytochem 51(9): 1119-30. VIII References 127 Schiffner, S., B. M. Braunger, et al. (2014). "Tg(Grm1) transgenic mice: a murine model that mimics spontaneous uveal melanoma in humans?" Exp Eye Res 127: 59-68. Schmid, M. C. and J. A. Varner (2010). "Myeloid cells in the tumor microenvironment: modulation of tumor angiogenesis and tumor inflammation." J Oncol 2010: 201026. Seftor, R. E., A. R. Hess, et al. (2012). "Tumor cell vasculogenic mimicry: from controversy to therapeutic promise." Am J Pathol 181(4): 1115-25. Seftor, R. E., E. A. Seftor, et al. (2001). "Cooperative interactions of laminin 5 gamma2 chain, matrix metalloproteinase-2, and membrane type-1-matrix/metalloproteinase are required for mimicry of embryonic vasculogenesis by aggressive melanoma." Cancer Res 61(17): 6322-7. Shields, C. L., S. Kaliki, et al. (2013). "American Joint Committee on Cancer classification of posterior uveal melanoma (tumor size category) predicts prognosis in 7731 patients." Ophthalmology 120(10): 2066-71. Shields, C. L., J. Manalac, et al. (2014). "Choroidal melanoma: clinical features, classification, and top 10 pseudomelanomas." Curr Opin Ophthalmol 25(3): 177-85. Sica, A. (2010). "Role of tumour-associated macrophages in cancer-related inflammation." Exp Oncol 32(3): 153-8. Sica, A. and A. Mantovani (2012). "Macrophage plasticity and polarization: in vivo veritas." J Clin Invest 122(3): 787-95. Singh, A. D., C. L. Shields, et al. (2001). "Prognostic factors in uveal melanoma." Melanoma Res 11(3): 255-63. Singh, A. D., M. E. Turell, et al. (2011). "Uveal melanoma: trends in incidence, treatment, and survival." Ophthalmology 118(9): 1881-5. Singh, P. and A. Singh (2012). "Choroidal melanoma." Oman J Ophthalmol 5(1): 3-9. Sisley, K., I. G. Rennie, et al. (1990). "Cytogenetic findings in six posterior uveal melanomas: involvement of chromosomes 3, 6, and 8." Genes Chromosomes Cancer 2(3): 205-9. Stout, R. D. and J. Suttles (2005). "Immunosenescence and macrophage functional plasticity: dysregulation of macrophage function by age-associated microenvironmental changes." Immunol Rev 205: 60-71. Syed, N. A., J. J. Windle, et al. (1998). "Transgenic mice with pigmented intraocular tumors: tissue of origin and treatment." Invest Ophthalmol Vis Sci 39(13): 2800-5. Timar, J. and J. Toth (2000). "Tumor sinuses- vascular channels." Pathol Oncol Res 6(2): 836. Timmermans, F., J. Plum, et al. (2009). "Endothelial progenitor cells: identity defined?" J Cell Mol Med 13(1): 87-102. Toivonen, P., T. Makitie, et al. (2004). "Microcirculation and tumor-infiltrating macrophages in choroidal and ciliary body melanoma and corresponding metastases." Invest Ophthalmol Vis Sci 45(1): 1-6. Tolleson, W. H., J. C. Doss, et al. (2005). "Spontaneous uveal amelanotic melanoma in transgenic Tyr-RAS+ Ink4a/Arf-/- mice." Arch Ophthalmol 123(8): 1088-94. Tormo, D., A. Ferrer, et al. (2006). "Rapid growth of invasive metastatic melanoma in carcinogen-treated hepatocyte growth factor/scatter factor-transgenic mice carrying an oncogenic CDK4 mutation." Am J Pathol 169(2): 665-72. van den Bosch, T., E. Kilic, et al. (2010). "Genetics of uveal melanoma and cutaneous melanoma: two of a kind?" Dermatol Res Pract 2010: 360136. van Rooijen, N. and E. van Kesteren-Hendrikx (2003). ""In vivo" depletion of macrophages by liposome-mediated "suicide"." Methods Enzymol 373: 3-16. VIII References 128 Verheul, H. M., E. E. Voest, et al. (2004). "Are tumours angiogenesis-dependent?" J Pathol 202(1): 5-13. Virgili, G., G. Gatta, et al. (2007). "Incidence of uveal melanoma in Europe." Ophthalmology 114(12): 2309-15. Wang, R., K. Chadalavada, et al. (2010). "Glioblastoma stem-like cells give rise to tumour endothelium." Nature 468(7325): 829-33. Yamaura, H. and T. Matsuzawa (1980). "Decrease in capillary growth during aging." Exp Gerontol 15(2): 145-50. Zhang, S., D. Zhang, et al. (2006). "Morphologic research of microcirculation patterns in human and animal melanoma." Med Oncol 23(3): 403-9. 