Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
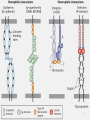
1 • • • • • • THE EXTRACELLULAR MATRIX (ECM) Cells are embedded in a jelly of proteins and polysaccharides called the extracellular matrix or the ground substance In epithelial tissues, cells are tightly attached together to form sheets; ECM is scanty and forms the basal lamina The basal lamina forms a supporting layer underlying the epithelia and helps prevent the cells from ripping apart In connective tissues ECM forms a larger space and carries the mechanical stress to which the tissue is subjected The ECM serves as a reservoir for many extracellular signaling molecules that control cell growth and differentiation In addition, it provides a network through or on which cells can move during tissue assembly 2 • The cells in animal tissues are “glued” together by cell adhesion molecules (CAMs), proteins embedded in their surface membranes • CAMs mediate cell-cell and cell-ECM interactions • CAMs fall into four major families: the cadherins, immunoglobulin (Ig) superfamily, integrins and selectins • CAMs mediate, through their extracellular domains, adhesive interactions between cells of the same type (homotypic adhesion) or between cells of different types (heterotypic adhesion) • A CAM on one cell can directly bind to the same kind of CAM on an adjacent cell (homophilic binding) or to a different class of CAM (heterophilic binding) • Clustered CAMs form cell junctions • The cytosolic domains of CAMs form connections with cytoskeletal elements 3 4 • Three types of molecules are abundant in the extracellular matrix of all tissues: Highly viscous proteoglycans –a group of glycoproteins that cushion cells and bind many extracellular molecules Fibrous proteins–which provide mechanical strength Soluble multiadhesive matrix proteins, which bind to and cross-link cell-surface adhesion receptors and other ECM components The extracellular matrix of epithelial tissues • All epithelial cells in a sheet are connected to one another and the extracellular matrix by specialized cell junctions consisting of dense clusters of CAMs • Anchoring junctions and tight junctions perform the key task of holding cells together into tissues • Gap junctions permit the rapid diffusion of small, watersoluble molecules between the cytoplasm of adjacent cells • The primary CAMs in the cell-cell interactions of epithelia are cadherins; some cell–cell many cell–matrix interactions are 5 mediated by integrins • The basal lamina can be a single layer (intestine, skin,…) or it can surround each cell (muscle, adipocytes,…) • Most of the ECM components in the basal lamina are synthesized by the cells that rest on it • Four protein components are found in basal laminae: Type IV collagen –trimeric molecules with both rodlike and globular domains that form a two-dimensional network Laminins –a family of multiadhesive proteins that form a fibrous two-dimensional network with type IV collagen and that also bind to integrins Entactin –a rodlike protein that cross-links type IV collagen and laminin and helps incorporate other components Perlecan –a large proteoglycan that binds to and cross-links many ECM components and cell-surface molecules • Type IV collagen, is one of more than 20 types of collagen that participate in the formation of the ECM in various tissues 6 7 • Collagen accounts for around 25 % of the total protein content of the body • All collagens are fibrous proteins made from three polypeptides called collagen α chains • Unlike the α helix secondary structure of many proteins, the structure of collagen is a left-handed helix • The three chains wrap around each other to form a righthanded triple helix • The collagen triple helix can form because of an unusual abundance of three amino acids: glycine, proline and hydroxyproline They make up the characteristic repeating pattern Gly-X-Y, where X and Y can be any amino acid but are often proline and hydroxyproline and less often lysine and hydroxylysine • Glycine is essential because its small side chain, a hydrogen atom, is the only one that can fit into the crowded center of 8 the three stranded helix • Although the rigid peptidylproline and peptidyl hydroxyproline linkages are not compatible with formation of a classic single-stranded helix, they stabilize the distinctive three-stranded collagen helix The hydroxyl group of hydroxyproline (by forming hydrogen bonding with members of other chains) helps hold its ring in a conformation that stabilizes the triple helix • The triple helix of collagen IV is interrupted at several points by non-helical segments that give flexibility to the structure • Type IV collagen is a sheet/network forming type Triple helices associate with each other through the globular domains at the C and N terminals • Laminin is a heterotrimeric cross-shaped protein • Different regions of laminin bind to cell-surface receptors and various matrix components e.g., collagen, proteoglycans, integrins,… 