Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Molecular mimicry wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Adaptive immune system wikipedia , lookup
Atherosclerosis wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
TOWARDS A DETAILED UNDERSTANDING OF
THE RED BLOOD CELL STORAGE LESION
and its consequences for in vivo survival following
transfusion
ANDREAS HULT
Department of Community Medicine and Rehabilitation
Department of Surgical and Perioperative Sciences
Department of Integrative Medical Biology
Umeå 2015
Responsible publisher under swedish law: the Dean of the Medical Faculty
This work is protected by the Swedish Copyright Legislation (Act 1960:729)
ISBN: 978-91-7601-288-8
ISSN: 0346-6612
Cover illustration by MD, PhD Gustav Andersson. Copyright with artist.
Elektronisk version tillgänglig på http://umu.diva-portal.org/
Printed by: Print & Media
Umeå, Sweden 2015
Till Simon och Elvira
i
ii
Table of Contents
Table of Contents .......................................................................... iii
Abstract......................................................................................... v
Abbreviations ...............................................................................vii
Enkel sammanfattning på svenska .................................................ix
List of original papers .................................................................. xiii
Introduction................................................................................... 1
The leukocytes .................................................................................... 1
Innate immunity ................................................................................................. 2
Pattern recognition receptors (PRRs) ................................................................ 3
Adaptive immunity ............................................................................................. 4
Tolerance vs. Immunity ...................................................................................... 5
The Red blood cell............................................................................... 5
Function ............................................................................................................. 5
Erythropoiesis .................................................................................................... 7
The life and death of RBCs ................................................................................. 8
Eryptosis ............................................................................................................. 9
Recognition and phagocytosis of RBCs .............................................................. 9
Immune cells of the spleen .............................................................................. 11
RBC transfusion .................................................................................13
History .............................................................................................................. 13
Demographics .................................................................................................. 14
Effects of RBC transfusion ................................................................................ 14
RBC storage conditions .................................................................................... 15
The storage lesion ............................................................................................ 15
Risks associated with transfusion .................................................................... 17
How to study RBCs .............................................................................18
Human studies ................................................................................................. 18
Animal models ................................................................................................. 18
Aims ............................................................................................ 20
Materials and methods ................................................................ 21
Human studies (paper 1) ....................................................................21
Donation and reinfusion .................................................................................. 21
Blood analysis................................................................................................... 21
Maximal aerobic capacity (VO2max) assessment............................................. 22
Animal studies (paper 2 and 3) ...........................................................22
Labeling of RBCs ............................................................................................... 22
The mouse model for RBC storage and transfusion......................................... 23
In vitro characteristics of stored RBCs ............................................................. 24
In vitro phagocytosis ........................................................................................ 24
Ethics ................................................................................................................ 25
iii
Statistical analysis ............................................................................................ 25
Results ......................................................................................... 26
Paper 1 ..............................................................................................26
Transfusion of cryopreserved RBCs leads to acute accelerated RBC breakdown
......................................................................................................................... 26
Absent pro-inflammatory response in the acute phase after transfusion ....... 26
Additional results ............................................................................................. 27
Paper 2 ..............................................................................................28
Stored human RBCs contain a subset of Ca2+-high cells................................... 28
Characteristics of eryptotic murine RBCs ......................................................... 28
In vivo uptake of Ca2+-RBCs is located to the splenic marginal zone in a CD47independent manner ....................................................................................... 28
Additional results ............................................................................................. 29
Paper 3 ..............................................................................................30
Characteristics of the murine model for storage and transfusion ................... 30
Phagocytosis of stored human and mouse RBCs is serum dependent and
mediated by scavenger receptors .................................................................... 30
Additional results ............................................................................................. 31
Discrimination of RBC age fractions by in vivo biotinylation ....................... 31
Role of the RBC age at the start of storage for their 24 h survival post
transfusion ................................................................................................... 32
Phthalates during storage ............................................................................ 32
CD47 on stored murine RBCs does not influence the rapid elimination that
occurs during the first 24 h .......................................................................... 33
Discussion .................................................................................... 33
Frozen vs. liquid stored RBCs ........................................................................... 34
Inflammation after transfusion? ...................................................................... 34
The mouse model for storage and transfusion ................................................ 35
The role of calcium uptake in the storage lesion ............................................. 36
Impact of storage on RBCs - Senescence, eryptosis, or both? ......................... 37
How are stored RBCs presented to the immune system upon transfusion? ... 38
CD47 in senescence, eryptosis and the storage lesion .................................... 39
Scavenger receptor mediated phagocytosis of stored RBCs............................ 39
The role of serum in phagocytosis of stored cells ............................................ 40
Concluding remarks ......................................................................................... 41
Conclusions .................................................................................. 41
Acknowledgements ..................................................................... 43
References ................................................................................... 46
iv
Abstract
Red blood cells (RBCs) are vital for oxygen delivery to tissues and constitute the vast
majority of all cells in blood. After leaving the red bone marrow as mature cells, RBCs
have a lifespan of approximately 120 days before they are removed from the
circulation by macrophages, mainly in the spleen and liver. RBC transfusion is a
common therapy in modern healthcare. Major surgery, numerous cancer treatments
and other, often lifesaving, interventions would be unthinkable without available
blood supply. For this reason, hospitals store donated RBCs in blood banks.
The metabolic and structural changes that occur during prolonged storage of RBCs
(the storage lesion) have been studied in detail in vitro and include oxidative stress,
a reduction in glycolysis, increased membrane rigidity and shedding of
microparticles from the RBC membrane. Stored RBCs share several features of
senescent RBCs, but also with RBCs undergoing an apoptotic-like process called
eryptosis. A consequence of the storage lesion is the fact that as much as 25% of
stored RBCs could be rapidly removed from the circulation within 24 hours after
transfusion. The mechanisms behind this rapid macrophage-mediated recognition
and removal of stored RBCs, and its immunological consequences, remain largely
unknown. Therefore, the aims of this thesis were to investigate if cryopreserved
human RBCs induced an inflammatory response following autologous transfusion
into healthy volunteers, and to further understand the mechanisms behind
macrophage recognition of stored RBCs in vitro and in vivo.
Autologous transfusion of two units of cryopreserved RBCs into healthy human
recipients was found to be associated with an increased extravascular RBC
elimination already at 2 hours after transfusion. However, there were no signs of an
increased production of any of the investigated pro-inflammatory cytokines,
indicating that an increase in the destruction of RBCs per se did not induce an
inflammatory response.
Eryptosis is a form of induced RBC death associated with an increased cytoplasmic
Ca2+ uptake. We found that a subset of human RBCs increased their Ca2+
permeability during prolonged storage at +4°C. Using a murine model, to further
understand how RBCs with an increased Ca2+ permeability were eliminated by
phagocytic cells in the spleen, it was found that such RBCs were taken up by marginal
zone macrophages and dendritic cells (DCs) in a manner distinct from that of
naturally senescent RBCs. The DC population particularly efficient in this process
expressed CD207 and are known for their ability to promote immunological
tolerance. Eryptotic cell uptake was not regulated by the phagocytosis-inhibitory
protein CD47 on the RBCs.
v
To investigate how RBCs damaged during liquid storage are recognized and taken up
by macrophages, a model to store and transfuse murine RBCs was developed. This
storage model generated murine RBCs with several characteristics similar to that of
stored human RBCs (i.e. loss of ATP, formation of RBC microparticles and rapid
clearance of up to 35% of the RBCs during the first 24 h after transfusion). In vitro
phagocytosis of human as well as murine stored RBCs was serum dependent and
could be inhibited by blocking class A scavenger receptors using fucoidan or dextran
sulphate.
In conclusion, the findings of this thesis contribute to further understanding how
changes inflicted to RBCs during storage direct the fate of these cells in their
interaction with cells of the immune system after transfusion. The observation of an
increased Ca2+ permeability of stored RBCs, and the possible recognition of such cells
by tolerance-promoting DCs, in combination with the findings that class A scavenger
receptors and serum factors may mediate recognition of stored RBCs, may result in
novel new directions of research within the field of transfusion medicine.
vi
Abbreviations
2,3-DPG
Ab
APC
AS
ATP
CPD
CTL
CTLA-4
DC
EPO
ERK-1/2
FCS
FcγR
FSC
Hb
Hct
HS
Ig
IL
LPS
MARCO
MCP-1
MHC
MIP-1β
MP
MZ
MZM
NAbs
NK-cell
NTBI
PAMP
P38-MAPK
pCO2
pO2
PI3-Kinase
Poly(I:C)
PRR
PS
RBC
SAG-M
2,3- Diphosphogylcerate
Antibody
Antigen presenting cell
Additive solution
Adenosine triphosphate
Citrate-Phosphate-Dextrose
Cytotoxic T-lymphocyte
Cytotoxic T-lymphocyte-associated protein 4
Dendritic cell
Erythropoietin
Extracellular signal-regulated kinase-1/2
Follicular calf serum
Fc gamma receptor
Forward scatter
Hemoglobin
Hematocrit
Human serum
Immunoglobulin
Interleukin
Lipopolysaccharide
Macrophage receptor with collagenous structure
Monocyte chemoattractant protein 1
Major Histocompatibility Complex
Macrophage inflammatory protein 1 beta
Microparticle
Marginal zone
Marginal zone macrophages
Naturally occurring antibodies
Natural killer cell
Non-transferrin bound iron
Pathogen-associated molecular patterns
P38 mitogen-activated protein kinase
Partial pressure of carbon dioxide
Partial pressure of oxygen
phosphatidylinositide 3-kinase
Polyinosinic polycytidylic acid
Pattern recognition receptor
Phosphatidylserine
Red Blood Cell
Saline-Adenine-Glucose-Mannitol
vii
SIGLEC
SR
TCR
TH-cell
TLR
TNF-α
TReg-cell
TSP
VO2max
Sialic acid-binding immunoglobulin-type lectins
Scavenger receptor
T-cell receptor
T-helper lymphocyte
Toll-like receptor
Tumor necrosis factor- alpha
Regulatory T-lymphocyte
Thrombospondin
Maximal aerobic capacity
viii
Enkel sammanfattning på svenska
Mot en ökad förståelse för lagringspecifika förändringar hos röda
blodkroppar, vilka kan påverka dess överlevnad efter transfusion
Röda blodkroppar (RBK) är celler som saknar cellkärna och många andra
cellulära komponenter och vars primära syfte är att transportera syre från
lungor till kroppens celler. De cirkulerar i c:a 120 dagar innan de har ådragit
sig sådana åldersskador att de känns igen och bryts ner av immunförsvarets
makrofager i lever och mjälte. Kunskapen om de åldersskador som leder till
att cellerna tas upp och bryts ned är i dagsläget begränsad. Den normala
livslängden hos RBK kan förkortas i situationer som skadar RBK, exempelvis
sjukdom eller olika typer av läkemedel. Exakt hur makrofager känner igen
skadade RBK är delvis oklart. Blodtransfusioner är en vanlig
behandlingsform inom modern sjukvård, och direkt avgörande för att kunna
utföra stora operationer, vid flertalet cancerbehandlingar samt för att rädda
traumapatienter med stora blödningar. För att tillgodose behovet på blod vid
dessa ingrepp tappas blodgivare på blod som separeras i olika komponenter
(vanligtvis RBK, blodplättar och plasma) och lagras i +4°C vid blodcentraler
i väntan på transfusion. Det är sedan länge känt att under lagringstiden, som
i dagsläget är maximalt 42 dagar, ådrar sig RBK en rad skador. Detta leder
till att en substantiell andel av lagrade RBK (upp till 25%) försvinner kort
efter transfusion då de känns igen av mottagarens makrofager. De möjliga
riskerna för mottagaren som kan vara kopplat till detta är i dagsläget
omdebatterat i den vetenskapliga litteraturen. Även om många
lagringsspecifika skador har identifierats hos RBK så är mekanismerna
varmed makrofager känner igen och bryter ner vissa RBK efter transfusion
fortfarande mycket oklara.
Syftet med denna avhandling har därför varit att öka förståelsen för de
igenkänningsmekanismer som immunförsvarets celler använder vid kontakt
med RBK, samt hur immunförsvaret reagerar på denna igenkänning. Initialt
studeras detta i transfusionsförsök utförda på frivilliga försökspersoner. Det
finns dock inneboende etiska problem med att i tillräckligt stor detalj
studera dessa förlopp i människokroppen, vilket gör djurmodeller och
studier i cellkulturer till en mer framkomlig väg. Vi har därför satt upp en
musmodell för lagring och transfusion av RBK.
I det första delarbetet undersöktes vilken effekt transfusion av två blodpåsar
med fryslagrade RBK hade på inflammation i det akuta skedet (2 timmar)
samt två dygn efter transfusion. Vi fann en ökad makrofagmedierad
ix
nedbrytning av RBK i akutskedet efter transfusion, men att detta inte ledde
till mätbara nivåer av inflammatoriska substanser i blodet.
Skadade (även kallade eryptotiska) RBK kännetecknas av ett ökat upptag av
kalcium, vilket följs av att dessa celler snabbt tas upp av makrofager. Vi
observerade att en liten andel humana RBK vid lagring får förhöjda
kalciumnivåer, vilket var relaterat till en lång lagringstid. Delarbete två
utforskade med hjälp av en musmodell hur sådana skadade RBK känns igen
av immunförsvarets celler. Det visade sig att eryptotiska RBK togs upp av
marginalzonsmakrofager och dendritiska celler (dessa celler är viktiga för
reglering av immunförsvaret) i mjälten. Detta upptag liknar mycket hur
uttjänta kroppsegna celler (apoptotiska celler) tas upp, men skiljer sig från
upptag av normalt åldrade RBK. Upptag av apoptotiska celler motverkar
som regel inflammation och hjälper snarare till att förhindra
immunförsvaret från att attackera sina egna celler och vävnader. Då en liten
del av lagrade RBK påvisade eryptotiska drag, öppnas möjligheten att dessa
skadade celler faktiskt kan ha en positiv inverkan på mottagaren av en
blodtransfusion. Att eryptos kan utgöra en del av lagringsskadan stärks av
vårt fynd att upptag av lagrade RBK i mjälten delar likheter med både det
som ses hos normalt åldrade och eryptotiska RBK. Även om CD47 är ett
cellyteprotein som kan förhindra upptag av normala värdceller, så fann vi att
varken eryptotiska eller lagrade cellers upptag påverkades av mängden CD47
på dessa cellers yta.
Det tredje delarbetet visade att det finns anmärkningsvärda likheter varmed
lagrade RBK från mus och människa kan tas upp av makrofager, något som
stärker relevansen för upptäckter i vår musmodell. När färska RBK från mus
och människa blandades med musmakrofager skedde inget upptag. Däremot
så togs lagrade RBK upp oberoende om de kom från mus eller människa.
Makrofagernas upptag visade sig vara serumberoende, vilket indikerar att
det finns någon komponent i serum som krävs för makrofagernas
igenkänning av lagrade celler. Vidare kunde vi förhindra upptaget genom att
blockera en viss typ av receptorer (klass-A scavenger receptorer) på
makrofagernas yta. Detta tyder på att lagrade RBK känns igen med hjälp av
dessa receptorer hos makrofagerna, samt att upptaget involverar någon
serumkomponent.
Sammanfattningsvis så har detta avhandlingsarbete påbörjat en kartläggning
av interaktionen mellan lagrade RBK och kroppens immunceller, vilket
medfört en utökad förståelse för de skador som uppkommer hos RBK under
lagring och konsekvenserna av detta efter återinförsel av dessa celler i
blodet. Vi har funnit att ökad nedbrytning av RBK efter transfusion i sig inte
leder till inflammation. Vidare har avhandlingsarbetet kunnat koppla en del
x
av lagringskadorna som uppstår till så kallad eryptos, vilket skulle kunna
innebära att vissa lagringsskadade RBK kan ha en dämpande effekt på
kroppens immunförsvar. Avslutningsvis tyder arbetet på att makrofagers
upptag av lagrade RBK är beroende av komponenter i serum och klass-A
scavengerreceptorer på makrofagernas yta.
I en förlängning hoppas vi att kunskap om vilka förändringar hos lagrade
RBK som är relevanta för deras igenkänning av makrofager kan leda till
framtagandet av en mer optimerad lagringsmiljö för RBK. Sådana strategier
kan leda till att minska mängden RBK som behöver överföras till patienter i
samband med transfusioner om fler fungerande RBK kan behållas i
cirkulationen.
xi
xii
List of original papers
1. Transfusion of cryopreserved human red blood cells into
healthy humans is associated with rapid extravascular
hemolysis without a proinflammatory cytokine response
Hult A., Malm C. and Oldenborg P.A.
Transfusion, 2013. 53(1): p. 28-33.
2. Splenic uptake of RBCs with an elevated cytoplasmic Ca2+concentration primarily involves marginal zone
macrophages and CD207+ dendritic cells
Larsson A., Hult A., Nilsson A., Olsson M. and Oldenborg, P.A.
Submitted
3. Phagocytosis of liquid-stored red blood cells in vitro
requires serum and macrophage scavenger receptors
Hult A., Toss F., Malm C., and Oldenborg P.A.
