Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
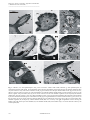
Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ The antennal sensory function in the oldest pterygote insects: an ultrastructural overview M. Rebora, S. Piersanti and E. Gaino Department of Cellular and Environmental Biology, University of Perugia, Via Elce di Sotto 1, 06100 Perugia, Italy Paleoptera (Odonata and Ephemeroptera) represent the oldest pterygote insects. In consideration that antennae are one of the main site of not-visual insect perception, ultrastructural investigations under SEM and TEM have been recently performed on the flagellum of species belonging to several families of Odonata and Ephemeroptera, to clarify the sensory function of the antennae in Paleoptera. These antennae appear very reduced and are constituted by scape, pedicel and an aristate flagellum, mainly monoarticulated in Ephemeroptera and composed of 1-4 flagellomeres in Odonata. Several sensory structures have been identified on the ventro-lateral side of the flagellum in both orders with two main possible functions: thermo-hygroreception and olfaction. Studies on the sensory biology of these aquatic insects can contribute to clarify interesting aspects of their biology. In addition, studies on Paleoptera sensilla light into the evolution of insect sensory abilities. Keywords: sensory biology; thermo-hygroreceptors; olfactory receptors; aquatic insects; Paleoptera; Odonata; Ephemeroptera 1. Introduction Odonata (damselflies and dragonflies) and Ephemeroptera (mayflies) constitute an important component of the freshwater lentic and lotic ecosystems. Both these insect orders develop their larvae in water while the winged adults fly over streams, lakes and small ponds. Dragonflies and damselflies are predators, both as larvae and adults, and many of them are protected in Europe as threatened species, mainly for environmental degradation. Mayflies larvae are algae grazers or detritivorous while the adults do not feed and are short-lived. Many species are important bio-indicators used in European and American monitoring protocols for freshwater ecosystems. Traditionally, Ephemeroptera and Odonata have been classified as Paleoptera (old wings), based on their inability to fold the wings over the abdomen. The remainders of the pterygote insects, able to fold their wings over the abdomen with the inner surfaces facing the latter, constitute the large clade Neoptera (new wings). Together with their inability to fold the wings over the abdomen, Odonata and Ephemerotera share other common features such as big eyes and short, reduced antennae. For this reason these insects have always been considered mainly visual dependent and many studies have been published on their visual abilities while other sensory modalities have been disregarded. In consideration that antennae are one of the main sites of insect perception, we decided to perform ultrastructural investigations under SEM and TEM on the antennal flagellum of some Ephemeroptera and Odonata species [1-4] to clarify the sensory function of these appendixes. This paper reviews such data and reports some additional investigations on other Odonata and Ephemeroptera species belonging to the Italian fauna. This in order to lay the basis for further investigations on the not-visual sensory abilities of these old insects. 2. Material and Methods Mayfly adults of Rhithrogena semicolorata (Curtis, 1834) - Electrogena lateralis (Curtis, 1834) – Ecdyonurus venosus (Fabricius, 1775) (Heptageniidae), Habrophlebia eldae Jacob & Sartori, 1984 (Leptophlebiidae), Baetis rhodani (Pictet 1843) - Cloeon dipterum (Linnaeus, 1761) (Baetidae), Caenis luctuosa (Burmeister) (Caenidae), Siphlonurus lacustris (Eaton) (Siphlonuridae), Serratella ignita (Poda, 1771) (Ephemerellidae), Ephemera danica Müller 1764 (Ephemeridae), dragonfly adults of Libellula depressa Linnaeus - Sympetrum striolatum (Charpenter, 1840) (Libellulidae), Onychogomphus forcipatus (Linnaeus, 1761) (Gomphidae), Aeshna cyanea Müller 1764 (Aeshnidae), Somatochlora metallica Van Der Linden, 1825 (Cordulidae), Cordulegaster boltonii Donovan 1807 (Cordulegastridae) and damselfly adults of Coenagrion puella Linnaeus 1756 - Ischnura elegans (Van Linden,1820), (Coenagrionidae), Platycnemis pennipes (Pallas, 