Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
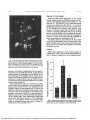
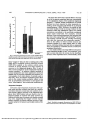
Investigative Ophthalmology & Visual Science, Vol. 30, No. 3, March 1989 Copyright © Association Tor Research in Vision and Ophthalmology Sensory and Sympathetic Innervation of the Mammalian Cornea A Retrograde Tracing Study Carl F. Marfurr,* Robert E. Kingsley,t oncl Stephen E. Echtenkamp^: The method of retrograde transport of horseradish peroxidase-wheat germ agglutinin (HRP-WGA) was used to study the locations, numbers and somata diameters of cornea! afferent and efferent neurons in four different mammalian species. A 2% solution of HRP-WGA was applied to the central cornea of the rat, rabbit, cat and monkey and the animals were perfusion-fixed 48-96 hr later. HRP-WGA-labeled sensory neurons were distributed relatively uniformly throughout the ophthalmic region of the ipsilateral trigeminal ganglion of the rat, cat and monkey. In marked contrast, labeled cells in the rabbit trigeminal ganglion were clustered in a sharply defined longitudinal column located in the midregion of the ophthalmic area. Occasional cells in some cats and monkeys were observed near, and possibly within, the maxillary region of the ganglion. There was no evidence for a dorsoventral somatotopy of corneal afferent neurons in any species. The majority of the labeled afferent somata were small or medium in size, although some larger diameter neurons were also observed. Modest numbers of labeled neurons were observed in the ipsilateral superior cervical ganglion (SCG) of the rabbit and cat; however, only occasional labeled neurons were observed in the SCG of the rat, and none were seen in the monkey. The labeled SCG cells, when present, were concentrated in the rostral half of the ganglion, although many cells in the cat SCG were also found further caudally. No labeled neurons were found in the middle cervical or stellate ganglia of any animal. The results of this study have revealed the existence of subtle interspecies differences in the organization of the mammalian corneal afferent and efferent innervations. Invest Ophthalmol Vis Sci 30:461-472,1989 modify corneal sensitivity. 7 The mechanism by which corneal nerves exert these influences is unknown, although the release of axonally transported, biologically active substances such as peptides and neurotransmitters may be involved. Information about the cell bodies that give origin to these different components of the corneal innervation is limited. The corneal sensory fibers in mice, rats, cats and monkeys originate from neurons located, with rare exceptions, in the ophthalmic (medial) region of the ipsilateral trigeminal ganglion.8"13 However, the distribution of corneal afferent somata in the rabbit, a popular animal model in ophthalmological research, has not been described. In addition, there is little information concerning the relative numbers of sensory cells that contribute to the corneal innervation in different species. While it is widely reported that comeal afferent perikarya are small or medium in size, the cell size spectra for these neurons' 4 1 5 in most species remain unknown. Knowledge of the origin(s) of the corneal sympathetic innervation is even more fragmentary,716 and analysis of the limited available data suggests that there may be significant interspecies differences. The mammalian cornea is richly supplied by sensory nerve fibers from the trigeminal ganglion (TG) and, to a lesser extent, by sympathetic fibers from the superior cervical ganglion (SCG). Both populations of nerve fibers play unique roles in maintaining the overall structural and functional integrity of the cornea. Corneal afferent nerves, in addition to signalling sensory information, exert important trophic influences on the corneal epithelium 1 and stimulate wound healing following epithelial lesions.2 Corneal sympathetic nerves, in turn, regulate epithelial ion transport processes,3 exert inhibitory influences on epithelial mitogenesis and wound healing4"6 and may From the Departments of *Anatomy and ^Physiology, Northwest Center for Medical Education, Indiana University School of Medicine, Gary, Indiana, and the fDepartment of Physiology, South Bend Center for Medical Education, Indiana University School of Medicine, University of Notre Dame, Notre Dame, Indiana. Supported in part by PHS grants EY-05717 to CFM and HL37223 to SFE. Submitted for publication: June 24, 1988; accepted October 5, 1988. Reprint requests: Dr. Carl F. Marfurt, Northwest Center for Medical Education, 3400 Broadway, Gary, IN 46408. 461 Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 462 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / March 1989 In the current investigation, we have used the method of retrograde transport of horseradish peroxidase-wheat germ agglutinin (HRP-WGA) to study the afferent and efferent corneal innervation of four mammalian species commonly used in ophthalmological research: the rat, rabbit, cat and monkey. The specific goals of the current study were to determine: (1) the distribution within the trigeminal and superior cervical ganglia of corneal sensory and sympathetic nerve cell bodies; (2) the relative numbers of trigeminal and SCG cells that contribute to the corneal innervation in different species; and (3) the somata diameters of the corneal afferent neurons. The data generated from this study will increase our understanding of the anatomy of the mammalian corneal innervation and will provide a useful database of information for future studies by