Submitted
The published paper was reproduced by permission from the publisher John
Wiley & Sons Ltd.
xiii
xiv
Introduction
The human body contains, depending on body size, between 4 and 6 liters of
blood [4]. The main components of blood are the cell free plasma (50-60%)
and erythrocytes (red blood cells/RBCs; 40-50%), while white blood cells
(leukocytes) and platelets (thrombocytes) together accounts for only ~1% of
total blood volume [5]. The cellular components of blood originate from
pluripotent hematopoietic stem cells situated in the red bone marrow.
Hematopoietic stem cells give rise to erythrocytes, megakaryocytes (which
produce thrombocytes important for the coagulation process), and
leukocytes of lymphoid or myeloid origin [6]. The formation of blood cells, or
hematopoiesis, is strictly regulated by different hormones and cytokines in
order to meet the demand of the ever shifting physiological milieu of the
body, i.e. to produce more leukocytes upon infection or more erythrocytes
when tissue oxygenation is low [5].
The leukocytes
The immune system is built up by a variety of lymphoid tissues, leukocytes
and molecules, which are designed to protect us from infectious agents such
as bacteria, viruses, fungi and parasites. In addition to protecting us from
pathogens, the immune system also helps in maintaining body homeostasis
by removing and restoring damaged endogenous cells and tissues. After
differentiation and maturation, predominantly in the bone marrow,
leukocytes migrate to the blood, peripheral tissues and the lymphatic system.
In all these tissues, leukocytes carry out their various immunologic
functions. The immune system can be divided into two branches; the innate
and the adaptive system. The innate immune system is a rapidly responding
system that recognizes and eliminates a wide range of pathogen-associated
molecular patterns (PAMPs) found on various infectious agents. PAMPs are
identified by their binding to different pattern recognition receptors (PRRs)
expressed by the immune cells [6, 7]. The adaptive immune system is
initially much slower, but instead more efficient in clearing the body from
infectious agents. Furthermore, the adaptive system has a memory, and the
ability to generate long lasting immunity against different pathogens. Via
this memory function, pathogens will be swiftly recognized and eliminated if
invading a second time [6].
1
Innate immunity
The cellular components of the innate immune system derive mainly from
myeloid progenitors and include mast cells, monocytes, macrophages,
dendritic cells (DCs), and neutrophil, eosinophil and basophil granulocytes.
Despite their lymphoid origin, natural killer cells (NK-cells) are also
considered as innate immune cells which identify and kill virus-infected or
cancerous endogenous cells. Mast cells, macrophages and DCs are mainly
found in various tissues, while monocytes and granulocytes are mainly found
in the blood of a healthy individual. The neutrophil granulocyte is the most
abundant leukocyte in blood, constituting 50-70% of all leukocytes. It is
readily recruited to infected areas by inflammatory chemokines and
cytokines and play a key role in the initial defense against pathogens. Blood
monocytes eventually leave the circulation and migrate to organs and
peripheral tissues where they mature into macrophages. As they mature,
macrophages will display a unique receptor repertoire depending on their
microenvironment, making for instance a liver macrophage somewhat
phenotypically and functionally different from a marginal zone macrophage
in the spleen [8]. Upon recognition of infectious agents, the innate immune
system launches an inflammatory response by the release of histamine, proinflammatory cytokines and chemokines in order to recruit and activate
other immune cells to the infected area for the participation in the fight of
the infection [6]. The innate immune system also includes the ability of
antigen presentation, whereby macrophages or DCs scavenge foreign or
endogenous antigens in peripheral tissues by phagocytosis or
macropinocytosis for later presentation to the adaptive immune system.
In addition to fighting infectious agents, the innate immune system (DCs
and macrophages in particular) is also involved in maintaining normal
homeostasis in its microenvironment by engulfment of senescent or
apoptotic endogenous cells and cell fragments, a process mostly resulting in
anti-inflammatory responses [9]. The discrimination between (modified) self
and non-self is dependent on which receptor(s) that recognize(s) the cell or
substance destined for phagocytosis (or endocytosis) by the phagocytic cells
and ultimately determines if the cell will inhibit or induce an immune
response [7]. Aside from the cellular components of the innate immune
system, there is also the complement system which consists of several
plasma proteins with the ability to bind to various pathogens and either
initiate lysis of the pathogen (mainly bacteria) or elicit an inflammatory
response by the release of pro-inflammatory molecules and the formation of
opsonins on the surface of the pathogen. Opsonins target pathogens for
elimination by phagocytic cells that carry complement receptors (CRs) [10].
2
Pattern recognition receptors (PRRs)
The cells of the innate immune system carry a panel of receptors, PRRs, that
recognize evolutionary conserved patterns on pathogens - PAMPs as well as
molecules from damaged endogenous cells and tissues (danger-associated
molecular patterns - DAMPs) [11, 12]. These PRRs include the Toll-like
receptors (TLRs), which recognize bacterial patterns like lipopolysaccharide
(LPS) as well as viral RNA. Another member of the PRRs is the C-type lectin
receptors (CLRs). These receptors recognize carbohydrate structures on
pathogens. Endogenous carbohydrate structures, capable of binding to
CLRs, are often hidden by terminal sialic acid, thus hindering autorecognition [6]. Binding to TLRs and CLRs can elicit production of the proinflammatory cytokines interleukin-1β (IL-1β), tumor necrosis factor-α
(TNF-α) and IL-6 [11] , (Figure 1).
Scavenger receptors (SRs) also belong to the PRRs and can be involved in
recognition of PAMPs on pathogens as well as ligands on apoptotic cells [13].
The discrimination between self and non-self will ultimately determine if the
Pathogen
PRRs
PAMP
Recognition
Phagocytic cell
Degradation
of pathogen
Release of
pro-inflammatory cytokines
Nucleus
Cytokine
production
IL-1β, IL-6, TNF-α
Figure 1. PRR mediated pathogen recognition by a phagocytic cell.
PAMP recognition by PRRs results in; 1, Phagocytosis and subsequent degradation of
the pathogen. 2, Transcription, production and secretion of pro-inflammatory
cytokines e.g. IL-1β, TNF-α and IL-6. The release of pro-inflammatory cytokines will
initiate an inflammatory response that will recruit and activate other leukocytes to
the site of infection.
3
phagocytic cell will generate a pro-, or anti-inflammatory response. SRs are a
heterogeneous family of receptors with an extremely diverse ligand
repertoire, where most ligands are polyanionic and include proteins,
polyribonucleotides, polysaccharides and lipids [13, 14]. SRs are subdivided
into classes (class A-I, each with several members) and are known to partner
with various co-receptors for their function [13]. Macrophages express
several SR classes, including class A-SRs. SR-A1 and MARCO are two
receptors that belong to the class A-SRs found in the spleen. SR-A1 is
expressed by most macrophages while the expression of MARCO is restricted
to marginal zone macrophages (MZM) [13].
Adaptive immunity
The cells of the adaptive immune system derive from a common lymphoid
progenitor cell and can be divided into B-lymphocytes (B-cells) and Tlymphocytes (T-cells). Lymphocytes are, in contrast to leukocytes of the
innate immune system, highly specific in their ability to recognize antigen in
that it will only bind to one specific motif. During development, the genes
that are coding for the variable region of the B- or T-cell receptor undergoes
rearrangements and hypermutations, resulting in a wide range of receptor
variability and subsequent pathogen recognition within the lymphocyte
population. Each individual lymphocyte, however, is still highly specific in its
antigen recognition. Lymphocytes that recognize self-antigens undergo
apoptosis (programmed cell death) during development in order to avoid
autoimmune reactions. Those that survive this selection are allowed to
migrate from the bone marrow (or thymus in the case of T-cells) into the
blood and lymphatic system, including the spleen and lymph nodes [6].
When a B-cell has bound an antigen to its receptor (called B-cell
receptor/BCR), the antigen is internalized and later presented on MHC class
II to T-cells for T-cell dependent activation. Upon activation, the B-cell starts
to proliferate and changes phenotype into either a memory cell or a plasma
cell. The plasma cell starts to produce immunoglobulins (antibodies or ab:s)
which are then distributed via the lymph system out into the body, with the
ability to bind a specific antigen. Binding of ab:s to their antigens induces
opsonization of the pathogen which can then be recognized by phagocytic
cells having Fcγ-receptors (FcγR) that recognizes the constant region of the
ab:s. The binding of an ab to its antigen can, in addition, initiate activation of
the complement system which in turn increases the opsonization and leads
to further promotion of phagocytosis [10]. The memory B-cells can swiftly be
activated if presented with the antigen upon a second infection and hence
establishes a long lasting immunity against its specific antigen [6].
4
T-cells recognize its antigen via the T-cell Receptor (TCR). Antigens are
presented to T-cells by macrophages or DCs. When T-cells recognizes its
antigen, it can be activated and will then differentiate into one of the
following subsets; 1) Cytotoxic T-lymphocytes (CTL) whose main function is
to kill infected host cells, 2) T-helper cells (TH-cells) who are involved in coactivation of B-cells as well as other immune cells and, 3) Regulatory T-cells
(TReg-cells) which can promote suppression of lymphocytes. [6] This is of
course a simplified description of the extremely complex functions of the
immune system in general and the adaptive immune system in particular.
Tolerance vs. Immunity
DCs are particularly efficient antigen presenting cells (APCs) that control the
T-cell mediated immune response via paracrine and contact-dependent
signaling mechanisms. When a DC has taken up and processed an antigen, it
will migrate to the T-cell areas of secondary lymphoid organs and present
the MHC class II-bound antigen to naïve T-cells. If the engulfed antigen was
recognized as foreign by the DC, it will also express co-stimulatory receptors
(e.g. CD80 and CD86) and secrete pro-inflammatory cytokines (i.e.
interleukin 12 (IL-12) and IL-18) to promote activation of TH-cells and CTLs
[15] (Figure 2A). On the other hand, if the antigen was recognized as a selfantigen, the DC will display the antigen to T-cells together with lower levels
of CD80 and CD86, which will not be accompanied by pro-inflammatory
cytokine production. Instead, the anti-inflammatory cytokine IL-10 will be
released. Presentation of self-antigens may result in either T-cell anergy or
the formation of TReg-cells and promotion of tolerance against the selfantigen (Figure 2B) [16]. The activation of T-cells is hence a two-step process
in which binding of the T-cell receptor to the MHC-bound antigen on DCs is
the first step. The second step is either the ligation of CD80/CD86 on the DC
to CD28 on the T-cell to promote an immune response, or binding of
CD80/CD86 to CTLA-4 on the T-cell to promote tolerance.
The Red blood cell
Function
Erythrocytes (from now on referred to as RBCs), is the most abundant cell
type in the human body, and play a key role in oxygen delivery from the
lungs to tissues throughout the body. The transport of oxygen is mediated by
the iron-containing metalloprotein hemoglobin (Hb), which is highly
abundant in RBCs. The tertiary structure of adult Hb consists of four
polypeptide chains (two α-globin and two β-globin) which all carry an ironcontaining heme group to which oxygen is reversibly bound, forming
5
Pathogen
A
PRRs
Antigen presentation
on MHC class II
PAMP
Recognition
Degradation of
pathogen
Nucleus
APC
Naїve
T-cell
TCR
CD28
Pro-inflammatory
Co-receptors
Pro-inflammatory cytokines
IL-1β, IL-6, IL-12, IL-18, TNF-α
Apoptotic cell
B
PRRs
Antigen presentation
on MHC class II
Recognition of
modified self
Nucleus
Degradation
of cell
APC
Naїve
T-cell
TCR
CTLR-4
Anti-inflammatory
Co-receptors
Anti-inflammatory cytokines
IL-10
Figure 2. Recognition patterns on ingested material determine how
antigens are co-presented to T-cells.
Ingestion of antigen by an APC is degraded and later presented on MHC class II on the
cell surface. A, Recognition of foreign (e.g. pathogens) by APCs induce a proinflammatory response and subsequent T-cell mediated immunity. B, Recognition of
modified self (e.g. apoptotic cells) by APCs induces T-cell mediated tolerance in an
anti-inflammatory manner.
oxyhemoglobin (hemoglobin that has not bound oxygen is denoted
6
deoxyhemoglobin). The affinity of oxygen to hemoglobin is affected by the
partial pressure of oxygen (pO2) and carbon dioxide (pCO2), temperature,
pH and the intracellular concentration of 2,3-diphosphoglycerate (2,3-DPG).
Thus, the affinity regulation of oxygen to Hb optimizes oxygen uptake in the
lungs, and oxygen delivery to metabolically active tissues, dependent on the
physiological milieu at the respective site [17, 18].
RBCs also facilitate CO2 transport in two different forms; a small fraction of
CO2 is bound to the globin part of Hb, while the bulk is converted by the
enzyme carbonic anhydrase into carbonic acid (H2CO3) which dissociates
into H+ and bicarbonate (HCO3-) in the cytosol of RBCs. The HCO3- is then
exchanged with chloride (Cl-) across the plasma membrane by the anion
transporter membrane protein Band 3 (an abundant membrane protein in
RBCs) to further be transported in the blood plasma to the lungs where the
reaction is reversed, allowing CO2 to diffuse into the alveoli of the lungs and
finally be exhaled. [5]
The delivery of O2 to, and the removal of CO2 from, the tissues enables
adenosine triphosphate (ATP) production from oxidative phosphorylation to
occur in the mitochondria of the target cells and thus permitting an efficient
energy utilization throughout the body. Because mature RBCs do not contain
any organelles, including mitochondria, it will not consume any of the
oxygen it carries and are hence solely dependent on anaerobic glycolysis for
ATP production, leaving lactate as an end product. Lactate is finally expelled
from the RBC via symport of hydrogen ions in a pH dependent manner [5,
19].
Erythropoiesis
The maturation of RBCs from hematopoietic stem cells involves several
intermediates as the precursor cells divide and proliferate, starting to
produce Hb and extrudes organelles (including their nucleus), to finally
reach the state of reticulocytes. Still carrying ribosomes and mRNA for
protein synthesis, the reticulocytes are then released from the red bone
marrow into the blood stream. During the first couple of days in circulation,
the reticulocytes maturate into RBCs which have lost their capability to
produce proteins. At any given time, 1-2 % of all circulating RBCs are
reticulocytes. The reticulocyte concentration can hence be used to assess the
rate of the RBC production, since a higher percentage of reticulocytes is
indicative of increased erythropoiesis [20].
The regulation of erythropoiesis under normal conditions is highly
dependent on the peptide hormone erythropoietin (EPO), which is mainly
7
synthesized in the kidneys. EPO promotes committed erythroblasts in the
red bone marrow to proliferate and differentiate to RBCs [21]. The EPO
production is in turn regulated by the oxygen availability in the EPO
producing cells, thus creating an efficient feedback loop in where more EPO,
and consequently more RBCs, are produced when tissue oxygenation is low
[20, 22]. However, in cases of increased hemolysis and chronic anemia, EPO
regulation of erythropoiesis is not sufficient to compensate for the vast
reduction of RBCs. Instead, glucocorticoids and stem cell factor have been
suggested to facilitate the recovery from severe anemia by stimulating
extended proliferation of early erythroid progenitor cells [23].
The life and death of RBCs
The mature human RBC has a lifespan of approximately 120 days before it is
targeted for destruction by macrophages, mainly in the spleen and liver.
Although not fully understood, the senescence of RBCs is likely to involve
several structural changes on the cell surface, which ultimately leads to
macrophage recognition and phagocytosis [24]. Following ingestion of RBCs,
the macrophages recycle iron from Hb which is then transported back to the
bone marrow by the transporter protein transferrin in order to facilitate
continued erythropoiesis. In situations with increased RBC breakdown,
transferrin can become saturated. The highly reactive iron ions, capable of
forming reactive oxygen species and tissue damage in its unbound state, is
then found as non-transferrin bound iron (NTBI) in the plasma [25]. The
remaining heme-group of Hb (without iron) is converted to bilirubin in
macrophages and later secreted in the blood plasma [26]. Changes in the
plasma bilirubin concentration and transferrin saturation can hence be used
as indicators to assess the rate of RBC phagocytosis.
Senescence markers of RBCs include the clustering and/or degradation of
the Band 3 membrane protein which has been shown to be formed after
various oxidation processes. This may be attributed to the binding of
oxidized Hb to the cytoplasmic domain of Band 3 [24, 27, 28]. Alterations in
Band 3 can also be induced trough the activation of calcium- dependent
proteases like calpain, at least in the elderly [29]. RBCs shrink in size and
increase their density as they age, which can be seen by lower forward scatter
(FSC) as assessed by flow cytometry and by density centrifugation
fractionation [30]. The decrease in cell size is contributed to by the shedding
of microparticles (MPs) throughout the lifespan of the RBC [31]. MPs
contain various amounts of Hb and their formation seems to be, at least
partly, facilitated by the spleen [31, 32]. The MPs display the phospholipid
phosphatidylserine (PS) on their outer membrane leaflet and are quickly
removed from the circulation by macrophages [33]. If human RBCs
8
themselves externalize PS as a natural part of their senescence is
questionable [30], but cannot be completely ruled out due to the fast
recognition of PS+ cells by macrophages, leaving them little time to be
detected in circulation. Reductions in terminal sialic acids [34, 35] and in the
expression of the cell surface glycoprotein CD47 [36] have also been
observed in the membrane of senescent RBCs. Increased non-enzymatic
glycation (forming advanced glycation end products (AGEs)) of Hb as well as
membrane proteins is also found to occur as RBCs age [37, 38]. Finally, the
metabolic rate of RBCs decrease over time as seen by reduced activity of
glycolytic enzymes and a consequent reduction in ATP and lactate
production [39].
