Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
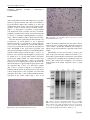
Struct Chem (2008) 19:203–208 DOI 10.1007/s11224-007-9273-8 ORIGINAL RESEARCH Demonstration of a protein with enhanced resistance to proteinase K in transmissible cytopathic condition elicited by cell-free lysate of free-living ameba Naegleria gruberi Are microbial proteins capable of converting mammalian proteins into prion-like pathogens? A probable pathomechanism for some postinfectious degenerative sequelae and for natural prion disease in sheep, goat and cervids Miklos Füzi Æ Judit Pászti Æ Ágnes Gyuris Æ Erika Orosz Æ János Minárovits Æ Zsuzsa Szénási Received: 12 October 2007 / Accepted: 14 November 2007 / Published online: 19 January 2008 Ó Springer Science+Business Media, LLC 2008 Abstract The cell-free lysate of free-living amebae Naegleria gruberi and Naegleria fowleri were reported to elicit cytopathic effect in various cell lines that could be indefinitely transmitted by the culture media. The causative agent showed sensitivity to treatments detrimental to proteins while resisted exposures damaging to nucleic acids. Here we demonstrate that subsequent to exposure to N. gruberi lysate mild digestion with proteinase K reveals the presence of a protein band in HeLa cells absent from control cell lines. Though the small quantity of this protein with enhanced resistance to proteinase K relative to the total protein content of the sample has proved to date insufficient for its purification, we suppose that it is a human cellular protein that assumed altered conformation in a prion-like fashion. The conformational conversion could have been trigerred by an ameba protein in the lysate. In addition, we showed that HeLa cells treated with N. gruberi lysate display elevated cathepsin B activity which is assumed to be a secondary response to the accumulation of the proteinase K-resistant protein. We propose that a number of degenerative sequelae following previous microbial infections in mammals could have a similar pathomechanism. Moreover, epidemiological data strongly suggest that natural prion disease in sheep, goat and cervids may also have an etiology linked to prior infection/colonization with a microbe, as it had already been proposed by one of us. M. Füzi (&) J. Pászti Á. Gyuris E. Orosz J. Minárovits Z. Szénási National Center for Epidemiology, Gyali út 2-6, 1097 Budapest, Hungary e-mail: [email protected] Keywords Prion Ameba Postinfectious Transmission Proteinase K resistant Cathepsin B Autoimmune Rheumatic fever Introduction It is well established that some proteins are capable of turning into infectious agents and cause diseases in both mammals and fungi [1, 2]. These proteinaceous pathogens are called prions after Stanley B. Prusiner’s landmark discovery published in 1982 [3]. Since the number of prions has been expanding in the last two decades it is reasonable to suppose that there remain many more unrecognized proteins capable of assuming an infectious character. Between 1971 and 1999 a series of papers were published by T. H. Dunnebacke and her coworkers on the cytopathic effect elicited by the cell-free lysate of two free-living amebae N. gruberi and N. fowleri [4–14]. The cytopathic effect which turned out to be the consequence of apoptosis could be demonstrated in various mammalian and avian cell lines and proved indefinitely transmissible with the culture media of cells having undergone cytopathogenicity. A comprehensive investigation of the causative agent revealed its proteinaceous nature: physico-chemical treatments detrimental to proteins destroyed the agent while various exposures damaging nucleic acids failed to impact infectivity [10–14]. In addition, the agent’s size proved so small— initial measurements yielded a mol. wt. in the 50,000 D range [10], subsequent estimates gave a mol. wt. of 36,000 D [12]—that it is inconceivable that it could harbor sufficient amount of nucleic acid to code for its protein content. It was supposed that an ameba