ourselves and other workers in such areas as nerve cell loss following corneal wounding, the development of experimental animal models of herpes virus infections, and the role of the sympathetic nervous system in the regulation of corneal physiology. Materials and Methods A total of ten rats, six rabbits, four cats and four monkeys were used in this investigation. Some of the data from the rat and monkey experiments have been presented in preliminary fashion as parts of larger studies on corneal sympathetic innervation in the rat16 and brainstem termination sites of corneal afferent neurons in monkeys.13 AH experiments were performed in accordance with the guidelines outlined in the ARVO Resolution on the Use of Animals in Research. Each animal was anesthetized with ketamine (10 mg/kg, I.M.) or sodium pentobarbital (35-45 mg/kg, I.P. or I.V.), and the central area of the corneal epithelium was gently scratched in a checkerboard fashion with a sterile number 11 scalpel blade. Partial disruption of the epithelium was found in preliminary experiments to be necessary in order to permit satisfactory penetration of the tracer substance into the cornea and to obtain good retrograde transport within the corneal nerve fibers. The scratched area was in every case restricted to the central one-fourth to one-half of the cornea and never involved the peripheral cornea or the coraeoscleral limbus. The prepared corneal areas in cats were, on the average, smaller than those in the other species due to the tendency for the cat eyeballs to be drawn back into the orbit by contraction of the retractor bulbi muscles. The procedure was performed unilaterally in at least two animals from each species, and bilaterally in the remaining animals. ' Vol. 30 In the rabbits, cats and monkeys, the scratched area was encircled with a 2 mm high vaseline wall constructed by squeezing a thin strand of vaseline through a 25-gauge syringe needle. Thin wafers of gelfoam surgical sponge were then saturated with 5-8 n\ of 2% HRP-WGA in saline, placed on the scratched corneal surface, and manipulated until they completely filled the area inside the vaseline retaining wall. The preparation was continuously monitored through a dissecting microscope for 15-30 min, after which the gelfoam pledgets were removed and the corneal surface was rinsed for 2-3 min with sterile saline. The eyes were then gently dabbed with cotton applicator sticks and the eyelids were closed with butterfly bandages or sutures. In the rats, the method of tracer application was similar to that used in the larger animals, except that a modified corneal trephine with an inside diameter slightly larger than that of the scratched corneal surface was used to hold the HRP-WGA solution in place. Postoperative recovery for all of the animals was unremarkable and daily monitoring of their progress revealed no evidence of epiphora, photophobia or ocular irritation of any sort. At least one eye from one animal of each species was used as a control to check the specificity of the experimental design and to test for possible leakage of tracer away from the central corneal application site. In the latter cases, 3-5 n\ of 2% HRP-WGA was applied in dropwise fashion to the central areas of normal, nondamaged corneas and was permitted to flow freely over the entire cornea and surrounding extraocular tissues. Thirty minutes later, the tracer was removed by rinsing the eye repeatedly with sterile saline, dabbing it gently with cotton applicator swabs, and closing the eyelids with butterfly bandages or sutures as described above. Forty-eight to 96 hr later, all animals were reanesthetized and perfused through the left ventricle or retrogradely through the descending aorta with 200-500 ml of warm, phosphate-buffered saline, pH 7.4, containing 5000 units/kg heparin and 3% procaine-HCl, followed in sequence by 1-2 1 of warm fixative containing 1% paraformaldehyde-2% glutaraldehyde in 0.1 M phosphate buffer, and 1 1 of icecold 0.1 M phosphate buffer with 10% sucrose. The trigeminal ganglia, superior cervical ganglia, corneas and irides were immediately removed and placed in fresh 0.1 M phosphate buffer. In at least two animals of each species, the ciliary, middle cervical and stellate ganglia were also removed. Sections through each ganglion were cut serially at 40 pm in a cryostat, mounted directly on chrome alum-gelatin-coated slides, air-dried and reacted for the presence of HRP activity according to the tetramethylbenzidine Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 No. 3 CORNEAL INNERVATION / Morfurr er al (TMB) procedure of Mesulam.17 All tissues were then carefully examined with darkfield optics for the .presence of HRP labeled neurons. The corneas and irides from every animal were also examined in order to observe the application sites. Rat and monkey corneas and the irides of all four species were reacted whole with TMB histochemistry as free-floating preparations; the thicker cat and rabbit corneas were split along natural cleavage planes in the corneal stroma into two or three layers prior to TMB processing to permit better penetration of the incubation media components. Four radial slits were made in each tissue and they were then flat-mounted on chrome alum-gelatin-coated slides and air-dried under weighted coverslips. The total number of labeled neurons in each ganglion was determined by counting labeled somata containing visible nuclei. Other cells were also counted if their maximum profiles were contained entirely within the thickness of a section but were so heavily filled with reaction