Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
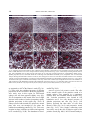
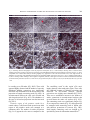
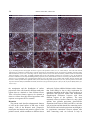
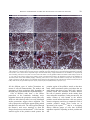
Indian Journal of Experimental Biology Vol. 48, April 2010, pp. 346-353 Modulation of cell surface architecture in gastrulating chick embryo in response to altered fibroblast growth factor (FGF) signaling Seema Borgave & Surendra Ghaskadbi* Division of Animal Sciences, Agharkar Research Institute, Pune 411 004, India Received 15 October 2009 Gastrulation is a fundamental process that results in formation of the three germ layers in an embryo. It involves highly coordinated cell migration. Cell to cell communication through cell surface and the surrounding molecular environment governs cell migration. In the present work, cell surface features, which are indicative of the migratory status of a cell, of an early gastrulating chick embryo were studied using scanning electron microscopy. The distinct ultrastructural features of cells located in the various regions of the epiblast are described. Differences in the surface features of cells from distinct embryonic regions indicate differences in their migratory capacities. Further, the dynamic nature of these cell surface features by their response to altered fibroblast growth factor (FGF) signaling, experimentally created by using either excess FGF or inhibition of FGF signaling are demonstrated. Keywords: Altered FGF signaling, Cell surface architecture, Chick embryo In amniotes the epiblast cells ingress through the developing primitive streak and give rise to mesodermal and endodermal cells during gastrulation. Gastrulation involves coordinated movements of cells either individually or as cell sheets1 and cell-cell communication is crucial during such migratory processes2. Cells directly communicate with one another through their cell surface; this often decides their respective path of differentiation3. The threedimensional cellular architecture as well as the surface features of individual cells can be studied with scanning electron microscopy (SEM). In spite of being one of the primary processes, gastrulation is not completely understood in amniotes. The organization and surface architecture of cells of early gastrulating chick embryos has been described4-9. These studies point towards differences in shape and migration rates of cells located within and outside the primitive streak, probably due to the differences in requirement for moving inside through the streak and lateral movements between ectoderm and endoderm, postinvagination. In some of these reports mesodermal and endodermal cells from ventral surface have been studied using scanning electron microscopy (SEM)6-8. However, there are only a few studies wherein the ________________ *Correspondent author Telephone: +91-20-25653680, 25654357 (Ext. 278) Fax: +91-20-25651542 E-mail: [email protected] dorsal surface i.e. epiblast of the early chick embryos is described4,5. Study of dorsal surface is crucial for understanding the migratory potentials of epiblast cells that eventually ingress through the primitive streak from dorsal side, undergo epithelial to mesenchymal transition and move ventrally to form either mesoderm or endoderm. Earlier, Bancroft and Bellairs4 have studied the surface features of epiblast cells of embryos from un-incubated egg up to 30 h of incubation. On the other hand, Jacob et al.5 have described the morphological features of epiblast cells from prospective neural plate, anterior primitive streak and lateral prismatic region. In the present study the differences in the ultrastructural features of the epiblast cells from four developmentally significant regions of an early chick embryo, viz, Hensen’s node, prospective neural plate, anterior and posterior regions of the primitive streak have been studied. To our knowledge, such a detailed account of cell surface morphology from discrete embryonic areas does not exist in literature. Altered fibroblast growth factor (FGF) signaling causes abnormal development of nervous system and mesodermal structures in early chick embryo have been demonstrated10. The abnormal development was associated with rapid modulation of expression of brachyury, noggin10 and goosecoid11, genes which are essential for normal development of the nervous system and mesodermal structures. Brachyury12 and BORGAVE & GHASKADBI: ALTERED FGF SIGNALING & CELL SURFACE CHANGES goosecoid13 encode