protein capable of replicting itself in 123 204 Struct Chem (2008) 19:203–208 mammalian and avian cells could correspond to the agent [9– 13]. Moreover, the investigators hinted that the infectious protein could be a cathepsin B-like cystein proteinase [14]. In this study, we prepared cell-free N. gruberi lysate and tested its ability to elicit cytopathic effect. We could reproduce the cytopathogenicity reported earlier by Dunnebacke and coworkers in HeLa cell line [13] and confirmed that the pathogenic effect was transmissible with the culture media of damaged cells. In addition, we demonstrated that a protein showing enhanced resistance to proteinase K is present in cell lines showing cytopathogenicity. This protein is conspicuously absent in control cells. Furthermore, in contrast to control cell lines an elevated cathepsin B activity could be demonstrated in cell lines undergoing cytopathic degeneration. However, the protein showing partial resistance to proteinase K had a higher mol. wt. than cathepsin B, thus, the two proteins must not be identical. The elevation of cathepsin B activity could be the consequence of the accumulation of the partly proteinase K-resistant protein. The day before treatment with lysate, the cells were subcultured (1 9 105 cells/3 mL complete medium). Experiments were performed in 75 mm2 flasks (Greiner) with cells reaching semiconfluency. At treatment the medium was replaced by medium containing the N. gruberi lysate. The physical condition of the cells were followed by daily microscopical investigation. After cytopathogenicity became evident the supernatants of the control and treated cells were removed, cleared from cell debris by centrifugation and kept at -70 °C until use. The cells were removed from the surface of the culture flask by rubber, washed in complete medium and stored at -70 °C until further use. For treating the cells N. gruberi lysate (prepared from 1200–1800 cystas/lL in 2 mL of phosphate buffer) was used in 1009 dilution in fresh medium. The cells showed cytopathic effect following 2–4 days. When the cytopathic effect was transmitted between HeLa cell lines 25% or 50% of the normal cell line’s medium was replaced with medium from the cytopathic cell line. Cells showed cytopathogenicity within 2 or 4 days, respectively. Materials and methods Demonstration of protein showing enhanced resistance to proteinase K The N. gruberi strain was purchased from ATCC (accession number:30133). HeLa cell line was bought from ATCC (CCL-2) and cultured in Eagle’s Minimum Essential Medium (Sigma-Aldrich, St. Louis, USA) with 10 per cent of bovine serum. Naegleria gruberi was grown on PAGE agar (Nebotrade, Budapest, Hungary). After an incubation of about 30 days at 22 °C cells were counted using a counting chamber then the amebae were harvested in 2 mL of PPYG medium (Nebotrade, Budapest, Hungary) and centrifuged with 1200g for 5 min at 4 °C. The pellet was suspended in 2 mL of 0.01 M phosphate buffer (0.01 M Na2HPO4 9 2H2O pH 7.2) and centrifuged with 1200g for 5 min at 4 °C. The collected amebae were suspended again in 2 mL of phosphate buffer and subjected to repeated freeze-thawing (4 times). The resulting lysate was clarified by centrifugation (10,000g, 30 min 40 °C). The supernatant fluid was passed through a 0.45 lm Millipore filter. The lysate was then divided into samples and stored at -70 °C. Cells beginning to show cytopathic effect were removed and mixed with lysis buffer at a ratio of 1:4. (Lysis buffer: Triton X-100 0.5%; sodium deoxycholate 0.5%; Tris HCl 5 mM (pH 7.4); NaCl 150 mM; EDTA 5 mM) then centrifuged at 1000g at 4 °C for 5 min. The determination of total protein content was made by the Bradford method. Bradford reagent was purchased from Sigma-Aldrich (St. Louis, USA). Tests were performed according to manufacturer’s instruction. Some of the samples were treated with proteinase K (20 lg/mL; 30 min; 37 °C). Digestion with proteinase K was stopped with 1mM of PMSF on ice. The samples were then ultracentrifuged (230,000g for 45 min). All reagents were purchased from Sigma-Aldrich (St. Louis, USA). Control HeLa cells were grown under same conditions until decay became apparent (10–11 days) then processed like lysate treated samples. SDS-PAGE was performed as described by Laemmli [15] with an acrylamide concentration of 12%. Protein bands were visualised with PageBlue Protein Staining Solution (Fermentas, Vilnius, Lithuania). Demonstration of cytopathic effect Measurement of cathepsin B activity HeLA cells were kept in CO2 thermostate at 37 °C and were subcultured twice weekly. The activity of cathepsin B was examined by Cathepsin B Detection Kit; Substrate III (Calbiochem-Merck, Preparation of N. gruberi lysate 123 Struct Chem (2008) 19:203–208 Darmstadt, Germany) recommendation. according 205 to manufacturer’s Results After treating HeLa cell lines with ameba lysate cytopathic effect was apparent from the 2nd to 4th day of incubation. Cytopathogenicity implied the rounding up of cells and detachment from the surface followed by complete disintegration. Control cell lines always remained orthodox. (Figs. 1, 2). Subsequent to inoculation of normal HeLa cells with media from cytopathic cell lines an indistinguishable cytopathic effect could be observed from the 2nd to 4th day of incubation. Indistinguishable cytopathogenicity could be transmitted with no deterioration in effect. Since the infectious agent had been demonstrated to be proteinaceous, we tested whether or not it displayed enhanced resistance to proteinase K, a feature characteristic for prions. We examined if a protein with enhanced resistance to proteinase K was present in the cytopathic cell lines. SDS-PAGE of the lysate-treated cytopathic and untreated normal HeLA cell lines proved indistinguishable without treatment with proteinase K (Fig. 3, lanes 1, 2). However, after treatment with 20 lg/mL of proteinase K for 30 min at 37 °C a protein band could be observed in lysate-treated samples which was clearly distinct from those in the control sample (Fig. 3, lanes 3, 4; band indicated by arrow). Comparing the proteinase K-treated and untreated samples it is apparent that though proteins in both the cytopathic (Fig. 3, lanes 2, 4) and control samples (Fig. 3, lanes 1, 3) began to degrade as a consequence of treatment with proteinase K, degradation was faster in the control sample relative to the cytopathic one at the band denoted by arrow (Fig. 3, lane 4). This band denser than its equivalent in the control sample (Fig. 3, lane 3). In Fig. 1 HeLa control cell line/incubation time: 2 days Fig. 2 Cytopathic effect on HeLa cell line exposed to N. gruberi lysate/incubation time: 2 days addition, the band located directly below it in the control is absent from the cytopathic sample. Thus, this band obviously contains a protein that should be more resistant to digestion by proteinase K than its equivalent in the control sample. Unfortunately, we have not been able to obtain a pure band of this protein showing enhanced resistance to proteinase K by applying higher concentrations of the enzyme or subsequent to extended period of incubation. (Data not shown.) This could be the consequence of its relatively small quantity in the sample compared to those of other proteins. Fig. 3 Lane 1: Negative control HeLA cell line Lane 2: Cytopathic HeLa cell line exposed to ameba lysate Lane 3: Negative control HeLA cell line treated with 20 lg/mL proteinase K for 30 min at 37 °C Lane 4: Cytopathic HeLa cell line exposed to ameba lysate treated with 20 lg/mL proteinase K for 30 min at 37 °C 123 206 Struct Chem (2008) 19:203–208 The mol. wt. of the partially proteinase K-resistant protein is about 60,000 D, however, the detected band could comprise an already truncated protein, thus, its real (original) mol. wt. remains unknown. The fluorogenic substrate specific for cathepsin B (Calbiochem-Merck, Darmstadt, Germany) showed an elevated activity of the enzyme in cell lines undergoing cytopathic effect. In contrast, no increase in the activity