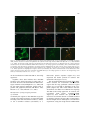
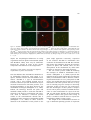
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Springer 2005 Plant Molecular Biology (2005) 58:109–122 DOI 10.1007/s11103-005-4618-4 Identification and characterization of AtCASP, a plant transmembrane Golgi matrix protein Luciana Renna1, , Sally L. Hanton1, , Giovanni Stefano1, , Lauren Bortolotti1, Vikram Misra2 and Federica Brandizzi1,* 1 Department of Biology, 112 Science Place, University of Saskatchewan, Saskatoon, SK S7N 5E2, Canada; (*author for correspondence; e-mail [email protected]); 2Department of Veterinary Sciences, 52 Campus Drive, University of Saskatchewan, Saskatoon, SK S7N 545B, Canada; These authors have contributed equally to the work Received 14 February 2005; accepted in revised form 28 March 2005 Key words: CASP, membrane Golgi matrix proteins, plants Abstract Golgins are a family of coiled-coil proteins that are associated with the Golgi apparatus. They are necessary for tethering events in membrane fusion and may act as structural support for Golgi cisternae. Here we report on the identification of an Arabidopsis golgin which is a homologue of CASP, a known transmembrane mammalian and yeast golgin. Similar to its homologues, the plant CASP contains a long N-terminal coiled-coil region protruding into the cytosol and a C-terminal transmembrane domain with amino acid residues which are highly conserved across species. Through fluorescent protein tagging experiments, we show that plant CASP localizes at the plant Golgi apparatus and that the C-terminus of this protein is sufficient for its localization, as has been shown for its mammalian counterpart. In addition, we demonstrate that the plant CASP is able to localize at the mammalian Golgi apparatus. However, mutagenesis of a conserved tyrosine in the transmembrane domain revealed that it is necessary for ER export and Golgi localization of the Arabidopsis CASP in mammalian cells, but is not required for its correct localization in plant cells. These data suggest that mammalian and plant cells have different mechanisms for concentrating CASP in the Golgi apparatus. Introduction The Golgi apparatus is a vital organelle dedicated to processing and sorting of secretory cargo in all eukaryotic cells (Palade, 1975). In plants, vesicles containing protein cargo are targeted to both the cis- and trans-cisternae from other organelles (reviewed by Jürgens, 2004). It is likely that vesicles also move between cisternae as may occur in other species (Volchuk et al., 2004). How the specificity of vesicular traffic between individual cisternae is conferred has not yet been well-characterized in plants, although a considerable amount of evidence regarding the function and localization of SNAREs (Sanderfoot et al., 2001; Uemura et al., 2004) and Rabs (Drew et al., 1993; Ueda et al., 1996; Batoko et al., 2000; Inaba et al., 2002; Saint-Jore et al., 2002; Grebe et al., 2003) in the Golgi apparatus has been presented. Further investigations based on those already carried out will doubtless clarify the molecular mechanisms involved. Plant cells have numerous Golgi stacks dispersed in the cytoplasm (Boevink et al., 1998; Nebenführ et al. 1999). Each stack contains cis-totrans cisternae and moves on an underlying membrane network, the endoplasmic reticulum (ER), by means of actin-myosin motors (Boevink et al., 1998; Brandizzi et al., 2002a; Saint-Jore 110 et al., 2002). The Golgi apparatus and ER export sites (ERES, daSilva et al., 2004) form secretory units that move together, although the ER to Golgi transport of cargo molecules is independent of the movement (Brandizzi et al., 2002a). It is currently unknown how the plant Golgi apparatus maintains its structure despite the rapid transport of proteins to and from this organelle (Brandizzi et al., 2002a) and while it is moving over the ER (daSilva et al., 2004). It has been proposed that the mammalian Golgi apparatus may maintain its form and ensure vesicular trafficking via a proteinaceous structure known as the matrix or scaffold (Slusarewicz et al., 1994). Matrix proteins are involved in binding to and linking adjacent membranes in vesicle docking during protein transport, and linking cisternae during Golgi stack formation (Sonnichsen et al., 1998; Shorter and Warren, 1999). Long coiled-coil proteins such as golgins are components of this matrix. Golgins were initially identified on the Golgi and on endosomes, mostly as auto-antigens in autoimmune disorders (Dohlman et al., 1993). Based on their restricted intracellular distributions and their predicted rod-like structure, it has been proposed that these proteins play a role in tethering vesicles to target Golgi membranes prior to fusion. However, such proteins may also play a structural role as scaffolds for the assembly of other factors important for fusion, such as Rab proteins (reviewed in Barr and Short, 2003). The large number of different golgins suggests that they may perform more than one general function in the Golgi. Indeed, tethering and organizational functions for a number of golgins have been shown in in vitro systems. For instance, Uso1p and its mammalian homologue p115 are required for ER-to-Golgi transport in vivo, and reconstitution of this process in vitro has shown that the requirement is at the stage of vesicle tethering to the cis-Golgi (Cao et al., 1998; Seemann et al., 2000; Alvarez et al., 2001). Two other coiled-coil proteins, GM130 and giantin, have been shown to recruit p115 to Golgi membranes (Sonnichsen et al., 1998; Lesa et al., 2000; Puthenveedu and Linstedt, 2001). These proteins also seem to contribute to cisternal structure in vivo and are required in an in vitro assay system that reconstitutes the stacking of Golgi cisternae (Shorter and Warren, 1999; Puthenveedu and Linstedt, 2001). Most golgins are peripheral membrane proteins and often have short non-coiled-coil regions at either end of the protein that mediate targeting and other interactions. For example, the C-terminus of GM130 binds to GRASP65, a lipid-anchored protein on the cis-Golgi (Barr et al., 1998). Similarly, several golgins share a C-terminal GRIP domain that is sufficient to target them to the trans-Golgi (Barr, 1999; Kjer-Nielsen et al., 1999; Munro and Nichols, 1999; Barr and Short, 2003). In contrast, three golgins, CASP (CCAAT-displacement protein alternatively spliced product), giantin and golgin-84 have been experimentally proved to be integral membrane proteins anchored to the bilayer via a C-terminal transmembrane domain (Linstedt and Hauri, 1993; Bascom et al., 1999; Misumi et al., 2001; Gillingham et al., 2002). Ultrastructural evidence points towards the existence of a Golgi matrix in plant cells (Staehelin and Moore, 1995; Neumann et al., 2003) and the existence of DNA sequences in the Arabidopsis genome encoding homologues of mammalian matrix proteins has been reported (Gillingham et al., 2002; Rose et al., 2004). However, to date there is little data on the localization and dynamics of these proteins. Only recently, the