transcription factors that are required for the gastrulation movements and differentiation of mesoderm. Appropriate levels of FGFs are thought to be essential for the normal morphogenesis of vertebrate embryos14. Hence it is likely that the effects of altered FGF signaling in early chick embryo10,11 are reflected at the level of cell surface. In the present work, the effects of altered FGF signaling have been studied on the surface features of embryos that reflect the migratory status of cells. Also the dynamic nature of the cell surface in four important embryonic regions have been described. Materials and Methods Freshly laid White Leghorn chicken eggs were obtained from a local hatchery. Human recombinant bFGF and suramin were procured from Sigma (St. Louise, USA). In vitro culture and treatment of chick embryo explants⎯Eggs were incubated for 18 h at 37.5°C to obtain Hamburger Hamilton15 (HH) stage 4 chick embryos. Embryos were dissected in Pannet-Compton saline16 (PC saline, pH 7.4), cultured in vitro by New's single ring technique17 and staged on the basis of standard morphological criteria15. The cultures were randomly divided into 4 groups. Embryos were treated as described before10,11,18. Two controls, treated either with PC saline alone or with PC saline containing BSA, were maintained. Thus, embryos were treated with either PC saline, BSA (100 ng/culture), 6 nM bFGF (10 ng/culture) or 2 mM suramin (200 nmoles/culture) as described earlier10,11. After addition of the test chemical the embryos were left at room temperature for 30 min to allow proper diffusion of the chemical and then incubated at 37.5°C for 2 h. These embryos were fixed for SEM. All experiments were carried out at least in triplicate. Scanning electron microscopy⎯At the end of 2 h, the embryos were processed for SEM as described before9,19. Embryos were fixed in 2.5% glutaraldehyde (pH. 7.4) for 24 h at 4°C. Embryos were washed with PBS to remove traces of glutaraldehyde, dehydrated in a graded series of ethanol and transferred to amyl acetate gradually. Amyl acetate was replaced with liquid CO2 in a E3000 critical point drier (Bio-Rad). Three cycles of critical point drying were carried out to ensure complete drying. Embryos were mounted flat with their dorsal side up on metal stubs, coated with gold in E5200 347 automatic sputter coater (Bio-Rad), scanned with a Stereoscan S120 SEM (Cambridge Instruments) and photographed. A minimum of 15 representative embryos were scanned from each group. Results Cells from the Hensen’s node, prospective neural plate, anterior and posterior regions of the primitive streak of stage 4 chick embryos (Fig. 1) were studied by SEM. Simultaneously, alterations in the cellular arrangement and cell surface features after 2 h of treatment with either exogenous FGF or suramin were studied. Hensen’s node⎯Cells of the Hensen’s node of a control embryo were arranged in a uniform sheet (Fig. 2A-C). Cells exhibited various types of surface projections. Vesicular projections described earlier as ‘glass-bell-like formations’20, appeared as beads along the cell boundaries. The cells also exhibited numerous globular projections and a few small, thin thread-like connecting cords (Fig. 2B, C). At the end of 2 h, FGFtreated embryos exhibited distorted cell surface features (Fig. 2D-F). The cells appeared to be crawling over each other and surface features were not clearly discernible. The cells exhibited varied sizes and shapes with more connecting cords as compared to controls (Fig. 2A-C). In suramin-treated embryos, the sheet of cells in Hensen’s node appeared wavy (Fig. 2G). Surface protrusions increased to such an extent that the cell surfaces were barely visible. The cells were covered with globular projections (Fig. 2H, I). The surface protrusions were abnormal; they were shorter, ended in bulb like structures and appeared hollow which have been described as ‘bowllike projections’5 (Fig. 2I). Prospective neural plate⎯In a control embryo the cells of the prospective neural plate were arranged as a uniform sheet (Fig. 3A-C). These cells possessed Fig. 1⎯Diagrammatic representation of HH stage 4 embryo showing the regions that were scanned for cell surface ultrastructure studies. (A) Hensen's node, (B) prospective neural plate, anterior (C) and posterior (D) parts of the primitive streak. 