129 IX Summary Marta Kilian The Role of Macrophages in Murine Intraocular Melanoma Uveal melanoma is the most common intraocular tumour in the adult Caucasian population. While local treatment of the primary ocular tumour is very effective, prognosis worsens dramatically when metastatic disease appears. Hepatic micrometastases may establish early and remain dormant for decades. Understanding its pathways and preventing hematogeneous metastasis is a major goal uveal melanoma research. Several prognostic factors have been determined so far, including clinical (e.g. age), histopathologic, (e.g. tumour size, mean vascular density [MVD], vasculogenic mimicry/ extravascular matrix patterns, tumour associated macrophages [TAM]) and genetic characteristics. However, in vivo studies in mice remain challenging. All murine models are subject to restrictions as spontaneous uveal melanoma has not been described in wild type mice, yet. In search for spontaneous ocular melanomas, among other groups, we examined the eyes of transgenic mice which exhibit spontaneous skin melanoma. Apart from an incidental and unusual finding of heavily pigmented cells in the cornea, Hgf-Cdk4 mice did not show any melanocytic proliferations in the uvea (chapter VI). However, this underlines genetic and clinical differences between uveal and skin melanoma. Besides mouse models in which human uveal melanoma cells are injected into the eye of immunodeficient mice, there are also other well established mouse models like intraocular inoculation with murine cutaneous melanoma cells. Despite genetic differences, such a model IX Summary 130 allows investigations on intraocular tumour biology in an immunocompetent animal. We modified such an established mouse model by injecting HCmel12 murine cutaneous melanoma cells into the ocular vitreous of CX3CR1+/GFP mice and flt-1/eGFP BAC mice of different age groups. We were particularly interested in the role of macrophages in intraocular melanoma of aged mice (10-12 months). Intraocular tumours of HCmel12 melanoma cells showed solid and aggressive growth with angiotropism, angiogenesis, vasculogenic mimicry/ extravascular matrix patterns, inflammatory cell infiltration with pronounced macrophage proportions, but no hematogenously dispersed metastases (chapter III). In our studies, old mice exhibited more aggressive tumours (histologically) and an increased number of M2 macrophages (flow cytometry analysis) compared to young mice. Thus, aged mice better resembled uveal melanomas growth characteristics. Hence, our intraocular tumour model in mice of advanced age offers the opportunity to study the tumour microenvironment in a semiclosed system (eye, blood but no lymphatic vessels), which can be manipulated iatrogenically by systemic or intraocular injections. To investigate the general role of macrophages in intraocular melanoma we examined histologically tumour growth characteristics of systemically macrophage-depleted mice in comparison to untreated mice. The frequency of prognostically significant extravascular matrix patterns (chapter IV) and active angiogenesis (chapter V) was increased in untreated mice compared to macrophage-depleted mice. These results suggest that TAM promote different tumour vascularisation strategies in intraocular melanoma and thereby contribute to tumour progression. Further, we studied the role of polarised macrophages which can be functionally subdivided into a M1 phenotype with tumouricidal and a M2 phenotype with tumour-favouring properties. HCmel12 cells were incubated in vitro in a M1- or M2dominated environment and then injected into the mouse eye. Tumours of