Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Durante lo sviluppo, l’esperienza gioca un ruolo
determinante nel guidare l’emergere del
comportamento in maniera individuo specifica all’interno
di un trend maturativo comune alla specie.
La presenza di un determinato background genetico
costituisce la situazione al contorno in cui l’esperienza
opera.
Costituisce, ovvero, la base della formazione dei circuiti
neurali sulla cui maturazione l’esperienza agisce per
guidare l’emergenza del comportamento
Patrimonio genetico = potenzialità
ma anche vincolo, limite
per l’azione dell’esperienza
E’ sempre più evidente che fattori genetici e fattori
ambientali non sono indipendenti tra loro nel controllare
lo sviluppo del sistema nervoso e del comportamento;
ciò che è importante è l’interazione tra essi.
Donald Hebb riassunse molto efficacemente
questa idea rispondendo a un giornalista che gli
chiedeva chi, tra geni e ambiente, contribuisse
maggiormente alla personalità. Hebb rispose
domandando al giornalista se per l’area di un rettangolo
fosse più importante la base o l’altezza.
Studi che esaminano nell’uomo le interazioni
geni-ambiente nella probabilità di sviluppare
disturbi del comportamento o del tono
dell’umore
Approcci allo studio del ruolo di fattori genetici nel
comportamento e nei suoi disturbi
La storia recente della ricerca sul ruolo di fattori genetici nello sviluppo del
comportamento e nei suoi disturbi ha seguito tre grandi approcci, ciascuno con la sua
logica e le sue assunzioni.
Il primo approccio assume che vi sia una relazione diretta fra geni e
comportamento.
In particolare, assume che i geni causino i disturbi
del comportamento.
Lo scopo di questo approccio è stato di correlare la presenza di disturbi nel
comportamento con differenze individuali nella sequenza del DNA. “This has been
attempted using both linkage analysis and association analysis, with regard to many
psychiatric conditions such as depression, schizophrenia and addiction. Although a few
genes have accumulated replicated evidence of association with disorder, replication
failures are routine and overall progress has been slow.” (Caspi et al., 2006)
“Because of inconsistent findings, many scientists have despaired of the search for a
straightforward association between genotype and diagnosis, that is, for direct main
effects.” (Caspi et al., 2006).
Caspi et al. Nature Reviews Neuroscience 7, 583–590 (July 2006) | doi:10.1038/nrn1925
Il secondo approccio ha cercato di aggirare questo scoglio
cercando correlazioni fra il genotipo individuale e fenotipi
intermedi, chiamati endofenotipi.
“Endophenotypes are heritable neurophysiological, biochemical,
endocrinological, neuroanatomical or neuropsychological
constituents of disorders (es. from ASDs).” (Caspi et al., 2006)
Si assume che gli endofenotipi abbiano cause genetiche più
semplici rispetto al fenotipo completo. Quindi, l’assunzione dietro
questo approccio è che è più semplice identificare i geni associati
con endofenotipi che identificare i geni associati con il disturbo
con cui tali endofenotipi sono correlati.
Anche questo approccio, però, cerca un effetto principale di fattori
genetici nelle differenze interindividuali.
Caspi et al. Nature Reviews Neuroscience 7, 583–590 (July 2006) | doi:10.1038/nrn1925
Il terzo e più nuovo approccio cerca di incorporare nello studio delle
differenze interindividuali nel comportamento l’effetto dell’ambiente.
Questo approccio, detto approccio “interazioni geni-ambiente” differisce in
maniera forte dai due approcci precedenti, che cercano un “effetto principale”
dei fattori genetici.
“Main-effect approaches assume that genes cause disorder, an
assumption carried forward from early work that identified single-gene
causes of rare Mendelian conditions. “ (Caspi eta l., 2006).
L’approccio “interazioni geni-ambiente” (Gene x Environment, G x E) invece
assume che patogeni ambientali possano causare disturi del comportamento e
che il genotipo influenzi la suscettibilità a tali patogeni.
Si assume quindi che i geni siano fattori di rischio per la suscettibilità a
patogeni ambientali.
In contrasto con gli studi che cercano effetti principali del genotipo, secondo
l’approccio G xE non c’è attesa per una associazione diretta genecomportamento in assenza di patogeni ambientali (e viceversa).
Caspi et al. Nature Reviews Neuroscience 7, 583–590 (July 2006) | doi:10.1038/nrn1925
L’approccio G x E è scaturito da due tipi di
osservazioni:
1. Disturbi del comportamento e del tono
dell’umore hanno cause legate all’ambiente ed
all’esperienza individuale;
2. C’è eterogeneità nella risposta a “patogeni”
ambientali (resilienza/vulnerabilità)
“Like other non-communicable diseases that have common prevalence
in the population and complex multi-factorial aetiology, most mental
disorders have known non-genetic, environmental risk factors (that
is, predictors whose causal status is unproven).