Eryptosis
Besides normal senescence, RBCs can also enter a programed cell death
(apoptotic)-like process called eryptosis, which can be induced by an number
of endogenous and xenobiotic substances, as well as by oxidative stress,
osmotic chock and energy depletion [40]. Eryptosis is not a pure analogue of
apoptosis in nucleated cells, as RBCs lack mitochondria and nucleus.
Eryptosis and apoptosis yet share many common features like cell shrinkage,
MP formation and PS externalization, which in RBCs is induced by increased
cytosolic Ca2+ levels [41, 42]. The actions of increased cytosolic Ca2+ involves
the activation of K+ ion channels and subsequent reduction in cell size,
cytoskeleton degradation (and membrane blebbing) by activated calpain,
and an increased scrambling activity of the phospholipids in the lipid bilayer
resulting in PS-externalization [42]. Eryptosis has also been reported to
occur in a number of diseases, including type-2 diabetes [43, 44],
thalassemia [45] and heart failure [46], often related to oxidative events.
Recognition and phagocytosis of RBCs
The mechanisms behind macrophage removal of normally senescent RBCs,
reaching the 120 day limit of their life-span, are still not completely
understood [2]. This lack of detailed knowledge is one important reason why
there is an even greater deficit in the understanding of the mechanisms
behind the rapid post-transfusion clearance of stored RBCs, which will be
discussed further in a later section. In addition to the biochemical changes
that occur during RBC senescence, clearance of senescent RBCs requires
molecular changes to the cell surface which can then be recognized by
macrophages (Figure 3). Most likely, senescent RBCs gradually accumulate
these cell surface changes, suggested to include loss of terminal sialic acids
and the subsequent exposure of underlying carbohydrate structures [34], a
reduction of the anti-phagocytic protein CD47 on the cell surface [36],
9
binding of naturally occurring antibodies (NAbs) [30, 47] and oxidative
damage to Hb and phospholipids [48]. Many of these changes may well be
recognized by NAbs, but the most convincing target for such antibodies so
far is the conformational changes that take place in the Band 3 protein as a
result of oxidative damage to hemoglobin [27, 28, 49]. Although NAbs are of
low affinity and at low concentrations in circulation, binding of NAbs could
result in FcγR-mediated phagocytosis of senescent RBCs but also
complement deposition and CR participation [50]. Finally, while being
questioned [30], senescent RBCs may also, like nucleated apoptotic cells,
eventually expose PS on the surface to promote their phagocytosis by
macrophages [51]. PS can be recognized by several receptors on phagocytic
cells, either directly by TIM1, TIM4 or stabilin-2, or via bridging molecules
like Gas-6, thrombospondin (TSP) and lactadherin, following interaction
with integrins on the macrophage [2].
Normal RBC
Senescent RBC
Sialic acid
SIGLEC
PS
CD47
SIRP-α
Tim-4
Anti-phagocytic
receptors
TSP
Integrin
Band 3
NAbs
FcγR
C3b
CR
Pro-phagocytic receptors
Macrophage
Figure 3. Pro-, and anti-phagocytic receptors on macrophages.
Abbreviations: SIGLEC - Sialic acid binding immunoglobulin lectins, SIRP-α Signal-regulatory protein-α, PS – Phosphatidylserine, TIM-4 - T-cell immunoglobulin mucin protein 4, TSP - Thrombospondin, NAbs - Naturally occurring
antibodies, FcγR – Fc gamma receptor, CR – complement receptor, C3b –
complement protein 3b.
Modified from de Back et al. [2].
10
In addition to the pro-phagocytic receptors mentioned above, macrophages
also carry anti-phagocytic receptors (figure 3). Signal-regulatory protein α
(SIRPα) is an inhibitory receptor expressed on phagocytic cells that
recognizes the widely expressed “self-antigen” CD47 [52]. Mouse RBCs
lacking CD47 was shown to be rapidly phagocytized by red pulp
macrophages in the spleen, indicating the anti-phagocytic properties of the
CD47-SIRPα interaction. Another family of receptors that mediates
recognition of self and inhibits phagocytosis is the sialic acid binding
immunoglobulin lectins (SIGLECs) that recognizes terminal sialic acids on
host cells [53]. A reduction in sialic acid on RBCs (by neuraminidase
treatment) leads to the rapid clearance of these cells by macrophages [54]. A
reduction in CD47 and terminal sialic acid on aging RBCs have been
observed [34-36], which may well contribute to macrophage recognition of
senescent RBCs.
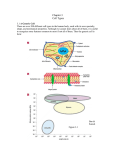
Immune cells of the spleen
Principally, the spleen has two separate compartments; the immunological
and lymphocyte-rich white pulp, and the RBC-rich red pulp having more of a
filtering function [1, 55, 56]. The splenic white pulp is divided into separate
T-cell and B-cell areas. Macrophages can be found in both the red and white
pulp, as well as in the area separating these two domains – the marginal
zone (MZ). Separate DC subsets are also distributed within the red pulp, MZ
and white pulp (T-cell) areas. In the murine spleen, three major macrophage
populations can be identified; the F4/80+ red pulp macrophages, the
MARCO+ MZ macrophages and the MOMA-1+ marginal metallophilic
macrophages (MMM) [1, 3] (Figure 4). F4/80+ macrophages are mainly
involved in the clearance of cells trapped in the red pulp, whereas the other
two macrophage populations are more specialized in trapping microbes and
PS+ apoptotic cells. Due to their close localization to the splenic white pulp,
MARCO+ and MOMA-1+ macrophages (together with DCs) are more actively
interacting with lymphocytes to regulate the adaptive immune system. When
investigating macrophage phagocytosis of RBCs, it is important to remember
that macrophages in vivo are very heterogeneous as a group, with many
specific functions depending on their phenotype and anatomical location [8].
Adding on to this complexity, activated macrophages may also promote as
well as dampen inflammation, making analysis of specific macrophage
subsets even more important [8].
11
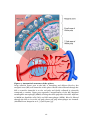
Figure 4. Anatomical structure of the spleen.
Large arteriole (lower part to the left) is branching and delivers blood to the
marginal zone (MZ) and sinusoids of the spleen. Blood is then filtered through the
MZ or arteriole sinusoids in to the red pulp and finally collected in sinusoids
connected to venules. Upper part represent a magnification of the MZ. Marginal
metallophilic macrophages (MMM) is lining the white pulp side of the MZ. Adjacent
to MMM are dendritic cells (DCs) and MZ macrophages (MZM). Blood is filtered
through the MZ an in to the red pulp where red pulp macrophages are situated.
(Modified from Junqueira et al., [1] and Cyster [3]).
12
RBC transfusion
History
The first successful blood transfusion in humans was carried out by
obstetrician Dr. James Blundell in 1818 when he transfused approximately
400 ml of whole blood, which was collected from multiple donors and
administrated ten times over a 40 min period of time [57]. The patient,
suffering from gastric carcinoma (described at the time as “scirrhosity of the
pylorus”), showed clear symptoms of anemia and was barely conscious at the
time of transfusion, but displayed marked improvement in the following 24 h
post transfusion. His symptoms then reemerged and were followed by death
a little over two days after the transfusion. Dr. Blundell continued his
pioneering work in blood transfusion with a total of 10 documented
transfusions of which 5 were successful [58].
The next big step towards modern transfusion medicine was taken by Karl
Landsteiner in the early 20th century. He discovered that sera from different
donors sometimes, but not always, agglutinated blood cells of other donors.
He proposed the ABO blood group system (although Dr. Landsteiner at the
time classified blood into groups A, B and C) [59] as a method to conduct
compatible blood transfusions, a discovery that ultimately rendered him the
Nobel prize in physiology or medicine in 1930. The identification of the
Rhesus blood group system (the second most important blood group system
after the ABO system) in the late 1930’s [60] is also to be considered an
important step on the way to safer and more compatible blood transfusions.
Today, more than 300 antigens belonging to over 30 blood group systems
have been described[61]. Blood group antigens are proteins or carbohydrate
structures present on the cell membrane of RBCs. Alloimmunization against
blood group antigens can occur after RBC transfusion, owing to blood group
incompatibilities between donor and recipient [61]. Alloimmunization
results in ab-mediated immunity against the foreign antigen. In most cases,
the production of ab:s against blood group antigens is an induced response
which follows after contact with the respective antigen. In contrast, ab:s
against A and B antigens of the ABO blood group system starts to be
produced during the first year after birth [62], irrespective of contact with
the antigens. Enzymatic removal of the B-antigen from RBCs of blood group
B has been described, making them transfusable to donors of all blood
groups [63]. The combined removal of both the A and B antigens have later
been reported [64], opening the door for universal RBCs that can be
transfused to everyone, regardless of the ABO-blood group of the donor or
recipient.
13
The use of citrate as an anticoagulant in donated blood made transfusions
easier to manage as the risk for coagulation was vastly reduced [65]. The
addition of glucose (or dextrose) in combination with citrate enabled
prolonged storage of blood possible [66]. The introduction of first AcidCitrate-Dextrose (ACD) and later Citrate-Phosphate-Dextrose (CPD) as a
combined anticoagulant and energy source to the collected blood (stored in
plastic bags) enabled donation and transfusion to be separated in time by as
much as 28 days when stored at +4°C [67], thereby making storage of blood
in blood banks a routine in many hospitals.
Demographics
According to the World Health Organization (WHO), 108 million units of
blood were donated worldwide in 2012, with approximately half of these
donations occurring in high-income countries where only 18% of the world’s
population lives. In low-income countries, a majority of the collected blood is
stored and later on transfused as whole blood, as compared to high income
countries where 95% of the donated blood is further processed into RBC-,
plasma-, and thrombocyte concentrates [68].
Effects of RBC transfusion
Blood transfusions are used both in the setting of acute and chronic
bleeding, as well as in patients with bone marrow suppression, e.g. cytostatic
treatment, and in patients with hemoglobinopathies (e.g. thalassemia and
sickle cell disease) [69, 70]. Symptoms of anemia (i.e. fatigue, tachycardia
and dyspnea) usually only arises at Hb concentrations below 90-100 g/L due
to compensatory mechanisms that increase cardiac output and replenish
blood plasma volume [71]. However, symptom onset is largely dependent on
patients’ related factors such as age, cardiorespiratory fitness, rate of anemia
development and functional requirements. The recommendations to
transfuse hemodynamically stable patients is generally recommended at Hb
70-80 g/L [71]. The restoration of blood volume and viscosity after profound
hemorrhagic shock and hemodilution is also important in order to restore a
functional microcirculation by increasing capillary blood perfusion [72].
Moreover, the oxygen carrying capacity following transfusion has been
documented to increase in non-anemic men as assessed by increased arterial
oxygen content and greater resistance to hypoxic conditions during exercise
[73], an observation that has opened up for the misuse of blood transfusions
as a doping agent in endurance sports [74].
14
RBC storage conditions
To allow for a more efficient use of donated blood, blood donation and
transfusion have to be optimally separated in place and time. This can be
achieved by avoiding coagulation and assuring a sufficient ex vivo storage
milieu with respect to nutrients and buffered pH conditions [66]. The first
step involves collecting donated blood in CPD. As mentioned above, donated
blood is today often separated into its different components. In this process,
RBCs are isolated from the other cellular components of the blood, as well as
from the majority of the plasma, by centrifugation and filtration techniques
carried out inside a closed system containing the necessary components. The
last step in this process involves the addition of an additive solution (AS) to
the packed RBCs in order to optimize storage conditions. In Europe, a
solution of Saline-Adenine-Glucose-Mannitol (SAG-M) is widely used, while
closely related solutions (abbreviated AS-1, AS-3 and AS-5) are used in
America. Blood stored at +4°C in these solutions can be kept for up to 42
days and typically display an in vivo recovery of 78-82% 24 h after
transfusion into healthy individuals [75]. This may, however, not reflect the
24 h recovery in a clinical setting, since there are reports involving
transfusions to patients with various underlying diseases showing 24 h
recoveries after transfusion of around 75% already after 25-35 days of
storage [76, 77]. Recently, a new additive solution (AS-7) has been approved
for prolonged storage of RBCs for up to 8 weeks [78]. The AS-7 differs from
the previous storage solutions by containing NaHCO3 and NaPO4 but no
saline, having a higher pH and lower osmolality and displaying an in vivo
24-h recovery of 88% at day 42 and 82% at day 56 of storage [78].
The storage lesion
RBCs in liquid storage undergo time-dependent metabolic and structural
changes, collectively referred to as the storage lesion. The storage lesion has
been well characterized, based on in vitro measurements on RBC
concentrates over time. Metabolic alterations are manifested by the
reduction in ATP and 2,3-diphosphoglycerate (2,3-DPG) content. As RBCs
are purely glycolytic, a time-dependent increase in levels of lactate and a
corresponding decrease in pH is also noticed in the storage media. Structural
changes, partly due to oxidative damage, include conformational changes in
the Band 3 anion transport protein, IgG binding as well as increased rigidity
and osmotic fragility. Shape shifting and a reduction in size is also noted in a
subset of stored RBCs, attributed to the shedding of MPs and by reduced
levels of cytoplasmic K+ [76, 79-82]. The amount of CD47 may also be
reduced during storage [83, 84], although as to which extent is not clear.
Thus, stored RBCs show several similarities to what has been described for
15
normally senescent RBCs in circulation (Figure 5). ATP and 2,3-DPG levels
have been shown to recover shortly after transfusion, indicating that at least
part of the storage lesion is reversible in vivo following transfusion [85].
However, it is presently unclear if the storage lesion simply reflects an
accelerated aging of the RBCs, if it is merely a stress-induced eryptosis, a
combination of both, or something else. Perhaps more important, the
physiological consequences in vivo (after transfusion) remains largely
unknown [86]. It is also unclear if the storage lesion affects all age-fractions
of the RBCs in a random way, or if it primarily affects the older cells within
the RBC population. This distinction may be of importance to understand
the storage lesion and in the long run to develop strategies to prevent it.
In addition to liquid storage, RBCs can be frozen (cryopreserved) for
extended periods of time, using glycerol as a cryoprotectant. Cryopreserved
RBCs are more fragile and display increased hemolysis upon thawing and
washing, as compared to liquid stored RBCs, but instead maintain their ATP
and 2,3-DPG content better [87, 88]. In addition, RBC cryopreservation does
not result in MP formation, PS exposure or reduced levels of CD47 [89]. The
24 h in vivo recovery of cryopreserved RBCs has been reported to be ~85%,
making cryopreservation of RBCs a valuable alternative to liquid storage,
especially for patients with rare blood groups or when there is a shortage of
liquid stored RBCs [88].
Senescent
RBC
Microparticle formation
Glycolysis
ATP
2,3-DPG
Lactate
Oxidative damage
Ca2+ influx
Band 3 aggregation/degradation
Phosphatidylserine exposure
IgG binding
Sialic acid
CD47
Stored
RBC
+
+
↓
↓
↓
↓
↓
?
+
+
+
?
?
+
+
?
?
+
+
↓
↓
?
↓(?)
Eryptotic
RBC
+
?
?
+
+
+
?
?
Figure 5. A comparison between senescent, stored or eryptotic RBCs.
Presented is a selection of metabolic and structural changes reported in the
literature.
16
Risks associated with transfusion
Transfusion-related acute lung injury (TRALI) is the leading cause of
fatalities directly related to transfusion, followed by hemolytic transfusion
reactions due to blood group incompatibility [71]. In a systemic review of 45
cohort studies, evaluating the efficacy of RBC transfusions in high-risk
patients, the risks associated with transfusion were found to outweigh the
benefits in all but three studies [90]. Firm conclusions of the relation of risks
and benefits of transfusions are however difficult to draw because of the
inherent bias in cohort studies as sicker patients tend to receive more
transfusions [91]. There has also been an ongoing debate as to whether RBCs
that have been stored for longer periods of time will have adverse effects
following transfusion in various clinical settings. These effects include
increased risks of infection, multiple organ failure, length of hospital stay
and mortality [92-94]. A compilation of 32 studies on the topic by Aubron et
al. [95] found that 18 studies reported clinically harmful effects and 14 did
not. The discrepancy is likely due to various confounders not taken in to
account, as well as problems with underpowered studies. It is also possible
that transfusion of older blood can be particularly harmful to certain patient
groups [92].
Recently, a large randomized controlled trial (Age of blood evaluation,
ABLE) was published in which over 2400 ICU-patients had been enrolled
and randomly selected to receive either fresh RBCs (stored for 6.1±4.9 days)
or standard issued RBCs (stored for 22.0±8.4 days) [96]. The authors found
neither an increased risk of 90 day mortality, nor any other risks that could
be associated with transfusion of stored RBCs. Similar findings have been
reported from randomized controlled trials conducted on cardiac surgery
patients (the RECESS trial) [97] and in premature infants (the ARIPI trial)
[98], indicating that there are no major risks associated with transfusion of
older blood, at least not for the patient groups that have been investigated.