localization of a green fluorescent protein fusion to the GRIP domain of an Arabidopsis gene (AtGRIP) has been described (Gilson et al., 2004). The C-terminal domain from AtGRIP is sufficient to target the Golgi apparatus in plant and mammalian cells, indicating that GRIP-proteins might be implicated in a targeting mechanism that is conserved amongst eukaryotes (Gilson et al., 2004). However, apart from the identification of genomic sequences, no other data are available on integral membrane Golgi matrix proteins in plants. Here we have studied a coiled-coil integral membrane protein from Arabidopsis thaliana (AtCASP) that shares a high degree of homology with human and yeast CASP, a known golgin in these organisms (Gillingham et al., 2002; Mansour et al., 2002; Barr and Short, 2003). AtCASP shows an overall domain structure that is strongly conserved across kingdoms. By means of fluorescent protein tagging, we have determined that AtCASP targets the Golgi apparatus in plant cells and also contains encoded information to target the Golgi apparatus in mammalian cells. Our results indicate that integral membrane Golgi matrix proteins exist in plant cells and belong to 111 a highly conserved family that shares overall structure and targeting domains with homologues in other kingdoms. Materials and methods Molecular Cloning Standard molecular techniques were used as described in Sambrook et al. (1989). The fluorescent proteins used in this study were based on fusions with either mGFP5 (Haseloff et al., 1997) or EYFP (Clontech Inc., California, USA). The spectral properties of mGFP5 allow efficient spectral separation from YFP (Brandizzi et al., 2002b). We used TMHMM Server v. 2.0 (Denmark) for prediction of transmembrane helices in proteins and Parcoil for prediction of coiled-coil regions (Berger et al., 1995). Similarity and identity values for the comparison of AtCASP with human and yeast proteins were established by BLASTP 2.2.10 analysis using BLOSUM62 matrix. To generate an AtCASP-fluorescent protein fusion, the cDNA of the Arabidopsis CASP (GenBank accession AY142680) was fused in frame downstream of a fluorescent protein sequence using unique BamHI and SacI sites and inserted in pVKH18En6. To generate the tyrosine mutant (residue 648), we performed site directed mutagenesis by altering the relevant codon to leucine. Fluorescent fusions of AtNCASP and AtCCASP were generated by PCR amplification of the DNA encoding for the predicted N-terminus domain of the protein (1–564aa) and the last 125 amino acids, respectively. The PCR products were fused in frame downstream of a fluorescent protein sequence using unique BamHI and SacI sites and inserted in pVKH18En6. All CASP fusions, including mutants, full length plus N- and C-terminal fusions, were generated as GFP and YFP fusions. The localization of each type of fusion was identical regardless of the GFP variant in use. For expression in mammalian cells, the fluorescent fusions of CASP and its mutants were amplified with primers containing XhoI and XbaI sites for subcloning in the expression vector pcDNA3 (Invitrogen). ST-RFP was generated as an in frame fusion of the last 58 amino acids of a rat sialyltrans- ferase fused to the monomeric RFP as explained in Saint-Jore et al. (2002). The DNA encoding for this fusion was subcloned into pcDNA3 using the XhoI and XbaI sites of its multiple cloning site. The plasmid encoding for the ER marker, luman (pcLuman-RFP, Lu and Misra, 2000) was generated by recovering the coding sequences of luman, without the stop codon, as well as the coding sequences of the monomeric red F1 protein by PCR and then assembling the two fragments between the BamHI and XbaI sites of pcDNA3. The primers for amplifying luman incorporated BamHI and EcoRI sites while the PCR product of RFP was bracketed by EcoRI and XbaI sites. The primer sequences used for the subcloning and mutagenesis indicated above are available upon request. Expression Systems Expression in plant cells With the only exception of the GFP-AtCASPY648L mutant, which has been expressed, transiently, for this work we have used plant transformants expressing stably and transiently all the other AtCASP and derivative constructs. We have obtained the same results with both systems. The confocal images produced for this work have been obtained from transient transformants. The cell fractionation has been performed on leaves of stable transformants for AtCCASP. Generation of stable plant transformants was achieved as described in Crofts et al. (1999), except that the Agrobacterium strain used was GV3101 and selection for transformants was on hygromycin. Transformants were screened using fluorescence microscopy. Four week old Nicotiana tabacum (cv Petit Havana) greenhouse plants grown at 25 C were used for Agrobacterium tumefaciens [strain GV3101]-mediated transient expression (Batoko et al., 2000). The bacterial optical density (OD600) used for plant transformation was 0.05–0.1 for CASP- and 0.2 for ERD2- and ST-tagged constructs. Transient expression in mammalian cells African green monkey (Vero, obtained from P. O’Hare, Marie Curie Institute, UK) cells were transfected with plasmid DNA by a modification of the technique described by Chen and Okayama 112 (1998) . Briefly, 2.5 · 105 Vero cells in Dulbecco’s minimum essential medium (Invitrogen) containing 10% newborn calf serum, penicillin and streptomycin were added to each well of a six well tissue culture plate (BD Falcon) containing quartz coverslips (VWR). After incubating overnight the culture medium was replaced and two hours later 5 lg of plasmid in transfection buffer (140 mM NaCl, 25 mM N,N-bis(2hydroxyethyl)-2-aminoethanesulfonic acid, 0.75 mM Na2HPO4 and 125 mM CaCl2) were added to cells in each well (Chen and Okayama, 1988). Sixteen hours later the medium was replaced and the next day cells on the coverslips were rinsed with phosphate buffered saline (PBS) and fixed for 10 min in 1% paraformaldehyde. The coverslips were mounted on glass slides in 50% glycerol in PBS. Cell Fractionation and Protein Gel Blot Analysis For cell fractionation experiments, we isolated protoplasts from leaves of stable transformants expressing AtCCASP or transient transformants expressing ssNGFP-HDEL as described in Phillipson et al. (2001). Protoplasts were then fractionated into cytosol and vacuoles (S1), microsomal contents (S2) and cell membranes (P) (Denecke et al., 1992). Protoplasts were gently resuspended in GFP extraction buffer (0.2 M NaCl, 0.1 M Tris pH 7.8, 1 mM EDTA pH 8, supplemented with 2% v/v b-mercaptoethanol immediately before use), and incubated on ice for 10 min. Samples labeled + Triton were resuspended in GFP extraction buffer supplemented with 0.2% v/v Triton X-100 to solubilize membranes. All samples were centrifuged for 15 min at 14,000 rpm and 4 C. The supernatant (S1) was recovered and the pellet was resuspended by sonication in GFP extraction buffer (with or without 0.2% v/v Triton X-100), then centrifuged for a further 15 min at 14,000 rpm and 4 C. The supernatant (S2) was recovered and the