Environmental risk factors for mental disorders discovered to date
include (but are not limited to):
maternal stress during pregnancy, maternal substance abuse during
pregnancy, low birth weight, birth complications, deprivation of normal
parental care during infancy,
childhood physical maltreatment, childhood neglect, premature parental
loss, exposure to family conflict and violence,
stressful life events involving loss or threat,
substance abuse, toxic exposures”
Caspi et al., 2006
I patogeni ambientali sono quindi in realtà non “causali”, ma
sono fattori di rischio. L’esposizione ad essi non sempre
causa un disturbo del comportamento.
Sia gli studi nell’uomo che gli studi in modelli animali hanno
consistentemente mostrato una grande variabilità
interindividuale nelle risposte comportamentali a patogeni
ambientali.
“Heterogeneity of response characterizes all known environmental
risk factors for psychopathology, including even the most
overwhelming of traumas.
Such response heterogeneity is associated with pre-existing
individual differences in temperament, personality, cognition and
autonomic physiology, all of which are known to be under genetic
influence.” (Caspi et al., 2006).
“The hypothesis of genetic moderation implies
that differences between individuals,
originating in the DNA sequence, bring about
differences between individuals in their
resilience or vulnerability to the environmental
causes of many pathological conditions of the
mind and body.”
Caspi et al., 2006
Esempi di interazioni geni x ambiente:
Sovraespressione del gene NMDA2B x ambiente arricchito,
(memoria migliore per animali wild type, ceiling effect per animali
con mutazione)
G x E nella suscettibilità a sviluppare dipendenza da sostanze:
Esempio: Cabib et al., 2000
Uso di linee di topi “Inbred”, largamente usati per la genetica del
comportamento.
Prima di entrare nel vivo dell’argomento, vi introduco il sistema
endogeno della ricompensa
Qui andare al file sistema endogeno della ricompensa
Saline
Drug of abuse
appaiato
Place preference
non appaiato
C57B6J
DBA
C57B6J
DBA
Conditioned place preference test, due background genetici diversi. C57, più
suscettibili a sviluppare dipendenza da sostanze d’abuso dei DBA
Descrizione del “conditioned Place Preference “test
Un periodo moderato di carenza di cibo (“food shortage”), che
costituisce una esperienza “ecologica” e piuttosto comune in
natura, può invertire o cancellare le differenze fra le linee C57
e DBA nella risposta comportamentale alla esposizione a
psicostimolanti. Quindi, una “sfida” ambientale può annullare la
differenza fra i genotipi.
Neuromodulatory system lesion
Ambiente normale
Food shortage
Place preference
C57
unpaired
paired
Place preference
DBA
unpaired
paired
“The period of food shortage occurred when the
animals were mature and was terminated before
the administration of amphetamine.
Strain differences in behavior appear highly
dependent on environmental experiences.
Consequently, to identify biological
determinants of behavior, in this case
resilience to addiction, an integrated
approach considering the interaction between
environmental and genetic factors needs to
be used.”
Cabib et al., 2000
Ventura et al., 2003
Trasformare i C57 in DBA modificando la
trasmissione cortico striatale
La corteccia prefrontale esercita un forte effetto
modulatorio sui meccanismi di dipendenza.
La trasmissione noradrenergica nella corteccia
prefrontale mediale (mpFC) modula ad esempio l’effetto
della anfetamina a livello del comportamento.
Il lavoro di Ventura et al mira a verificare che l’azione
modulatoria della noradrenaline (NE) a livello
prefrontale sia coinvolta nello sviluppo di
dipendenza dalle anfetamine.
Metodo:
Deplezione selettiva di NE nella mPFC nei C57BL/6J,
(molto suscettibili allo sviluppo di dipendenza da
anfetamine).
Scopo:
Valutare lo sviluppo di dipendenza da anfetamine con il
conditioned place preference.
Risultati:
Assenza di conditioned place preference nei topi C57
con deplezione prefrontale di NE (C57 sono diventati
DBA).
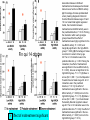
Effects of prefrontal cortical norepinephrine depletion on the preference scores shown by saline (A)
and amphetamine (B)-treated groups of C57 mice in conditioned place preference test. All data are
expressed as mean ± SE. *p < 0.05 compared with the unpaired compartment.
Ventura et al., 2003
In un secondo gruppo di esperimenti, Ventura et al.,
(2003) hanno mostrato che la deplezione
noradrenergica a livello prefrontale riduce fortemente
l’effetto della anfetamina nell’aumentare il rilascio di
dopamina dalla VTA al nucleo accumbens, in linea con
l’assenza di dipendenza.
“These results indicate that noradrenergic prefrontal
transmission, by allowing increased dopamine
release in the nucleus accumbens induced by
amphetamine, is a critical factor for the rewardingreinforcing effects of this drug.”
Ventura et al., 2003
“Later studies have shown that C57 and DBA mice differ
in their sensitivity to other drugs of abuse (cocaine and
morphine) in terms of drug induced conditioned place
preference and suggest that the two strains differ in
sensitivity to the positive incentive properties of
drugs of abuse (Orsini et al., 2005).” (Pascucci et al.,
2007)
Sembrerebbe quindi che lo sviluppo di dipendenza
dall’assunzione di sostanze, legato al rilascio di
dopamina nel nucleo accumbens (dopamina che è
rilasciata dai neuroni della VTA), sia sotto il controllo
opponente del sistema noradrenergico e dopaminergico
a livello della corteccia prefrontale. (Pascucci et al.,
2007).