Still, because the maximum storage of RBCs in AS ranges from 42 to 56 days,
it could be argued that the average storage times of 22 days (ABLE), 28 days
(RECESS) or 14.6 days (ARIPI) may not be relevant to asses “older” RBCs.
This argument becomes particularly important in case the possible adverse
effects related to prolonged RBC storage would be manifested closer to the
end of the maximum storage time [99]. In addition, despite the relatively
large number of participants, the above mentioned studies have been
challenged as underpowered, thus failing to detect possible small but
clinically relevant differences [99, 100].
Several animal studies have been conducted in the search for the
mechanistic explanation to the supposed risks with transfusion of stored
17
RBCs. Hod et al. reported that mice receiving the equivalent of 2 units of
stored (but not fresh) RBCs manifested with high levels of pro-inflammatory
cytokines and NTBI in the plasma 2 h after transfusion [101]. In addition,
they found that the pro-inflammatory response to lipopolysaccharide (LPS)
stimulation was enhanced by the transfusion of stored, but not fresh RBCs
[101]. Comparable inflammatory responses were obtained in a similar study
in canines [102]. “The iron hypothesis” have been suggested to explain the
inflammatory response seen in animal studies. This states that inflammation
is mediated by an increased erythrophagocytosis and iron overload in the
macrophages, resulting in release of reactive iron ions in a quantity that
exceeds transferrin binding capacity [103]. However, in a human study
where 1 unit of RBCs stored for 42 days was transfused, there were
indications of NTBI in the plasma but no signs of a pro-inflammatory
response [104].
How to study RBCs
Human studies
As mentioned earlier, the current (poor) understanding of the storage lesion
and its effects upon transfusion is largely based on in vitro studies of stored
RBC concentrates, which may or may not be relevant to explain the swift
macrophage recognition of these cells after transfusion. In vivo studies are
usually limited to non-invasive monitoring of 24 h survival assessments
using 51Cr or biotin in vitro-labeled RBCs [105, 106]. As for the efficacy of
transfusion, there are indications that stored RBCs actually decrease tissue
oxygenation upon transfusion [107, 108]. This is possibly attributed to a
reduced vasodilation activity as a consequence of a decreased ATP content
and nitric oxide (NO) availability [109]. However, there are limitations in the
studies conducted to measure tissue oxygenation in humans, which
complicate the interpretation of these findings [110].
Animal models
Human physiology or pathophysiology is, of course, best studied directly in
humans. However, that is not always practical or possible to do, in part due
to ethical considerations. Therefore, the use of the mouse as a model system
is common in order to enable studies not easily performed in humans. The
mouse offers advantages by its small size, short generation time and both
homologous and modifiable genetic backgrounds. Although mice are not a
man, they are still a close relative when comparing the genomes with ~99%
of the human genes having mouse homologues [111]. Even though the
immune system of humans is quite similar to that of mice, it is nonetheless
18
important to mention that there are differences between species which needs
to be taken in to account before making generalizations of findings in mice
and its potential impact for humans. For instance; the main leukocyte cell
type in human blood is neutrophil granulocytes which constitute 50-70% of
total leukocytes. In mice, neutrophils only account for 10-25% and
lymphocytes are instead the most abundant among blood leukocytes.
Immunological differences between species also includes the occurrence and
distribution of various kinds of Toll-like receptors (TLRs), CD4 and CD33, Fc
receptors and immunoglobulin subclasses, as well as differences in cytokines
secreted by the cells of the immune system. This can be seen as a reflection
of different strategies used by the two species to cope with the environmental
demands that they have been subjected to during the 65 to 75 million years
of evolution that separates us [112].
Mouse RBCs have a shorter lifespan than human RBCs, typically about 50
days [113]. Senescent mouse RBCs are recognized by macrophages in spleen
and liver [114]. The senescence mechanisms seem to be similar in mice and
humans with respect to rheological properties and cell shrinkage [115].
Although at an accelerated rate, human and mouse RBCs also display similar
storage characteristics [116]. Taken together, this indicates that a mouse
model of storage and transfusion of RBCs could prove useful in getting a
deeper understanding of implications of the human red blood cell storage
lesion after transfusion.
19
Aims
Conflicting results have been reported regarding inflammation as a potential
side effect of transfusion of stored RBCs.
• The aim was to investigate if cryopreserved human RBCs induce an
increased erythrophagocytosis and subsequent inflammatory response
following autologous transfusion into healthy volunteers.
The storage lesion of RBCs may resemble senescence, but could potentially
also include eryptosis.
• We aimed at explore the role of eryptosis in the storage lesion and then
investigate the splenic uptake of eryptotic cells after transfusion.
Although the biochemical changes described in stored RBCs may have an
impact on their survival after transfusion, it seems logical that there is
another level of the problem that needs to be understood in order to explain
the rapid loss of RBCs after transfusion, namely; what molecular changes to
the cell surface of the stored RBCs are recognized by macrophages and
eventually results in RBC phagocytosis?
• The final aim was to study the receptor-ligand interaction between liquid
stored RBCs and macrophages in order to better understand the
consequences of the storage lesion after transfusion.
20
Materials and methods
Human studies (paper 1)
Ten healthy male participants, age 22 to 44 years, with Hb in the range of
140-155 g/L, were included in the study. The subjects, most of them
committed to recreational training, were not allowed to compete in any
events associated with the Swedish sports confederation during the entire
time period of the study, including 4 weeks after reinfusion, due to the fact
that they would potentially have a competitive advantage over any opponent
related to the increase in oxygen carrying capacity after reinfusion. In
addition, 7 control participants, matching the inclusion criteria, were also
included for the evaluation of maximal aerobic capacity (VO2max). Control
participants, which did not donate or reinfuse RBCs, conducted the VO2max
tests at the same time points as the transfusion group.
Donation and reinfusion
Study participants donated one unit of blood (450 ml) on two occasions
separated by one week. Blood was collected in CPD, followed by leukocyte
and thrombocyte reduction, after which the majority of the plasma was
removed by centrifugation. One hundred ml of SAG-M was then added to the
remaining cells after which they were stored for 48 to 72 h before
cryopreservation. Glycerolization of RBCs was carried out using the ACP 215
automated system (Hemonetics, Inc., Tamarac, FL) by the addition of a 57%
glycerolyte solution (Fenwal, Lake Zurich, IL), after which the cells were kept
frozen (-80°C) for 15 to 16 weeks. RBCs were then thawed, deglycerolized
and washed, resulting in a post thaw recovery of ~80%. Reinfusion of RBC
units was carried out by transfusing each unit under approximately 30 min.
Venous whole blood samples from study participants were collected in
EDTA, gel or Li-Heparin containing tubes (BD Vacutainer, Franklin Lake,
NJ) at 2 h before, and at 2 and 48 h post transfusion for whole blood, serum
and plasma analysis, respectively.
Blood analysis
Whole blood samples were used to analyze blood cell counts and Hb
concentrations (Sysmex 2100, Sysmex, Kobe, Japan), while serum samples
were used in the analysis of bilirubin, transferrin and serum iron (Vitros 5.1,
Ortho Clinical Diagnostics, Inc., Raritan, NJ). Results from the two latter
were combined to calculate transferrin saturation [117]. Serum samples were
also used for the analysis of pro-, and anti-inflammatory cytokines, using a
21
cytokine bead array, custom designed to detect and quantify IL-1β, IL-6, IL8, IL-10, monocyte chemoattractant protein 1 (MCP-1), macrophage
inflammatory protein 1β (MIP-1β) and TNF-α (7-Plex kit, Bio-Rad, Hercules,
CA). Analyses were made according to the manufactures instructions, with
the addition of an extra concentration for the generation of the standard
curve in order to increase detection of cytokines of minute concentrations.
Plasma samples were used for the quantification of total haptoglobin by an
immunoturbidimetric assay (Cobas 6000, Roche Diagnostics Scandinavia
AB, Bromma, Sweden).
Maximal aerobic capacity (VO2max) assessment
Study participants conducted treadmill running to exhaustion on a constant
speed (individually set to be maintainable for 10 km) with a fixed stepwise
incline increase (0.5-1°) each minute. Oxygen consumption (VO2) was
measured on a Jaeger Oxycon Pro (Erich Jaeger GmbH, Hoechberg,
Germany). VO2max tests were performed before and after blood donation, as
well as before and (48 h) after reinfusion.
Animal studies (paper 2 and 3)
In the subsequent experiments, we used adult (8-25 weeks of age) male and
female CD47-/-, CD47+/-, or wild type (wt) Balb/c mice, or wild type
C57BL/6J mice. Mice were kept under low pathogen conditions and bred in
our local facility with free access to water and food in accordance with local
guidelines. All the experiments were performed in compliance with relevant
Swedish and institutional laws and guidelines and approved by the Umeå
research animal ethics committee (A14-12).
Labeling of RBCs
Labeling of RBCs before transfusion enabled the tracing of these cells after
transfusion. We have used two different methods to label mouse RBCs. First,
in vitro labeling with the fluorescent lipophilic dye PKH26 (according to the
manufacturer’s instructions, Sigma Aldrich, St. Louis, MO). The PKH26 dye
readily incorporates in the lipids of the RBC cell membrane [118]. Second, in
vivo biotinylation with biotin (N-hydroxysuccinimido-biotin, EZ-Link NHSBiotin, Thermo Scientific, Rockford, IL) which forms irreversible amine
bonds to membrane proteins with exposed amine containing side chains
(commonly lysine) and the N-terminal of proteins and polypeptides [106].
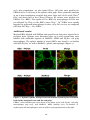
PKH26 labeling of RBCs was preferably used to identify splenic uptake of
transfused cells. Tissue sections of spleens from transfused animals were
22
incubated with fluorescently labeled ab:s against surface markers of
phagocytic cells in order to visualize specific cell populations, i.e. F4/80 on
red pulp macrophages, MARCO on MZM, MOMA-1 on MMM and CD11c on
dendritic cells. Sections were then analyzed by laser scanning confocal
microscopy (Leica TSP-2, Heidelberg, Germany).
We used biotin-labeled cells in survival experiments, where blood samples
could be drawn from mice at different time points after transfusion. Blood
samples were then incubated with fluorochrome-conjugated streptavidin,
which forms a strong bond to biotin. In this way, the fraction of the labeled
cells, i.e. the transfused cells, could be monitored over time by flow
cytometry (FACS Calibur, BD Biosciences, San Jose, CA). In vivo
biotinylation was carried out by tail vein injection of 0.4 to 1.5 mg of biotin,
dissolved in 150 µl of sterile PBS. This efficiently labeled all circulating cells.
The mice were sacrificed the following day, and blood (with >98% biotinlabeled cells) collected by heart puncture. This blood was used for storage
and transfusion. The double in vivo biotinylation technique enabled
discrimination between different age fractions within the RBC population
[119]. This was made possible by the injection of 1.1 mg of biotin per day for
three days and then by waiting until half of the RBC population had been
regenerated (~25 days), after which a lower dose of biotin (0.4 mg) was
injected. This resulted in half of the RBCs binding higher levels of biotin
(biotinhi; the older fraction) and the other half binding intermediate levels of
biotin (biotinint; the young fraction). The two fractions of biotinylated cells
can easily be discriminated individually from biotin negative RBCs of the
recipient mice after transfusion.
We also labeled human RBCs with FLUO-3 (Molecular Probes) in order to
quantify intracellular Ca2+ uptake. This was done by incubating RBCs with
FLUO-3 in Ca2+-free media for 1 h in 37°C. Cells were then washed and later
suspended in Ca2+-containing media, followed by incubation for 1 h in 37°C.
Fluorescence intensity was then determined by flow-cytometric analysis
(FACSCalibur flow cytometer, BD Biosciences).
The mouse model for RBC storage and transfusion
Whole blood was collected in CPD (blood to CPD-ratio - 7:1) by heart
puncture of euthanized mice and spun at +4°C and 500 x g for 5 min. Plasma
was then transferred to a separate tube, while the buffy coat and the top
layer of the RBCs were discarded. Plasma was spun at +4°C and 1300 x g for
10 min, in order to remove any residual cells, after which the supernatant
was transferred into a new tube. The RBCs were washed 3 times in PBS and
finally mixed with SAG-M and plasma (ratio 3:1) and either used for direct
23
transfusion or stored for 10 days at +4°C in 0.5 ml test tubes. For Phthalate
experiments, test tubes were coated in plastic film containing DEPHphthalates.
In eryptosis experiments, the RBCs were incubated with the calcium
ionophore ionomycin. For this, RBCs were suspended in Krebs-Ringer
Hepes (KRH) medium at a hematocrit of 0.5% with 1µM ionomycin for 1 h at
37°C, followed by 3 washes in PBS. Ionophore-treated RBCs were finally
resuspended in sterile PBS at a hematocrit 0.5% for phagocytosis
experiments, or at 20% hematocrit for transfusion experiments.
Mice anesthetized with isofluorane (Baxter, Deerfield, IL) were transfused
with 150 µl of fresh or 10 day stored RBCs, administered by tail vein
injection. Blood samples from the opposite tail vein were taken at multiple
time points after transfusion and analyzed by flow cytometry for the
presence of transfused cells as described above.
In vitro characteristics of stored RBCs
To elucidate if the mouse model for RBC storage shared characteristics with
that of human stored RBCs, in vitro measurements on intracellular ATP and
the formation of MPs were conducted. For ATP measurements, 5 x 107
murine RBCs, stored for 0, 4, 7, or 10 days, were lysed in hypotonic buffer.
An ATP standard curve was established by diluting ATP samples of known
concentrations. Samples were mixed with ATP-reagent, containing luciferin
and luciferase, followed by analysis of peak luminescence using a Triathler
luminometer (Hidex, Turku, Finland). For MP quantification, fresh and 10
day stored murine RBC concentrates were labeled with fluorescein
isothiocyanate (FITC)-conjugated anti-mouse Ter-119 (ImmunoTools,
Freisoythe, Germany) and analyzed for MPs by logarithmic scaling for FSC
and SSC on a FACSCalibur (BD Biosciences) flow cytometer.
In vitro phagocytosis
Mouse peritoneal macrophages were used for phagocytosis experiments,
basically as described elsewhere [120]. Fresh or stored human and mouse
RBCs, or eryptotic mouse RBCs, were then incubated together with adherent
macrophages for 1 h at 37°C, 5% CO2. Incubation was carried out with or
without the addition of 10% FCS or human serum (HS). The effect of the
class A-SR-inhibitors fucoidan and dextran sulphate, or the control
substance chondroitin sulphate, was also evaluated. For signaling inhibitor
experiments, macrophages were pre-incubated for 30 min in the presence or
absence of the phosphatidylinositide 3-kinases (PI3-kinase)–inhibitor
24
LY294002 (50µM), the P38 mitogen-activated protein kinase (p38 MAPK)inhibitor SB203580 (30 µM), or the extracellular signal-regulated kinase-1/2
(ERK1/2)-inhibitor PD98059 (30 µM). When phagocytosis was terminated,
non-ingested RBCs were lysed with dH2O for 30 sec, followed by washing,
fixation and staining, using May-Grunewald/Giemsa. In every experiment, at
least two technical replications per condition were made. Macrophages with
ingested RBCs were then counted to generate a phagocytosis index by
dividing the number of phagocytized RBCs with the total number of
macrophages.
Ethics
The human transfusion study was conducted in accordance with the
declaration of Helsinki and approved by the Swedish central ethics
committee (Dnr Ö 1-2009). For in vitro experiments, human RBCs were
acquired from blood bags (containing saline-adenine-glucose-mannitol
(SAG-M), kept at a constant temperature of +4˚C and stored in the normal
deposit at the Blood Bank at Norrlands University Hospital (Umeå, Sweden).
A part of a plastic hose, filled with ~1 ml RBCs, was welded off from blood
bags stored for up to six weeks. The acquired samples were anonymized and
carried only information of blood group and the date of donation. Since the
results obtained in the present study could not be traced back to a specific
blood donor, ethical approval was not needed according to Swedish
legislation (41, SFS 2003:460). All animal experiments were approved by
Umeå research animal ethics committee (A14-12).
Statistical analysis
Two tailed student’s paired and unpaired t-test was used for comparisons
between samples and groups respectively, when normal distribution could be
assumed. Wilcoxon´s related samples signed rank test were used of paired
comparisons when data were not normally distributed (IBM SPSS statistics
Base 19, IBM, Armonk, NY, USA and JMP 11.0, SAS Campus Drive, Cary,
NC).
25
Results
Paper 1
Transfusion of liquid-stored RBCs has been associated with worsened
outcomes after transfusion [92, 121]. In addition, an animal study has shown
that transfusion of the equivalent of 2 units of liquid-stored RBCs resulted in
a profound proinflammatory response, suggested to be the result of an
increased RBC uptake by macrophages shortly after transfusion [101]. To
investigate the response to transfusion of cryopreserved RBCs in humans, we
transfused 2 units of cryopreserved RBCs into 10 healthy human volunteers.
Indicators of erythrophagocytosis, inflammation and the number of
leukocytes in blood were assessed at 2 h and 48 h after transfusion and
compared with that at 2 h before transfusion.