pellet was resuspended by sonication in GFP extraction buffer as before (P). All samples were resuspended to a 20-fold lower volume than that of the original cell suspension. Protein extracts were diluted 50:50 with 2X SDS loading buffer (Crofts et al., 1999) and boiled for 5 min. Equal volumes of all extracts were loaded on SDS-polyacrylamide gels, transferred to nitrocellulose membrane by electroblotting and blocked with PBS, 0.05% Tween 20 and 1% milk powder for 2 h. The filter was then incubated in blocking buffer with anti-GFP serum from rabbit (Molecular Probes) at a dilution of 1:1000 overnight. Further steps were performed as in Crofts et al. (1999). The anti-GFP serum is known to recognize all the GFP variants (Molecular Probes). Proteinase K treatment Microsomes were isolated from transgenic protoplasts by osmotic shock in GFP extraction buffer without b-mercaptoethanol, followed by incubation on ice for 10 min. Microsomes were then incubated on ice with or without 0.3 mg/ml proteinase K for 30 min. After incubation the samples were boiled for 10 min to inactivate the proteinase. Extracts were diluted 50:50 with 2X SDS loading buffer and the samples analysed by western blot. Tunicamycin treatment Transgenic protoplasts were incubated in 20 lg/ml tunicamycin for 24 h before extraction by sonication in GFP extraction buffer supplemented with 0.2% v/v Triton X-100. Extracts were diluted 50:50 with 2X SDS loading buffer and the samples analysed by western blot. Sampling and Imaging Transformed leaves were analysed 48 h after infection of the lower epidermis. Confocal imaging was performed using an upright Zeiss LSM 510 META confocal microscope and a 63 · water immersion objective. For imaging expression of GFP constructs, YFP constructs or both, we used imaging settings as described in Brandizzi et al. (2002b) with a 1–3 lm optical slice. Appropriate controls were done to exclude the possibility of energy transfer between fluorochromes and cross-talk. Time-lapse scanning was acquired with imaging system software of the microscope. Fluorescence intensity measurements (plus tracking) and post acquisition image processing were done with Zeiss confocal and PaintShop Pro 7.0 software, respectively. 113 Results Plants have a homologous protein to the human golgin CASP Four integral membrane golgins have been identified in animal cells: giantin, golgin-84, golgin-67 and CASP (Linstedt and Hauri, 1993; Bascom et al., 1999; Jakymiw et al., 2000; Misumi et al., 2001; Gillingham et al., 2002; Barr and Short, 2003). Their identity as membrane spanning proteins has been ascertained experimentally for giantin, golgin-84 and CASP (Bascom et al., 1999; Misumi et al., 2001; Gillingham et al., 2002; Figure 1A), while golgin-67 has been predicted to have a membrane spanning region at the C-terminus (Jakymiw et al., 2000). Giantin, golgin-84 and CASP share a common topology with a cytosolic Figure 1. The plant CASP has an overall structure similar to homologues from other species. (A) Diagrams of the human (hs) giantin, golgin-84, CASP and Arabidopsis (At) CASP indicating the potential coiled-coils and the TMDs. All these golgins share a common topology with an extensive coiled-coil N-terminal region and a C-terminal region. Coiled-coil regions and transmembrane domains were established by Paircoil and TMHMM software, respectively. As predicted by the Paircoil program, the extensive coiled-coil region occupies 65% of the entire protein. (B) Diagram of entire AtCASP (residues 1–689). The large white box indicates the coiled-coil regions which are differentiated graphically from the non-coiled-coil regions by a black line. The black box indicates the transmembrane domain of the protein (TMD). An arrowhead points at residue 564 which delimits the AtNCASP (residues 1–564) and AtCCASP (residues 565–689). At the bottom of the panel there is a colour coded alignment of the amino acid composition of the transmembrane domain of CASP from different species. Note the presence of the conserved tyrosine residue (648aa; arrowhead). (C) Diagrams of the AtCASP fluorescent constructs used in this study indicating the position of a fluorescent protein (FP, green rectangle) with respect to the amino acid residues of AtCASP. For NGFP-AtCASP, a blue rectangle indicates the peptide bearing a glycosylation motif. 114 N-terminus and a membrane spanning domain at the C-terminus (Figure 1A; see also Gillingham et al., 2002). These proteins also contain several residues in their transmembrane domain that are strongly conserved between different species (Gillingham et al., 2002). In a protein-protein Blast search (NCBI Blast) in the Arabidopsis genome using human and yeast CASP homologues, we have identified an Arabidopsis protein that is a homologue of the CASP of these species (AtCASP; Figure 1; see also Gillingham et al., 2002). The putative Arabidopsis protein shows 32% identity, 55% similarity with the human homologue and 26% identity, 45% similarity with the yeast CASP. AtCASP is a protein of 689 residues and a predicted molecular mass of 80 kDa. The predicted structure of the Arabidopsis protein indicates the presence of short non-coiled-coil regions at either end of the protein and coiled-coil domains over most of its length (Figure 1A). The program TMHMM (Krogh et al., 2001) predicts a transmembrane domain (TMD) between residues 643– 661 (Figure 1B). A comparison of the TMD domain of the Arabidopsis CASP with homologues from different species indicates the presence of highly conserved residues among different CASP proteins (Figure 1B, see also Gillingham et al., 2002). Strikingly, several residues central in the TMD are invariant across species. Furthermore, a comparison of the putative TMD region of CASP to those of golgin-84 and giantin from different species reveals that some of these residues are also conserved in both golgin 84 and giantin (Gillingham et al., 2002). These observations on CASP structure and the comparison with the other golgins suggest that AtCASP may be a plant membrane protein that shares structural properties with known CASP of different species as well as with known integral membrane golgins such as golgin-84 and giantin. and giantin of other kingdoms (Linstedt and Hauri, 1993; Bascom et al., 1999; Jakymiw et al., 2000; Misumi et al., 2001; Gillingham et al., 2002), we fused AtCASP to a fluorescent protein (Figure 1C) and expressed it in tobacco leaf epidermal cells (Figure 2). Laser confocal microscopy observations on a green fluorescent protein fusion of AtCASP (GFP-AtCASP) indicated that the fusion labelled punctate structures (Figure 2) which were motile in the cell (data not shown). The observed pattern was similar to the known distribution of Golgi stacks in tobacco leaf epidermal cells and BY-2 cells (Boevink et al., 1998; Nebenführ et al., 1999). To ensure the identity of the GFP-AtCASPlabelled punctate structures, tobacco leaf epidermal cells were co-transformed with the CASP fusion and a known Golgi marker, the Arabidopsis H/KDEL receptor (ERD2) fused to the yellow fluorescent