product that the cell nucleus was obscured. Corrections for double-counting of split neurons in the trigeminal ganglia were calculated by using the formula of Abercrombie,18 however, double-counting of the relatively small numbers of SCG neurons was not a problem since it was possible to identify the same cell in adjacent serial sections. The locations of every HRP-labeled neuron in three randomly selected trigeminal and superior cervical ganglia from each species were carefully plotted onto line drawings made by using a drawing tube attached to the microscope. Measurements were determined for 500 HRP-labeled trigeminal ganglion neurons in each species by using an ocular micrometer and averaging the maximum and minimum cell diameters through the equatorial region of each cell. To insure randomness of sample, cell diameters were obtained from at least four different ganglia for each species, and within each ganglion the labeled cells were measured in every third section to eliminate the possibility of measuring the same cell twice in adjacent sections. Results Corneal Injection Sites Examination of the corneas and irides from the experimental animals revealed a dense staining of the epithelium and stroma of the central scarified cornea, with no reaction product in the peripheral cornea, limbus or iris (Fig. 1). The percentage of the corneal area stained with TMB reaction product varied from animal to animal, but in general covered the central one-quarter to one-half of the cornea, and was, on the average, slightly less in the cats than in the other spe- 463 Fig. 1. Brightfield light micrograph of a rabbit corneal whole mount illustrating the central location and restricted size of a typical HRP-WGA application site (arrow). The application site in this particular animal covers approximately 48.5% of the total corneal area. Trigeminal Ganglia HRP-WGA application to the central cornea of the experimental animals resulted in the production of numerous well labeled somata and fiber processes in the ipsilateral trigeminal ganglia of all animals (Fig. 2). Labeled cells were never observed in the contralateral trigeminal ganglia. An average of 143 HRP-labeled neurons were observed in the trigeminal ganglia of the rat (n = 10), 172 in the monkey (n = 7), 277 in the cat (n = 5), and 449 in the rabbit (n = 6) (Fig. 3). The mean diameters of the labeled corneal afferent perikarya were 23.2 /im for the rat, 30.0 fim for the rabbit, 32.9 fim for the cat, and 30.8 jum for the monkey (Fig. 4). The cell frequency distribution of the cat was relatively symmetrical with respect to the mean (skewness = 0.86); however, the frequency of distributions of the rat, rabbit and monkey were skewed to the left of the mean (skewness = 1.58, 1.36 and 1.62, respectively). The labeled sensory neurons in all four species were located in the ophthalmic region of the ipsilateral trigeminal ganglion (Fig. 5) and were intermingled extensively among other neurons in the ophthalmic region that did not contain reaction product. In rats, cats and monkeys, the labeled cells were distributed relatively uniformly throughout both the mediolateral and anteroposterior dimensions of the ophthalmic region, except that cells in the ventral part of the cat and monkey trigeminal ganglia tended to be more numerous posteriorly. In marked contrast, labeled cells in the rabbit trigeminal ganglion were much more restricted in their distribution and Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 464 Vol. 30 INVESTIGATIVE OPHTHALMOLOGY G VISUAL SCIENCE / Morch 1989 Superior Cervical Ganglia Following HRP-WGA application to the central cornea, modest numbers of labeled neurons were observed in the ipsilateral SCGs of the rabbits (x = 53) and cats (x = 48), however, only occasional labeled neurons were observed in the rats (x = 1.1), and no labeled SCG cells were observed in the monkeys (Fig. 7). The labeled SCG neurons, when present, were stellate or spindle shaped and possessed variable numbers of radiating dendrites (Fig. 8). In all three species in which labeled cells were observed, the cells were concentrated in the rostral half of the ganglion; however, in cats, many cells were also observed in more caudal regions of the ganglion (Fig. 9). No labeled neurons were observed in the middle cervical or stellate ganglia of any of the animals. One faintly labeled neuron was found in the ciliary ganglion of one cat; all other ciliary ganglia were devoid of labeled neurons. Controls HRP-WGA application to the corneal surfaces of nondamaged, control eyes of each species labeled 23 and 35 neurons, respectively, in the trigeminal gan- Fig. 2. Darkfield light micrograph illustrating several HRPWGA-labeled corneal afferent somata (large arrows) in the ophthalmic region of a rabbit trigeminal ganglion, 72 hr after HRPWGA was applied to the central cornea. The labeled cell in the upper center of the figure gives origin to an axon that bifurcates (open arrowhead) into a peripheral and a central process. O 500- C bO 400- formed a well denned, longitudinal column approximately 0.4 mm in width that did not, except near the ventral surface of the ganglion, extend all the way to the medial edge of the ganglion (Fig. 5). There was no strong and consistant evidence for a somatotopic distribution of corneal afferent neurons within the dorsoventral axis of the trigeminal ganglion in any of the species examined (Fig. 6). Occasional labeled cells were observed in all four species within the transitional zone representing the apprpximate border between the ophthalmic and maxillary regions of the ganglion (Fig. 5), and it was difficult on the basis of somata location alone to classify these neurons as belonging to one region of the ganglion of the other. However, by serial analyses of adjacent sections, it was determined that the majority of the "border cells" were connected to fibers that entered the ophthalmic division of the trigeminal nerve. In one monkey, however, six neurons gave origin to peripheral axons that clearly entered the medial part of the maxillary nerve. bO 100- RAT CAT MONKEY Fig. 3. Histogram illustrating the mean number of HRP-WGAlabeled neurons ± SEM observed in the trigeminal ganglia of each species following tracer application to the central cornea. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 No. 465 CORNEAL INNERVATION / Morfurr er ol 3 RAT RABBIT X=30.8t9.0 X=23.3t5.7 32 3 6 4 0 4 4 4 8 52 5 6 6 0 6 4 68 1 2 1 6 2 0 24 28 32 3 6 4 0 4 4 4 9 52 5 6 6 0 6 4 0) g 100 -, M O N K E Y CAT X=30.8t6.3 X=33.1±7.7 2 0 2 4 2 8 3 2 3 6 4 0 4 4 48 52 5 6 6 0 6 4 6 8 soma diameter (\im) Fig. 4. Histogram illustrating the size distributions of 500 HRP-WGA-labeled, randomly selected, corneal afferent somata from each of the four species. glia of the rat, eight and six cells, respectively, in the trigeminal ganglia of the rabbit, and one cell in the trigeminal ganglion of one cat. Labeled cells were absent from the trigeminal ganglion of the second control cat, the trigeminal ganglia of the monkeys, and the SCGs of all control animals. Discussion The results of the present investigation provide detailed information on the origins, numbers and afferent somata diameters of the sensory and sympathetic neurons that innervate the central corneas in four species of mammals: rat, rabbit, cat and monkey. The data reported here extend the observations of'dther workers in this area and provide several important, new pieces of information, including: (1) the location of the corneal afferent neurons within the trigeminal ganglion of the rabbit; (2) somatotopic maps of the trigeminal ganglia of the rat, cat and monkey that provide more detail than those published pre- viously9"13 by virtue of the fact that in the current study the location of all labeled corneal afferent neurons, and not just those in representative sections, have been meticulously plotted; (3) detailed analyses of the dorsoventral distribution of corneal afferent neurons in the trigeminal ganglia of all four species, (4) comprehensive cell size analyses of 500 randomly selected corneal afferent somata from each species; and (5) comparative data on the numbers and locations of SCG neurons that provide sympathetic innervation to the mammalian cornea. Technical Considerations Specificity of the experimental design: Examination of the TMB-reacted corneal whole mounts and irides from the experimental animals (eg, Fig. 1) revealed that HRP-WGA applied to the central cornea remained in the central one-fourth to one-half of the cornea without spreading into surrounding tissues. If any tracer diffused into the peripheral cornea, the Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 466 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / March 1989 Vol. 30 Monkey Cat Rabbit Fig. 5. Schematic illustrations showing the locations of HRP-WGA-labeled corneal afferent neurons (solid circles) in the trigeminal ganglia of the monkey, cat, rabbit and rat. The figures are arranged in series with the most dorsal section to the left and progressively more ventral sections to the right. Each drawing represents a composite of six or seven consecutive sections and, taken collectively, each series of drawings accurately depicts the total number of cells seen in that ganglion. See text for details of cell distribution. Oph, ophthalmic nerve; max, maxillary nerve; mand, mandibular nerve. Rat corneoscleral limbus or iris, it did so in quantitities so small that we were unable to demonstrate its presence by using the sensitive TMB technique. The absence of labeled cell bodies from all but one of the ciliary ganglia examined (and in the latter case, only one faintly labeled cell was detected) further suggests that tracer leakage from the cornea into the neighboring tissues of the iris and ciliary body was negligible. Thus, it may be concluded that the labeled neurons observed in the trigeminal and superior cervical gan- glia of the current investigation resulted predominantly if not solely from the uptake and retrograde transport of tracer by nerve fibers that terminated in the central cornea. Controls: The small number of labeled neurons observed in some of the trigeminal ganglia of the control animals were probably labeled via HRPWGA uptake and retrograde transport in corneal afferent fibers that terminated near the epithelial surface. The control-labeled neurons were in all cases Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 467 CORNEAL INNEIWATION / Morfurr er ol No. 3 number of cells 0 10 I 1 20 I L 30 40 i I 10 I 0-. 20 1 30 40 1 1 J RAT » w* ¥ * rt (ft IT' * 1 RABBIT i V X r V i .•.:••-:.