348 INDIAN J EXP BIOL, APRIL 2010 Fig. 2⎯Scanning electron micrographs of cells of Hensen's node. HH stage 4 chick embryos were incubated in presence of either 6 nM FGF or 2 mM suramin for 2 h at 37ºC and processed for scanning electron microscopy. (A) A control embryo showing uniform sheet of cells. (B,C) Magnified views of cells from the same region of (A). (B) Note vesicular projections at cell boundaries (hollow arrow), globular projections (arrow) and small thread like connecting cords (half-filled arrowhead). (D-F) Comparable area of FGF-treated embryos showing distorted surface features of cells. (E) The cells appear to crawl over each other and exhibit variation in size and shape (asterisk). Note increased abundance of connecting cords (half-filled arrowhead in E) and globular protrusions (arrow in F). (G-I) Cells of the Hensen's node region of suramin-treated embryo. Note wavy nature of the cell sheet (G) and significant increase in the abundance of small connecting cords (half-filled arrowhead in H) and globular projections (arrow in I). (I) bowl-like surface projections are also seen (hollow arrow). Scale bar in (G) represents 20 μm (for A,D,G), in (H) 10 μm (for B,E,H) and in (I) 5 μm (for C,F,I). more number of lengthy thread-like connecting cords as compared to cells of the Hensen’s node (Fig. 2AC). These cells also exhibited presence of globular projections on the cell surface. Bowl-like projections were rarely seen in this region. In FGF-treated embryos, the cell sheet appeared slightly wavy (Fig. 3D). In comparison to the control, there was a slight increase in the number of surface protrusions, mainly globular projections in this region (Fig. 3D-F). In embryos treated with suramin, the prospective neural plate cells were not arranged in one plane but exhibited an undulating pattern (Fig. 3G). Cells were irregular in shape with uneven surfaces and appeared to be crowded (Fig. 3G,H). The abundance of bowl- like projections was more than controls (Fig. 3H,I, for control Fig. 3B,C). Anterior region of the primitive streak⎯The cells of the anterior region of the primitive streak of a control embryo were arranged in a continuous uniform sheet and exhibited smooth surfaces (Fig. 4A). Vesicular protrusions were located at cell boundaries and a few cells showed presence of globular projections and cilia (Fig. 4A-C). Cell surfaces appeared flat than those of cells from Hensen’s node (Fig. 2A-C) and prospective neural plate (Fig. 3A-C). Also, lengthy connecting cords were rarely seen in the cells of anterior region of the primitive streak (Fig. 4B,C). In FGF-treated embryos, BORGAVE & GHASKADBI: ALTERED FGF SIGNALING & CELL SURFACE CHANGES 349 Fig. 3⎯Scanning electron micrographs of cells of prospective neural plate (A-C) A control embryo showing cells as a uniform sheet. Cellular projections are mainly in the form of connecting cords (half-filled arrowhead in B) and globular projections (arrow in C). (D-F) Comparable area of FGF-treated embryos showing slightly wavy appearance of the cell sheet (asterisk in D). Note slight increase in abundance of connecting cords (half-filled arrowhead in E) and small globular projections (arrow in F). (G-I) A suramin-treated embryo. (G) Note crowding of cells resulting in their irregular arrangement (asterisk). Cells appear with uneven surfaces (half-filled arrowhead in H) with bowl-like protrusions (arrow in I). Scale bar in (G) represents 20 μm (for A,D,G), in (H) 10 μm (for B,E,H) and in (I) 5 μm (for C,F,I). the cells were arranged in a wavy sheet and seemed to be crawling over each other (Fig. 4D-F). These cells appeared highly distorted and the number of unevenly distributed globular protrusions was significantly increased. FGF treatment led to an increase also in the proportion of lengthy connecting cords (Fig. 4E,F). In suramin-treated embryos the cells were crowded and with irregular surfaces (Fig. 4G-I). The abundance of all types of cell surface projections was also increased (Fig. 4G-I). Posterior region of the primitive streak⎯In a control embryo, cells located in the posterior one third region of the primitive streak were arranged as a continuous sheet. The number of surface projections (Fig. 5A-C) appeared to be more as compared to the anterior region of the primitive streak (Fig. 4A-C). The connecting cords in this region were more lengthy than any other embryonic region. These cells also exhibited presence of globular projections and microvilli. In FGF-treated embryos, globular projections were slightly more abundant (Fig. 5D-F). There was a considerable increase in the number of surface protrusions in suramin-treated embryos that covered almost the entire cell surfaces (Fig. 5G-I). The connecting cords were significantly longer (Fig. 5I) as compared to controls (Fig. 5C) and appeared to be entangled with connecting cords of distant cells with tuberous protrusions in between (Fig. 5H,I). Such lengthy connecting cords are thought to be specialized systems for information transmission4,5. 