1761) (Platycnemidae), Lestes barbarus Fabricius 1798 - Lestes viridis (Vander Linden 1825) (Lestidae), Calopteryx virgo Linnaeus 1758 - Calopteryx haemorrhoidalis (Van der Linden,1825 (Calopterigidae), were obtained in the laboratory from mature larvae. Mayfly and dragonfly larvae were collected in the Nera River (Perugia, Central Italy) and in a natural pond in Lisciano Niccone (Perugia, Central Italy) during the spring 2007-2008. In the laboratory, the larvae were kept in plastic containers with water, detritus and flora from the collecting site at 25±2°C, LD12:12 h. Dragonfly larvae were fed ad libitum with plankton. ©FORMATEX 2010 137 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ Antennal flagella were dissected from anaesthetised specimens and fixed for 12 hours in 2.5% glutaraldehyde in cacodylate buffer, pH 7.2. For scanning electron microscopy (SEM) analysis, the fixed material, repeatedly rinsed in the same buffer, was then dehydrated by using ethanol gradients, followed by critical-point drying in a critical-point dryer CPD 030 Bal-Tec (BalTec Union Ltd., Balzers, Liechtenstein). Specimens were mounted on stubs with silver conducting paint, sputter-coated with gold-palladium in a sputterer Emitech K550X (Emitech, Ashford, England), and observed with a Philips XL30 (Philips, Eindhoven, Netherlands), at an accelerating voltage of 18kV. For SEM observations of the inner cuticular wall of Odonata antennae, the flagella were longitudinally sectioned with a razor blade and cleaned with KOH-solution. These flagella were then dehydrated in a graded ethanol series, dried in an oven and glued onto the SEM specimen supports in order to allow the observation of the inner side of the antenna. The specimens were then sputter-coated with gold-palladium and observed with a Philips XL30 (Philips, Eindhoven, Netherlands), at an accelerating voltage of 18kV. For transmission electron microscopy (TEM), the fixed antennal flagella were repeatedly rinsed in cacodylate buffer and post-fixed for 1 hour at 4 °C in 1% osmium tetroxide in cacodylate buffer. Afterward the material was repeatedly washed in the same buffer, dehydrated by using ethanol gradients and finally embedded in an Epon-Araldite mixture resin. Ultrathin sections, cut on a Leica EM UC6 ultracut (Leica Microsystem GmbH, Wien, Austria), were collected on formvar-coated copper grids, stained with uranyl acetate and lead citrate, and examined with a Philips EM 208 (Philips, Eindhoven, Netherlands). 138 ©FORMATEX 2010 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ Fig. 1. Odonata (a-c) and Ephemeroptera (d-f) antennae under SEM. (a) Libellula depressa antenna consisting of a scape (S), a pedicel (P) and a flagellum constituted of 4 segments. Arrow points out the sensilla on the latero-ventral side of the flagellum; (b) Lestes viridis antenna consisting of a scape (S), a pedicel (P) and a monoarticulated flagellum (F); arrow points out the sensilla on the latero-ventral side of the flagellum; (c) Detail of the latero-ventral side of the flagellum of Sympetrum striolatum showing sensilla located in pits (arrow); (d) Habrophlebia eldae antenna consisting of a scape (S), a pedicel (P) and a monoarticulated flagellum (F); (e) Siphlonurus lacustris antenna showing the pedicel (P) and the flagellum (F) constituted of numerous segments; (f) Detail of the latero-ventral side of the flagellum of Cloeon dipterum showing sensilla located in pits (arrow) in its proximal portion. ©FORMATEX 2010 139 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ Fig. 2. Odonata (a-b, d-f) and Ephemeroptera (c, g, h) flagellar sensilla under SEM. (a-c) Coeloconic sensilla of Libellula depressa (a), Ischnura elegans (b) and Rhithrogena semicolorata (c) showing pores (P) on their cuticular surface; (d) Openings of the deep cavities in Libellula depressa; (e) Inner cuticular wall vision of the flagellum of Sympetrum striolatum showing the shape of the convoluted deep cavities hosting sensilla styloconica (possible thermo-hygroreceptors); (f) Type-1 (T1) and type-2 (T2) sensilla styloconica located inside the deep cavities in a sectioned convoluted cavity (cleaned with KOH-solution) in the antenna of Onichogomphus forcipatus; (g, h) Possible thermo-hygroreceptors of Siphlonurus lacustris (g) and Habrophlebia eldae (h) showing a more or less conical shape. 