•• m Y I ,^t,r,M; 05 0.5 ',: iI] J 1 bO Fig. 6. Histogram illustrating the relatively uniform dorsoventral distribution of the corneal afferent neurons in the trigeminal ganglia of the species examined in the current study. in 1.0- ,, J ±"4:-:::i * J 40 J 10 1 0 CAT 20 I 30 I 40 I MONKEY .:.,..;.;^,j,,, 0.5 r -i i uyi^r n^ni i rj^jfTru i iJTS " .i..A.i:.:..M 0.5i —i—I 1.0 very faintly labeled and their somata diameters and location within the trigeminal ganglia were identical to the labeled corneal cells observed in the experimental animals. Cell counts: The numbers of trigeminal and superior cervical ganglion neurons labeled in the current investigation undoubtedly represent conservative estimates of the total number of cells that innervate the mammalian cornea since, as mentioned above, the tracer applications were restricted to the central cornea and did not include more peripheral areas of the tissue. However, since the aim of the current investigation was to study the sensory and sympathetic in- nervation of the cornea, we intentionally restricted the size of the application area in order to prevent nonspecific tracer uptake by nerve fibers in the corneoscleral limbus, sclera, ciliary body and iris. Nevertheless, since most large corneal nerve bundles converge radially on the central cornea and the central cornea is more densely innervated than the peripheral cornea,19 it is likely that HRP-WGA as applied in this study reached the terminal or preterminal portions of a large percentage of the nerve fibers that supply the cornea. The lack of labeled SCG cells in the monkey experiments merits special comment. It is doubtful that Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 468 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / March 1989 100' u .£ u T3 60- % ^3 4oH M-H o a! RAT RABBIT CAT MONKEY Fig. 7. Histogram illustrating the mean number of HRP-WGAlabeled neurons ± SEM observed in the superior cervical ganglia of each species following tracer application to the central cornea. Vol. 30 We have also shown that corneal afferent neurons in all of the species examined here are concentrated in the ophthalmic region of the ipsilateral trigeminal ganglion.9"13 Similar medial localizations of corneal neurons have been reported in other mammals, including, mice8 and guinea pigs (Keller et al, personal communication, 1988). However, the results of this and other studies provide no strong evidence for the existence within the ophthalmic region of a refined somatotopy according to the particular peripheral target organ innervated. For example, corneal neurons in cats overlap extensively with neurons that give rise to the supraorbital10 and ethmoidal20 nerves, and monkey corneal neurons overlap extensively with cells that innervate the extraocular muscles.21 Even in the rabbit, in which corneal neurons are clustered together to a larger degree than in other species, individual labeled corneal cells are separated from one another by unlabeled neurons of unknown peripheral distribution. A very small number of corneal afferent neurons in some trigeminal ganglia of cats12 and monkeys13 give rise to axons that enter the cornea via the maxillary division of the trigeminal nerve. Theoretically, these these "negative" data are due to inadequacies of the HRP-WGA retrograde tracing technique because HRP-WGA applied to the monkey central cornea resulted in strong labeling of a large number of neurons in the trigeminal ganglion (Fig. 3) and of their central projections to the trigeminal brainstem nuclear complex.13 In addition, similar methods of corneal preparation and tracer application produced labeled cells in both the trigeminal and superior cervical ganglia of all other species of animals used in this study. Thus, the absence of labeled monkey SCG cells in the current studies probably reflects a true lack of sympathetic innervation in this species and not a "false negative" due to technical insufficiencies. Trigeminal Ganglion Cell counts and somatotopic localization: The current study provides evidence that there is a positive correlation between corneal size and numbers of corneal primary afferent neurons in the trigeminal ganglion. More HRP-labeled cells are found in the trigeminal ganglia of animals with large corneas (eg, rabbits and cats) than of animals with smaller corneas (eg, rats), suggesting that larger corneas require greater numbers of primary afferent neurons to adequately subserve the sensory and trophic needs of the tissue. Fig. 8. Darkfield micrograph illustrating two HRP-WGA-labeled neurons (arrows) in the superior cervical ganglion of a rabbit. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 No. 3 469 CORNEAL INNEIWATION / Morfurr er ol fibers may provide important sources of preserved corneal sensitivity following total or subtotal ophthalmic nerve injury, although the available evidence1213 suggests that the contribution made by these fibers to the overall corneal innervation, when present, is very small. In contrast to the findings of Morgan and coworkers'2 who found that 4% of all corneal-innervating cells in the cat are located in the maxillary division, we found that all labeled neurons in the trigeminal ganglia of cats were located in the ophthalmic division. This variability may be the result of differences in the type of tracer used, methods of tracer application and percentage of the cornea exposed to the solution. The results of this study have the revealed that corneal afferent neurons are distributed relatively uniformly in the dorsoventral axis of the trigeminal ganglia of all four species examined. According to one current theory of trigeminal ganglion somatotopy, cells that innervate oral and perioral regions of the face are found in greater numbers in ventral regions of the trigeminal ganglion, and cells that supply peripheral regions of the head are concentrated dorsally.10'22-23 Thus, the distribution of corneal afferent neurons observed in this study may reflect the "central" position of this tissue in the orofacial somatotopic map. Interestingly, the uniform dorsoventral distribution of corneal neurons that we observed in the