protein (ERD2-YFP, Boevink et al., 1998; Brandizzi et al., 2002a, b; Figure 3A–C). ERD2-YFP labelled punctate structures (Figure 3B, arrowheads) that co-localized with those labelled by GFP-AtCASP (Figure 3C, arrowheads). ERD2 overexpression can lead to redistribution of the receptor to the ER in mammalian cells (Tang et al., 1993), but there is currently no published evidence for this in plants. However, to rule out the possibility of interference of overexpressed ERD2 with the distribution of GFP-AtCASP, we analysed the subcellular localization of GFPAtCASP with another known Golgi marker, ST-YFP (Brandizzi et al., 2002a, b). As shown in Figure 3D–F, GFP-AtCASP localized at the Golgi apparatus labelled by ST-YFP, confirming the results obtained with ERD2-YFP (Figure 3A–C) AtCASP targets the Golgi apparatus in plant cells and the targeting motifs are encoded in its C-terminus Targeting to the plant Golgi apparatus In order to ascertain whether AtCASP localizes at the Golgi apparatus like CASP and the other integral membrane golgins, golgin-67, golgin-84 Figure 2. When expressed alone in leaf epidermal cells, GFPAtCASP shows a punctate distribution. Bar = 5 lm. 115 Figure 3. AtCASP localizes at the Golgi apparatus in tobacco leaf epidermal cells. (A–C) A GFP fusion to AtCASP (A) targets small punctate structures in tobacco leaf epidermal cells. (B) The known Golgi marker, ERD2-YFP (Boevink et al., 1998; Brandizzi et al., 2002a, b), labels the Golgi apparatus (arrowheads) and the ER network (arrow). (C) Merged image of A and B, showing colocalization of the GFP-AtCASP and ERD2-YFP exclusively at the Golgi apparatus (arrowheads). (D–F) GFP-AtCASP (D) in co-expression with ST-YFP (E) colocalized at the Golgi apparatus highlighted by ST-YFP as shown in the merged image (F). (G) The N-terminal domain of AtCASP fused to GFP localizes in the cytoplasm. As typical for a cytosolic GFP fluorescence (daSilva et al., 2004), organelles are highlighted in negative contrast (arrowheads). (H–J) The C-terminal domain of AtCASP fused to YFP targets punctate structures (H). These colocalize with ERD2-GFP (I) at the Golgi apparatus as shown in the merged image of H and I (J, arrowheads). Scale bars in C, F, G, J = 5 lm. on the localization of GFP-AtCASP at the Golgi apparatus. Together, these data indicate that AtCASP localizes at the Golgi apparatus in plant cells, as has been shown for the mammalian CASP homologue in human cells (Gillingham et al., 2002) and for the other integral membrane golgins such as giantin and golgin-84 (Linstedt and Hauri, 1993; Bascom et al., 1999; Misumi et al., 2001). Identification of Golgi targeting domains of AtCASP To identify the regions of AtCASP that target the protein to the Golgi apparatus, we fused the DNA encoding either the N-terminal residues (1–564aa) or the C-terminal residues (565–689aa) to a fluorescent protein sequence (Figure 1C), and expressed the fusion proteins in tobacco leaf epidermal cells (Figure 3 G–J). The N-terminal GFP fusion (GFP-AtNCASP), which contains the N-terminal non-coiled-coil region and the long predicted coiled-coil region (Figure 1C), was found to distribute in the cytoplasm (Figure 3G). This result indicates that the N-terminus of CASP is not sufficient for targeting AtCASP to the Golgi apparatus. In contrast, the C-terminal YFP fusion (YFP-AtCCASP; Figure 1C) that contains the predicted noncoiled-coil C-terminal domain of CASP localized to punctate structures (Figure 3H), as observed for the full length fusion. Subsequent co-localization experiments using the Golgi marker ERD2-GFP 116 confirmed that these punctate structures labelled by YFP-AtCCASP are in fact Golgi bodies (Figure 3I–J). These data indicate that the C-terminus rather than the N-terminus of AtCASP has encoded information for entering the endomembranes and localizing at the Golgi apparatus, as is the case for the human CASP (Gillingham et al., 2002). To confirm that the C-terminal sequence of AtCASP contains the integral membrane domain of the protein as predicted by structural analysis software, we carried out subcellular fractionation experiments in the presence or absence of the detergent Triton X-100 on tobacco leaves expressing either a YFP-AtCCASP fusion or ssNGFP-HDEL, a soluble secreted GFP bearing a glycosylation peptide (Batoko et al., 2000; daSilva et al., 2005) and a retention/retrieval HDEL signal (Figure 4A). The fusion proteins were detected on a western blot with antibodies c Figure 4. AtCASP is a type II membrane protein (A) Western blot with GFP antiserum of protoplasts expressing ssNGFPHDEL or YFP-AtCCASP and fractionated into cytosol and vacuoles (S1), microsomal contents (S2) and cell membranes (P). In control samples ()Triton) the signal due to YFP-AtCCASP (empty arrowhead) was found mostly in the pellet containing the cell membranes, while that due to ssNGFP-HDEL was found mostly in the microsomal contents. In samples treated with Triton X-100 (+Triton) the signal due to YFPAtCCASP was found mainly in the soluble phase of the cell extracts as an obvious consequence of solubilization of the cell membranes. Similarly, a significant proportion of the signal for ssNGFP-HDEL was found in S1 due to disruption of the microsomal membranes. Negative control (UT, untransformed protoplasts). The YFP-AtCCASP band detected in S2 is likely due to incomplete sedimentation of the microsomes in the cell fractionation procedure. Partial rupture of the microsomal membranes in the absence of Triton X-100 resulted in the release of some ssNGFP-HDEL into the soluble S1 phase. (B) Western blot on proteinase K treatment of microsomes containing YFP-AtCCASP and ssNGFP-HDEL. Treatment with proteinase K (+) resulted in complete loss of signal for YFP-AtCCASP compared with the sample in the absence of proteinase K ()), whereas the ssNGFP-HDEL signal was present in both lanes, although a slight reduction in signal intensity was observed in the presence of the proteinase, due to partial rupture of the microsomes during preparation. This indicates that the YFP portion of the AtCCASP fusion is exposed on the external face of the microsomes. (C) Western blot showing tunicamycin treatment of NGFP-AtCASP. No shift in molecular weight is observed for NGFPAtCASP when 20 lg/ml tunicamycin was added (+), while a significant change can be seen for ssNGFP-HDEL. against the fluorescent protein. Consistent with the prediction that the plant CASP is an integral membrane protein, YFP-AtCCASP was detected mainly in the membrane pellet of the cell extracts (Figure 4A AtCCASP)Triton, lane P). In contrast, ssNGFP-HDEL was found predominantly in the soluble fraction extracted from the microsomes (figure 4A HDEL)Triton, lane S2). Weak signals were detected in the S1 and P lanes, probably due to rupturing of some of the microsomes during the initial osmotic shock (S1) or trapping of the protein in the membrane fraction (P). To further ascertain the distribution of the protein fusion into membranes, we treated the cell extracts with Triton X-100, a detergent that solubilizes the membranes. As a