In ultima analisi, il bilancio fra attività
noradrenergica e dopaminergica a livello
prefrontale potrebbe, regolando il rilascio di
dopamina nel nucleo accumbens, determinare la
suscettibilità allo sviluppo di dipendenza da
sostanze, attraverso una interazione geni x
ambiente.
Risultati simili per la risposta allo stress.
Esposizione a stressors inibisce il rilascio di dopamina
nel nucleo accumbens.
Nei C57, questo effetto è accompaganto da una forte
attivazione del sistema dopaminergico mesocorticale; a
livello comportamentale, questo corrisponde ad una
messa in atto di comportamenti “rinunciatari” e alla
presenza di anedonia.
Nei DBA si ha invece una risposta neuromodulatoria
prefrontale e comportamentale opposta.
“There may be a genetic control over the balance
between mesocortical and mesoaccumbens
dopamine responses to drugs and stress, which
sets the level of susceptibility for drug addiction,
stressful events and ultimately, interactions with
the environment.”
(Pascucci et al., 2007)
I polimorfismi: fattori di rischio, fattori
protettivi nei confronti di “patogeni”
ambientali.
(Si parla di polimorfismo genetico quando una variazione genetica ha una prevalenza maggiore
dell'1% nella popolazione. La variazione genetica può essere determinata da sostituzioni, delezioni
o inserzioni di basi nel DNA e può riguardare regioni codificanti e regioni non codificanti.)
Lavoro capostipite: Caspi et al., 2002
Role of genotype in the cycle of violence in maltreated children
“Childhood maltreatment is a universal risk factor for antisocial
behavior. Boys who experience abuse--and, more generally, those
exposed to erratic, coercive, and punitive parenting--are at risk of
developing conduct disorder, antisocial personality symptoms, and of
becoming violent offenders.
The earlier children experience maltreatment, the more likely
they are to develop these problems. But there are large differences
between children in their response to maltreatment. Although
maltreatment increases the risk of later criminality by about 50%,
most maltreated children do not become delinquents or adult
criminals.
The reason for this variability in response is largely unknown,
but it may be that vulnerability to adversities is conditional, depending
on genetic susceptibility factors.”
In questo studio, viene indagata l’ipotesi di una
suscettibilità genetica al maltrattamento infantile.
Si ipotizza in particolare che differenze interindividuali
nella funzionalità del gene per la monoamminoossidasi A
(MAOA), dovute ad un polimorfismo nel promotore del
gene stesso, influenzino la resilienza ad un ambiente
avverso precoce.
Ipotesi: “polimorphism in MAOA gene modifies the
influence of maltreatment on children's development
of antisocial behavior.”
Il gene per la MAOA è localizzato sul cromosoma X.
Esso codifica per l’enzima MAOA, che metabolizza
neurotrasmettitori quali la Noradrenalina (NE), serotonina (5-HT),
e dopamina (DA), rendendoli inattivi e facendone quindi terminare
l’azione a livello sinaptico.
Una mutazione che determina la produzione di una versione
meno funzionale della MAOA prolunga la vita media del
neurotrasmettitore, aumentandone l’efficacia sinapitca.
Deficit nella attività della MAOA dovuti a polimorfismi
genetici sono stati messi in relazione con fenotipi a maggior
aggressività sia in modelli animali che nell’uomo.
1: Increased aggression and increased levels of brain NE, 5-HT,
and DA were observed in a transgenic mouse line in which the
gene encoding MAOA was deleted, and aggression was
normalized by restoring MAOA expression.
2: In humans, a null allele at the MAOA locus was linked
with male antisocial behavior in a Dutch kindred.
Because MAOA is an X-linked gene, affected males with a single
copy produced no MAOA enzyme--effectively, a human knockout.
However, this mutation is extremely rare.
Evidence for an association between MAOA
and aggressive behavior in the human general
population remains inconclusive.
Circumstantial evidence suggests the hypothesis
that childhood maltreatment predisposes most
strongly to adult violence among children whose
MAOA is insufficient to constrain maltreatmentinduced changes to neurotransmitter systems.
Indeed, animal studies document that
maltreatment stress (e.g., maternal deprivation, peer
rearing) in early life alters NE, 5-HT, and DA
neurotransmitter systems in ways that can persist into
adulthood and can influence aggressive behaviors
(Harlow, Meaney).
“Maltreatment has lasting neurochemical correlates in
human children: psychobiological sequelae of child
maltreatment may be regarded as an
environmentally induced complex developmental
disorder, and although no study has ascertained
whether MAOA plays a role, it exerts an effect on all
aforementioned neurotransmitter systems.
Posttraumatic stress disorder in maltreated children is
associated with dysregulation of biological stress
systems, adverse brain development, and neuronal loss
in the anterior cingulate region of the medial prefrontal
cortex.”
(Caspi et al., 2000)
Deficient MAOA activity may dispose the
organism toward neural hyperreactivity to threat.
McDermott et al., 2009
Monoamine oxidase A gene (MAOA) has earned the nickname
“warrior gene” because it has been linked to aggression in
observational and survey-based studies.