rat trigeminal ganglia differs from the pronounced dorsal concentration of corneal cells observed in an earlier study in this species1' and provides evidence for a previously unrecognized interanimal variability as regards the distribution of these cells. Corneal afferent somata diameters: Our results have shown that corneal afferent neurons in all four species examined constitute a population of predominantly small and medium sized neurons. The mean diameter of the cat corneal afferent neurons (33.1 (un) is nearly identical to the figure reported by Nishimori and coworkers14 (33.3 nm), however, the mean diameter of the rat corneal afferent neurons (23.3 um) is approximately 3-4 fim larger than the range reported by Sugimoto et al.15 Since the mean diameter of normal adult rat trigeminal ganglion neurons is 26.1 nm24 corneal afferent cells in the latter species are among the smaller neurons in the ganglion. The small sizes of the corneal afferent somata reported here are consistant with electron microscopic studies showing that the corneal innervation is derived almost exclusively from unmyelinated and finely myelinated axons.25 Electrophysiological observations have shown that the majority of corneal sensory axons conduct in the lower A delta fiber Cat Monkey Fig. 9. Schematic illustrations showing the distribution of HRPWGA-labeled neurons (solid circles) in representative superior cervical ganglia from each species. range, with some cold-sensitive units conducting in the C fiber range.2627 In all of the animals examined in the current investigation, a small percentage of the labeled somata were of relatively large size (greater than 40 fim in diameter), suggesting that they may serve a mechanoreceptive function. Clinical testing suggests that some tactile sensibility is present in the human eye, but that it is normally masked by the dominant sensation of corneal pain. 2829 A definite tactile component of corneal innervation has been described in patients with trigeminal tractotomies; cutting the spinal tract of V at the level of the obex in these individuals produces corneal analgesia while maintaining tactile sensibility.3031 Alternatively, the large diameter corneal afferent neurons may, like the more numerous smaller diameter neurons, signal some form of corneal pain. In the latter case, the size of the perikarya may be correlated with the total volume of the peripheral and central nerve branches supported by the cell, and not by the sensory modality subserved by that cell. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 470 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / March 1989 Sympathetic Innervation of the Cornea Our results have shown that the cornea of the rat, rabbit, and cat is innervated by small to moderate numbers of cells located in the ipsilateral superior cervical ganglion. These observations confirm and extend previous reports of corneal sympathetic nerve fibers in these species as revealed by fluorescent staining techniques32"35 and HRP-WGA anterograde transport procedures16 and the resultant disappearance of these fibers following superior cervical ganglionectomy.36 The negative findings reported here for the monkey also substantiate previous reports of a lack of a sympathetic innervation to the primate cornea, as evidenced by the inability to label corneal adrenergic fibers in this species via fluorescent staining 35 - 3738 or neuropeptide Y immunocytochemistry.39 We have also shown that the SCG neurons that supply the cornea are located primarily in the rostral half of the ganglion. The mammalian SCG is somatotopically organized; cells that project through the internal carotid nerve are concentrated near the rostral pole of the ganglion, whereas cells that project through the external carotid nerve aggregate further caudally.40"42 However, we found no evidence for somatotopic localization of SCG neurons according to the particular peripheral target tissue innervated. In the rat SCG, for example, neurons that supply the pineal gland,43'44 anterior eye chamber,45 Miiller's smooth muscle46 and cornea (current investigation) intermingle extensively with one another in the rostral region of the ganglion. Based on estimated numbers of neurons in the rat SCG of 29,000,47 in the rabbit, 76,000,48 and in the cat, 66,000,49 the number of labeled neurons that supply the cornea as indicated by our results (Fig. 7) represent only 0.01%, 0.1% and 0.2%, respectively, of the total SCG cell population. The data presented here also show that neither the middle cervical nor the stellate ganglia contribute to the corneal innervations in the species examined. The latter observation is compatible with what is known of the organization of the cervical sympathetic chain ganglia, ie, some cells in the middle cervical and stellate ganglia send their axons through the SCG and into the external carotid nerve to reach target tissues in the head and face,4150 but only cells in the SCG send their axons through the internal carotid nerve to supply the orbit. Functional considerations: The functional significance of the sympathetic innervation of the mammalian cornea is unclear, although several interesting hypotheses have been proposed. The literature in this area has recently been reviewed in detail16 and will only be commented on here briefly. There is a good Vol. 30 deal of experimental evidence to suggest that corneal sympathetic nerve fibers in rabbits modulate ion transport processes in the corneal epithelium,3 and, therefore, may assist the endothelium in regulating stromal hydration and corneal