350 INDIAN J EXP BIOL, APRIL 2010 Fig. 4⎯Scanning electron micrographs of anterior region of the primitive streak (A-C) A control embryo. Note cells with smooth surfaces that are arranged in a uniform sheet. Vesicular projections at cell boundaries (arrow in B) and a few globular protrusions (halffilled arrowhead in C) are seen. Connecting cords are rarely observed. (D-F) Cells from the anterior primitive streak of FGF-treated embryo arranged in a wavy sheet. (E) Highly distorted shape of the cells and their crawling over each other is seen. (E) Significant increase in abundance of unevenly distributed globular projections (arrow) and localized increase in connecting cords (hollow arrow) are observed. (G-I) Comparable area of suramin-treated embryos. (H) Cells appear to be crowded with irregular surfaces (asterisk) and increase in surface protrusions like globular bodies (arrow) and connecting cords (hollow arrow) is seen. (I) Note presence of bowl-like projections in these embryos (half-filled arrowhead). Scale bar in (G) represents 20 μm (for A,D,G), in (H) 10 μm (for B,E,H) and in (I) 5 μm (for C,F,I). Overall, these results show distinct differences in the arrangement and the distribution of surface projections of the cells located in different embryonic regions that are destined to form different tissues. These cell surface features appear to be dynamic in nature as evident from their altered appearance after exposure to exogenous FGF or suramin. Discussion The present work describes ultrastructural features of cells on the dorsal surface of HH stage 4 chick embryo. Cells of the Hensen's node, prospective neural plate and anterior and posterior regions of the primitive streak, which contribute to different tissues of the embryo, were studied. The cells from different embryonic regions exhibited distinct surface features that could mainly be due to their requirement for migration depending on their fate. These results are in agreement with previous studies that demonstrate morphological differences between cells from different regions of the epiblast4,5. The most commonly observed surface features in early chick epiblast were globular projections, glass-bell-like formations in the form of either bowl-like or vesicular cell projections and thin thread like connecting cords. The bowl-like and vesicular projections are thought to be cytoplasmic connections between adjacent cells while connecting cords are believed to help in BORGAVE & GHASKADBI: ALTERED FGF SIGNALING & CELL SURFACE CHANGES 351 Fig. 5⎯Scanning electron micrographs of cells located in the posterior region of the primitive streak. (A-C) A control embryo showing cells arranged in a continuous sheet. (B) Note presence of globular projections (hollow arrow), microvilli and long thread like connecting cords (arrow). (D-F) Cells from the comparable region of FGF-treated embryo. Slight enhancement in the abundance of comparatively short connecting cords (arrow in E) that cover most of the cell surface in F. The globular bodies look swollen in these embryos (arrow in F). (G-I) Cells of the posterior region of the primitive streak of suramin-treated embryo. (H) Significant increase in the abundance of microvilli (hollow arrow) and extremely lengthy connecting cords (arrow) is seen. (H,I) Cell surface is barely seen. Note presence of lengthy connecting cords that appear to be entangled with projections from neighboring cells with tuberous structures in between (arrow in H,I). Scale bar in (G) represents 20 μm (for A,D,G), in (H) 10 μm (for B,E,H) and in (I) 5 μm (for C,F,I). communication between cells at longer distances4,5. All the different types of surface protrusions are means of cell-cell communication. The number and appearance of these projections differ depending on the embryonic region and the developmental stage4,5. Cells of Hensen's node show a few surface protrusions at the boundaries indicating active migration whereas the prospective neural plate cells, which are with relatively more but evenly distributed surface protrusions suggest slower migration. The cells of prospective neural plate appear tightly packed than Hensen’s node cells. The