of cathepsin B could be demonstrated in control cell lines (Figs. 4, 5). Discussion We confirmed the earlier results of Dunnebacke and coworkers [13] that the cell-free lysate of the free-living ameba N. gruberi elicits a transmissible cytopathic effect on HeLa cell lines. Investigating the etiology of the transmissible cytopathic effect, we have demonstrated the presence of a protein showing enhanced resistance to proteinase K in the cytopathic samples that is absent in control cell lines. Moreover, we proved that in contrast to control samples, the activity of cathepsin B is elevated in the cytopathic cell lines. The quantity of the proteinase K-resistant protein is certainly small relative to the total protein content of the sample. This could be the consequence of a short period of multiplication since the protein can accumulate for merely 2–4 days before cytopathogenesis destroys the cell line. The detected protein with enhanced resistance to proteinse K must be distinct from cathepsin B. Human cathepsin B has a mol. wt. of about 38,000 D, while our protein displayed a much higher mol. wt. in SDS-PAGE even following partial enzymatic degradation. Fig. 4 Demonstration of cathepsin B activity in negative control HeLa cell line with Cathepsin B fluorogenic substrate III. /Calbiochem/ after two days of incubation 123 Fig. 5 Demonstration of cathepsin B activity in HeLa cell line exposed to N. gruberi lysate with Cathepsin B fluorogenic substrate III. /Calbiochem/ after 2 days of incubation Though it was earlier supposed that an ameba protein could replicate itself in the cytopathic cell lines [9–14] we consider this possibility to be remote because there is no conceivable mechanism for a protein to multiply itself and maintain its amino acid sequence in a cell void of its gene. It is much more probable that a human protein with altered conformation has been detected. The conformational conversion could have been triggered by a microbial protein in the ameba lysate conferring enhanced resistance to proteinase K to the human protein and the ability of selfsustaining replication in a prion-like fashion. That a microbial prion can be capable of eliciting conformational change in a protein of another species has recently been demonstrated by Tessier and Lindquist [16]. Though Tessier and Lindquist’s work concerns closely related fungi there is no reason to doubt that triggering of conformational change can occur also between proteins of more distant species, including microbes and humans. It is well-known that proteins of various species are capable of adhering to each other. There is no reason to doubt that conversion into a prion-like conformational state in a protein can be triggered by a normal protein of a distant species. We suppose that the pathomechanism proposed by us is common and many degenerative sequelae developing subsequent to microbial infection could have similar etiologies. The best-documented postmicrobial sequel is probably rheumatic fever linked to previous infection with group A streptococci. It is generally accepted that rheumatic fever is caused by the so-called M protein of group A streptococcus, a filamentous molecule protruding from the surface of the bacterium. M protein shows homology to cardiac myosin and to various extracellular matrix proteins and has been proved to adhere to and aggregate human Struct Chem (2008) 19:203–208 collagen [17, 18]. This characteristic of M protein seems to be crucial to the development of rheumatic fever since aggregated collagen was demonstrated to be a primary autoantigen in affected patients [18, 19]. The question arises that if the proteinase K-resistant protein detected by us is really reminiscent of the infectious prion, i.e., prion protein with altered conformation (PrPSc), could prion disease also have a similar ‘‘microbial pathomechanism’’ described by us. The answer is certainly yes. As it was earlier shown by one of us [20, 21] epidemiological data obtained with scrapie (prion disease in sheep and