9 10 11 • Proteoglycans are the products of the covalent linkage of core proteins with glycosaminoglycans (GAG) • They contain more carbohydrates than glycoproteins • GAGs are composed of repeating disaccharide units the first sugar of the disaccharide is either N-acetylglucosamine or N-acetylgalactosamine the second sugar is usually either glucuronate, iduronate or galactose the sugars are usually sulfated • GAGs are classified based on the nature of the repeating disaccharide unit: heparan sulfate, chondroitin sulfate, dermatan sulfate, keratan sulfate and hyaluronan • A hypersulfated form of heparan sulfate called heparin, produced mostly by mast cells, plays a key role in allergic reactions 12 • Heparin is used medically as an anticlotting agent because a pentasaccharide unit of heparin binds and activates antithrombin III 13 14 15 • The glycosylation of core proteins begins in the lumen of the endoplasmic reticulum and continues in the Golgi • A link trisaccharide containing xylose and two galactose residues can be the first to attach through O-glycosidic bond with a Serine of the core proteins and then the other sugares are added • Or the modified sugars could be directly be attached through N-glycosidic bond with Asparagine residue of the core proteins • Once chain formation is over, sulfotransferases add sulfate which is obtained from (PAPS) The abundance of negative charges increases polarity The proteoglycans bind large amounts of water and fill the gaps between the other components of the ECM in the form of a hydrated gel • Epimerization of glucuronate to iduronate also takes place 16 after the chain is synthesized • Specific types of GAGs are abundant in certain tissues: Hyaluronate – especially in embryonic tissue; allows migration of cells during morphogenesis and wound healing; vitreous humor = 1 % hyaluronate and 98 % water Chondroitin sulfate – along with hyaluronate, gives compressibility to the bone and cartillage Keratan and dermatan surface –lie between collagen fibrils and play a role in corneal transparency o Heparin –can dislodge LPL from capillaries • GAGs are degraded by lysosomal enzymes: glycosidases and sulfatases • Various degrees of deficiency of these enzymes leads to different types of mucopolysaccharidoses Rare diseases inherited in an autosomal recessive manner e.g. Hurler Syndrome - deficiency of α-L- iduronidase Hunter Syndrome - iduronate sulfatase 17 • • • • • • The extracellular matrix of non-epithelial tissues Connective tissues are composed of ECM for the most part The main components include hyalruornate, various proteoglycans and collagen Rubber-like elastin fibers, which can be stretched and relaxed, also are present in deformable sites (e.g., skin, tendons, heart, lungs) Multi adhesive proteins known as fibronectins are characteristic of connective tissue The various components of the ECM of connective tissues are produced largely by cells called fibroblasts 80-90% of the collagen in the body consists of types IIII –fiber forming collagen The triple helices of type I-III collagen associate into collagen fibrils, which in turn often aggregate into larger bundles called collagen fibers 18 • Collagen is first synthesized as preprocollagen which contains a signal sequence • In the lumen of the endoplasmic reticulum, preprocollagen is modified: removal of the signal sequence, hydroxylation of proline and lysine residues and glycosylation of some hydroxylysine residues preprocollagen is changed to procollagen • In the Golgi apparatus procollagen is associated into a triple helix (initiated through disulfide bonds between C-terminal residues) and released to the extracellular space • Extracellular peptidases remove the N- and C-terminal propeptides to give tropocollagen • Tropocollagens associate laterally to generate fibrils • In fibrils, adjacent collagen molecules are displaced from one another by about one-quarter of their length 19 Striation is observed in electron micrographs 20 • The fibrils are strengthened through covalent cross-links Lysine and hydroxylysine side chains are modified by extracellular lysyl oxidases to form aldehydes (allysine and hydoxyallysine) in place of the amine group at the end of the side chain The aldehydes form covalent crosslinks with lysine, hydroxylysine and histidine residues in adjacent molecules • The cross-links stabilize the side-by-side packing of collagen molecules and generate a strong fibril • Removal of the propeptides and covalent cross-linking take place in the extracellular space to prevent the potentially catastrophic assembly of fibrils within the cell • Type I, II and III collagen fibers are characteristic of the ECM of the (skin, bone and tendons); cartilage and arteries, respectively 21 22 23 Disorders of collagen synthesis • Scurvy: in addition to its antioxidant properties, vitamin C plays an essential role in collagen synthesis prolyl hydroxylase and lysyl hydroxylase reactions require O2, α-ketoglutarate, ascorbic acid and the