cells ingressed through the Hensen’s node move anteriorly to form the foregut, head process and notochord21. Cells from the anterior region of the primitive streak possess the least number of surface protrusions. Cells of the posterior region of the primitive streak, on the other hand, exhibit maximum surface projections that are much longer than those of any other cells. Anterior primitive streak cells contribute to somites whereas the cells of posterior primitive streak mainly form extra-embryonic mesoderm22. In the present study, the surface features of cells located in the anterior primitive streak indicate that these cells may possess extensive migratory activities as compared to cells of posterior primitive streak. These results are in agreement with reports4,5,23,24 that in a developing embryo different groups of cells have different migration capacities depending on their position and fate. 352 INDIAN J EXP BIOL, APRIL 2010 Endogenous FGF levels were altered with addition of either exogenous FGF or suramin. Although suramin affects a variety of growth factors, it predominantly inhibits FGF action10,11,25-29. FGF is thought to act through genes like brachyury10 and goosecoid11. Brachyury is involved in induction and differentiation of mesoderm12 whereas goosecoid regulates gastrulation movements and formation of body axis13. Modulation of expression of both the genes within 2 h in response to altered FGF signaling was obseved. FGF signaling is indeed necessary for cell movements during streak formation in chick embryo30. Present results indicate that altered FGF signaling may hamper cellular movements during gastrulation to varying extent in distinct embryonic areas. FGF is believed to coordinate gastrulation movements14 through non-canonical Wnt11 signaling pathway31, Wnt3a signaling pathway and by maintaining Ecadherin levels32. FGF-treated embryos exhibited down-regulation of brachyury expression in the primitive streak10. The observations of the present study suggest that anterior region of the primitive streak is the active site for ingression of epiblast cells in early chick embryos. Crowding of cells observed in this region of FGF-treated embryos may be due to lack of sufficient amounts of brachyury as Brachyury is essential for cells to undergo convergent extension movements12. On the other hand, a few suramintreated embryos showed absence of brachyury transcripts10 and presence of abnormally lengthy connecting cord like projections in the posterior onethird region of the primitive streak. These observations suggest that cells from this region may have lost their ability to migrate. The surface architecture of cells of Hensen's node was also differentially altered due to FGF and suramin. Differential modulation of goosecoid expression was observed in the Hensen's node due to FGF and suramin within 2 h11. The altered cell surface features together with modulated expression of brachyury and goosecoid may lead to improper cellular movements during gastrulation and result in abnormal morphogenesis of embryos treated with FGF and suramin. The present work demonstrates the distinct differences in the migratory capacities of cells from various embryonic regions as indicated by cell surface architecture and dynamic changes in these ultrastructural features in response to altered FGF signaling. Earlier it has been shown that embryos with altered levels of FGF signaling develop abnormal neural and mesodermal structures10. Collectively, these studies shows that the abnormal development in FGFand suramin-treated embryos is not only accompanied by but appears to be an outcome of the rapid modulation of brachyury10 and goosecoid11 as well as cell surface architecture. Acknowledgement Thanks are due to Drs. Saroj Ghaskadbi, Hemant Ghate and Vidya Patwardhan for discussions and Mr. Rajdeep Dongre for technical help with SEM. This work was supported by Agharkar Research Institute, Pune. References 1 Mittenthal J E & Jacobson A G, The mechanics of morphogenesis in multicellular embryos, in Biomechanics of active movement and deformation of cells (Spring-Verlag, Berlin, Heidelberg) 1990, 295. 2 Gilbert S F, Developmental biology, seventh edition (Sinauer Associates, Inc., Sunderland, Massachusetts) 2003. 3 Fraser S E, Green C R, Bode H R & Gilula N B, Selective disruption of gap junctional communication interferes with a patterning process in hydra, Science, 237 (1987) 49. 4 Bancroft M & Bellairs R, The onset of differentiation in the epiblast of the chick blastoderm (SEM and TEM), Cell Tiss Res, 155(1974) 399. 