goat) and with chronic wasting disease (prion disease in cervids) strongly suggest that these infections are transmitted by a vector under natural conditions which could be a microbe residing in the alimentary tract of the animals. This microbe that we are not in a position to identify might be responsible for the horizontal transmission of natural scrapie and natural chronic wasting disease. Since horizontal spread of infection has not been observed in other species with prion disease the microbe should command a limited host range and should not be capable of colonizing (infecting) the alimentary tract of species other than sheep, goat and cervids. The microbe is not a vector in the sense we use this term when describing the epidemiology of infectious diseases. It should not carry the proteinaceous agent itself. It should, however, harbor a structural protein capable of adhering to normal prion protein (PrPC) and altering its conformation turning it thereby into an infectious prion. A recent paper provides a strong circumstantial evidence for a microbe playing a crucial role in the transmission and triggering of scrapie and chronic wasting disease under natural conditions. Mathiason et al. [22] reported that the saliva of animals with natural chronic wasting disease proved infectious when administered perorally to healthy deer. If we consider that the amount of PrPSc is very low in the salivary glands [23] the most plausible explanation for Mathiason et al’s [22] observation remains—ast it was already suggested [21]—that it is not PrPSc that transmits infection with the saliva. The saliva of sheep, goat and cervids with natural prion disease should harbor the ‘‘triggering microbe’’ [21]. The microbe if introduced perorally may colonize the oral cavity of an animal and convert the conformation of PrPC [21]. Though unlike prion disease rheumatic fever is considered an autoimmune condition autoantibodies have also been demonstrated in animals and humans with prion disease [24–27]. In addition, streptococcal M protein commands more than 100 serotypes displaying slightly diverse amino acid sequences and eliciting rheumatic fever with distinct efficiencies [17]. This feature of M protein is strongly reminiscent of the way the strains of prion emanate, thus, the microbial protein eliciting natural scrapie 207 and chronic wasting disease could also have multiple variants (serotypes) converting PrPC into prions with diverse conformations. Finally, we have demonstrated that the activity of cathepsin B is elevated in cell lines exposed to the lysate of N. gruberi and showing cytopathic effect. We consider this finding a response to the accumulation of the detected host protein with enhanced resistance to proteinase K. The exact role of cathepsin B in the cytopathic condition remains to be established. However, it is certainly not an accident that the activity cathepsin B is elevated also in prion disease [28]. Acknowledgements We thank Thelma D. Dunnebacke for her advice and Ákos Toth and Csaba Bognár for their excellent technical contributions. This study was funded by Diagon Ltd. Budapest, Hungary. References 1. Gains MJ, LeBlanc AC (2007) Canadian association of neurosciences review: prion protein and prion diseases: the good and the bad. Can J Neurol Sci 2:126–145 2. Wickner RB, Edskes HK, Shewmaker F, Nakayashiki T (2007) Prions of fungi: inherited structures and biological roles. Nat Rev Microbiol 8:611–618 3. Prusiner SB (1982) Novel proteinaceous infectious particles cause scrapie. Science 4542:136–144 4. Dunnebacke TH, Schuster FL (1971) Infectious agent from a free-living soil amoeba, Naegleria gruberi. Science 8:516 5. Dunnebacke TH, Schuster FL (1974) An infectious agent associated with amebas of the genus Naegleria. J Protozool 2:327 6. Schuster FL, Dunnebacke TH (1974) Growth at 37 °C of the EGs strain of the amoeboflagellate Naegleria gruberi containing viruslike particles. I. Nuclear changes. J Invertebr Pathol 2:172 7. Schuster FL, Dunnebacke TH (1974) Growth at 