McDermott and coworkers have performed an experiment,
synthesizing work in psychology and behavioral economics, which
suggests that aggression occurs with greater intensity and
frequency as provocation is experimentally manipulated upwards,
especially among low activity MAOA (MAOA-L) subjects.
Low MAOA activity may be particularly
problematic early in life, because there is
insufficient MAOB (a homolog of MAOA with
broad specificity to neurotransmitter amines) to
compensate for an MAOA deficiency.
Based on the hypothesis that MAOA
genotype can moderate the influence of
childhood maltreatment on neural systems
implicated in antisocial behavior, Caspi et al.
tested
whether antisocial behavior would be
predicted by an interaction between a gene
(MAOA) and an environment (maltreatment).
A well-characterized variable number
tandem repeat (VNTR) polymorphism exists
at the promoter of the MAOA gene, which is
known to affect gene expression.
Caspi’s group genotyped this polymorphism in
members of the Dunedin Multidisciplinary Health
and Development Study.
This birth cohort of 1,037 children (52% male)
has been assessed at ages
3, 5, 7, 9, 11, 13, 15, 18, and 21 and was virtually
intact (96%) at age 26 years.
The Dunedin Multidisciplinary Health and Development Study
(often referred to as the Dunedin Longitudinal Study) is a longrunning cohort study of 1037 people born over the course of a
year in Dunedin, New Zealand.
The original pool of study members were selected from those born
between 1 April 1972 and 31 March 1973 and still living in the
Otago region 3 years later.
Study members were assessed at age 3, and then at ages 5, 7, 9,
11, 13, 15, 18, 21, 26, 32 and, most recently, at age 38 (20102012). Future assessments are scheduled for ages 44 and 50.
Avshalom Caspi and Terrie Moffitt
During an assessment, study members are brought back to
Dunedin from wherever in the world they live. They participate in a
day of interviews, tests and surveys.
Sub-studies of the Dunedin Study include the on-going Parenting
Study which focuses on the Dunedin Study member and their first
three-year-old child;
and the Next Generation Study which involves the offspring of
Dunedin Study members as they turn 15 and looks at the
lifestyles, behaviours, attitudes and health of today's teenagers,
and aims to see how these have changed from when the original
Study Members were 15 (in 1987-88).
This means that information across three generations of the same
families will be available.
Great emphasis is placed on retention of study
members.
At the most recent (age 38) assessments, 96% of
all living eligible study members, or 961 people,
participated.
This is unprecedented for a longitudinal study,
with many others worldwide experiencing 20–
40% drop-out rates.
The Caspi et al. 2002 study.
The study offers three advantages for testing gene-environment
(G × E) interactions.
First, in contrast to studies of clinical samples, this study of a
representative general population sample avoids potential
distortions in association between variables.
Second, the sample has well-characterized environmental
adversity histories.
Between the ages of 3 and 11 years, 8% of the study children
experienced "severe" maltreatment, 28% experienced "probable"
maltreatment, and 64% experienced no maltreatment.
(Maltreatment groups did not differ on MAOA activity, suggesting
that genotype did not influence exposure to maltreatment.)
Third, the study has ascertained antisocial outcomes
rigorously.
Antisocial behavior is a complicated phenotype, and each method and data
source used to measure it (e.g., clinical diagnoses, personality checklists,
official conviction records) is characterized by different strengths and
limitations.
Using information from independent sources appropriate
to different stages of development, Caspi et al.,
examined four outcome measures.
A common-factor model fit the four measures of antisocial behavior well, with
factor loadings ranging from 0.64 to 0.74, showing that all four measures index
liability to antisocial behavior.
1 Adolescent conduct disorder was assessed according
to criteria of the Diagnostic and Statistical Manual of
Mental Disorders (DSM-IV);
2 Convictions for violent crimes were identified via the
Australian and New Zealand police;
3 A personality disposition toward violence was
measured as part of a psychological assessment at age
26;
4 Symptoms of antisocial personality disorder were
ascertained at age 26 by collecting information about the
study members from people they nominated as
"someone who knows you well."
Means on the composite
index of antisocial
behavior as a function of
MAOA activity and a
childhood history of
maltreatment .
MAOA activity is the gene
expression level associated
with allelic variants of the
functional promoter
polymorphism, grouped into
low and high activity;
childhood maltreatment is
grouped into 3 categories of
increasing severity.
The antisocial behavior
composite is standardized
(z score) to a M = 0 and
SD = 1; group differences
are interpretable in SD unit
differences (d).
A test of the interaction between MAOA activity and maltreatment revealed a significant G × E interaction
(P = 0.01). This interaction within each genotype group showed that
effect of childhood maltreatment on antisocial behavior was significantly weaker
among males with high MAOA activity (P = 0.03) than among males with low
MAOA activity (P < 0.001).
*
*
*
Fin qui 14 ottobre
*
*
*
Effect of maltreatment significant
Association between childhood
maltreatment and subsequent antisocial
behavior as a function of MAOA activity.