result of this treatment, the AtCCASP fusion should partition into the soluble phase of the cell extracts. As expected, treatment with Triton X-100 shifted the YFP-AtCCASP signal to the soluble 117 phase of the cell extracts (Figure 4A AtCCASP + Triton, lane S1). Similarly, a redistribution of a significant portion of ssNGFP-HDEL to the cytosolic phase was observed due to the disruption of the microsomal membranes (Figure 4A HDEL + Triton, lane S1). In order to confirm the prediction data on the orientation of the TMD of CASP, we examined the topology of the YFP-AtCCASP fusion by means of proteinase K protection. As a control to establish the integrity of the membranes, we used the secreted form of GFP bearing the ER retention signal HDEL, ssNGFP-HDEL. The rationale of this experiment was that if the TMD assumed a type II orientation, the YFP-AtCCASP signal would be exposed on the outside of the microsomes and would therefore not be protected from the protease. As shown in Figure 4B, the peptide due to YFP-AtCCASP was visible in the untreated sample signal but it was not present in the sample treated with proteinase K. The signal of ssNGFPHDEL was only partially affected, indicating that the microsomes were mostly intact in the experiment; the slight reduction observed is likely due to the rupturing of some microsomes during the initial osmotic shock (compare with Figure 4A HDEL)Triton). This control rules out the possibility that the digestion of YFP-AtCCASP with proteinase K was due to complete rupture of the microsomes. This experiment indicates that the TMD of AtCASP assumes a type II orientation. To further demonstrate the orientation of the TMD of AtCASP, we treated cells expressing a fusion of GFP containing an N-glycosylation site (NGFP; Batoko et al., 2000) to the N-terminus of AtCASP (NGFP-AtCASP; Figure 1C) with tunicamycin, an inhibitor of N-glycosylation (Shamu, 1997). NGFP-AtCASP localized at the Golgi apparatus as does GFP-AtCASP (data not shown). The N-terminal domain of CASP possesses a glycosylation consensus sequence in addition to that on the GFP moiety, increasing the chance that a shift in molecular weight would be observed if this part of the protein were found within the lumen of the secretory pathway. However, according to the predicted type II orientation of the protein, the glycosylation sites would both be in the cytosol and would therefore not be glycosylated. Concomitant with this hypothesis, Figure 4C shows that treatment with tunicamycin has no effect on the molecular weight of NGFP- AtCASP, while treatment of cells expressing ssNGFP-HDEL with tunicamycin gives a significant shift in the molecular weight of the protein synthesized during the drug treatment. This indicates that neither the NGFP moiety nor AtCASP itself was able to be glycosylated due to the cytosolic location of the N-terminus, while the NGFP fused to the HDEL motif is found within the ER lumen where proteins responsible for glycosylation are active. Taken together, these results show that AtCASP is a protein of the Golgi apparatus. The absence of a signal peptide at the N-terminus of the protein sequence and the orientation of the TMD with the N-terminus in the cytosol and a C-terminus in the lumen of the Golgi apparatus suggest that the plant CASP is likely to be a type II membrane protein, as is the human homologue (Gillingham et al., 2002). This is the same structure as determined experimentally not only in the mammalian CASP but also in other integral membrane golgins such as golgin-84 and giantin (Linstedt and Hauri, 1993; Bascom et al., 1999; Misumi et al., 2001; Gillingham et al., 2002). The Arabidopsis CASP is able to target the mammalian Golgi apparatus AtCASP shares a large degree of homology with its mammalian counterpart. To further explore the identity of AtCASP as a putative Golgi matrix protein, we aimed to investigate whether it could target the mammalian Golgi apparatus. To do so, we subcloned a GFP-AtCASP fusion in a mammalian expression vector and transiently expressed it in monkey cells (Figure 5A). Observations carried out with a laser scanning confocal microscope indicated that the fluorescence of GFP-AtCASP localized at a large perinuclear structure that resembled the mammalian Golgi apparatus (Ward and Brandizzi, 2004). To confirm the identity of this structure, we co-transformed monkey cells with GFP-AtCASP and a red fluorescent protein fusion of a TMD domain and cytosolic tail of a rat sialyl transferase (ST-RFP; Figure 5B), which is known to target the mammalian and plant Golgi apparatus (Munro, 1995; Saint-Jore et al., 2002). The ST-RFP fluorescence was distributed to the Golgi apparatus (Figure 5B) and it colocalized with that of GFP-AtCASP (Figure 5C). 118 Figure 5. AtCASP targets the mammalian Golgi apparatus. An African green monkey cell coexpressing GFP-AtCASP (A) and ST-RFP (B). ST-RFP is a known marker of the Golgi apparatus. (C) GFP and RFP fluorescence signals overlap, indicating that AtCASP localizes at the mammalian Golgi apparatus. Scale bar = 5 lm. These data indicate that the plant CASP contains domains that can efficiently target to the mammalian Golgi and reinforce the identity of the plant CASP as a transmembrane matrix protein. A tyrosine residue that is crucial for Golgi localization of CASP in mammalian cells has no effect on the Golgi distribution of this protein in plants The TMD of CASP contains a tyrosine residue which is conserved across species (Figure 1B). A GFP fusion of the human CASP bearing mutation of the tyrosine to leucine was found to localize at the ER and not the Golgi apparatus in mammalian cells (Gillingham et al., 2002). To ascertain what role this conserved amino acid plays in AtCASP for localization at the mammalian Golgi, we mutated the conserved tyrosine and expressed it as a GFP fusion in monkey cells (GFP-AtCASPY648L, Figures 1C, 6A–C). We observed that the GFP-AtCASPY648L mutant was distributed to the ER in these cells, as shown by co-localization experiments with an RFP fusion to a known mammalian ER marker, luman (Lu and Misra, 2000). These data confirm that the plant CASP has a tyrosine motif residue in the TMD that influences the localization of the protein in mammalian cells in a similar manner to that of the human homologue (Gillingham et al., 2002). To investigate whether the mechanisms of CASP localization to the Golgi apparatus are conserved between animals and plants, we expressed a GFP fusion of AtCASPY648L in tobacco leaf epidermal cells. Unlike its localization in mammalian cells, the plant mutant was found to label exclusively the Golgi apparatus (Figure 6 D–F) of tobacco leaf epidermal cells. These data indicate that although plant CASP contains domains that are conserved among kingdoms, sorting requirements for reaching the plant Golgi apparatus appear to be independent of the tyrosine motif, in great contrast to the situation in mammalian cells. This suggests that ER to Golgi transport may not rely entirely on conserved mechanisms between plants and mammals. Discussion Identification of a plant