transparency. Sympathetic fibers in rats and rabbits have been shown to exert antimitogenic effects on the corneal epithelium 4 ' 551 and inhibitory effects on corneal wound healing.6-52 Finally, corneal sympathetic fibers may play a role in modulating the sensitivity of the cornea by interacting with the terminal or preterminal portions of trigeminal sensory nerve fibers.733 However, solid experimental evidence in support of the latter hypothesis is lacking. In humans with Homer's syndrome (in which the sympathetic outflow from the SCG to the eye has been interupted), the corneal epithelium of the affected eye shows abnormal function: epithelial thickness in the Homer's eye is slightly increased, and when subjected to an hypoxic "stress test" it shows significantly more epithelial greying, microcytic edema and a slower rate of desweUing.53 The significance of the pronounced interspecies differences in SCG innervation of the mammalian cornea as revealed in the current investigation is uncertain and will require additional study. Perhaps animals with very large corneas (eg, rabbits and cats) require greater numbers of SCG neurons than animals with small corneas (eg, rats) in order to maintain similar degrees of neural control over target tissues of disproportionate size. However, this theory is untenable when one considers that the monkey cornea, intermediate in size between that of the rat and the rabbit, is totally lacking in sympathetic innervation. Alternatively, sympathetic fibers may play more "important" roles in regulating certain aspects of corneal physiology and metabolism in some species than in others. However, it is unclear how the corneas of monkeys and humans, which contain few or no sympathetic nerve fibers,37'54 differ anatomically, metabolically or biochemically from the corneas of rabbits and cats, which contain modest numbers of sympathetic fibers. Key words: corneal innervation, trigeminal ganglion, superior cervical ganglion, herpes simplex virus, ocular nerves Acknowledgments The authors would like to thank Mr. Mark A. Jones for excellent technical assistance rendered during the course of these experiments and Ms. Kathleen Drajus for caring for the cats used in these experiments. References 1. Duke-Elder S and Leigh AG: Neurotrophic keratitis. In System of Ophthalmology, Vol. VIII, Diseases of the outer eye, Part 2, Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 No. 3 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. CORNEAL INNERVATION / Marfurr er ol Duke-Elder S and Leigh A, editors. St. Louis, The C.V. Mosby Co., 1965, pp. 803-810. Beuerman RW and Schimmelpfennig B: Sensory denervation of the rabbit cornea affects epithelial properties. Exp Neurol 69:196, 1980. Klyce SD and Crosson CE: Transport processes across the rabbit corneal epithelium: A review. Curr Eye Res 4:323, 1985. Friedenwald JS and Bushke W: The effects of excitement, of epinephrine and of sympathectomy on the mitotic activity of the corneal epithelium in rats. Am J Physiol 141:689, 1944. Mishima S: The effects of the denervation and the stimulation of the sympathetic and trigeminal nerve on the mitotic rate of the corneal epithelium in the rabbit. Jpn J Ophthalmol 1:65, 1957. Perez E, Lopez-Briones L, Gallar J, and Belmonte C: Effects of chronic sympathetic stimulation on corneal wound healing. Invest Ophthalmol Vis Sci 28:221, 1987. Morgan C, de Groat WC, and Jannetta PJ: Sympathetic innervation of the cornea from the superior cervical ganglion. An HRP study in the cat. J Auton Nerv Syst 20:179, 1987. Arvidson B: Retrograde axonal transport of horseradish peroxidase from cornea to trigeminal ganglion. Acta Neuropathol (Berlin) 38:49, 1977. Morgan CW, Nadelhaft I, and de Groat WC: Anatomical localization of corneal afferent cells in the trigeminal ganglion. Neurosurgery 2:252, 1978. Marfurt CF: Somatotopic organization of the cat trigeminal ganglion as determined by the horseradish peroxidase technique. Anat Rec 201:105, 1981. Marfurt CF and Del Toro DR: Corneal sensory pathway in the rat: A horseradish peroxidase tracing study. J Comp Neurol 261:450, 1987. Morgan C, Jannetta PJ, and deGroat WC: Organization of corneal afferent axons in the trigeminal nerve root entry zone in the cat. Exp Brain Res 68:411, 1987. Marfurt CF and Echtenkamp SF: Central projections and trigeminal ganglion somatotopy of corneal afferent neurons in the monkey, Macaca fascicularis. J Comp Neurol 272:370, 1988. Nishimori T, Sera M, Suemune S, Yoshida A, Tsuru K, Tsuiki Y, Akisaka T, Okamoto T, Dateoka Y, and Shigenaga Y: The distribution of muscle primary afferents from the masseter nerve to the trigeminal sensory nuclei. Brain Res 372:375, 1986. Sugimoto T, Takemura M, and Wakisaka S: Cell size analysis of primary neurons innervating the cornea and tooth pulp of the rat. Pain 32:375, 1988. Marfurt CF: Sympathetic innervation of the rat cornea as demonstrated by the retrograde and anterograde transport of horseradish peroxidase-wheat germ agglutinin. J Comp Neurol 268:147, 1988. Mesulam MM: Tetramethylbenzidine for horseradish peroxidase neurohistochemistry: A non-carcinogenic blue reactionproduct with superior sensitivity for visualizing neural afferents and efferents. J Histochem Cytochem 26:106, 1978. Abercrombie M: Estimation of nuclear population from microtome sections. Anat Rec 94:239, 1946. Rozsa AJ and Beuerman RW: Density and organization of free nerve endings in the corneal epithelium of the rabbit. Pain 14:105, 1982. Lucier GE and Egizii R: Central projections of the ethmoidal nerve of the cat as determined by the horseradish