(A) Percentage of males (and standard
errors) meeting diagnostic criteria for
Conduct Disorder between ages 10 and
18. In a hierarchical logistic regression
model, the interaction between
maltreatment and MAOA activity was in
the predicted direction, P = 0.06. Probing
the interaction within each genotype
group showed that the effect of
maltreatment was highly significant in the
low-MAOA activity P < 0.001, and
marginally significant in the high-MAOA
group P = 0.09). (B) Percentage of males
convicted of a violent crime by age
26. The G × E interaction was in the
predicted direction, p = 0.05. Probing the
interaction, the effect of maltreatment
was significant in the low-MAOA activity
P < 0.001), but was not significant in the
high MAOA group P = 0.17. (C) Mean z
scores (M = 0, SD = 1) on the Disposition
Toward Violence Scale at age 26. G × E
interaction was in the predicted
direction, P = 0.10); the effect of
maltreatment was significant in the lowMAOA activity P = 0.002) but not in the
high MAOA group P = 0.17). (D) Mean z
scores (M = 0, SD = 1) on the Antisocial
Personality Disorder symptom scale at
age 26. The G × E interaction was in the
predicted direction P = 0.04); the effect of
maltreatment was significant in the lowMAOA activity P < 0.001) but not in the
high MAOA group P = 0.12).
Although individuals having the combination of
low-activity MAOA genotype and maltreatment were
only 12% of the male birth cohort, they accounted for
44% of the cohort's violent convictions, yielding an
attributable risk fraction (11%) comparable to that of
the major risk factors associated with cardiovascular
disease.
Moreover, 85% of cohort males having a lowactivity MAOA genotype who were severely
maltreated developed some form of antisocial
behavior.
“These findings provide initial evidence that a
functional polymorphism in the MAOA gene moderates
the impact of early childhood maltreatment on the
development of antisocial behavior in males.” (Caspi et al., 2000)
Questo lavoro è stato il primo a mostrare
chiaramente che un fattore genetico poteva rendere
resilienti nei confronti di un patogeno ambientale.
“With regard to research in psychiatric
genetics, knowledge about environmental context
might help gene-hunters refine their phenotypes.
Genetic effects in the population may be
diluted across all individuals in a given
sample, if the effect is apparent only among
individuals exposed to specific environmental
risks.
Environment effect in the population may be
masked across all individuals in a given
sample, if the effect is apparent only among
individuals carrying a specific polymorphism”
“With regard to research on child health, knowledge
about specific genetic risks may help to clarify risk
processes.
The search has focused on social experiences that may
protect some children, overlooking a potential protective role
of genes.
Genes are assumed to create vulnerability to
disease, but from an evolutionary perspective they
are equally likely to protect against environmental
insult.
Maltreatment studies may benefit from
ascertaining genotypes associated with sensitivity
to stress, and the known functional properties of
MAOA may point toward hypotheses, based on
neurotransmitter system development, about
how stressful experiences are converted into
antisocial behavior toward others in some, but not
all, victims of maltreatment.
Caspi et al., 2003
Influence of Life Stress on Depression: Moderation by a
Polymorphism in the 5-HTT Gene
“Across the life span, stressful life events that
involve threat, loss, humiliation, or defeat influence the
onset and course of depression.
However, not all people who encounter a stressful
life experience succumb to its depressogenic effect.”
Costello et al., 2002
The NIMH convened a multidisciplinary Workgroup of
scientists to review the field and the NIMH portfolio and to
generate specific recommendations.
To encourage a balanced and creative set of proposals,
experts were included within and outside this area of research, as
well as public stakeholders.
The Workgroup identified the need for expanded knowledge of
mood disorders in children and adolescents, noting important
gaps in understanding the onset, course, and recurrence of earlyonset unipolar and bipolar disorder. Recommendations included
the need for a multidisciplinary research initiative on the
pathogenesis of unipolar depression encompassing genetic and
environmental risk and protective factors.
Whether specific genes exacerbate or buffer the effect of
stressful life events on depression is beginning to be unravelled.
In this study, a functional polymorphism in the
promoter region of the serotonin transporter gene (5-HTT)
was used to characterize genetic vulnerability to depression
and to test whether 5-HTT gene variation moderates the
influence of life stress on depression.
The 5-HT system provides a logical source of candidate
genes for depression, because this system is the target of
selective serotonin reuptake–inhibitor drugs that are effective in
treating depression.
The 5-HTT has received particular attention because it is involved
in the reuptake of serotonin at brain synapses.
The promoter activity of the 5-HTT gene is
modified by sequence elements within the
proximal 5' regulatory region, designated
the 5-HTT gene-linked polymorphic region
(5-HTTLPR).
The short ("s") allele in the 5-HTTLPR is
associated with lower transcriptional
efficiency of the promoter compared with
the long ("l") allele.
Up to Caspi’s work, evidence for an association between
the short promoter variant and depression was
inconclusive.
Although the 5-HTT gene may not be directly
associated with depression, it could moderate the
serotonergic response to stress.
Three lines of experimental research suggested this
hypothesis of a gene-by-environment (G x E) interaction.
First, in mice with disrupted 5-HTT, homozygous
and heterozygous (5-HTT –/– and +/–) strains
exhibited more fearful behavior;
in response to stress they showed greater
increases in the stress hormone (plasma)
adrenocorticotropin compared to homozygous (5HTT +/+) controls, but in the absence of stress no
differences related to genotype were observed.