3. Results The antennal flagellum in Anisoptera shows a smooth surface and is constituted of 2-4 segments (Fig. 1a) while in Zygoptera it is monoarticulated and shows cuticular scales (Fig. 1b). In both suborders, the flagellum show sensilla located in pits on its latero-ventral side (Fig. 1c). Ephemeroptera have a monoarticulated flagellum (Fig. 1d), which is segmented in some families such as Siphlonuridae (Fig. 1e). The flagellar surface has a texture of cuticular scales (Fig. 1f). As in Odonata, the latero-ventral side of the flagellum shows sensilla located in pits (Fig. 1f). In both the insect Orders the sensory structures are more numerous in the proximal portion or in the more proximal segments of the flagellum. In Odonata (dragonflies and damselflies) and in Ephemeroptera the sensilla located on the latero-ventral side of the flagellum are represented by porous sensilla similar to olfactory sensilla (Figs. 2a-c) and by aporous sensilla whose 140 ©FORMATEX 2010 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ structure recalls that of thermo-hygroreceptors (Figs. 2f-h). The porous sensilla in Odonata are pegs located inside pits (Figs. 2a,b) while in Ephemeroptera they are lobe-shaped pegs, located on the cuticle surface between the scales (Fig. 2c). The possible thermo-hygroreceptors in Odonata are sensilla styloconica (with a sensory peg or cone located on a protrusion or stylus emerging from the cuticle) located inside deep cavities visible from the outside of the antenna as simple openings (Fig. 2d). These cavities are more or less developed and can be very deep and convoluted, especially in Anisoptera (Fig. 2e). Inside the cavities it is possible to distinguish between two kinds of sensilla styloconica, type-1 and type-2 (Fig. 2f). In Ephemeroptera the possible thermo-hygroreceptors are located on the flagellar surface and show a more or less conical shape (Figs. 2g,h). Odonata porous sensilla appear innervated by three unbranched neurons, whose outer dendritic segments enter the peg (Figs. 3a-c). At the base of the peg, the dendrite sheath becomes very thin and tends to disappear, leaving only some traces in the peg (Fig. 3a). The cuticle of the peg shows wide pore-like structures at the base of which the continuity of the cuticle is interrupted by actual pores (Fig. 3b). The space between the pores and the dendrites is narrow and is filled with a material of medium electron-density in which pore tubules are visible (Fig. 3b). Mayfly porous sensilla are innervated by three neurons whose outer dendritic segments enter the peg (Figs. 3d-f). These three neurons are enveloped by the dendrite sheath (Fig. 3f) that opens at the entrance into the peg, leaving the dendrites immersed in the sensillum liquor (Figs. 3d,e). In the peg, the neurons are in contact with the outside by pores present in the cuticle (Figs. 3d,e). Pore tubules are located among pores and dendrites in the sensillum liquor (Fig. 3e). No socket is visible at the insertion of the peg (Fig. 3d). In Odonata, type-1 sensilla styloconica are innervated by four unbranched neurons whose outer dendritic segments are enveloped by the dendrite sheath (Figs. 4a,c). All four dendrites enter the peg, but only three of them reach the apex of the sensillum (Figs. 4b,c); the fourth ends beneath it, where the dendrite sheath is interrupted. The accessory cells are well developed and, around the outer dendritic segments, form long microvilli delimiting numerous electron-lucid vesicles (some species, such as Onichogomphus forcipatus, show an astonishing number of vesicles) (Fig. 4c). At the tip of the cone, the cuticle of the peg is interrupted and a cap-like structure made of amorphous material covers the apex of the dendrites (Figs. 4a and inset,b). From the interrupted cuticle, cuticular fingers emerge to cover the cap-like structure (Figs. 4a and inset,b). No socket and no pores connecting the dendrites with the outside are visible. Type-2 sensilla styloconica are innervated by three unbranched neurons enveloped by three accessory cells (Figs. 4d-f)) producing an abundant electron-dense secretion which is interspersed among their microvilli (Figs. 4d, f). The three outer dendritic segments enter the stylus enveloped by the dendrite sheath. One of the three dendrites stops short of the cone where