M2-conditioned IX Summary 131 melanoma cells showed an increased TAM infiltration, more collagen IV stroma patterns and a higher MVD compared to M1-conditioned tumours. Thus, aggressive, pro-angiogenic and tumour promoting factors such as inflammatory cell infiltration, stroma remodelling and MVD could be directly attributed to the existence of TAM, in particular to M2 macrophages (chapter IV). In flt-1/eGFP BAC mice we further found that the majority of tumour vessels were CD31+GFP+ and thus of angiogenic host origin (chapter V). Hence, the main vascularisation strategy in our tumour model was co-option of host vasculature followed by induction of angiogenesis from existing host vessels. However, some vessels were observed to be CD31+GFP- and thus of vasculogenic or tumour-derived origin. Ongoing studies using stem cell and melanoma markers might reveal their final origin and offer potentially new animal models to distinguish between angiogenesis and vasculogenesis in tumours or other pathologies. Taken together, an aged immunoprofile with an increased number of M2 macrophages provides a tumour-favouring microenvironment and might be a basis for tumour progression and metastasis. These findings could be a target for future therapeutic interventions in different tumour entities. 132 X Zusammenfassung Marta Kilian Rolle der Makrophagen beim intraokularen Melanom der Maus Das uveale Melanom ist der häufigste intraokulare Tumor bei der erwachsenen kaukasischen Bevölkerung. Die Behandlung des primären okularen Tumors bietet vielversprechende Therapiemöglichkeiten, jedoch verschlechtert sich die Prognose des Patienten enorm, wenn Metastasen klinisch in Erscheinung treten. Mikrometastasen in der Leber können sich sehr früh etablieren, zugleich aber für mehrere Jahrzehnte “stumm” verbleiben. Das Augenmerk vieler Studien über das uveale Melanom richtet sich daher auf die Signalwege und die Prävention der hämatogenen Metastasierung. Zahlreiche Eigenschaften von Aderhautmelanomen konnten bislang statistisch mit der Prognose des Patienten korreliert werden. Hierzu gehören klinische (z.B. Alter des Patienten), histopathologische (z.B. Tumorgröße, Gefäßdichte, vasculogenic mimicry, tumor-assoziierte Makrophagen) und genetische Charakteristiken. In vivo Studien in Mäusen gestalten sich jedoch nach wie vor schwierig: Alle bisherigen Mausmodelle besitzen gewisse Einschränkungen, da spontane uveale Melanome in Wildtyp-Mäusen noch nicht beschrieben wurden und demnach als adäquates Modell nicht zur Verfügung stehen. Neben zahlreichen anderen Arbeitsgruppen untersuchten auch wir eine neue transgene (HgfCdk4) Mauslinie, welche spontan Hautmelanome entwickelt und deren Augen bisher noch nicht auf die Entstehung von spontan auftretenden uvealen Melanomen überprüft wurde. Abgesehen vom Zufallsbefund stark pigmentierter Zellen in der Cornea der Hgf-Cdk4 Mäuse, X Zusammenfassung 133 konnten wir keinerlei Hinweise auf melanozytäre Proliferationen in der Uvea finden (Kapitel VI). Dies unterstreicht nochmals die Unterschiede in der Pathogenese zwischen kutanen und uvealen Melanomen, die sich sowohl genetisch als auch durch ihr klinisches Verhalten unterscheiden. Neben Mausmodellen, bei denen humane Aderhautmelanomzellen in das Auge immunsupprimierter Mäuse injiziert werden, ist die intraokulare Injektion von murinen Hautmelanomzellen gut etabliert. Trotz gewisser genetischer Unterschiede zwischen Aderhaut- und Hautmelanomzellen, ermöglicht diese Methode Untersuchungen zur intraokularen Tumorbiologie in einem immunologisch funktionsfähigen Organismus. Wir modifizierten daher ein bisher etabliertes Mausmodell und injizierten HCmel12 Zellen (murine, kutane Melanomzellen) in den Glaskörperraum des Auges von CX3CR1+/GFP und flt1/eGFP BAC Mäusen zweier Altersklassen, um insbesondere den Einfluss