5-HTT KO mice
Behaviour in the elevated plus maze
The 5-HT1A receptor antagonist, WAY 100635,
produced anxiolytic-like effects in the EPM
Second, in rhesus macaques, whose length
variation of the 5-HTTLPR is analogous to that of
humans, the short allele is associated with decreased
serotonergic function [lower cerebro-spinal fluid (CSF)
concentrations of 5-HT metabolites] among monkeys
reared in stressful conditions but not among
normally reared monkeys.
Monkeys with deleterious early rearing experiences
were differentiated by genotype in cerebrospinal
fluid concentrations of the 5-HT metabolite, 5hydroxyindoleacetic acid, while monkeys reared
normally were not.
Questi risultati dimostrano che l’effetto del
genotipo (s/s, l/l, s/l) del 5-HTT sulla funzione
serotoninergica nel SNC dipende dall’ambiente.
Una cosa molto importante è che anche la
risposta allo stress è modificata in s/s carriers
esposti a stress precoce (peer rearing)
Caspi et al. Nature Reviews Neuroscience 7, 583–590 (July 2006) | doi:10.1038/nrn1925
Influence of exposure to early stress (peer rearing) on subsequent exaggerated responses
of the HPA responses to stress is conditioned by serotonin transporter gene promoter
variation (rh-5HTTLPR) in rhesus macaques. When exposed to stress later in life, peer-reared
animals with the s/l genotype had higher ACTH levels than animals with the l/l genotype. There
were no differences between genotypes among animals reared with their mothers
l/l
Trattatemi con
cura, sono un
s/s
s/s
An alert monkey and a depressed monkey.
"Depressed monkeys do not appear responsive to potential threats. It's that slumped,
collapsed body posture accompanied by a lack of responsiveness to environmental
events.“
As in humans, monkey depression is deeper than just mopey behavior. The depressed
macaques had less body fat, higher levels of stress hormones, lower bone density
(consistent with osteoporosis), higher cholesterol concentrations and high heart rates.
They were often subordinate in the community hierarchy and socially stressed by
dominant animals.
Third, human neuroimaging research suggests that the fear
response (and therefore the stress response) is mediated by
variations in the 5-HTTLPR. Humans with one or two copies of
the s allele exhibit greater amygdala neuronal activity to
fearful stimuli compared to individuals homozygous for the l
Effect of 5-HTT genotype on right
allele.
amygdala activity. Bar graphs
represent the mean BOLD fMRI
percent signal change in a region of
interest (ROI) comprising the entire
right amygdala in the s (n = 14) and
l (n = 14) groups collapsed across
both cohorts. Individual circles
represent the activity for each
subject in this ROI. Consistent with
the statistical parametric maps (Fig.
2), which identified significant voxels
within the right amygdala, analysis
of variance for the entire amygdala
ROI, including voxels that were not
differentially activated according to
statistical parametric mapping, still
revealed significant group
differences in the mean (±SEM)
BOLD fMRI percent signal change
[s group = 0.28 ± 0.08 and l
group = 0.03 ± 0.05;
F(1,26) = 6.84, P = 0.01].
Presi nel loro insieme, questi risultati
costituiscono un sufficiente razionale per
formulare l’ipotesi che variazioni a carico del
gene del 5-HTT possano moderare le reazioni ad
esperienze avverse, rendendo gli individui
resilienti o vulnerabili.
Caspi tested this G x E hypothesis among members of
the Dunedin Multidisciplinary Health and Development
Study. This representative birth cohort of 1037 children
(52% male) has been assessed at ages 3, 5, 7, 9, 11,
13, 15, 18, and 21 and was virtually intact (96%) at the
age of 26 years.
Stressful life events occurring after the 21st birthday
and before the 26th birthday were assessed with the
aid of a life-history calendar, a highly reliable method for
ascertaining life-event histories.
The 14 events included employment, financial, housing,
health, and relationship stressors.
30% of the study members experienced no
stressful life events; 25% experienced one event;
20%, two events; 11%, three events; and 15%,
four or more events.
There were no significant differences between
the three genotype groups in the number of
life events they experienced, F(2,846) = 0.56, P
= 0.59, suggesting that 5-HTTLPR genotype
did not influence exposure to stressful life
events.
* Main effect of
5-HTT genotype significance
* (p 0.06)
Results of multiple regression analyses estimating the association between number of stressful life
events (between ages 21 and 26 years) and depression outcomes at age 26 as a function of 5HT T genotype.
(A) Self-reports of depression symptoms. The main effect of 5-HT TLPR (i.e., an effect not conditional on
other variables) was marginally significant (P = 0.06), the main effect of stressful life events was
significant (P < 0.001), and the interaction between 5-HT TLPR and life events was in the predicted
direction (P = 0.02). The interaction showed that the effect of life events on self-reports of
depression symptoms was stronger among individuals carrying an s allele (P < 0.001 among
s/s homozygotes, and P < 0.001 among s/l heterozygotes) than among l/l homozygotes ( P = 0.08).