Transfusion of cryopreserved RBCs leads to acute accelerated
RBC breakdown
As expected, these RBC transfusions resulted in a significant increase in
blood Hb. The number of circulating neutrophil granulocytes was
significantly higher at 2 h and 48 h after transfusion. This was accompanied
by an increase in circulating monocytes at 48 h after transfusion, thus
indicating a transfusion-mediated effect on the immune system. We also
found evidence of an acute clearance of transfused RBCs as the serum iron
concentration, the transferrin saturation and the serum bilirubin
concentration were all significantly increased at 2 h post transfusion, but had
then returned toward baseline at 48 h. We further noticed a decrease in
serum haptoglobin levels at 2 h after transfusion, which again returned
towards baseline levels at 48 h, indicating that part of the hemolysis was in
fact intravascular.
Absent pro-inflammatory response in the acute phase after
transfusion
After concluding that an acute hemolysis occurred after transfusion, we next
investigated if there were any signs of inflammation. For this, we screened
for known pro-inflammatory cytokines; IL-1β, IL-6, IL-8, MCP-1, MIP-1β,
TNF-α, and the anti-inflammatory cytokine IL-10. None of the investigated
cytokines increased in serum samples, neither at 2 h nor at 48 h after
transfusion.
26
Change from initial value (%)
Additional results
Paper 1 is based on data that originates from a larger study aimed at
evaluating the enhanced aerobic capacity as a result of autologous
transfusion and to identify biological markers of blood doping. Maximal
aerobic capacity (VO2max) was evaluated before and after blood donation as
well as reinfusion. Donation of 900 ml of blood led to a significant reduction
in VO2max of 8±3%. Surprisingly, this reduction was not compensated for
during the 15 weeks of recovery after donation even though Hb
concentration was restored (Figure 6). In contrast, reinfusion of the
cryopreserved RBCs led to a 13±8% increase in VO2max, close to the starting
value.
Maximal oxygen uptake (VO2max)
4
2
0
-2
-4
-6
*
-8
-10
-12
R
-14
0
Change from initial value (%)
*
D
2
4
10
6
8
10
12
14
16
Blood Hb concentration
5
0
-5
D*
-10
Control
Transfusion
R
-15
0
2
4
6
8
Week
10
12
14
16
Figure 6. Restoration of [Hb] 15 weeks after donation does not result in
restored maximal aerobic capacity (Vo2MAX).
VO2MAX and blood Hb concentration was measured before and after blood donation
as well as reinfusion. Values for control group (dotted line, n=7) and transfusion
group (continuous line, n=9), are given as means±SEM. Unpaired student’s t-test
was used for group comparisons. Indicated is the time of blood donation (D) and
time of RBC reinfusion (R).
27
Paper 2
Eryptosis, a Ca2+ dependent stress-induced cell death in RBCs, results in a
shorter than normal lifespan and could theoretically be a part of the RBC
storage lesion. However, the understanding on how eryptotic RBCs are taken
up by phagocytic cells in the spleen is virtually unknown. Thus, this study
aimed at first investigating if there were any signs of eryptosis in liquid store
RBCs, and second to investigate what phagocytic populations of the spleen
that mediate uptake of eryptotic cells.
Stored human RBCs contain a subset of Ca2+-high cells
It is currently unknown if eryptosis plays a role in the storage lesion. Because
calcium uptake is a hallmark of eryptosis, we tested if liquid stored human
RBCs display an increased Ca2+ permeability. RBCs, stored in SAG-M for 0,
2, 4 and 6 weeks were labelled with the FLUO-3 calcium indicator followed
by incubation in Ca2+ containing medium for 1 h at 37°C. At the beginning of
storage, human RBCs stored in SAG-M contained a low fraction of cells with
an increased cytoplasmic Ca2+-level (0.7±0.3%). However, we found a timedependent increase in the fraction of cells with increased Ca2+, reaching
3.6±0.8% after 6 weeks of storage. This suggests that part of the storage
lesion could be associated with some RBCs having an eryptotic phenotype.
Characteristics of eryptotic murine RBCs
A mouse model was used to investigate the clearance of transfused eryptotic
cells, as this is difficult to assess in humans. It was found that RBCs treated
with 1 µM ionomycin increased their intracellular Ca2+ concentration and
that around 50% of the cells were PS+ as accessed with Annexin V binding.
This was accompanied by a reduction in cell size as seen by lower FSC in flow
cytometric analysis.
In vivo uptake of Ca2+-RBCs is located to the splenic marginal
zone in a CD47-independent manner
Transfusion of untreated RBCs, labeled with PKH26 or biotin, resulted in a
rather linear clearance over the entire expected lifespan of 50 days, where
the splenic uptake of these cells was located to F4/80+ macrophages in the
red pulp area . The fact that the clearance kinetics of PKH26 or biotin labeled
cells was virtually identical indicated that the labeling per se had no effect on
the removal of the transfused cells. In contrast to that for freshly isolated
RBCs, transfusion of PKH26-labeled Ca2+-RBCs (ionophore treated) resulted
in a swift clearance of the majority of the transfused cells within 30 minutes.
The splenic uptake of Ca2+-RBCs was located to the MZ, and appeared to be
preferentially mediated by MARCO+ MZ macrophages (MZM). Uptake of
Ca2+-RBCs was also seen in CD11c+ DCs in the MZ at 1 h after transfusion. At
28
24 h after transfusion, we also found CD11c+ DCs that were positive for
PKH26 in the T-cell area of the splenic white pulp. Flow cytometric analysis
at 24 h after transfusion revealed that more than 20% of the total CD11chi
DCs, and about half of the CD11chi/CD207hi DC subset, were positive for
PKH26+ Ca2+-RBCs. The uptake of Ca2+-RBCs by macrophages or DCs was
not affected by CD47 on the RBCs since CD47-/- Ca2+-RBCs were equally
ingested by peritoneal macrophages in vitro, or by DCs in vivo, as compared
with that for CD47+/+ Ca2+-RBCs.
Additional results
Murine RBCs, labeled with PKH26 and stored for 10 days were injected in to
recipient mice. Spleens were harvested after 24 h and cryosections were
labeled with antibodies against of MARCO+ MZM and F4/80+ red pulp
macrophages. The splenic uptake of stored RBCs was found to co-localize
with both F4/80+, as well as MARCO+, splenic macrophages (Figure 7.)
Figure 7. Splenic uptake of liquid stored murine erythrocytes is located
both in the marginal zone and the red pulp.
PKH26+ stored erythrocytes were shown to be taken up by both F4/80+ red pulp
macrophages (top row), and MARCO+ MZM (bottom row). Co-location of
erythrocytes and macrophages is seen as yellow on merged pictures to the right.
29
Paper 3
Much is still unknown regarding the storage lesion and the mechanisms
mediating rapid RBC elimination after transfusion. The lack of detailed
knowledge can in part be explained by ethical limitations in human studies.
To circumvent these limitations, we have adopted a murine model of RBC
storage and transfusion. This model was designed to mimic that of the
human setting as closely as possible, e.g. by using CPD and SAG-M in the
collection and storage of the RBCs. In addition to the in vivo model of RBC
transfusions in mice, we also investigated if scavenger receptors were
involved in the phagocytosis of stored murine or human RBCs, using an in
vitro mouse macrophage phagocytosis assay.
Characteristics of the murine model for storage and transfusion
Murine RBCs have a shorter lifespan and are generally considered as more
fragile than human RBCs [116], which was also reflected in the shorter
storage time needed to induce the storage lesion (10 days for mouse RBCs
and 42 days for human RBCs). At this time point, stored murine RBCs
displayed similar characteristics as previously described for stored human
RBCs, including about 50% reduction in ATP content and formation of MP.
In addition, about 35% of the stored murine RBCs were eliminated from the
circulation within 24 h, while the RBCs surviving this time period showed
normal long-term clearance kinetics after transfusion.
Phagocytosis of stored human and mouse RBCs is serum
dependent and mediated by scavenger receptors
The initial rapid clearance kinetics of stored RBCs likely represents
molecular changes caused to a fraction of the RBCs during storage, which are
efficiently recognized by macrophages leading to phagocytosis after
transfusion. We hypothesized that these changes may not be speciesspecific, but rather that the recognition mechanisms could be part of the
evolutionary conserved systems that identify damaged cells. To test this
hypothesis, both human and murine RBCs (fresh or stored) were incubated
with murine peritoneal macrophages in an in vitro phagocytosis assay. When
macrophages were incubated with fresh RBCs, neither murine nor human
RBCs were phagocytized. In contrast, stored murine and human RBCs were
readily phagocytized by macrophages in the presence of heat inactivated FCS
(and heat inactivated HS for human RBCs). Interestingly, phagocytosis of
stored murine or human RBCs was markedly reduced by 70-75% under
serum-free conditions. To investigate if phagocytosis of stored RBCs was
dependent on class A-SR, we studied phagocytosis in the presence of the
class A-SR ligands fucoidan and dextran sulphate. The results showed that
both fucoidan and dextran sulphate inhibited phagocytosis of stored murine
30
or human RBCs by more than 90%. Phagocytosis of stored RBCs was also
significantly reduced in the presence of the PI3 kinase-inhibitor LY294002.
In addition, the ERK1/2-inhibitor PD98059 and the p38MAPK-inhibitor
SB203580 each reduced phagocytosis of stored RBCs by 30% and 36%,
respectively.
Additional results
Discrimination of RBC age fractions by in vivo biotinylation
Double in vivo biotinylation was carried out with 25 days in between biotin
injections. This enabled the RBCs to be separated into age fractions. Biotinhi
cells represented RBCs >25 days old and biotinint cells represented the
remaining RBCs which were <25 days old (figure 8A). On day 26, the mice
were bled and RBCs were isolated before transfusion into recipient mice.
Blood samples from transfused mice showed a close to linear clearance of
>25 days old biotinhi cells during the first 25 days after transfusion. The <25
days old biotinint RBCs were, as expected, initially cleared slower and could
be detected up to 55 days after transfusion (figure 8B).
B. 120
>25 days old
<25 days old
>25 days old
<25 days old
Total
100
RBC recovery (%)
Streptavidin-PE
A.
80
60
40
20
0
0
FSC
20
40
60
Time (days)
Figure 8. The use of biotin to label circulating erythrocytes in donor
mice and subsequent analysis of their clearance following transfusion
into recipient mice.
A, Donor mice were injected i.v. with 1.1 mg biotin at day 0, 1 and 2. At day 25,
another biotin injection of 0.4 mg was made. On day 26, mice were bled,
erythrocytes were stained with streptavidin-PE and analyzed by flow cytometry. A
representative dot-plot shows that >25 days old biotinhi erythrocytes can be
separated from <25 days old biotinint erythrocytes. B, Recovery of freshly isolated
>25 days old (closed symbols) or <25 days old RBCs (open symbols) after
transfusion into syngeneic recipient mice. Grey symbols represents recovery of all
transfused RBCs. Data are mean±SD of three separate recipient mice.
31
Role of the RBC age at the start of storage for their 24 h survival post
transfusion
RBCs from double in vivo biotinylated mice, freshly isolated or stored for 10
days, were transfused into recipient mice in order to investigate if RBCs >25
days old at the start of storage would be more severely affected by storage
and therefore eliminated to a larger extent during the first 24 h after
transfusion. The 24 h recovery of freshly transfused RBCs was high among
both younger and older fractions (figure 9 A and B). Interestingly, when 10
days stored RBCs were transfused, we found that virtually all cells
disappearing rapidly within the first 24 h belonged to the fraction of RBCs
>25 days old at the start of storage (figure 9 B). When comparing the
survival kinetics of the fraction of >25 days old freshly transfused or stored
RBCs that remained in circulation at 24 h after the transfusion, we found
that old fresh and stored RBCs had virtually similar recovery and expected
life span (figure 9 C).
B
Young RBC
120
Fresh
Stored
80
60
40
Fresh
Stored
RBCs remaining after 24 h
120
Old Fresh
100
100
RBC recovery (%)
RBC recovery (%)
100
C
Old RBC
120
RBC recovery (%)
A
80
60
40
Old stored
80
60
40
20
20
20
0
0
0
0
10
20
30
40
Time (days)
Figure 9. Virtually all of the stored murine RBCs cleared within 24 h
belong to the fraction of older RBCs.
Murine RBCs were labeled twice with biotin in vivo to enable separation of RBCs
with an age of <25 days (young) or >25 days (old). Isolated RBCs were either
freshly transfused (Fresh) or stored in SAG-M and autologous plasma at +4°C for
10 days before transfusion. (A) Recovery of fresh or stored younger RBCs, or (B)
fresh or stored older RBCs, at 24 h after transfusion. (C) Recovery kinetics of older
fresh or older stored RBCs, where the amount of each RBC fraction remaining in
circulation at 24 h after transfusion was set to 100 per cent. Data are means±SD of
4 separate recipient mice.
Phthalates during storage
Plastic softeners or phthalates are known to be released from blood bags in
to the membranes of the stored RBCs [122-124]. Trying to investigate a
potential effect of phthalates in our murine storage model, RBCs were stored
in test tubes coated with or without phthalate containing PVC-plastic film.
32
Transfusion of phthalate stored RBCs demonstrated near identical 24 h
survival characteristics as RBCs stored without phthalates (Figure 10A.) This
finding could indicate that plastic softeners may not affect murine RBCs
during storage with respect to their chances of surviving after transfusion.
CD47 on stored murine RBCs does not influence the rapid elimination that
occurs during the first 24 h
The cell surface protein CD47 has been shown to serve as an anti-phagocytic
ligand, protecting RBCs from being phagocytized by macrophages [52].
Furthermore, the amount of CD47 on the cell surface has been implicated to
decrease during storage of human RBCs [83]. To test if a reduction in CD47
on stored RBCs could affect the rate of rapid 24 h elimination after
transfusion, stored RBCs from Wt (CD47+/+) or CD47+/- mice (in which the
CD47 expression is 50 % of that in CD47 Wt) were transfused into Wt mice.
The 24 h survival was nearly identical when comparing Wt and CD47+/RBCs, indicating that CD47 may not be involved in regulating the acute
phagocytosis of stored erythrocytes (figure 10B).
A
B
120
100
RBC recovery (%)
100
RBC recovery (%)
120
80
60
40
80
60
40
20
20
Phthalates
Control
CD47+/- RBCs
0
0
0
4
8
CD47+/+ RBCs
12
16
20
0
24
4
8
12
16
20
24
Time (hours)
Time (hours)
Figure 10. Acute clearance of stored murine RBCs is not affected by
phthalates of RBC CD47.
(A) RBCs were stored in the presence or absence of phthalates before transfusion
to syngenic mice. Data are means±SD of 2 separate recipient mice. (B) Transfusion
of stored Wt (CD47+/+) or CD47+/- RBCs displays near identical RBC recovery
during the first 24 h after transfusion Data are means±SD of 4 separate recipient
mice.
33
Discussion
Frozen vs. liquid stored RBCs
At present, the vast majority of RBC concentrates are stored under liquid
conditions [125]. Cryopreservation of RBCs are mainly restricted to donors
with rare blood groups and for autologous transfusions [126]. Cryopreserved
RBCs have been shown to be spared from many of the hallmarks of the liquid
storage lesion including a reduction in ATP and 2,3-DPG and the formation
of MPs [87-89]. Since little is known about the clinical consequences of the
storage lesion, it is problematic to evaluate the two storing methods based on
in vitro characteristics, but the phenotype of cryopreserved RBCs suggest
that the method may have advantages over liquid storage. On the other
hand, the time consuming procedure (3-4 hours) of thawing RBCs limits its
use to planned procedures. Our data indicated that transfusion of two units
of cryopreserved RBCs had a modest effect on the Hb concentration, which
was only significantly elevated at 2 h after transfusion (from 145.4±4.9 g/L
to 151.1±4.6 g/L). After 48 h, it had dropped 3 g/L and was no longer
significantly different from the Hb concentration before transfusion. On the
other hand, aerobic capacity assessment at 48 h after transfusion, revealed a
13% increase in VO2max as compared to values obtained the week before
transfusion (Figure 6). Thus, differences in the venous Hb concentration
may not be the best assessment of neither RBC volume nor aerobic capacity
after RBC transfusion to non-anemic men, possibly explained by the capacity
of the spleen to store RBCs [127, 128].
Inflammation after transfusion?
There is at present a controversy regarding risks with transfusion of RBCs
stored for prolonged periods of time [92, 93]. Alarming results from animal
studies have also emerged, in were Hod et al. [101] could show a profound
pro-inflammatory cytokine storm and the presence of NTBI 2 h after
transfusion of the equivalent of two units of stored RBCs into mice. The
cytokine release was suggested to originate from the large amount of iron
delivered to macrophages upon the rapid phagocytosis of a fraction of the
transfused RBCs [103]. However, no clear evidence has been presented for
such a response in humans, with the exception of a report in preterm infants
[129]. To test if increased erythrophagocytosis alone could provoke such a
response in healthy humans, we conducted experiments where human
subjects received two units of cryopreserved RBCs. Although the storage
conditions was not the same (frozen vs. liquid stored), it has been shown that
approximately 15 % of the previously frozen, transfused RBCs are rapidly
34
cleared from circulation upon transfusion [88, 130]. Indeed, our results also
showed a marked increase in serum bilirubin, iron and transferrin saturation
at 2 h post transfusion, which all indicate increased erythrophagocytosis.