5 Jacob H J, Christ B, Jacob M & Bijvank G J, Scanning electron microscope (SEM) studies on the epiblast of young chick embryos, Z Anat Entwickl Gesch 143 (1974) 205. 6 England M A & Wakely J, Scanning electron microscopy of the development of the mesoderm layer in chick embryos, Anat Embryol, 150 (1977) 291. 7 Wakely J & England M A, Development of the chick embryo endoderm studied by S.E.M., Anat Embryol, 153 (1978) 167. 8 Critchley D R, England M A, Wakely J & Hynes R O, Distribution of fibronectin in the ectoderm of gastrulating chick embryos, Nature, 280 (1979) 498. 9 Karandikar A & Ghaskadbi S, β-Microseminoprotein-related molecules may participate in formation of the mesoderm in the chick embryo, Dev Growth Diff, 45 (2003) 309. 10 Khot S & Ghaskadbi S, FGF signaling is essential for the early events in the development of the chick nervous system and mesoderm, Int J Dev Biol, 45 (2001) 877. 11 Borgave S & Ghaskadbi S, Fibroblast growth factor regulates early mesoderm and neural development in chick embryo through its action on brachyury, goosecoid, ERNI and noggin, Curr Sci, 96 (2009) 1217. 12 Conlon F L & Smith J C, Interference with Brachyury function inhibits convergent extension, causes apoptosis, and reveals separate requirement in FGF and activin signaling pathways, Dev Biol, 213 (1999) 85. 13 Izpisúa-Belmonte J C, De Robertis E M, Storey K G & Stern C D, The homeobox gene goosecoid and the origin of organizer cells in the early chick blastoderm, Cell, 74 (1993) 645. BORGAVE & GHASKADBI: ALTERED FGF SIGNALING & CELL SURFACE CHANGES 14 Yamaguchi T P, Conlon R A & Rossant J, Expression of the fibroblast growth factor receptor FGFR-1/flg during gastrulation and segmentation in the mouse embryo, Dev Biol, 152 (1992) 75. 15 Hamburger V & Hamilton H L, A series of normal stages in the development of the chick, J Morphol, 88 (1951) 49. 16 New D A T, The culture of vertebrate embryos (Logos Press, London) 1966, 47. 17 New D A T, A new technique for the cultivation of chick embryo in vitro, J Embryol Exp Morphol, 3 (1955) 326. 18 Ghaskadbi S & Mulherkar L, Effects of cytochalasin H on chick embryo explants cultured in vitro, Toxicology, 33 (1984) 323. 19 Patwardhan V & Ghaskadbi S, Activin disrupts somitogenesis in cultured chick embryos, Dev Genes Evol, 211 (2001) 478. 20 Klika E & Jelinek R, “Glass-Bell-Like” formations as modified cellular junctions in the developing neural tube and ectoderm of the chick embryo, Folia Morphol (Praha), 19 (1971) 137. 21 Schoenwolf G C, Garcia-Martinez V & Dias M S, Mesoderm movement and fate during amphibian gastrulation and neurulation, Dev Dyn, 193 (1992) 235. 22 Smith J L & Schoenwolf G C, Getting organized: New insights into the organizer of higher vertebrates, Curr Top Dev Biol, 40 (1998) 79. 23 Keller R E, Shih J & Sater A, The cellular basis of convergence and extension of the Xenopus neural plate, Dev Dyn, 193 (1992) 199. 353 24 Psychoyos D & Stern C D, Fates and migratory routes of primitive streak cells in the chick embryo, Development, 122 (1996) 1523. 25 Aoki T, Kato S, Fox J C, Okamoto K, Sakata K, Morimatsu M & Shigemori M, Inhibition of autocrine fibroblast growth factor signaling by the adenovirus-mediated expression of an antisense transgene or a dominanat negative receptor in human glioma cells in vitro, Int J Onco, 21 (2002) 629. 26 Palamakumbura A H, Sommer P & Trackman P C, Autocrine growth factor regulation of lysyl oxidase expression in transformed fibroblasts, J Biol Chem, 278 (2003) 30781. 27 Mitrani E, Gruenbaum Y, Shohat H & Ziv T, Fibroblast growth factor during mesoderm induction in the early chick embryo, Development, 109 (1990) 387. 28 Riese J, Zeller R & Dono R, Nucleo-cytoplasmic translocation and secretion of fibroblast growth factor-2 during avian gastrulation, Mech Dev, 49 (1995) 13. 29 Cirillo A, Chifflet S & Villar B, Neural retina of chick embryo in organ culture: effects of blockade of growth factors by suramin, Cell Tissue Res, 304 (2001) 323. 30 Chuai M, Zeng W, Yang X, Boychenko V, Glazier J A & Weijer C J, Cell movement during chick primitive streak formation, Dev Biol, 296 (2006)137. 31 Tada M, Concha M L & Heisenberg C-P, Non-canonical Wnt signaling and regulation of gastrulation movements, Seminars in Cell and Developmental Biology, 13 (2002) 251. 32 Ciruna B & Rossant J, FGF signaling regulates mesoderm cell fate specification and morphogenetic movements at the primitive streak, Dev Cell, 1 (2001) 37.