(B) Probability of major depressive episode. The main effect of 5-HT TLPR was not significant (P = 0.29),
the main effect of life events was significant (P < 0.001), and the G x E was in the predicted direction
(P = 0.056). Life events predicted a diagnosis of major depression among s carriers (P = 0.001
among s/s homozygotes, and P < 0.001 among s/l heterozygotes) but not among l/l
homozygotes (P = 0.24).
Probability of suicide ideation or attempt. The main effect of 5-HT TLPR was not
significant (P = 0.99), the main effect of life events was significant (P < 0.001), and the
G x E interaction was in the predicted direction (P = 0.051). Life events predicted
suicide ideation or attempt among s carriers (, P = 0.09 among s/s homozygotes,
and P < 0.001 among s/l heterozygotes) but not among l/l homozygotes (P = 0.62).
Caspi reasoned that if measure of life events
represents environmental stress, then the timing
of life events relative to depression must follow
cause-effect order and life events that occur after
depression should not interact with 5-HTTLPR to
postdict depression.
Caspi tested this hypothesis by substituting the age-26
measure of depression with depression assessed in
this longitudinal study when study members were 21
and 18 years old, before the occurrence of the
measured life events between the ages of 21 and 26
years.
Whereas the 5-HTTLPR x life events interaction
predicted depression at the age of 26 years, this same
interaction did not postdict depression reported at age
21 nor at the age of 18 years, indicating that the above
related finding is a true G x E interaction.
If 5-HTT genotype moderates the depressogenic
influence of stressful life events, it should moderate the
effect of life events that occurred not just in adulthood
but also of stressful experiences that occurred in earlier
developmental periods.
Based on this hypothesis, Caspi tested whether
adult depression was predicted by the interaction
between 5-HTTLPR and childhood maltreatment that
occurred during the first decade of life. Consistent
with the G x E hypothesis, the longitudinal prediction
from childhood maltreatment to adult depression was
significantly moderated by 5-HTTLPR.
The interaction showed that childhood
maltreatment predicted adult depression only
among individuals carrying an s allele but not
among l/l homozygotes.
Results of regression analysis estimating the association between childhood maltreatment
(between the ages of 3 and 11 years) and adult depression (ages 18 to 26), as a function of 5HT T genotype. Among the 147s/s homozygotes, 92 (63%), 39 (27%), and 16 (11%) study members were in
the no maltreatment, probable maltreatment, and severe maltreatment groups, respectively. Among the 435 s/l
heterozygotes, 286 (66%), 116 (27%), and 33 (8%) were in the no, probable, and severe maltreatment groups.
Among the 265 l/l homozygotes, 172 (65%), 69 (26%), and 24 (9%) were in the no, probable, and severe
maltreatment groups. The main effect of 5-HT TLPR was not significant (b = –0.14, SE = 0.11, z = 1.33, P =
0.19), the main effect of childhood maltreatment was significant (b = 0.30, SE = 0.10, z = 3.04, P = 0.002), and the
G x E interaction was in the predicted direction (b = –0.33, SE = 0.16, z = 2.01, P = 0.05). The interaction showed
that childhood stress predicted adult depression only among individuals carrying an s allele (b = 0.60, SE
= 0.26, z = 2.31, P = 0.02 among s/s homozygotes, and b = 0.45, SE = 0.16, z = 2.83, P = 0.01 among s/l
heterozyotes) and not among l/l homozygotes (b = –0.01, SE = 0.21, z = 0.01, P = 0.99).
Evidence of a direct relation between the 5-HTTLPR
and depression had been inconsistent, perhaps because
prior studies have not considered participants' stress
histories.
In this study, no direct association between the 5-HTT gene
and depression was observed.
Previous experimental paradigms, including 5-HTT
knockout mice, stress-reared rhesus macaques, and human
functional neuroimaging, have shown that the 5-HTT gene can
interact with environmental conditions, although these
experiments did not address depression.
Caspi’s study demonstrates that this G x E interaction
extends to the natural development of depression in a
representative sample of humans.
Cosa abbiamo imparato dai lavori di Caspi e da
altri simili
“The study of gene–environment
interactions has been the province of
epidemiology, in which genotypes,
environmental pathogen exposures and
disorder outcomes are studied as they
naturally occur in the human population. “
However, genetic epidemiology is limited for
understanding the biological mechanisms
involved in an interaction, and therefore its
potential will be better realized when it is
integrated with experimental neuroscience
(psychobiology).
Psychobiology can complement psychiatric
genetic epidemiology by specifying the more
proximal role of nervous system reactivity in the
gene–environment interaction.
Caspi et al. Nature Reviews Neuroscience 7, 583–590 (July 2006) | doi:10.1038/nrn1925
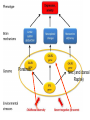
The building blocks
correspond to the three
elements of the triad: the
disorder, the
environmental pathogen
and the genotype. First,
evidence is needed about
which neural substrate
is involved in the
disorder. Second,
evidence is needed that
an environmental cause
of the disorder has
effects on variables
indexing the same
neural substrate. Third,
evidence is needed that a
candidate gene has
functional effects on
variables indexing that
same neural substrate.