enzymes contain Fe+2 Proline (lysine) changed to hydroxyproline (hydroxylysine); α-ketoglutarate is decarboxylated to succinate Ascorbate is not directly involved in the reaction The hydroxylases can also decarboxylate α-ketoglutarate without hydroxylating proline and lysine During this reaction, the Fe+2 becomes oxidized, and the oxidized form of the enzyme is unable to hydroxylate proline or lysine The ascorbate consumed in the reaction presumably functions to reduce the heme iron and restore enzyme 24 activity • If vitamin diet is deficient in vitamin C , hydrogen bonds cannot be formed between strands in the collagen triple helix • the melting point of collagen fibers decreases –loss of stability • Scurvy leads to lesions in the skin and blood vessels, bleeding gums, swelling joints, poor wound healing, and, in 25 its advanced stages, it can lead to grotesque disfiguration • Genetic abnormalities in collagen synthesis include: Osteogenesis imperfecta – “brittle bone syndrome” Abnormally Short chains make up the collagen I triple helix ; or other amino acids may take the place of glycine Ehlers-Danlos syndrome –about ten types lack of peptidases; resistance to peptidases; insufficient synthesis of collagen; rapid degradation;… recurrent dislocation of joints, hyperextensible skin • In general, collagen fibers make connective tissues resistant to stretching; the swelling caused by the water associated with hyaluronate gives compressibility • The predominant proteoglycan in cartilage, called aggrecan, assembles with hyaluronan into very large aggregates 26 27 • Elastic fibers are composed of elastin and microfibrils • Elastin is synthesized as a soluble monomer known as tropoelastin • Unlike procollagen, tropoelastin does not have extension peptides, Gly-X-Y sequences, triple helices or attached carbohydrates; little hydroxyproline and no hydroxylysine • Lysyl oxidase in the extracellular space oxidatively deaminates lysine residues of tropoelastin • Cross links are formed between elastin molecules making use of three modified lysine residues (aldehydes) and one unmodified lysine (amino) the linkage may be desmosine or isodesmosine • The insoluble cross-linked elastin molecules are deposited on a network of microfibrils 28 29 • Microfibrils are polymers of the glycoprotein fibrillin • Mutations in the gene for fibrillin lead to Marfan’s syndrome dislocated lens, long digits and dilated aorta o Please read about the elastase in the lungs and its inhibitor • Fibronectins help attach cells to the extracellular matrix by binding to other ECM components, particularly fibrous collagens and heparan sulfate proteoglycans, and to cell surface adhesion receptors such as integrins • Fibronectins are dimers of two similar polypeptides linked at their C-termini by two disulfide bonds • Each chain comprises six functional regions with different ligand-binding specificities • Integrins are usually bound by repeats of Arg-Gly-Asp (RGD) sequences of fibronectins 30 31 32 • After adhesive interactions in epithelia form during differentiation, they often are very stable and can last for the lifetime of the cells or until the cells become loosely-associated • The formation and breaking of adhesive interactions is a characteristic mainly of certain non-epithelial tissues e.g., during morphogenesis, inflammation,… The movement of leukocytes into tissues • To fight infection and clear away damaged tissue, leukocyte must move rapidly from the blood, where they circulate as unattached, into the underlying tissue at sites of infection • This process is known as extravasation • First inflammatory signals (chemokines from the endothelium and leukocytes) cause activation of the endothelium The synthesis of the CAMs is part of the activation of the endothelium 33 • The lectin domain of P-selectin found on endothelial cells will bind the oligosaccharides present in the glycoproteins and glycolipids of leukocytes • Because of the force of the blood flow and the rapid “on” and “off” rates of P-selectin binding to its ligands, the leukocytes are slowed but not stopped and roll along the endothelium • For tight adhesion to occur, certain integrins on leukocytes are activated by chemokines and PAF the integrin is changed to a high affinity form • Activated integrins on leukocytes then bind to each of two distinct Ig-CAMs (ICAM 1 and 2) • The integrin–ICAM interactions stop leukocytes from rolling; leukocytes spread on the surface of the endothelium • The adhered cells move between adjacent endothelial cells 34 and into the underlying tissue 35 o If The connection of cytoskeletal elements with the ECM is defective, disease conditions may occur The muscular dystrophies •Duchenne muscular dystrophy (DMD) the most common type • cardiac or respiratory failure in young boys • mutations in the gene encoding dystrophin –a cytosolic adapter protein, binding to actin filaments and to an adhesion receptor called dystroglycan (which, in its turn, is connected to laminin) 36