only two dendrites, closely adherent to each other, are visible (Fig. 4e); these are enveloped by a tightly adherent dendrite sheath which is in close contact with the surrounding cuticle (Fig. 4e). In cross section, the cuticle appears solid in its proximal portion, while it shows round clefts in its distal portion (Fig. 4e). No socket and no connection of the dendrites with the outside are visible. In mayflies the possible thermo-hygroreceptors are innervated by four neurons (Fig. 4i), two of which are unbranched and enter the peg (Fig. 4g,h), while the others stop before entering the peg and one of them is branched. The outer dendritic segments are enveloped by a thick dendrite sheath (Figs. 4g, i). At the base of the peg, the dendrite sheath fuses with the peg cuticle that penetrates deeply inside the sensillum (Fig. 4g). In the peg the dendrites are in close contact with the thick cuticle that shows irregular small clefts (Fig. 4h). There is no connection of the dendrites with the outside in the peg. No socket is evident at the insertion of the peg. In some cases the small pegs are innervated by only two unbranched neurons; the outer dendritic segments of these neurons enter the peg, whose internal structure is like that previously described in the small pegs innervated by four neurons. Both in Odonata and Ephemeroptera antennal flagellum campaniform sensilla are occasionally visible (Figs. 3g,h). The dendrite stops at the base of the cuticle and forms a tubular body in its apical portion (Figs. 3g,h). Above the dendrite, the cuticle is in contact with the dendrite sheath of the tubular body by the socket septum. No difference in the flagellar sensilla number has been so far identified between males and females both in Odonata and in Ephemeroptera. ©FORMATEX 2010 141 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ Fig. 3. Odonata (a-c) and Ephemeroptera (d-f) porous coeloconic sensilla under TEM; Odonata (g) and Ephemeroptera (h) campaniform sensilla under TEM. (a) Longitudinal section showing the dendrites (D) entering the peg; the dendrite sheath becomes very thin and leaves only some traces at the base of the peg (arrow head). Note the pore-like structures on the peg cuticle (arrows); (b) Cross section at the level of the peg. Note the dendrites (D) without dendrite sheath and the pore-like structures (arrows) on the cuticle, together with pore tubules (PT); (c) Cross section at the level of the outer dendritic segments (D) enveloped by the dendrite sheath (DS). Note the accessory cells (AC); (d) Longitudinal section of a peg showing three dendrites (D) immersed in the sensillum liquor; the dendrite sheath (DS) opens at the entrance of the peg leaving the dendrites in contact with the outside by pores (arrow) on the peg cuticle; (e) Cross section of the peg showing dendrites (D), pore tubules (PT) and pores (arrow); (f) Cross section of the outer dendritic segments (D) enveloped by a thick dendrite sheath (DS). AC, accessory cell; (g,h) Longitudinal sections showing the tubular body (TB) at the apex of the dendrite (D). The dendrite sheath (DS) of the tubular body is in contact with the cuticle by the socket septum (SS). 142 ©FORMATEX 2010 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ Fig.4. Odonata (a-f) and Ephemeroptera (g-i) possible thermo-hygroreceptors. (a) Longitudinal section of the peg of a type-1 sensillum showing the dendrites (D) enveloped by the dendrite sheath (DS) entering the peg. AC, accessory cells, CC, ciliary constriction. In the inset the apical region of the peg shows the dendrite (D) with the cap-like structure (Ca) surrounded by cuticular fingers (CF); (b) Cross section of a type-1 sensillum at the level of the apical portion of the peg showing the three dendrites (D) with the cap-like structure (Ca) surrounded by cuticular fingers (CF); (c) Cross section of a type-1 sensillum at the level of the four outer dendritic segments (OD) with the dendrite sheath (DS) surrounded by numerous electron-lucid vesicles (V); (d) Longitudinal section of the peg of a type-2 sensillum showing the dendrites (D), wrapped by the dendrite sheath (DS), entering the cone. AC, accessory cells; (e) Cross section of a type-2 sensillum at the level of the cone showing the two outer dendritic segments (D) entering the peg, enveloped by the dendrite sheath (DS) in close contact with the surrounding cuticle with round clefts (arrow) in its distal portion; (f) Cross section of a type-2 sensillum at the level of the outer dendritic segments (OD) wrapped by the dendrite sheath (DS) and the accessory cells (AC); (g) Longitudinal section of the peg showing the peg cuticle (C) that penetrates deeply into the sensillum and fuses with the dendrite sheath (DS) enveloping the dendrites (D); (h) Cross section of a peg showing two dendrites (D) in close contact with the peg cuticle (C); note the irregular clefts of the cuticle (arrow); (i) Cross section at the level of the four outer dendritic segments (D) enveloped by the dendrite sheath (DS) and the accessory cells (AC). ©FORMATEX 2010 143 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ 4. Discussion On the ventro-lateral side of the antennal flagellum of Ephemeroptera and Odonata, sensilla have been identified in all the examined species representatives of the main Italian families [1-4 and the data presented in this paper]. Notwithstanding the differences in shape and location, these sensilla share relevant common features in the two insect orders: they are more concentrated on the proximal portion of the unarticulated flagella, or on the proximal segments in the articulated ones; most of them tend to be short, closely adherent to the flagellar surface, located inside pits, and can be classified as sensilla coeloconica (= pit peg). The presence of coeloconic sensilla in Paleoptera antennae is particularly interesting owing to the ancient origin of this kind of sensilla in insects [5,6]. The external and internal cuticular morphology of the flagellar sensilla of both Odonata and Ephemeroptera is compatible with interpreting them as single-walled olfactory sensilla [7,8] and thermo-hygroreceptors [see review in 911]. Occasionally campaniform sensilla are visible on the dorsal antennal surface, and presumably senses the stretching movements of the flagellum. The importance of hygroreceptors perceiving changes in humidity is obvious in both Insect Orders being aquatic insects: they must detect humidity gradient in air to find the adequate laying places. It is well known that Odonata and Ephemeroptera can “see” the presence of water, as many aquatic insects, on the basis of the horizontally polarized light reflected from the water surface [12-14]. Indeed, the ventral region of the eye in many aquatic insects is sensitive to the polarization of light in the visible and/or ultraviolet spectral ranges [15,16]. Polarised light reflected by water can aid the orientation of these insects from a distance where other cues are still ineffective [17]. Evidently, as reported by Corbet [18], in Odonata the more general cues are detected visually, and the final, more specific cues by other sensory modalities. At a closer range, indeed, hygroreception could have an important role in Odonata and Ephemeroptera habitat selection and oviposition. Insect hygroreceptors are typically associated with thermoreceptors in peculiar sensilla mainly located on the antennae [see review in 9-11]. As far as thermoreceptors are concerned, the ability to perceive changes in temperature is fundamental in Odonata, which rely on direct sunshine for their thermoregulation [19], and in the fragile adult mayflies, able to perform mating flights only in a peculiar temperature range [14]. The importance of olfactory receptors in Paleoptera is more questionable. Indeed neuroanatomical studies on insect brain have prompted researchers to hypothesize that the extant paleopteran insects are probably all anosmic with respect to airborne odours; indeed, they lack glomerular antennal lobes, which typically receive olfactory receptor neurons in Neoptera [20,21]. A recent neuroanatomical study on the ground plan of the insect mushroom body suggested that the simplicity of these structures in mayflies may not be entirely attributable to a primitive character state, but rather related to the highly derived lifestyle of Ephemeroptera, with aquatic larvae and short non-feeding adults, in which olfaction is redundant [22]. Olfaction could be considered redundant also in Odonata, where vision plays an important role for the adult behaviour [18], but our results suggest that mayflies, dragonflies and damselflies could perceive odours and could use these ability for some aspects of their biology. To understand the function of the antennal sensilla of paleopterans, further molecular, electrophysiological, neuroanatomical and behavioural studies are in progress. Such studies on paleopteran sensory abilities could add new important data to trace phylogenetic relationships between the two insect orders and could shed light onto the evolution of insect olfaction, a research field so far in a fairly initial phase. 