tumor-assoziierter Makrophagen auf das Tumormikromilieu unter Berücksichtigung des Lebensalters zu untersuchen. Intraokulare Tumore von HCmel12 Zellen zeichneten sich aus durch angiotropes Wachstum, Angiogenese, Vasculogenic Mimicry/ Mikrozirkulationsmuster und eine Immunzellinfiltration mit hohem Makrophagenanteil. Diese Eigenschaften sind allesamt für das humane Aderhautmelanom charakteristisch. Eine rein hämatogene Metastasierung konnte jedoch nicht nachgewiesen werden (Kapitel II). Im Vergleich zu jungen Mäusen zeigten alte Mäuse in unseren Studien ein aggressiveres Wachstumsmuster (histologisch) sowie das vermehrte Auftreten von M2 Makrophagen (durchflusszytometrische Analyse) und wiesen somit eine größere Ähnlichkeit zu humanen uvealen Melanomen auf (Kapitel IV). Demzufolge bietet das von uns etablierte Tumormodell in alten Mäusen die Möglichkeit, das intraokulare Tumormikromilieu in dem semigeschlossenem System des Auges (Blut- aber keine Lymphgefäße) zu untersuchen, welches iatrogen durch systemische oder intraokulare Injektionen manipuliert werden kann. X Zusammenfassung 134 Um die allgemeine Rolle der Makrophagen beim intraokularen Melanom untersuchen zu können, wurden histologische Tumorwachstumseigenschaften von systemisch Makrophagendepletierten Mäusen mit denen unbehandelter Tiere verglichen. Bei unbehandelten Mäusen war die Häufigkeit prognostisch signifikanter Mikrozirkulationsmuster (Kapitel IV) sowie der Anteil aktiver Angiogenese (Kapitel V) erhöht. Diese Ergebnisse lassen vermuten, dass tumor-assoziierte Makrophagen unterschiedliche Vaskularisationsstrategien des Tumors begünstigen und damit zur Tumorprogression beitragen können. Weiterhin untersuchten wir die Rolle von funktionell polarisierten Makrophagen, welche einen tumoriziden M1 Phänotyp oder einen Tumor-begünstigenden M2 Phänotyp annehmen können. Hierfür wurden HCmel12 Zellen in vitro in einem M1- oder M2-dominierten Mikromilieu inkubiert und anschließend ins Mausauge injiziert. Tumore aus M2-konditionierten Melanomzellen zeigten histologisch eine erhöhte Infiltration mit Makrophagen, ein verstärktes Auftreten von Kollagen 4-positiven Strukturen und eine höhere mittlere Gefäßdichte verglichen zu M1konditionierte Tumoren. Demnach konnten wir zeigen, dass tumor-assoziierte Makrophagen und hier insbesondere M2 Makrophagen – durch tumor-begünstigende Faktoren (wie stromale Umbaumechanismen im Sinne von Mikrozirkulationsmustern, Entzündungszellinfiltration und vermehrter Angiogenese) direkt zu einem aggressiveren Tumorphänotyp beitragen können (Kapitel IV). Zudem konnten wir in flt-1/eGFP BAC Mäusen feststellen, dass der Großteil der Tumorgefäße CD31+GFP+ war und demnach murinen Strukturen entstammte. Dies deutet auf “Co-option” von Wirtsgefäßen mit anschließender Angiogenese aus bereits existierenden Mausgefäßen als Hauptvaskularisationsstrategie des Tumors hin (Kapitel V). Allerdings zeigten sich auch vereinzelt CD31+GFP- Gefäße, welche folglich einer anderen Vaskularisierungsstrategie - Vaskulogenese oder aus Tumorzellendothelien - entstammt sein müssen. Laufende Untersuchungen mit Stammzell- und Melanommarkern sollen deren X Zusammenfassung 135 endgültigen Ursprung aufklären. Dies könnte dazu beitragen potentiell ein neues Modell zur Unterscheidung von Angiogenese und Vaskulogenese zu etablieren. Zusammenfassend, ein “gealtertes” Immunsystem mit einem erhöhten Anteil an M2 Makrophagen bietet eine tumor-begünstigende Grundlage. Diese Erkenntnisse können potentielle Ansatzpunkte für zukünftige therapeutische Interventionen darstellen, da sie möglicherweise auf andere Tumoren übertragbar sind. XI Acknowledgements 136 XI Danksagung Forschung: ein buntes Konvolut aus springenden Ideen, geduldigem Feilen an scheinbar zum Scheitern vorbestimmten Methoden, Momenten der unerwarteten Freude wie auch