37 °C of the EGs strain of the amoeboflagellate Naegleria gruberi containing viruslike particles. II. Cytoplasmic changes. J Invertebr Pathol 2:182 8. Schuster FL, Dunnebacke TH (1974) Virus-like particles and an unassociated infectious agent in amoebae of the genus Naegleria. Ann Soc Belg Med Trop 4–5:359 9. Schuster FL, Dunnebacke TH (1976) Development and release of virus-like particles in Naegleria gruberi EGS. Cytobiologie 1:131 10. Dunnebacke TH, Schuster FL (1977) The nature of a cytopathogenic material present in amebae of the genus Naegleria. Am J Trop Med Hyg 3:412 11. Dunnebacke TH, Schuster FL (1985) Morphological response of cultured cells to Naegleria amoeba cytopathogenic material. J Cell Sci 75:1–16 12. Dunnebacke TH, Dixon JS (1989) NACM, a cytopathogen from Naegleria ameba: purification of monoclonal antibody, and immunoreactive material in NACM-treated vertebrate cell cultures. J Cell Sci 93:391–401 13. Dunnebacke TH, Dixon JS (1990) NACM, a cytopathogenic protein from Naegleria gruberi, EGs; purification, production of monoclonal antibody, and the immunoidentification of a product that develops in NACM-treated vertebrate cell cultures. J Protozool 4:11 14. Dunnebacke TH, Walen KH (1999) A protein from Naegleria amoebae causes apoptosis in chick embryo and CHO cells after they become confluent. In Vitro Cell Dev Biol Anim 5:252 123 208 15. Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 5259:680–685 16. Tessier PM, Lindquist S (2007) Prion recognition elements govern nucleation, strain specificity and species barriers. Nature 7144:556 17. Guilherme L, Ramasawmy R, Kalil J (2007) Rheumatic fever and rheumatic heart disease: genetics and pathogenesis. Scand J Immunol 2–3:199–207 18. Dinkla K, Rohde M, Jansen WT, Kaplan EL, Chhatwal GS, Talay SR (2003) Rheumatic fever-associated Streptococcus pyogenes isolates aggregate collagen. J Clin Invest 12:1905–1912 19. Dinkla K, Nitsche-Schmitz DP, Barroso V, Reissmann S, Johansson HM, Frick IM, Rohde M, Chhatwal GS (2007) Identification of a streptococcal octapeptide motif involved in acute rheumatic fever. J Biol Chem 26:18686 20. Fuzi M (1999) Is the pathogen of prion disease a microbial protein? Med Hypotheses 2:91 21. Fuzi M (2007) Is natural prion disease in sheep, goat and cervids triggered by a microbial protein? Prion 2007 Book of abstracts p 99, http://www.prion2007.com 22. Mathiason CK, Powers JG, Dahmes SJ, Osborn DA, Miller KV, Warren RJ, Mason GL, Hays SA, Hayes-Klug J, Seelig DM, Wild MA, Wolfe LL, Spraker TR, Miller MW, Sigurdson CJ, Telling GC, Hoover EA (2006) Infectious prions in the saliva and blood of deer with chronic wasting disease. Science 5796:133 123 Struct Chem (2008) 19:203–208 23. Vascellari M, Nonno R, Mutinelli F, Bigolaro M, Di Bari MA, Melchiotti E, Marcon S, D’Agostino C, Vaccari G, Conte M, De Grossi L, Rosone F, Giordani F, Agrimi U (2007) PrPSc in salivary glands of scrapie-affected sheep. J Virol 9:4872 24. Sotelo J, Gibbs CJ Jr, Gajdusek DC (1980) Autoantibodies against axonal neurofilaments in patients with Kuru and Creutzfeldt-Jakob disease. Science 4466:190–193 25. Bahmanyar S, Liem RK, Griffin JW, Gajdusek DC (1984) Characterization of antineurofilament autoantibodies in Creutzfeldt-Jakob disease. J Neuropathol Exp Neurol 4:369–375 26. Toh BH, Gibbs CJ Jr, Gajdusek DC, Goudsmit J, Dahl D (1985) The 200- and 150-kDa neurofilament proteins react with IgG autoantibodies from patients with kuru, Creutzfeldt-Jakob disease, and other neurologic diseases. Proc Natl Acad Sci USA 10:3485–3489 27. Toh BH, Gibbs CJ Jr, Gajdusek DC, Tuthill DD, Dahl D (1985) The 200- and 150-kDa neurofilament proteins react with IgG autoantibodies from chimpanzees with kuru or Creutzfeldt-Jakob disease; a 62-kDa neurofilament-associated protein reacts with sera from sheep with natural scrapie. Proc Natl Acad Sci USA 11:3894–3896 28. Zhang Y, Spiess E, Groschup MH, Burkle A (2003) Up-regulation of cathepsin B and cathepsin L activities in scrapie-infected mouse Neuro2a cells. J Gen Virol 8:83