Golgi matrix protein Several different Golgi matrix proteins have been identified and characterized in yeast and mammals (reviewed by Barr and Short, 2003). In this study we have demonstrated the existence of a plant Golgi integral membrane matrix protein, an Arabidopsis homologue of the mammalian protein CASP (Gillingham et al., 2002). Golgi matrix proteins are thought to be important for maintenance of the structure of the Golgi apparatus, and may also be involved in tethering of transport vesicles through Rab GTPases (Barr, 1999; Moyer et al., 2001; Short et al., 2001; Valsdottir et al., 2001; Gillingham and Munro, 2003). The role of Rab GTPases is being unraveled in plants, and current data indicate roles of these proteins in various transport routes within the cell (Batoko et al., 2000; Ueda et al., 2000, 2001; Rutherford and Moore, 2002; Nahm et al., 2003; Bolte et al., 2004; Kotzer et al., 2004; Preuss et al., 2004). It therefore seems likely that further plant Golgi matrix proteins may exist that could specifically interact with different Rabs, but these have yet to be identified. The characterization of at least one plant Golgi matrix protein suggests that, 119 Figure 6. AtCASP contains signals that are important for localization of the protein at the Golgi apparatus in mammalian cells. (A–C) An African green monkey cell coexpressing the GFP-AtCASPY648L fusion (A) and an ER marker, luman-RFP (B). The signal due to GFP is found in the ER as shown in the merged image in C. (D–F) Tobacco leaf epidermal cells expressing a GFPAtCASPY648L fusion (D) show that the YFP fluorescence localizes at the Golgi apparatus as indicated by a colocalization with the Golgi marker, ERD2-YFP (E). No localization of CASP at the membranes of the ER was verified as shown by the absence of GFP labeling at the ER network highlighted by ERD2-YFP (arrowhead). (F) Merged image of D and E. Scale bars in C and F = 5 lm. despite the morphological differences in Golgi organization between plants and mammals (Ward and Brandizzi, 2004), there may be similarities between the systems in terms of the protein composition and function of the Golgi matrix. Targeting of AtCASP to the Golgi has different requirements in plants and animals Our data indicate that AtCASP has similarities to its mammalian homologue with regard to sequence, domain structure and intracellular localization. AtCASP is a type II transmembrane protein, with a short luminal domain and an extensive coiled-coil cytosolic domain that may be involved in interactions with other proteins, or in forming homodimers as has been shown for the mammalian homologue (Gillingham et al., 2002). Indeed, the homology between the plant and mammal CASP proteins is such that AtCASP is targeted to the Golgi in mammalian cells. However, when we mutated a conserved tyrosine residue in the transmembrane domain that is essential for Golgi targeting of mammalian CASP (Gillingham et al., 2002), no effect could be detected in the localization of the protein to the plant Golgi apparatus. Conversely, expression of the mutated AtCASP in mammalian cells resulted in its distribution in the ER. We conclude that despite the similarities between the transport systems of plants and animals, there are also discrepancies in the mechanisms by which some proteins are exported to the Golgi apparatus. The role of the tyrosine residue in mediating the transport of CASP in mammalian cells is unclear. Gillingham et al. (2002) reported that mutation of the tyrosine residue does not affect the ability of mammalian CASP to dimerize, indicating that dimerization is not a prerequisite for ER export. It is possible that the tyrosine is involved in interactions with other proteins that act as receptors for selecting cargo for transport to the Golgi apparatus. If this were the case, one could postulate that homologues of these putative receptors do not exist in plants, and that AtCASP therefore relies on other mechanisms for export from the ER. To gain further insights on the involvement of specific residues that govern ER sorting in plants, it would be useful in future to express the mammalian CASP in a plant system. The length of the transmembrane domain in type I membrane-spanning proteins is important 120 in determining their destinations (Brandizzi et al., 2002b), although no data have yet been published to show whether a similar rule applies to type II membrane-spanning proteins. The predicted 19amino acid transmembrane domain of AtCASP would likely be sufficient to transport the protein to the Golgi apparatus if type II membrane proteins behave in the same manner as type I. Our findings suggest that in plants the length of the transmembrane domain may supersede a tyrosine motif for export of proteins from the ER. Our data do not exclude that specific signals in the cytosolic domain of AtCASP may be responsible for mediating its transport to the Golgi apparatus. Two types of export signals have recently been identified in plant transmembrane proteins (Contreras et al., 2004; Yuasa et al., 2005), opening up the possibility that similar signals may be present in many other plant proteins. Misumi et al. (2001) have shown that the first 100 amino acids of the cytosolic domain adjacent to the transmembrane segment are essential for the correct targeting of several golgins in mammals. This corresponds to our finding that the C-terminal 125 amino acids of AtCASP (giving 78 amino acids in the cytosolic domain) are sufficient to instigate transport of the protein to the Golgi apparatus. Further investigation into the transport mechanisms of type II transmembrane proteins will be necessary in order to clarify the means by which AtCASP travels to the plant Golgi. Conclusions Our findings open fascinating new avenues of research on the role of plant matrix proteins in maintenance of the structure of the Golgi apparatus and on protein transport occurring in this organelle in plant cells. Deletion of the Saccharomyces cerevisiae CASP-homologue, COY1 does not affect viability, but restores normal growth to cells lacking the Golgi soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) Gos1p (Gillingham et al., 2002). It has also been established that CASP contains an internal coiled-coil motif that is required for cytohesin binding both in vitro and in COS-1 cells (Mansour et al., 2002). The specificity of the coiled-coil of CASP is not restricted to cytohesin, however, because it is also capable of interacting with other members of the cytohesin/ ARNO family, ARNO and ARNO3 (Mansour et al., 2002). The function of CASP in plant cells is unknown but some clues are provided by its Golgi localization, structure and sequence similarity to its homologues in other species and other golgins such as golgin-84 and giantin (Lindstedt and Hauri, 1993; Bascom et al., 1999; Gillingham et al., 2002). The exclusive localization of AtCASP at the Golgi apparatus suggests that the protein either leaves the ER rapidly or cycles with a slow rate from the Golgi apparatus. Therefore, At-CASP fusion proteins may serve as cleaner