peroxidase tracer technique. J Comp Neurol 247:123, 1986. Porter JD, Guthrie BL, and Sparks DL: Innervation of monkey extraocular muscles: localization of sensory and motor 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 471 neurons by retrograde transport of horseradish peroxidase. J Comp Neurol 218:208, 1983. Beaudreau DE and Jerge CR: Somatotopic representation in the Gasserian ganglion of tactile peripheral fields in the cat. Arch Oral Biol 131:247, 1968. Kerr FWL and Lysak W: Somatotopic organization of trigeminal ganglion neurones. Arch Neurol 11:593, 1964. Forbes DJ and Welt C: Neurogenesis in the trigeminal ganglion of the albino rat: A quantitative autoradiographic study. J Comp Neurol 199:133, 1981. Hoyes AD and Barber P: Ultrastructure of the corneal nerves in the rat. Cell Tissue Res 172:133, 1976. Belmonte C and Giraldez F: Responses of cat comeal sensory receptors to mechanical and thermal stimulation. J Physiol 321:355, 1981. Tanelian DL and Beuerman RW: Responses of rabbit comeal nociceptors to mechanical and thermal stimulation. Exp Neurol 84:165, 1984. Giraldez F, Geijo E, and Belmonte C: Response characteristics of corneal sensory fibers to mechanical and thermal stimulation. Pain 7:1, 1979. Lele PP and Weddell G: Sensory neurons of the cornea and cutaneous sensibility. Brain 79:119, 1959. Grant FC, Groff RA, and Lewy FH: Section of the descending spinal root of the fifth cranial nerve. AMA Arch Neurol Psychiatry 43:498, 1940. Rowbotham GF: Observations on the effects of trigeminal denervation. Brain 62:364, 1939. Ehinger B: Distribution of adrenergic nerves to orbital structures. Acta Physiol Scand 62:291, 1964. 33. Laties A and Jacobowitz D: A histochemical study of the adrenergic and cholinergic innervatidn of the anterior segment of the rabbit eye. Invest Ophthamol 3:592, 1964. 34. Ehinger B: Distribution of adrenergic nerve in the eye and some related structures in the cat. Acta Physiol Scand 66:123, 1966. 35. Laties AM and Jacobowitz D: A comparative study of the autonomic innervation of the eye in monkey, cat, and rabbit. Anat Rec 156:383, 1966. 36. Tervo T and Palkama A: Sympathetic nerves to the rat cornea. Acta Ophthalmol (Copenh) 54:75, 1976. 37. Ehinger B: A comparative study of the adrenergic nerves to the anterior eye segment of some primates. Z Zellforsch 116:157, 1971. 38. Ehinger B and Sj6berg N-O: Development of the ocular adrenergic neve supply in man and guinea-pig. Z Zellforsch 118:579, 1971. 39. Stone RA, Laties AM, and Emson PC: Neuropeptide Y and the ocular innervation of rat, guinea pig, cat, and monkey. Neuroscience 17:1207, 1986. 40. Bowers CW and Zigmond RE: Localization of Neurons in the rat superior cervical ganglion that project into different postganglionic trunks. J Comp Neurol 185:381, 1979. 41. Dail WG, Khoudary S, Barraza C, Murray HM, and Bradely C: The fate of adrenergic fibers which enter the superior cervical ganglion. In Histochemistry and Cell Biology of Autonomic Neurons, SIF Cells, and Paraneurons, Eranko O, Soinila S, and Paiva'rinta H, editors. New York, Raven Press, 1980, pp. 287-297. 42. Jacobowitz D and Woodward JK: Adrenergic neurons in the cat superior cervical ganglion and cervical sympathetic nerve trunk. A histochemical study. J Pharmacol Exp Ther 162:213, 1968. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017 472 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / March 1989 43. Bowers CW, Dahm LM, and Zigmond RE: The number and distribution of sympathetic neurons that innervate the rat pineal gland. Neuroscience 13:87, 1984. 44. Patrickson JW and Smith TE: Innervation of the pineal gland in the rat: An HRP Study. Exp Neurol 95:207, 1987. 45. Arvidson B: Retrograde transport of horseradish peroxidase in sensory and adrenergic neurons following injection into the anterior eye chamber. J Neurocytol 8:751, 1979. 46. Smith PG, Brucker JW, and Mills E: Reinnervation of Muller's smooth muscle by atypical sympathetic pathways following neonatal ganglionectomy in the rat: Structural and functional investigations of enhanced neuroplasticity. Neuroscience 23:781, 1987. 47. Smolen AJ, Wright LL, and Cunningham TJ: Neuron numbers in the superior cervical sympathetic ganglion of the rat: A critical comparison of methods for cell counting. J Neurocytol 12:739, 1983. 48. Purves D, Rubin E, Snider WD, and Lichtman J: Relation of animal size to convergence, divergence, and neuronal number in peripheral sympathetic pathways. J Neurosci 6:156, 1986. 49. Wolf G: The ratio of preganglionic neurons to postganglionic 50. 51. 52. 53. 54. Vol. 30 neurons in the visceral nervous system. J Comp Neurol 75:235, 1941. Thomander L, Aldskogius H, and Arvidsson J: Evidence for a sympathetic component in motor branches of the facial nerve: A horseradish peroxidase study in the cat. Brain Res 301:380, 1984. Butterfield LC and Neufeld AH: Cyclic nucleotides and mitosis in the rabbit cornea following superior cervical ganglionectomy. Expfcye Res 25:427, 1977. Beuerman RW, Klyce SD, Vigo MC, Duruy BM, and Crosson CE: Modulation of latency and velocity in cornea! epithelial wound healing. ARVO Abstracts. Invest Ophthalmol Vis Sci 26(Suppl):91, 1985. Sweeney DF, Vannas A, Holden BA, Tervo T, and Telaranta T: Evidence for sympathetic neural influence on human corneal epithelial function. Acta Ophthalmol (Copenh) 63:215, 1985. Toivanen M, Tervo T, Partanen M, Vanna A, and Hervonen A: Histochemical demonstration of adrenergic nerves in the stroma of human cornea. Invest Ophthalmol Vis Sci 28:398, 1987. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933147/ on 05/05/2017