befürchteter Enttäuschungen; Spontanität und Kreativität als auch langwierige Literaturrecherche; finanzielle, bauliche sowie erbauliche Rahmenbedingungen. Mit jedem einzelnen Punkt bin ich mittlerweile sehr vertraut, manchmal meines Widerwollens zum Trotz. Doch schließlich ist es die Freude am Hinterfragen und Entdecken sowie eine erfolgreiche Teamarbeit, die mich an der Wissenschaft begeisterte. Ich weiß mich sehr glücklich zu schätzen, dass ich Teil eines großartigen Labors der Augenklinik Bonn sein durfte, in dem ich diese Erfahrungen machen und meine Promotion abschließen konnte. Mein besonderer Dank gilt Frau Dr. M. Herwig, zum einen für die Möglichkeit an diesem spannenden Thema arbeiten zu dürfen, zum anderen für die wissenschaftliche Anleitung und insbesondere dafür, dass sie mich im Laufe dieses Projektes unentwegt betreut, beraten, kritisiert, motiviert und - vor allem - unterstützt hat. Mein ganz spezieller Dank geht an Frau Prof. K. Löffler, die durch ihre Expertise sowie immerwährende wertvolle Unterstützung das Projekt maßgeblich mitgestaltet und betreut hat. Sehr dankbar bin ich ebenfalls Frau Prof. C. Pfarrer, dass sie mir als „Doktormutter“ eine Promotion an der Tierärztlichen Hochschule Hannover ermöglichte und – trotz der Ferne – mich stets engagiert betreut und freundlich beraten hat. Damit hat sie meinen tierärztlichen Weg in die Forschung geebnet und mir die Möglichkeit gegeben meine Arbeit im tiermedizinischen Rahmen vorzustellen und zu verteidigen. Ebenso gilt großer Dank Herrn Prof. F. Holz, der mit Enthusiasmus und Nachhaltigkeit Forschungsarbeiten an der Universitäts-Augenklink Bonn unterstützt und stets motivierend die Möglichkeiten bot, die eigenen Forschungsfortschritte im Bonner Hause sowie auf nationalen und internationalen Kongressen präsentieren und diskutieren zu dürfen. Des Weiteren möchte ich unseren zahlreichen Kooperationspartnern aus anderen Instituten und Kliniken danken, die unser Projekt ermöglicht und mit Fachkompetenz unterstützt sowie beraten haben: Prof. C. Kurts (Institute of Experimental Immunology, Universität Bonn), Prof. T. Tüting (Labor für Experimentelle Dermatologie, Universität Bonn), Prof. D. Wenzel (Institut für Physiologie I, Universität Bonn), Prof. E. Latz (Institut für angeborene Immunität, Universität Bonn) und Prof. H. Grossniklaus (Emory Eye Center, Emory University, Atlanta, GA, USA), Tobias Höller (Institut für Medizinische Biometrie, Informatik und Epidemiologie, Universität Bonn). Besonderer Dank gilt nicht minder den Wissenschaftlern, technischen Angestellten und Mitarbeitern der genannten Institute, die mir stets helfend und beratend zur Seite standen: Dr. Andre Tittel, Dr. Daniel Engel, Andreas Dolf, Peter Wurst, Tobias Bald, Katia Herz, Anne Alfter, Florian Hoß. XI Acknowledgements 137 Herzlich möchte ich P. Widmar und C. Strack danken für ihre geduldigen Einweisungen, hilfreichen Ratschläge sowie tatkräftige Zusammenarbeit und die damit verbundene große Unterstützung bei der Verwirklichung des Projektes. Ein ganz besonderer Dank geht allgemein an das gesamte Laborteam der Augenklinik Bonn. Wenn auch verschiedenen AGs zugeordnet, so waren wir doch immer eine Gemeinschaft, die sich gegenseitig unterstützt, motiviert und geholfen hat, die zusammen geforscht und gelacht sowie auf Kongresse gefahren und Nachlesen gehalten hat. Und wenn man manchmal das Gefühl hatte, dass es um Leben und Toad, Gelingen oder Scheitern eines Versuches oder Vortrages ging, konnte man sich stets auf fachliche sowie menschliche Unterstützung verlassen, sodass es einen nicht all zu sehr aus der Kurve geworfen hat. Herzlichen Dank Karin Löffler, Alfred Wegener, Tim Krohne, Martina Herwig, Parand Widmar, Claudine Strack, Johanna Meyer, Carolina Brandstetter, Franziska Lörch, Wolf Harmening, Michael Linden, Niklas Domdei…