Golgi markers because they do not exhibit the weak ER staining observed for the other Golgi markers ST-YFP and ERD2-YFP. This feature may be at the basis of the involvement of this protein in events of either vesicular tethering or establishment and maintenance of the Golgi apparatus. The identification of the binding partners of CASP in plant cells will shed light on the role of this golgin in the organization, structure and trafficking pathways of the Golgi apparatus in plants. Acknowledgements We acknowledge for financial support the University of Saskatchewan and the Department of Biology, U of S, CFI and Canada Research Chair (CRC) grants to F.B for the development of this work. S.H. is supported by a CRC Provincial Operating Fund and a Department of Biology Post-Doctoral Award. CRC Provincial Operating Fund and Graduate College Studies Award are acknowledged for the support of G.S. CRC Provincial Operating Fund supports L.B. For her MSc studentship, L.R. is indebted to a University of Saskatchewan New Faculty Award. References Alvarez, C., Garcia-Mata, R., Hauri, H.P. and Sztul, E. 2001. The p115-interactive proteins GM130 and giantin participate in endoplasmic reticulum-Golgi traffic. J. Biol. Chem. 276(4) 2693–2700. Barr, F.A. 1999. A novel Rab6-interacting domain defines a family of Golgi-targeted coiled-coil proteins. Curr. Biol. 9(7) 381–384. 121 Barr, F.A., Nakamura, N. and Warren, G. 1998. Mapping the interaction between GRASP65 and GM130, components of a protein complex involved in the stacking of Golgi cisternae. EMBO J. 17(12) 3258–3268. Barr, F.A. and Short, B. 2003. Golgins in the structure and dynamics of the Golgi apparatus. Curr. Opin. Cell Biol. 15(4) 405–413. Bascom, R.A., Srinivasan, S. and Nussbaum, R.L. 1999. Identification and characterization of golgin-84, a novel Golgi integral membrane protein with a cytoplasmic coiledcoil domain. J. Biol. Chem. 274(5) 2953–2962. Batoko, H., Zheng, H.Q., Hawes, C. and Moore, I. 2000. A Rab1 GTPase is required for transport between the endoplasmic reticulum and Golgi apparatus and for normal Golgi movement in plants. Plant Cell 12(11) 2201–2218. Berger, B., Wilson, D.B., Wolf, E., Tonchev, T., Milla, M. and Kim, P.S. 1995. Predicting coiled coils by use of pairwise residue correlations. Proc. Natl. Acad. Sci. USA 92(18) 8259–8263. Boevink, P., Oparka, K., Santa Cruz, S., Martin, B., Betteridge, A. and Hawes, C. 1998. Stacks on tracks: the plant Golgi apparatus traffics on an actin/ER network. Plant J. 15(3) 441–447. Bolte, S., Brown, S. and Satiat-Jeunemaitre, B. 2004. The N-myristoylated Rab-GTPase m-Rabmc is involved in postGolgi trafficking events to the lytic vacuole in plant cells. J. Cell Sci. 117(6) 943–954. Brandizzi, F., Snapp, E.L., Roberts, A.G., LippincottSchwartz, J. and Hawes, C. 2002a. Membrane protein transport between the endoplasmic reticulum and the Golgi in tobacco leaves is energy dependent but cytoskeleton independent: evidence from selective photobleaching. Plant Cell 14(6) 1293–1309. Brandizzi, F., Frangne, N., Marc-Martin, S., Hawes, C., Neuhaus, J.M. and Paris, N. 2002b. In plants the destination for single pass membrane proteins is markedly influenced by the length of the hydrophobic domain. Plant Cell 14(5) 1077–1092. Cao, X., Ballew, N. and Barlowe, C. 1998. Initial docking of ER-derived vesicles requires Uso1p and Ypt1p but is independent of SNARE proteins. EMBO J. 17(8) 2156–2165. Chen, C.A. and Okayama, H. 1988. Calcium phosphatemediated gene transfer: a highly efficient transfection system for stably transforming cells with plasmid DNA. Biotechniques 6(7) 632–638. Contreras, I., Yang, Y., Robinson, D.G. and Aniento, F. 2004. Sorting Signals in the Cytosolic Tail of Plant p24 Proteins Involved in the Interaction with the COPII Coat. Plant Cell Physiol. 45(12) 1779–1786. Crofts, A.J., Leborgne-Castel, N., Hillmer, S., Robinson, D.G., Phillipson, B., Carlsson, L.E., Ashford, D.A. and Denecke, J. 1999. Saturation of the endoplasmic reticulum retention machinery reveals anterograde bulk flow. Plant Cell 11(11) 2233–2248. daSilva, L.L., Snapp, E.L., Denecke, J., Lippincott-Schwartz, J., Hawes, C. and Brandizzi, F. 2004. ER export sites and Golgi bodies behave as single mobile secretory units in plant cells. Plant Cell 16(7) 1753–1771. daSilva, L.L., Taylor, J.P., Hadlington, J.L., Hanton, S.L., Snowden, C.J., Fox, S.J., Foresti, O., Brandizzi, F. and Denecke, J. 2005. Receptor salvage from the prevacuolar compartment is essential for efficient vacuolar protein targeting. Plant Cell 17(1) 132–148. Denecke, J., Rycke, R.De and Botterman, J. 1992. Plant and mammalian sorting signals for protein retention in the endoplasmic reticulum contain a conserved epitope. EMBO J. 11(6) 2345–2355. Dohlman, J.G., Lupas, A. and Carson, M. 1993. Long chargerich alpha-helices in systemic autoantigens. Biochem. Biophys. Res. Commun. 195(2) 686–696. Drew, J.E., Bown, D. and Gatehouse, J.A. 1993. Sequence of a novel plant ras-related cDNA from Pisum sativum. Plant Mol. Biol. 21(6) 1195–1199. Fridmann-Sirkis, Y., Siniossoglou, S. and Pelham, H.R. 2004. TMF is a golgin that binds Rab6 and influences Golgi morphology. BMC Cell Biol. 5(1) 18. Gillingham, A.K., Pfeifer, A.C. and Munro, S. 2002. CASP, the alternatively spliced product of the gene encoding the CCAAT-displacement protein transcription factor, is a Golgi membrane protein related to giantin. Mol. Biol. Cell 13(11) 3761–3774. Gillingham, A.K. and Munro, S. 2003. Long coiled-coil proteins and membrane traffic. Biochim. Biophys. Acta 1641(2–3) 71–85. Gilson, P.R., Vergara, C.E., Kjer-Nielsen, L., Teasdale, R.D., Bacic, A. and Gleeson, P.A. 2004. Identification of a Golgilocalised GRIP domain protein from Arabidopsis thaliana. Planta 219(6) 1050–1056. Grebe, M., Xu, J., Mobius, W., Ueda, T., Nakano, A., Geuze, H.J., Rook, M.B. and Scheres, B. 2003. Arabidopsis sterol endocytosis involves actin-mediated trafficking via ARA6positive early endosomes. Curr. Biol. 13(16) 1378–1387. Haseloff, J., Siemering, K.R., Prasher, D.C. and Hodge, S. 1997. Removal of a cryptic intron and subcellular localization of green fluorescent protein are required to mark transgenic Arabidopsis plants brightly. Proc Natl Acad Sci USA 94(6) 2122–2127. Inaba, T., Nagano, Y., Nagasaki, T. and Sasaki, Y. 2002. Distinct localization of two closely related Ypt3/Rab11 proteins on the trafficking pathway in higher plants. J. Biol. Chem. 277(11) 9183–9188. Jakymiw, A., Raharjo, E., Rattner, J.B., Eystathioy, T., Chan, E.K. and Fujita, D.J. 2000. Identification and characterization of a novel Golgi protein, golgin-67. J. Biol. Chem. 275(6) 4137–4144. Jürgens, G 2004. Membrane trafficking in plants. Annu. Rev. Cell Dev. Biol. 20: 481–504. Kjer-Nielsen, L., Teasdale, R.D., Vliet, C.van and Gleeson, P.A. 1999. A novel Golgi-localisation domain shared by a class of coiled-coil peripheral membrane proteins. Curr. Biol. 9(7) 385–388. Kotzer, A.M., Brandizzi, F., Neumann, U., Paris, N., Moore, I. and Hawes, C. 2004. AtRabF2b (Ara7) acts on the vacuolar trafficking pathway in tobacco leaf epidermal cells. J. Cell Sci. 117(26) 6377–6389. Krogh, A., Larsson, B., Heijne, G.von and Sonnhammer, E.L 2001. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305(3) 567–580. Lesa, G.M., Seemann, J., Shorter, J., Vandekerckhove, J. and Warren, G. 2000. The amino-terminal domain of the golgi protein giantin interacts directly with the vesicle-tethering protein p115. J. Biol. Chem. 275(4) 2831–2836. Linstedt, A.D. and Hauri, H.P. 1993. Giantin, a novel conserved Golgi membrane protein containing a cytoplasmic domain of at least 350 kDa. Mol. Biol. Cell 4(7) 679–693. 122 Lu, R. and Misra, V. 2000. Potential role for luman, the cellular homologue of herpes simplex virus VP16 (alpha gene trans-inducing factor), in herpesvirus latency. J. Virol. 74(2) 934–943. Mansour, M., Lee, S.Y. and Pohajdak, B 2002. The N-terminal coiled coil domain of the cytohesin/ARNO family of guanine nucleotide exchange factors interacts with the scaffolding protein CASP. J. Biol. Chem. 277(35) 32302–32309. Misumi, Y., Sohda, M., Tashiro, A., Sato, H. and Ikehara, Y. 2001. An essential cytoplasmic domain for the Golgi localization of coiled-coil proteins with a COOH-terminal membrane anchor. J. Biol. Chem. 276(9) 6867–6873. Moyer, B.D., Allan, B.B. and Balch, W.E. 2001. Rab1 interaction with a GM130 effector complex regulates COPII vesicle cis-Golgi tethering. Traffic 2(4) 268–276. Munro, S. 1995. An investigation into the role of transmembrane domains in Golgi protein retention. EMBO J. 14(19) 4695–4704. Munro, S. and Nichols, B.J. 1999. The GRIP domain – a novel Golgi-targeting domain found in several coiled-coil proteins. Curr. Biol. 9(7) 377–380. Nahm, M.Y., Kim, S.W., Yun, D., Lee, S.Y., Cho, M.J. and Bahk, J.D. 2003. Molecular and biochemical analyses of OsRab7, a rice Rab7 homolog. Plant Cell Physiol. 44(12) 1341–1349. Nebenführ, A., Gallagher, L.A., Dunahay, T.G., Frohlick, J.A., Mazurkiewicz, A.M., Meehl, J.B. and Staehelin, L.A. 1999. Stop-and-go movements of plant Golgi stacks are mediated by the acto-myosin system. Plant Physiol. 121(4) 1127–1142. Neumann, U., Brandizzi, F. and Hawes, C. 2003. Protein transport in plant cells: in and out of the Golgi. Ann. Bot. 92(2) 167–180. Palade, G. 1975. Intracellular aspects of the process of protein synthesis. Science 189(4200) 347–358. Phillipson, B.A., Pimpl, P., daSilva, L.L., Crofts, A.J., Taylor, J.P., Movafeghi, A., Robinson, D.G. and Denecke, J. 2001. Secretory bulk flow of soluble proteins is efficient and COPII dependent. Plant Cell 13(9) 2005–2020. Pimpl, P., Movafeghi, A., Coughlan, S., Denecke, J., Hillmer, S. and Robinson, D.G. 2000. In situ localization and in vitro induction of plant COPI-coated vesicles. Plant Cell 12(11) 2219–2236. Preuss, M.L., Serna, J., Falbel, T.G., Bednarek, S.Y. and Nielsen, E. 2004. The Arabidopsis Rab GTPase RabA4b localizes to the tips of growing root hair cells. Plant Cell 16(6) 1589–1603. Puthenveedu, M.A. and Linstedt, A.D. 2001. Evidence that Golgi structure depends on a p115 activity that is independent of the vesicle tether components giantin and GM130. J Cell Biol 155(2) 227–238. Rose, A., Manikantan, S., Schraegle, S.J., Maloy, M.A., Stahlberg, E.A. and Meier, I. 2004. Genome-wide identification of Arabidopsis coiled-coil proteins and establishment of the ARABI-COIL database. Plant Physiol. 134(3) 927– 939. Rutherford, S. and Moore, I. 2002. The Arabidopsis Rab GTPase family: another enigma variation. Curr. Opin. Plant. Biol. 5(6) 518–528. Saint-Jore, C.M., Evins, J., Batoko, H., Brandizzi, F., Moore, I. and Hawes, C. 2002. Redistribution of membrane proteins between the Golgi apparatus and endoplasmic reticulum in plants is reversible and not dependent on cytoskeletal networks. Plant J. 29(5) 661–678. Sambrook, J., Fritsch, E.F. and Maniatis, T. 1989. Molecular Cloning: a laboratory manual. Cold Spring Harbour Laboratory Press, USA. Sanderfoot, A.A., Kovaleva, V., Bassham, D.C. and Raikhel, N.V. 2001. Interactions between syntaxins identify at least five SNARE complexes within the Golgi/prevacuolar system of the Arabidopsis cell. Mol. Biol. Cell 12(12) 3733–3743. Seemann, J., Jokitalo, E.J. and Warren, G. 2000. The role of the tethering proteins p115 and GM130 in transport through the Golgi apparatus in vivo. Mol. Biol. Cell 11(2) 635–645. Shamu, C.E. 1997. Splicing together the unfolded-protein response. Curr. Biol. 7(2) R67–70. Short, B., Preisinger, C., Korner, R., Kopajtich, R., Byron, O. and Barr, F.A. 2001. A GRASP55-rab2 effector complex linking Golgi structure to membrane traffic. J. Cell Biol. 155: 877–883. Shorter, J. and Warren, G. 1999. A role for the vesicle tethering protein, p115, in the post-mitotic stacking of reassembling Golgi cisternae in a cell-free system. J. Cell Biol. 146: 57–70. Slusarewicz, P., Nilsson, T., Hui, N., Watson, R. and Warren, G. 1994. Isolation of a matrix that binds medial Golgi enzymes. J. Cell Biol. 124(4) 405–413. Sonnichsen, B., Lowe, M., Levine, T., Jamsa, E., DiracSvejstrup, B. and Warren, G. 1998. A role for giantin in docking COPI vesicles to Golgi membranes. J. Cell Biol. 140(5) 1013–1021. Staehelin, L.A. and Moore, I. 1995. The plant Golgi apparatus: structure, functional organization and trafficking mechanisms. Annu. Rev. Plant Phys. Plant Mol. Biol. 46: 261–288. Tang, B.L., Wong, S.H., Qi, X.L., Low, S.H. and Hong, W. 1993. Molecular cloning, characterization, subcellular localization and dynamics of p23, the mammalian KDEL receptor. J. Cell Biol. 120(2) 325–328. Ueda, T., Anai, T., Tsukaya, H., Hirata, A. and Uchimiya, H. 1996. Characterization and subcellular localization of a small GTP-binding protein (Ara-4) from Arabidopsis: conditional expression under control of the promoter of the gene for heatshock protein HSP81y1. Mol. Gen. Genet. 250(5) 533–539. Ueda, T., Matsuda, N., Uchimiya, H. and Nakano, A. 2000. Modes of interaction between the Arabidopsis Rab protein, Ara4, and its putative regulator molecules revealed by a yeast expression system. Plant J. 21(4) 341–349. Ueda, T., Yamaguchi, M., Uchimiya, H. and Nakano, A. 2001. Ara6, a plant-unique novel type Rab GTPase, functions in the endocytic pathway of Arabidopsis thaliana. Embo J. 20(17) 4730–4741. Valsdottir, R., Hashimoto, H., Ashman, K., Koda, T., Storrie, B. and Nilsson, T. 2001. Identification of rabaptin5, rabex-5, and GM130 as putative effectors of rab33b, a regulator of retrograde traffic between the Golgi apparatus and ER. FEBS Lett. 508(2) 201–209. Volchuk, A., Ravazzola, M., Perrelet, A., Eng, W.S., Di Liberto, M., Varlamov, O., Fukasawa, M., Engel, T., Sollner, T.H., Rothman, J.E. and Orci, L. 2004. Countercurrent distribution of two distinct SNARE complexes mediating transport within the Golgi stack. Mol. Biol. Cell 15(4) 1506–1518. Ward, T.H. and Brandizzi, F. 2004. Dynamics of proteins in Golgi membranes: comparisons between mammalian and plant cells highlighted by photobleaching techniques. Cell Mol. Life Sci. 61(2) 172–185. Yuasa, K., Toyooka, K., Fukuda, H. and Matsuoka, K. 2005. Membrane-anchored prolyl hydroxylase with an export signal from the endoplasmic reticulum. Plant J. 41: 81–94.