However, the rise in transferrin saturation (from 28.1 ±1.3 % to 43.2 ±1.5 %)
did not indicate that non-transferrin bound iron were present in the plasma
since most of the transferrin were still not saturated with iron. It can
however not be ruled out that the peak of serum iron came about after the
measurement at 2 h post transfusion and that NTBI might have been present
at a later time point. A human study on transfusion of one unit of liquid
stored RBCs, stored for 42 days, to healthy recipients showed a larger
increase in transferrin saturation than observed in our study [104]. That
study also detected NTBI in plasma after transfusion, indicating that there
could be a difference as to how frozen and liquid stored RBCs are received
after transfusion [104]. This discrepancy is probably related to the storage
lesion which primarily is restricted to liquid stored RBCs.
To elucidate if the hemolysis was primarily intra- or extra-vascular (i.e. if the
red blood cells burst in circulation or were phagocytized in a controlled way
by macrophages), we measured the levels of haptoglobin in serum.
Haptoglobin binds free Hb and a diminished plasma level of haptoglobin is
an indirect marker for intravascular hemolysis, although profound
extravascular hemolysis could also lead to reduced haptoglobin levels [131].
We found that haptoglobin was reduced by 0.18 g/L at 2 h after transfusion,
which corresponds to the binding of approximately 0.8 g of Hb [132, 133].
Since the total amount of Hb transfused was about 80 g, intravascular
hemolysis only accounted for 1% of the transfused RBCs, strongly indicating
that the majority of the hemolysis was in fact extravascular. After concluding
that we had an acute, extravascular hemolysis, we investigated the presence
of pro-inflammatory cytokines in the plasma of transfused individuals, in
search of similarities with findings in the above mentioned animal study.
None of the investigated pro-inflammatory cytokines IL-1β, IL-6, IL-8, MCP1, MIP-1β or TNF-α increased in plasma at 2 h or 48 h after transfusion.
Similar findings were also reported after transfusion of one unit of liquid
stored RBCs in healthy recipients [104]. The discrepancy in the data
concerning signs of inflammation between animal and human studies could
be due to differences in the storage and/or administration procedure, but
also due to species differences [112].
The mouse model for storage and transfusion
In order to expand our understanding on post transfusion events related to
the storage lesion, we turned to an animal model. There are always
drawbacks when using model systems, and one must be cautious when
35
transferring findings in animal models to human physiology [112].
Nevertheless, due to ethical considerations, an animal model can present
opportunities to study mechanisms that are not easily assessed in human
subjects. The use of inbred mice is an attractive alternative due to practical
considerations and the genetic kinship between man and mouse [111].
Together with the increasing availability of genetic manipulations, the mouse
model offers unique possibilities compared to humans investigations.
The use of a mouse model in RBC storage and transfusion has been shown to
resemble human settings [134], albeit with shorter storage time of the RBCs
prior to transfusion. As expected, we observed that 10 days of storage
resulted in an ATP reduction, MP formation and 24 h survival kinetics
resembling that found in human RBCs after prolonged storage. Human
RBCs are stored in PVC-plastic bags that contain plasticizers (phthalates),
known to leak into the membranes of the RBCs and thus improve their
storage characteristics as they reduce the hemolysis incidence [122-124]. In
an attempt to evaluate the effect of phthalates on mouse RBC storage, we
coated the storage test tube with PVC-plastic film prior to the addition of the
cells. RBCs stored in phthalate plastic showed similar 24 h clearance kinetics
as control RBCs stored without phthalates (Figure 10A), indicating that
plasticizers may not have an major influence on the 24 h survival of stored
mouse RBCs after transfusion in our model.
The role of calcium uptake in the storage lesion
Increased intracellular Ca2+ is a hallmark of eryptotic cells, which leads to PS
externalization and the subsequent uptake by phagocytic cells [40]. The
observation that a small fraction of RBCs had an increased Ca2+ permeability
after 6 weeks of storage suggests that eryptosis could be a part of the RBC
storage lesion. It is not known if the fraction of Ca2+ RBCs could further
increase after transfusion, but the observation that human stored RBCs
starts to externalize PS after overnight incubation at 37°C suggests the
possibility that transfused cells may acquire additional removing signals
after transfusion [135].
The spleen consists of distinct populations of phagocytic cells with different
functions and locations [56]. By studying the splenic uptake of
experimentally induced eryptotic cells after transfusion, we found that the
vast majority of RBCs with increased Ca2+ were taken up in the marginal
zone (MZ), preferentially by MARCO+ MZM. This is in contrast to the uptake
of normally senescent RBCs, which generally are phagocytized by F4/80+ red
pulp macrophages. One explanation to the different micro-anatomical
locations of phagocytosis could be that a substantial amount of the arterial
36
blood in the murine (but also human) spleen flows through the MZ before
entering the red pulp, placing phagocytic cells of the MZ first in line to
recognize the eryptotic RBCs. The uptake of stored murine RBCs was found
to be located both in the marginal zone and in the red pulp (Figure 7) and as
such, agrees with the hypothesis that a part of the storage lesion could in fact
be explained by eryptosis. However, it also supports the view that the storage
lesion is multifactorial, and that other RBC changes lead to their recognition
by the red pulp macrophages.
Impact of storage on RBCs - Senescence, eryptosis, or both?
To further understand the effect of the storage lesion, we applied the double
in vivo biotinylation technique [119], in which RBCs in circulation can be
separated with regard to their age (Figure 8). This grants the ability to follow
younger and older RBCs separately, in order to evaluate the effect of the
storage lesion in separate age fractions. Interestingly, we found that almost
the entire loss of transfused stored RBCs during the first 24 h involved the
fraction of older RBCs (>25 days of age prior to storage), which strongly
indicates that the storage lesion has a more profound effect on aged RBCs
(Figure 9A, B). Markedly, the older cells that remained in circulation after 24
h showed similar clearance kinetics as freshly transfused RBCs of the same
age (Figure 9C). This therefore suggests that the storage lesion could have a
more profound effect on older RBCs, and that cells surviving the first 24h
after transfusion seem to be rather unaffected by the storage lesion.
It is known that some of the RBCs having reduced levels of ATP and 2,3-DPG
after storage may restore these levels within 24 h after transfusion [85]. Such
recovery could theoretically also be connected with restoring some of the
RBC cell surface changes that are involved in macrophage recognition and
clearance. Furthermore, it has been suggested that the shedding of PS+ MPs
from stored RBCs could reduce the amount of the externalized PS on the
RBCs and thus reduce the risk of phagocytosis of these cells [135]. To
investigate the possibility that some of the stored RBCs, which are normally
rapidly eliminated after transfusion, may have the capacity to recover in the
absence of macrophages, one could deplete recipient mice of macrophages in
the spleen and liver before transfusion. The efficient depletion of
macrophages in contact with the bloodstream by i.v. injection of clodronate
liposomes has been demonstrated [136]. An improvement in 24 h recovery of
stored RBCs in clodronate depleted macrophages have also been shown
[137]. By transfusing biotin-labeled stored RBCs, we could investigate how
different age fractions among the transfused cells remain in circulation over
time until macrophages start to return around day 6, and elucidate if all of
the rapidly cleared transfused RBCs are taken up by macrophages.
37
Transfused RBCs remaining in circulation can also be recovered by FACSsorting for further biochemical analysis.
How are stored RBCs presented to the immune system upon
transfusion?
DCs are professional APCs that trap foreign or endogenous antigens by
phagocytosis or macropinocytosis. The antigens are then intracellularly
processed, attached to MHC, and finally presented on the cell surface
together with costimulatory molecules (Figure 2). Depending on the
phenotype of the DC, and the recognition pattern of the antigen, the DC will
present the antigen in either an immunogenic or tolerogenic way [15, 138].
The balance between tolerance and immunity can further be affected by the
inflammatory state of the organism. Alloimmunization has been shown to be
propagated after RBC transfusion of transgenic hen egg lysosome (HEL)RBCs in mice, but only after co-stimulation with the immunostimulant
poly(I:C) [139]. In contrast, the administration of LPS together with HELRBC transfusion not only failed to increase alloimmunization, but was also
found to inhibit the same process [140]. Thus, inflammation as such may be
a too broad of a label to use when assessing risks of transfusions since
(mimicked) viral or bacterial inflammatory stimuli seem to have quite
different effects on immunomodulation in relation to alloimmunization, at
least in mice. This also highlights the importance of finding the receptor(s)
involved in recognition of stored RBCs by the immune cells.
A subset of DCs which express the cell surface proteins CD103 (integrin αE)
and CD207 (langerin) (CD207+ DCs) has been identified as responsible for
the uptake of apoptotic blood borne cells in the marginal zone of the (mouse)
spleen [141]. Their function is to maintain self-tolerance by migrating to the
T-cell area of the spleen and present self-antigens to naïve T-cells. Depletion
of MZ phagocytes has been shown to result in loss of immunosuppression
against apoptotic cells [142]. After stimulation with either poly(I:C) or LPS,
the number of CD207+ DCs was found to be drastically reduced in murine
spleens [141]. The finding in paper 2 of the present thesis, that stored human
RBCs may contain a population of eryptotic cells and that experimentally
eryptotic RBCs were recognized and taken up by CD207+ DCs in the spleen,
therefore opens the door for a somewhat unexpected possibility: that part of
the storage lesion may in fact have beneficial effects in terms of propagating
tolerance rather than immunity. Inflammatory stimuli are likely to interfere
with maintenance of this tolerance, again bringing the role of inflammation
after transfusion of stored RBCs to life. In a next step, it would be intriguing
to transfuse PHK26+ stored RBCs, collect spleens and (by flow cytometry)
determine to which degree different macrophage and DC subsets ingest
38
stored RBCs. Different activation markers on PKH26+ phagocytic cells can
then be assessed in order to understand the immunologic impact of
transfusion of stored RBCs.
CD47 in senescence, eryptosis and the storage lesion
The presence or absence of CD47 on experimentally eryptotic RBCs affected
neither macrophage phagocytosis in vitro nor DC uptake of these cells in
vivo. There have been indications in the literature that senescent, and
possibly also stored RBCs, may have reduced amounts of CD47 on their
surface [36, 83]. We found that CD47+/- RBCs (expressing half of normal
CD47 levels) had virtually identical clearance kinetics as normal CD47+/+
RBCs after storage and transfusion (Figure 10B). The long-term survival of
CD47+/- RBCs was on the other hand slightly reduced as compared to
CD47+/+ controls (data not shown). This could support the hypothesis that
senescent RBCs are recognized by phagocytic cells after opsonization by
NAbs, since IgG-opsonized CD47+/- RBCs were found to be cleared more
rapidly after transfusion than equally IgG-opsonized CD47+/+ RBCs in vivo
[143]. We have not specifically investigated the effect of CD47 in our
phagocytosis assay of stored RBCs. However, since macrophage phagocytosis
of stored RBCs could be efficiently blocked by addition of SR inhibitors , and
SR-mediated phagocytosis of oxidatively damaged RBCs has been shown to
be CD47 independent [120], there are no strong indications from our studies
that CD47 (or lack thereof) is influencing macrophage recognition of stored
RBCs.
Furthermore, the CD47-SIRPα interaction has been shown to be dependent
on RBC membrane flexibility, which could imply that the ability of CD47 to
inhibit phagocytosis could be further limited in senescent or stored RBCs
[144]. In addition, an alternative role of the CD47-SIRPα interaction has
been presented by Burger et al. [145], who showed that the serum protein
thrombospondin-1 (TSP-1) could bind to CD47 on senescent or stored RBCs
to promote their phagocytosis. A conformational change in CD47 enabled
binding of TSP-1 which resulted in SIRPα-dependent phagocytosis. This
indicates that the CD47-SIRPα interaction can have both pro-, and antiphagocytic properties, thus making quantification of total CD47 per se an
unsuitable indicator to predict the uptake of senescent and/or stored RBCs.
Scavenger receptor mediated phagocytosis of stored RBCs
Scavenger receptors are evolutionary conserved receptors that can mediate
endocytosis and phagocytosis of molecules or cells carrying certain
molecular signatures [13]. Fucoidan and dextran sulphate are ligands for
39
class A-SRs and can efficiently block binding of other ligands to these
receptors [146]. Interestingly, we found that both fucoidan and dextran
sulphate strongly inhibited phagocytosis of stored human as well as murine
RBCs in vitro. Thus, class A-SRs may be a previously unappreciated class of
receptors involved in macrophage phagocytosis of RBCs damaged by storage.
It has previously been shown that fucoidan and dextran sulphate can inhibit
phagocytosis of oxidatively damaged murine RBCs [120]. Inhibition of the
signal transduction proteins PI3-kinase, p38-MAPK and ERK1/2 was shown
to affect phagocytosis of IgG opsonized or oxidized RBCs [120]. Since we also
found that the same inhibitors reduced the uptake of stored RBC, this could
suggest that oxidative stress may in fact be one important contribution to the
storage lesion with respect to the 24 h recovery of transfused cells. PI3kinase is essential for phagocytosis of endogenous material as well as
pathogens [147], and has been suggested to also regulate phagocytosis of
stored RBCs [137]. Scavenger receptor class A-1 (SRA-1 or CD204) lacks a
signaling cytoplasmic domain and may only mediate binding of damaged
RBCs to the macrophage, while the intracellular signaling resulting in
phagocytosis could be contributed by other receptors binding to the same
RBC [148]. Ligation of fucoidan to SRA-1 has been shown to enhance TLR-4
mediated activation of nuclear factor-κβ (NF- κβ), resulting in TNF-α and
IL-1β production by LPS stimulated murine peritoneal macrophages [149].
Ligation of fucoidan alone did not, however, mediate an inflammatory
response. Importantly, the impact of this recognition mechanism in vivo, as
well as in the macrophage subpopulations normally involved in elimination
of stored RBCs in spleen or liver, has to be further investigated in both the
murine and human system.
The role of serum in phagocytosis of stored cells
Both human and murine stored RBCs required heat-stable serum factors for
efficient phagocytosis by macrophages in vitro, since removal of heatinactivated FCS strongly reduced RBC uptake. Phagocytosis of stored human
RBCs could also be restored by heat-inactivated human AB serum. Thus,
heat-stable serum factors seem to be required for macrophage phagocytosis
of stored RBCs. The fact that serum was required for phagocytosis of both
human and murine stored RBCs, and that stored RBCs of both species were
similarly recognized by murine macrophages, suggest that macrophages can
use evolutionary conserved species-independent mechanisms for uptake of
stored RBCs. Complement is not a likely candidate, since it is destroyed by
heat-inactivation [150]. However, several serum proteins have been
described to function as bridge molecules in macrophage recognition of
damaged or apoptotic cells [2, 151]. Thus, further investigations will be
40
required to narrow down on the serum component(s) and corresponding
receptor(s) that promote phagocytosis of RBCs affected by storage.
Concluding remarks
The findings in paper 3 of this thesis indicate that the recognition of liquid
stored RBCs is mediated by class A scavenger receptors. SR-A1 is a possible
candidate which is expressed on many different macrophage populations,
including murine peritoneal macrophages [149], which were used in our
experiments. Inhibition of phagocytosis in our in vitro model, both at
receptor and signaling transduction level, were shown to resemble that
found for oxidatively damaged RBCs [120]. Together with our observation
that the oldest RBCs are more prone to be rapidly removed after transfusion,
this indicates that oxidative damage may very well lead to important
phenotypic changes of stored RBCs that ultimately leads to their swift
recognition after transfusion.
The phagocytosis of stored murine or human RBCs was found to be serum
dependent. In addition, while SR-A1 lacks a signaling domain, PI3 kinase,
p38MAPK and ERK1/2 were found to be important for phagocytosis. This
suggests that (an)other receptor(s), possibly requiring serum proteins, could
provide signaling and partner with SR-A1 for phagocytosis of stored RBCs
(Figure 11). In contrast to phagocytosis of opsonized RBCs, CD47 does not
seem to inhibit SR mediated RBC phagocytosis [120]. Our observation that
stored CD47+/- RBCs (expressing half of normal CD47 levels) were cleared at
the same rate as normal (CD47+/+) RBCs, strengthen the hypothesis for the
involvement of SR in uptake of stored RBCs.
Stored
RBC
Serum protein
SR (SR-A1?)
Receptor x
Macrophage
PI3 kinase
P38 MAPK
ERK1/2
Figure 11. Working hypothesis for a two-receptor model mediating
phagocytosis of stored RBCs.
41
Conclusions
This thesis is based on human and murine in vivo and in vitro
studies. From human studies it is concluded:
•
Increased erythrophagocytosis did not provoke a pro-inflammatory
response in healthy men, thus questioning the iron hypothesis that
have been suggested to occur after transfusion.
•
A subset of liquid stored RBCs display a time dependent increase in
eryptotic-like phenotype.
From studies conducted in the murine model system it is
concluded:
•
Eryptotic cells are phagocytized in the splenic marginal zone by
macrophages and DCs in ways resembling uptake of apoptotic cells.
•
Liquid-stored RBCs is phagocytized by macrophages in the marginal
zone and red pulp of the spleen.