5. References [1] Rebora M, Piersanti S, Gaino E. The antennal sensilla of the adult of Libellula depressa (Odonata: Libellulidae). Arthropod Structure and Development. 2008;37: 504-510. [2] Rebora M, Piersanti S, Gaino E. A comparative investigation on the antennal sensilla of adult Anisoptera. Odonatologica. 2009a;38(4): 329-340. [3] Rebora M, Piersanti S, Gaino E. The antennal sensilla of adult mayflies: Rhithrogena semicolorata as a case study. Micron. 2009b;40: 571-576. [4] Piersanti S, Rebora M, Gaino E. A scanning electron microscope study of the antennal sensilla of adult damselflies (Odonata: Zygoptera). Odonatologica. 2010; in press. [5] Steinbrecht RA. Pore structures in insect olfactory sensilla: a review of data and concepts. International Journal of Insect Morphology & Embryology. 1997;26(3/4): 229-245. [6] Yao CA, Ignell R, Carlson JR. Chemosensory Coding by Neurons in the Coeloconic Sensilla of the Drosophila Antenna. The Journal of Neuroscience. 2005;25(37):8359–8367 [7] Altner H. Insect sensillum specificity and structure: An approach to a new typology. In: Le Magnen J, Mac Leod P, eds. Olfaction and Taste VI. Information Retrieval. London, 1977; pp. 295-303. [8] Altner H, Prillinger L. Ultrastructure of invertebrate chemo-, thermo- and hygroreceptors and its functional significance. International Review of Cytology. 1980;67:69-139. [9] Altner H, Loftus R. Ultrastructure and function of insect thermo- and hygroreceptors. Annual Review of Entomology. 1985;30:273-295. [10] Tichy H, Loftus R. Hygroreceptors in insects and a spider: Humidity transduction Models. Naturwissenschaften. 1996;83:255-263. 144 ©FORMATEX 2010 Microscopy: Science, Technology, Applications and Education A. Méndez-Vilas and J. Díaz (Eds.) ______________________________________________ [11] Yokohari F.. Hygro- and thermoreceptors. In: Eguchi E, Tominaga Y eds. Atlas of Arthropod sensory receptors: dynamic morphology in relation to function. Springer, Berlin, 1999; pp.191-210. [12] Horvath G. Reflection–polarization patterns at flat water surfaces and their relevance for insect polarization vision. Journal of theoretical Biology. 1995;175:27–37. [13] Horvath G, Bernath B, Molnar G. Dragonflies find crude oil visually more attractive than water: multiple-choice experiments on dragonfly polarotaxis. Naturwissenschaften. 1998;85: 292-297. [14] Kriska G, Horváth G, Andrikovics S. Why do mayflies lay their eggs en masse on dry asphalt roads? Water-imitating polarized light reflected from asphalt attracts Ephemeroptera. The Journal of Experimental Biology. 1998;201: 2273–2286. [15] Schwind R. Polarization vision in water insects and insects living on a moist substrate. Journal of Comparative Physiology A. 1991;169:5319. [16] Schwind R. Spectral regions in which aquatic insects see reflected polarized light. Journal of Comparative Physiology A.1995;177:439-448. [17] Bernáth B, Szedenics G, Wildermuth H, Horváth G, How can dragonflies discern bright and dark waters from a distance? The degree of polarization of reflected light as a possible cue for dragonfly habitat selection, Freshwater Biology 2002;47:17071719. [18] Corbet PS. Dragonflies behaviour and ecology of Odonata. Colchester: Harley Books; 1999. [19] May ML. Thermoregulation and adaptation to temperature in dragonflies (Odonata: Anisoptera). Ecology Monographs. 1976;46:1-32. [20] Farris SM. Evolution of insect mushroom bodies: old clues, new insights. Arthropod Structure and Development. 2005;34:211-234. [21] Strausfeld NJ, Hansen L, Li Y, Gomez RS, Ito K.. Evolution, discovery, and interpretations of Arthropod mushroom bodies. Learning Memory. 1998;5:11-37. [22] Strausfeld NJ, Sinakevitch I, Brown S, Farris S. Ground plan of the insect mushroom body: functional and evolutionary implications. Journal of Comparative Neurology. 2009;513: 265-291. ©FORMATEX 2010 145