•
The oldest fraction of liquid-stored murine RBCs is responsible for
the reduced 24 h survival seen after transfusion. A reduction of
CD47 on the cell surface did not affect the acute clearance of stored
RBCs after transfusion.
•
In vitro phagocytosis of stored human and mouse RBCs is serum
dependent and can be blocked by class A scavenger receptor
inhibition.
42
Acknowledgements
During my time as a PhD student, I have had the privilege to meet and work
with many interesting colleagues to whom I would like to express my
gratitude.
First, to my supervisor, Christer Malm who finally managed to get me
accepted as a PhD student. I am sincerely grateful for your support over the
years and for always having my back and allowing me to go my own way. You
are a warmhearted man and an inspiring entrepreneur. And thank you for
trying to teach me “good enough”. It has been helpful, even though I still
struggle with accepting this concept.
To my co-supervisor, Per-Arne Oldenborg for taking me in and letting me
try my wings as an experimental researcher. I am grateful for the many
discussions that we have had and for all the knowledge you have shared with
me. You have always been patient and supportive, and allowed me to try to
find the right way, even if it could take some time.
To my former co-supervisor, Lars Frängsmyr, who introduced me to the
field of proteomics where I started this scientific journey. Thank you for the
technical support over the years and for all the scientific and philosophical
discussions.
To my co-author Fredrik Toss at the unit for transfusion medicine,
Norrlands universitetssjukhus who have be extremely helpful and aided me
in getting a clinical insight regarding my research as well as helping me
getting material to do research on. I would also like to acknowledge Lilian
Falk and staff at Blodcentralen in Boden, Staffan Wikstrand at Sunderby
sjukhus, Hans Gulliksson, Stella Larsson and Linda Larsson and staff
at Blodkomponenten, Karolinska universitetssjukhuset and Karin
Skoglund and staff at Blodcentralen in Umeå, for all your help in acquiring
and administrating blood samples and blood bags from different corners of
Sweden.
To the staff of Winternet in Boden, especially Ann-Sofie Lindberg and
Anna Nordin for conducting the tests for aerobic capacity so skillfully. And
to Eva Edholm for taking all the blood samples.
To all my colleagues at the unit for sports medicine for making my years at
the unit so memorable. A special thanks to my fellow PhD-student colleagues
and friends Jonas Johansson and Tommy Henriksson, it would not
have been the same without you! And of course to my friend Apostolos
Theos for all your encouragement. And to my roommates, André
Brännström and Andreas Isaksson for all your artistic pictures and
collages that you have made… And my former roommate, Niklas Boman, I
43
really miss our philosophical discussions. And of course to Gunilla
Solander who have been amazingly helpful and kind during my years at
IME, you are truly missed. And to Britt-Marie Eliasson for your caring
way and administrative help during the more intense parts of my teaching
assignments. To Fia Lindberg for your support and inspiration, you are a
true friend. To Jenny Lindehall Hadrévi, who helped me in getting my
masters project, which ultimately led to me getting a PhD position. To
Michael B. Svensson for being such an inspiring person and to Kajsa
Gilenstam for your thoughtfulness and optimism. To Robert R.N 1,0
Nilsson for your extremely convincing argumentation skills. Or, come to
think of it, your way of never leaving anything up for discussion. My
appreciation also go to Håkan Alfredsson, Lars-Göran Fjällborg, Tom
Pietilä, Monica Sörman, Jonas Lorentzon, Lasse Stigell, Lisbeth
Wikström Frissén, Jiguo Yu, Claes-Göran Sundell, Ann Sörlin,
Anna Nordström, Peter Nordström, Helena Gustavsson, Nils
Berginström, Gabriel Högström, Stefan Sandlund, the staff of the
HAI-project; David, Jim and Magnus, and the lab personnel; Lennart,
Roger and Knappen for making IME such an great place to work at. And
of course to my former coworkers Per Jonsson, CJ Boraxbekk (formally
known as Olsson), Peter Forsgren, Erkki Jakobsson, Kicki Olsson
and Anna Oscarsson as you guys most definitely contributed to the
greatness of IME.
I would also like to thank my friends and colleagues at IMB. In anatomy: To
Anna-Karin Olofsson, I owe you so much for all the help and support
over the years. To Ludvig Backman for letting me run a few gels once in a
while but more important, for valuable discussions of non-scientific
importance. To Gustav Andersson for your absolutely magnificent
artwork on the cover of this thesis. To Mona Lindström, for being a good
listener and occasionally talking some sense in to me. To Anton Tjust for
stimulating academic discussions. To Lars-Eric Thornell for lending me a
pair of shorts so that we were allowed on the golf court in Georgia, USA
(together with the alligators). And to Lena Carlsson, Per Stål, Björn
Lindell, Sture Forsgren, Partik Danielsson, Jingxia Liu, Fatima
Pedrosa Domellöf, Yafeng Song, Eva Carlsson and Johan Bagge for
being great colleagues. In histology: Shrikant Kolan, for always being so
kind and helpful. I wish you the best of luck on your upcoming thesis
defense. To Barbro Borgström for excellent technical assistance. To
Anders Larsson, for sharing your methodological skills in the lab. And to
Åsa Stenberg, Sara af Bjerkén, Ana Virel, Maria Chermenina,
Sanaz Hashemian, Nina Nevalainen, Janne Ylärinne, Per
Linström, Anna Rehnmark, Ingrid Strömberg, Göran Dahlgren,
44
Saowaluck Sompet, Jan-Ove Sehlin, Anita Dreyer-Perkiömäki,
Ann-Lena Tallander and Selamawit Kefela.
To Kristina Lejon, Radha Thyagarajan, and Viqar Banday at the unit
for immunology for interesting lab meetings and journal clubs.
To Elisabeth, Jenny and Sandra at the animal facility for taking good
care of the mice. And to Siggan and Sofia for great company when working
with the mice.
To Börje Ljungberg and Sofia Wård at the dept. of surgical and
perioperative sciences.
Finally, I would like to acknowledge a few persons from outside the
academia that are of special importance to me.
To my fantastic sister Elin, and your wonderful family. You are like a second
family for me. And to my niece and goddaughter Wilma. If you still want to
be a scientist when you grow up, I will help you in any way I can.
To my mother and father, thank you for everything you have given me.
For all the encouragement and support over the years. I know that you have
always believed in me and for that I am truly grateful. This is one step closer
to the blue hall.
To my dear friend Erik. Thank you for all the adventures and for your
support when I have needed it. We have literally been through high and low
together. To Marie, for being such a good friend and listener. And to the
both of you for being an important part of my children’s lives. To Johanna,
finally, the three years have passed. I wish you all the best in the future.
Thank you for giving me the best things in my life, namely;
Simon och Elvira. Jag är så stolt och tacksam över att ni finns i mitt liv!
45
References
1.
Junqueira, L.C.U.a., J. Carneiro, and R.O. Kelley, Basic histology.
7th ed. 1992, Norwalk, Conn.: Appleton & Lange. x, 518 p.
2.
de Back, D.Z., et al., Of macrophages and red blood cells; a complex
love story. Front Physiol, 2014. 5: p. 9.
3.
Cyster, J.G., B cells on the front line. Nat Immunol, 2000. 1(1): p. 910.
4.
Feldschuh, J. and Y. Enson, Prediction of the normal blood volume.
Relation of blood volume to body habitus. Circulation, 1977. 56(4 Pt
1): p. 605-12.
5.
Alberts, B., Molecular biology of the cell. 4th ed. 2002, New York:
Garland Science. xxxiv, 1548 p.
6.
Murphy, K., et al., Janeway's immunobiology. 7th ed. 2008, New
York: Garland Science. xxi, 887 p.
7.
Medzhitov, R. and C.A. Janeway, Jr., Decoding the patterns of self
and nonself by the innate immune system. Science, 2002.
296(5566): p. 298-300.
8.
Gordon, S. and P.R. Taylor, Monocyte and macrophage
heterogeneity. Nat Rev Immunol, 2005. 5(12): p. 953-64.
9.
Wynn, T.A., A. Chawla, and J.W. Pollard, Macrophage biology in
development, homeostasis and disease. Nature, 2013. 496(7446): p.
445-55.
10.
Dunkelberger, J.R. and W.C. Song, Complement and its role in
innate and adaptive immune responses. Cell Res, 2010. 20(1): p.
34-50.
11.
Takeuchi, O. and S. Akira, Pattern recognition receptors and
inflammation. Cell, 2010. 140(6): p. 805-20.
12.
Areschoug, T., J. Waldemarsson, and S. Gordon, Evasion of
macrophage scavenger receptor A-mediated recognition by
pathogenic streptococci. Eur J Immunol, 2008. 38(11): p. 3068-79.
46
13.
Canton, J., D. Neculai, and S. Grinstein, Scavenger receptors in
homeostasis and immunity. Nat Rev Immunol, 2013. 13(9): p. 62134.
14.
Peiser, L. and S. Gordon, The function of scavenger receptors
expressed by macrophages and their role in the regulation of
inflammation. Microbes Infect, 2001. 3(2): p. 149-59.
15.
Reis e Sousa, C., Dendritic cells in a mature age. Nat Rev Immunol,
2006. 6(6): p. 476-83.
16.
Rutella, S., S. Danese, and G. Leone, Tolerogenic dendritic cells:
cytokine modulation comes of age. Blood, 2006. 108(5): p. 1435-40.
17.
Epstein, R.M. and E.M. Papper, Respiratory factors in the uptake
and excretion of anesthetics. 1965. Int Anesthesiol Clin, 1998. 36(4):
p. 121-35.
18.
MacDonald, R., Red cell 2,3-diphosphoglycerate and oxygen
affinity. Anaesthesia, 1977. 32(6): p. 544-53.
19.
Dubinsky, W.P. and E. Racker, The mechanism of lactate transport
in human erythrocytes. J Membr Biol, 1978. 44(1): p. 25-36.
20.
Williams, W.J., Hematology. 4th ed. 1990, New York: McGraw-Hill.
xxxvii, 1882 p., 6 p. of plates.
21.
Smith, B.R., Regulation of hematopoiesis. Yale J Biol Med, 1990.
63(5): p. 371-80.
22.
Mulcahy, L., The erythropoietin receptor. Semin Oncol, 2001. 28(2
Suppl 8): p. 19-23.
23.
Hattangadi, S.M., et al., From stem cell to red cell: regulation of
erythropoiesis at multiple levels by multiple proteins, RNAs, and
chromatin modifications. Blood, 2011. 118(24): p. 6258-68.
24.
Bosman, G.J., F.L. Willekens, and J.M. Werre, Erythrocyte aging: a
more than superficial resemblance to apoptosis? Cell Physiol
Biochem, 2005. 16(1-3): p. 1-8.
25.
Ganz, T. and E. Nemeth, Iron metabolism: interactions with normal
and disordered erythropoiesis. Cold Spring Harb Perspect Med,
2012. 2(5): p. a011668.
47
26.
Pimstone, N.R., et al., The enzymatic degradation of hemoglobin to
bile pigments by macrophages. J Exp Med, 1971. 133(6): p. 1264-81.
27.
Lutz, H.U. and A. Bogdanova, Mechanisms tagging senescent red
blood cells for clearance in healthy humans. Front Physiol, 2013. 4:
p. 387.
28.
Low, P.S., et al., The role of hemoglobin denaturation and band 3
clustering in red blood cell aging. Science, 1985. 227(4686): p. 5313.
29.
Schwarz-Ben Meir, N., T. Glaser, and N.S. Kosower, Band 3 protein
degradation by calpain is enhanced in erythrocytes of old people.
Biochem J, 1991. 275 ( Pt 1): p. 47-52.
30.
Franco, R.S., et al., Changes in the properties of normal human red
blood cells during in vivo aging. Am J Hematol, 2013. 88(1): p. 4451.
31.
Werre, J.M., et al., The red cell revisited--matters of life and death.
Cell Mol Biol (Noisy-le-grand), 2004. 50(2): p. 139-45.
32.
Willekens, F.L., et al., Hemoglobin loss from erythrocytes in vivo
results from spleen-facilitated vesiculation. Blood, 2003. 101(2): p.
747-51.
33.
Willekens, F.L., et al., Liver Kupffer cells rapidly remove red blood
cell-derived vesicles from the circulation by scavenger receptors.
Blood, 2005. 105(5): p. 2141-5.
34.
Vaysse, J., et al., Adhesion and erythrophagocytosis of human
senescent erythrocytes by autologous monocytes and their
inhibition by beta-galactosyl derivatives. Proc Natl Acad Sci U S A,
1986. 83(5): p. 1339-43.
35.
Bratosin, D., et al., Cellular and molecular mechanisms of senescent
erythrocyte phagocytosis by macrophages. A review. Biochimie,
1998. 80(2): p. 173-95.
36.
Khandelwal, S., N. van Rooijen, and R.K. Saxena, Reduced
expression of CD47 during murine red blood cell (RBC) senescence
and its role in RBC clearance from the circulation. Transfusion,
2007. 47(9): p. 1725-32.
48
37.
Cohen, R.M., et al., Red cell life span heterogeneity in
hematologically normal people is sufficient to alter HbA1c. Blood,
2008. 112(10): p. 4284-91.
38.
Vlassara, H., et al., Advanced glycosylation endproducts on
erythrocyte cell surface induce receptor-mediated phagocytosis by
macrophages. A model for turnover of aging cells. J Exp Med, 1987.
166(2): p. 539-49.
39.
Seaman, C., S. Wyss, and S. Piomelli, The decline in energetic
metabolism with aging of the erythrocyte and its relationship to
cell death. Am J Hematol, 1980. 8(1): p. 31-42.
40.
Lang, F., et al., Eryptosis, a window to systemic disease. Cell
Physiol Biochem, 2008. 22(5-6): p. 373-80.
41.
Bratosin, D., et al., Programmed cell death in mature erythrocytes:
a model for investigating death effector pathways operating in the
absence of mitochondria. Cell Death Differ, 2001. 8(12): p. 1143-56.
42.
Lang, F., E. Lang, and M. Foller, Physiology and pathophysiology of
eryptosis. Transfus Med Hemother, 2012. 39(5): p. 308-14.
43.
Maellaro, E., et al., Erythrocyte caspase-3 activation and oxidative
imbalance in erythrocytes and in plasma of type 2 diabetic
patients. Acta Diabetol, 2013. 50(4): p. 489-95.
44.
Calderon-Salinas, J.V., et al., Eryptosis and oxidative damage in
type 2 diabetic mellitus patients with chronic kidney disease. Mol
Cell Biochem, 2011. 357(1-2): p. 171-9.
45.
Basu, S., et al., Eryptosis in hereditary spherocytosis and
thalassemia: role of glycoconjugates. Glycoconj J, 2010. 27(7-9): p.
717-22.
46.
Mahmud, H., et al., Suicidal erythrocyte death, eryptosis, as a novel
mechanism in heart failure-associated anaemia. Cardiovasc Res,
2013. 98(1): p. 37-46.
47.
Kay, M.M., Mechanism of removal of senescent cells by human
macrophages in situ. Proc Natl Acad Sci U S A, 1975. 72(9): p. 35215.
48.
Kiefer, C.R. and L.M. Snyder, Oxidation and
senescence. Curr Opin Hematol, 2000. 7(2): p. 113-6.
49
erythrocyte
49.
Lutz, H.U., Naturally occurring anti-band 3 antibodies in clearance
of senescent and oxidatively stressed human red blood cells.
Transfus Med Hemother, 2012. 39(5): p. 321-7.
50.
Turrini, F., et al., Clustering of integral membrane proteins of the
human erythrocyte membrane stimulates autologous IgG binding,
complement deposition, and phagocytosis. J Biol Chem, 1991.
266(35): p. 23611-7.
51.
Boas, F.E., L. Forman, and E. Beutler, Phosphatidylserine exposure
and red cell viability in red cell aging and in hemolytic anemia.
Proc Natl Acad Sci U S A, 1998. 95(6): p. 3077-81.
52.
Oldenborg, P.A., et al., Role of CD47 as a marker of self on red blood
cells. Science, 2000. 288(5473): p. 2051-4.
53.
Paulson, J.C., M.S. Macauley, and N. Kawasaki, Siglecs as sensors of
self in innate and adaptive immune responses. Ann N Y Acad Sci,
2012. 1253: p. 37-48.
54.
Durocher, J.R., R.C. Payne, and M.E. Conrad, Role of sialic acid in
erythrocyte survival. Blood, 1975. 45(1): p. 11-20.
55.
Kraal, G., Cells in the marginal zone of the spleen. Int Rev Cytol,
1992. 132: p. 31-74.
56.
Mebius, R.E. and G. Kraal, Structure and function of the spleen. Nat
Rev Immunol, 2005. 5(8): p. 606-16.
57.
Blundell, J., Experiments on the Transfusion of Blood by the
Syringe. Med Chir Trans, 1818. 9(Pt 1): p. 56-92.
58.
Welck, M., P. Borg, and H. Ellis, James Blundell MD Edin FRCP
(1790-1877): pioneer of blood transfusion. J Med Biogr, 2010. 18(4):
p. 194-7.
59.
Landsteiner, K., On agglutination of normal human blood.
Transfusion, 1961. 1: p. 5-8.
60.
Levine, P. and R.E. Stetson, Landmark article July 8, 1939. An
unusual case of intra-group agglutination. By Philip Levine and
Rufus E Stetson. JAMA, 1984. 251(10): p. 1316-7.
61.
Matteocci, A. and L. Pierelli, Red blood cell alloimmunization in
sickle cell disease and in thalassaemia: current status, future
50
perspectives and potential role of molecular typing. Vox Sang,
2014. 106(3): p. 197-208.
62.
Mitra, R., N. Mishra, and G.P. Rath, Blood groups systems. Indian J
Anaesth, 2014. 58(5): p. 524-8.
63.
Goldstein, J., et al., Group B erythrocytes enzymatically converted
to group O survive normally in A, B, and O individuals. Science,
1982. 215(4529): p. 168-70.
64.
Liu, Q.P., et al., Bacterial glycosidases for the production of
universal red blood cells. Nat Biotechnol, 2007. 25(4): p. 454-64.
65.
Lewisohn, R., Twenty Years' Experience with the Citrate Method of
Blood Transfusion. Ann Surg, 1937. 105(4): p. 602-9.
66.
Mollison, P.L., The introduction of citrate as an anticoagulant for
transfusion and of glucose as a red cell preservative. Br J
Haematol, 2000. 108(1): p. 13-8.
67.
Gibson, J.G., 2nd, C.B. Gregory, and L.N. Button, Citratephosphate-dextrose solution for preservation of human blood: a
further report. Transfusion, 1961. 1: p. 280-7.
68.
WHO, Blood safety and availability. World health organization,
2014. Fact sheet No279.
69.
Hess, J.R., Red cell storage. J Proteomics, 2010. 73(3): p. 368-73.
70.
Greenwalt, T.J. and K.R. Zelenski, Transfusion support for
haemoglobinopathies. Clin Haematol, 1984. 13(1): p. 151-65.
71.
Goodnough, L.T., J.H. Levy, and M.F. Murphy, Concepts of blood
transfusion in adults. Lancet, 2013. 381(9880): p. 1845-54.
72.
Tsai, A.G., et al., Perfusion vs. oxygen delivery in transfusion with
"fresh" and "old" red blood cells: the experimental evidence.
Transfus Apher Sci, 2010. 43(1): p. 69-78.
73.
Pace, N., W.V. Consolazio, and E.L. Lozner, The Effect of
Transfusions of Red Blood Cells on the Hypoxia Tolerance of
Normal Men. Science, 1945. 102(2658): p. 589-91.
51
74.
Cacic, D.L., T. Hervig, and J. Seghatchian, Blood doping: the flip
side of transfusion and transfusion alternatives. Transfus Apher
Sci, 2013. 49(1): p. 90-4.
75.
Hess, J.R., An update on solutions for red cell storage. Vox Sang,
2006. 91(1): p. 13-9.
76.
Luten, M., et al., Survival of red blood cells after transfusion: a
comparison between red cells concentrates of different storage
periods. Transfusion, 2008. 48(7): p. 1478-85.
77.
Zeiler, T., Müller, J.T., Kretschmer, V., Flow-cytometric
determination of survival time and 24-hour recovery of transfused
red blood cells. Transfus Med Hemother, 2003. 30(1): p. 14-19.
78.
Cancelas, J.A., et al., Additive solution-7 reduces the red blood cell
cold storage lesion. Transfusion, 2015. 55(3): p. 491-8.
79.
Almizraq, R., et al., Storage of red blood cells affects membrane
composition, microvesiculation, and in vitro quality. Transfusion,
2013. 53(10): p. 2258-67.
80.
D'Alessandro, A., et al., Time-course investigation of SAGM-stored
leukocyte-filtered red bood cell concentrates: from metabolism to
proteomics. Haematologica, 2012. 97(1): p. 107-15.
81.
Kriebardis, A.G., et al., Storage-dependent remodeling of the red
blood cell membrane is associated with increased immunoglobulin
G binding, lipid raft rearrangement, and caspase activation.
Transfusion, 2007. 47(7): p. 1212-20.
82.
Relevy, H., et al., Blood banking-induced alteration of red blood cell
flow properties. Transfusion, 2008. 48(1): p. 136-46.
83.
Anniss, A.M. and R.L. Sparrow, Expression of CD47 (integrinassociated protein) decreases on red blood cells during storage.
Transfus Apher Sci, 2002. 27(3): p. 233-8.
84.
Stewart, A., et al., The application of a new quantitative assay for
the monitoring of integrin-associated protein CD47 on red blood
cells during storage and comparison with the expression of CD47
and phosphatidylserine with flow cytometry. Transfusion, 2005.
45(9): p. 1496-503.
52
85.
Heaton, A., T. Keegan, and S. Holme, In vivo regeneration of red
cell 2,3-diphosphoglycerate following transfusion of DPG-depleted
AS-1, AS-3 and CPDA-1 red cells. Br J Haematol, 1989. 71(1): p. 1316.
86.
Klein, H.G., D.R. Spahn, and J.L. Carson, Red blood cell transfusion
in clinical practice. Lancet, 2007. 370(9585): p. 415-26.
87.
Henkelman, S., et al., The effects of cryopreservation on red blood
cell rheologic properties. Transfusion, 2010. 50(11): p. 2393-401.
88.
Bandarenko, N., et al., Successful in vivo recovery and extended
storage of additive solution (AS)-5 red blood cells after
deglycerolization and resuspension in AS-3 for 15 days with an
automated closed system. Transfusion, 2007. 47(4): p. 680-6.
89.
Holovati, J.L., et al., The effects of cryopreservation on red blood
cell microvesiculation, phosphatidylserine externalization, and
CD47 expression. Transfusion, 2008. 48(8): p. 1658-68.
90.
Marik, P.E. and H.L. Corwin, Efficacy of red blood cell transfusion
in the critically ill: a systematic review of the literature. Crit Care
Med, 2008. 36(9): p. 2667-74.
91.
Carson, J.L., R.C. Reynolds, and H.G. Klein, Bad bad blood? Crit
Care Med, 2008. 36(9): p. 2707-8.
92.
Flegel, W.A., C. Natanson, and H.G. Klein, Does prolonged storage
of red blood cells cause harm? Br J Haematol, 2014. 165(1): p. 3-16.
93.
Wang, D., et al., Transfusion of older stored blood and risk of death:
a meta-analysis. Transfusion, 2012. 52(6): p. 1184-95.
94.
Sparrow, R.L., Red Blood Cell Storage Duration and Trauma.
Transfus Med Rev, 2015. 29(2): p. 120-126.
95.
Aubron, C., et al., Age of red blood cells and transfusion in critically
ill patients. Ann Intensive Care, 2013. 3(1): p. 2.
96.
Lacroix, J., et al., Age of Transfused Blood in Critically Ill Adults. N
Engl J Med, 2015. 372(15): p. 1410-1418.
97.
Steiner, M.E., et al., Effects of red-cell storage duration on patients
undergoing cardiac surgery. N Engl J Med, 2015. 372(15): p. 141929.
53
98.
Fergusson, D.A., et al., Effect of fresh red blood cell transfusions on
clinical outcomes in premature, very low-birth-weight infants: the
ARIPI randomized trial. JAMA, 2012. 308(14): p. 1443-51.
99.
Zimring, J.C., Established and theoretical factors to consider in
assessing the red cell storage lesion. Blood, 2015. 125(14): p. 218590.
100.
Pereira, A., Will clinical studies elucidate the connection between
the length of storage of transfused red blood cells and clinical
outcomes? An analysis based on the simulation of randomized
controlled trials. Transfusion, 2013. 53(1): p. 34-40.
101.
Hod, E.A., et al., Transfusion of red blood cells after prolonged
storage produces harmful effects that are mediated by iron and
inflammation. Blood, 2010. 115(21): p. 4284-92.
102.
Callan, M.B., et al., Transfusion of 28-day-old leucoreduced or nonleucoreduced stored red blood cells induces an inflammatory
response in healthy dogs. Vox Sang, 2013. 105(4): p. 319-27.
103.
Spitalnik, S.L., Stored red blood cell transfusions: iron,
inflammation, immunity, and infection. Transfusion, 2014. 54(10):
p. 2365-71.
104.
Hod, E.A., et al., Transfusion of human volunteers with older,
stored red blood cells produces extravascular hemolysis and
circulating non-transferrin-bound iron. Blood, 2011. 18(25): p.
6675-82.
105.
Mollison, P.L., Methods of determining the posttransfusion survival
of stored red cells. Transfusion, 1984. 24(2): p. 93-6.
106.
Mock, D.M., et al., Measurement of posttransfusion red cell survival
with the biotin label. Transfus Med Rev, 2014. 28(3): p. 114-25.
107.
Kiraly, L.N., et al., Transfusion of aged packed red blood cells
results in decreased tissue oxygenation in critically injured trauma
patients. J Trauma, 2009. 67(1): p. 29-32.
108.
Marik, P.E. and W.J. Sibbald, Effect of stored-blood transfusion on
oxygen delivery in patients with sepsis. JAMA, 1993. 269(23): p.
3024-9.
54
109.
Roback, J.D., Vascular effects of the red blood cell storage lesion.
Hematology Am Soc Hematol Educ Program, 2011. 2011: p. 475-9.
110.
Doctor, A. and P. Spinella, Effect of processing and storage on red
blood cell function in vivo. Semin Perinatol, 2012. 36(4): p. 248-59.
111.
Taft, R.A., M. Davisson, and M.V. Wiles, Know thy mouse. Trends
Genet, 2006. 22(12): p. 649-53.
112.
Mestas, J. and C.C. Hughes, Of mice and not men: differences
between mouse and human immunology. J Immunol, 2004. 172(5):
p. 2731-8.
113.
Horky, J., J. Vacha, and V. Znojil, Comparison of life span of
erythrocytes in some inbred strains of mouse using 14C-labelled
glycine. Physiol Bohemoslov, 1978. 27(3): p. 209-17.
114.
Knutson, M. and M. Wessling-Resnick, Iron metabolism in the
reticuloendothelial system. Crit Rev Biochem Mol Biol, 2003. 38(1):
p. 61-88.
115.
Waugh, R.E., et al., Rheologic properties of senescent erythrocytes:
loss of surface area and volume with red blood cell age. Blood,
1992. 79(5): p. 1351-8.
116.
Makley, A.T., et al., Murine blood banking: characterization and
comparisons to human blood. Shock, 2010. 34(1): p. 40-5.
117.
Vernet, M., Immunochemical assay of transferrin and iron
saturation in serum. Clin Chem, 1993. 39(11 Pt 1): p. 2352-3.
118.
Parish, C.R., Fluorescent dyes for lymphocyte migration and
proliferation studies. Immunol Cell Biol, 1999. 77(6): p. 499-508.
119.
Saxena, R.K., et al., A Double in vivo Biotinylation Technique for
Objective Assessment of Aging and Clearance of Mouse
Erythrocytes in Blood Circulation. Transfus Med Hemother, 2012.
39(5): p. 335-41.
120.
Olsson, M. and P.A. Oldenborg, CD47 on experimentally senescent
murine RBCs inhibits phagocytosis following Fcgamma receptormediated but not scavenger receptor-mediated recognition by
macrophages. Blood, 2008. 112(10): p. 4259-67.
55
121.
Goodnough, L.T. and M.F. Murphy, Do liberal blood transfusions
cause more harm than good? BMJ, 2014. 349: p. g6897.
122.
Hill, H.R., et al., The effects of polyvinyl chloride and polyolefin
blood bags on red blood cells stored in a new additive solution. Vox
Sang, 2001. 81(3): p. 161-6.
123.
Dumont, L.J., et al., Exploratory in vitro study of red blood cell
storage containers formulated with an alternative plasticizer.
Transfusion, 2012. 52(7): p. 1439-45.
124.
Haishima, Y., et al., Screening study on hemolysis suppression effect
of an alternative plasticizer for the development of a novel blood
container made of polyvinyl chloride. J Biomed Mater Res B Appl
Biomater, 2014. 102(4): p. 721-8.
125.
Wagner, C.T., et al., Biochemical stabilization enhances red blood
cell recovery and stability following cryopreservation. Cryobiology,
2002. 45(2): p. 153-66.
126.
Chaplin, H., Jr., The proper use of previously frozen red blood cells
for transfusion. Blood, 1982. 59(6): p. 1118-20.
127.
Kuwahira, I., et al., Splenic contraction-induced reversible increase
in hemoglobin concentration in intermittent hypoxia. J Appl
Physiol (1985), 1999. 86(1): p. 181-7.
128.
Sallet, P., et al., Study of a new indirect method based on absolute
norms of variation to detect autologous blood transfusion. Int J
Hematol, 2008. 88(4): p. 362-8.
129.
Keir, A.K., et al., Plasma cytokines and markers of endothelial
activation increase after packed red blood cell transfusion in the
preterm infant. Pediatr Res, 2013. 73(1): p. 75-9.
130.
Dumont, L.J. and J.P. AuBuchon, Evaluation of proposed FDA
criteria for the evaluation of radiolabeled red cell recovery trials.
Transfusion, 2008. 48(6): p. 1053-60.
131.
Beutler, E., Destruction of erythrocytes, in Williams Hematology,
8:th ed., K. Kaushasky, Lichtman, M.A., Beutler, E., Kipps, T.J.,
Seligsohn, U., Prchal, J.T., Editor. 2010, Mc Graw Hill: New York. p.
451-2.
56
132.
Sowemimo-Coker, S.O., Red blood cell hemolysis during processing.
Transfus Med Rev, 2002. 16(1): p. 46-60.
133.
Jayle, M.F. and J. Moretti, Haptoglobin: Biochmical, genetic and
physiopathologic aspects. Progr Hematol, 1962. 3: p. 342-59.
134.
Gilson, C.R., et al., A novel mouse model of red blood cell storage
and posttransfusion in vivo survival. Transfusion, 2009. 49(8): p.
1546-53.
135.
Burger, P., et al., Potassium leakage primes stored erythrocytes for
phosphatidylserine exposure and shedding of pro-coagulant
vesicles. Br J Haematol, 2013. 160(3): p. 377-86.
136.
van Rooijen, N. and E. Hendrikx, Liposomes for specific depletion of
macrophages from organs and tissues. Methods Mol Biol, 2010.
605: p. 189-203.
137.
Wojczyk, B.S., et al., Macrophages clear refrigerator storagedamaged red blood cells and subsequently secrete cytokines in vivo,
but not in vitro, in a murine model. Transfusion, 2014. 54(12): p.
3186-97.
138.
Belz, G.T., W.R. Heath, and F.R. Carbone, The role of dendritic cell
subsets in selection between tolerance and immunity. Immunol Cell
Biol, 2002. 80(5): p. 463-8.
139.
Hendrickson, J.E., et al., Recipient inflammation affects the
frequency and magnitude of immunization to transfused red blood
cells. Transfusion, 2006. 46(9): p. 1526-36.
140.
Hendrickson, J.E., et al., Discrete Toll-like receptor agonists have
differential effects on alloimmunization to transfused red blood
cells. Transfusion, 2008. 48(9): p. 1869-77.
141.
Qiu, C.H., et al., Novel subset of CD8{alpha}+ dendritic cells
localized in the marginal zone is responsible for tolerance to cellassociated antigens. J Immunol, 2009. 182(7): p. 4127-36.
142.
McGaha, T.L., et al., Marginal zone macrophages suppress innate
and adaptive immunity to apoptotic cells in the spleen. Blood, 2011.
117(20): p. 5403-12.
143.
Olsson, M., A. Nilsson, and P.A. Oldenborg, Dose-dependent
inhibitory effect of CD47 in macrophage uptake of IgG-opsonized
57
murine erythrocytes. Biochem Biophys Res Commun, 2007. 352(1):
p. 193-7.
144.
Sosale, N.G., et al., Cell rigidity and shape override CD47's "self"signaling in phagocytosis by hyperactivating myosin-II. Blood,
2015. 125(3): p. 542-52.
145.
Burger, P., et al., CD47 functions as a molecular switch for
erythrocyte phagocytosis. Blood, 2012. 119(23): p. 5512-21.
146.
Platt, N. and S. Gordon, Is the class A macrophage scavenger
receptor (SR-A) multifunctional? - The mouse's tale. J Clin Invest,
2001. 108(5): p. 649-54.
147.
Gillooly, D.J., A. Simonsen, and H. Stenmark, Phosphoinositides
and phagocytosis. J Cell Biol, 2001. 155(1): p. 15-7.
148.
Kelley, J.L., et al., Scavenger receptor-A (CD204): a two-edged
sword in health and disease. Crit Rev Immunol, 2014. 34(3): p. 24161.
149.
Yu, H., et al., Scavenger receptor A (SR-A) is required for LPSinduced TLR4 mediated NF-kappaB activation in macrophages.
Biochim Biophys Acta, 2012. 1823(7): p. 1192-8.
150.
Soltis, R.D., et al., The effect of heat inactivation of serum on
aggregation of immunoglobulins. Immunology, 1979. 36(1): p. 3745.
151.
Erwig, L.P. and P.M. Henson, Clearance of apoptotic cells by
phagocytes. Cell Death Differ, 2008. 15(2): p. 243-50.
58
1