It is this convergence of
environmental and
genotypic effects within
the same neural substrate
that allows for the
possibility of gene–
environment interactions.
Caspi et al. Nature Reviews Neuroscience 7, 583–590 (July 2006) | doi:10.1038/nrn1925
Psychobiology provides the building blocks for constructing hypotheses about gene–
environment interaction (a) that are tested against data (b), subsequently stimulating new
studies to illuminate the black box of biology (c) between the gene (G), the environmental
pathogen (E) and the disorder (D).
Several studies have sought to replicate the interaction
between the high- and low-activity MAOA genotypes and
maltreatment found by Caspi et al.; a recent metaanalysis revealed a significant pooled effect.
Positive replications of the interaction between 5HTT*long/5HTT*short genotypes and life stress have
also appeared, along with two failures to replicate.
Other studies have found G xE for other genes, such as
BDNF (Val/Met polimorphism), in relation to mood
disorders.
Brain galanin system genes interact with life
stresses in depression-related phenotypes.
Juhasz G, Hullam G, Eszlari N, Gonda X, Antal P, Anderson IM,
Hökfelt TG, Deakin JF, Bagdy G
Proc Natl Acad Sci U S A. 2014 Apr 22;111(16):E1666-73
Galanin is a stress-inducible neuropeptide and
cotransmitter in serotonin and norepinephrine neurons
with a possible role in stress-related disorders.
Galanin is widely distributed in the rodent and human
brain.
In rat it coexists with noradrenaline (NA) in the locus coeruleus
(LC) and with 5-HT in the dorsal raphe complex.
Like other peptide cotransmitters, galanin is released when
neurons fire in high-frequency bursts in response to strong
behavioral and pharmacological challenge
Variants in genes for galanin (GAL) and its receptors (GALR1,
GALR2, GALR3), conferred increased risk of depression and
anxiety in people who experienced childhood adversity or recent
negative life events in a European white population cohort totaling
2,361 from Manchester, United Kingdom and Budapest, Hungary.
The results suggest that the galanin pathway plays an important
role in the pathogenesis of depression in humans by increasing
the vulnerability to early and recent psychosocial stress.
The findings further emphasize the importance of
modeling environmental interaction in finding new
genes for depression
Forebrain
NAc (and dorsal
Raphe)
Caption for previous figure.
Galanin, a neuropeptide, and its receptors are colocalized in some
monoaminergic neurons in the brain. The galanin system is highly sensitive to
experimental and naturalistic stressors.
Recent analysis of human brain has shown that the GALR3 is the main
galanin receptor in NA-LC and probably 5-HT dorsal raphe nucleus cells, and
that the GALR1 is the main receptor in the forebrain.
Antidepressive effects may be achieved by (i) GALR3 antagonists, by
reinstating normal monoamine turnover in the brainstem, and by (ii) GALR1
antagonists in the forebrain by normalization of limbic system activity, or by
(iii) agonists at GALR2, promoting neuroprotection.
Genetic analysis suggests that GALR1 risk variants may compromise galanin
signaling during childhood, whereas GALR2 signaling may be influenced by
recent negative life events.
In addition, all four galanin system genes have relevant roles in the
development of depression-related phenotypes in those persons who were
highly exposed to life stressors.
Three-way interaction effect of 5-HTTLPR, BDNF
Val66Met, and childhood adversity on depression: a
replication study.
Comasco E, Åslund C, Oreland L, Nilsson KW.
Eur Neuropsychopharmacol. 2013 Oct;23(10):1300-6.
Both the serotonin transporter linked promoter region (5HTTLPR) and the brain-derived neurotrophic factor
(BDNF) Val66Met polymorphisms have been shown to
interact with unfavourable environment in relation to
depression symptoms and to depression diagnosis.
Several attempts have been made to study a three-way
interaction effect of these factors on depression, with
contradictory results.
This paper aimed to test the hypothesis of a three-way
interaction effect and to attempt at replication in an
independent population-based sample.
Family maltreatment and depression were self-reported
by an adolescent population-based cohort (N=1393)
from the county of Västmanland, Sweden.
DNA was isolated from saliva, and used for genotyping
of the 5-HTTLPR and BDNF Val66Met polymorphisms.
Neither 5-HTTLPR or BDNF genotypes separately, nor
in interaction with each other had any relation to
depression, however in an environment adjusted model
a two-way interaction and a three-way interaction effect
was found.
Both 5-HTTLPR and BDNF Val66Met interacted with
unfavourable environment in relation to depressive
symptoms.
Interazioni G x E nella lunghezza dei telomeri: effetti di
esperienze precoci avverse sulla integrità del DNA tramite i
telomeri
I telomeri impediscono la degradazione progressiva dei
cromosomi con rischio di perdita di informazione genetica.
Social disadvantage, genetic sensitivity, and
children's telomere length.
Mitchell C, Hobcraft J, McLanahan SS, Siegel SR, Berg A,
Brooks-Gunn J, Garfinkel I, Notterman D.
Proc Natl Acad Sci U S A. 2014 Apr 22;111(16):5944-9.
Disadvantaged social environments are associated with adverse
health outcomes. This has been attributed, in part, to chronic
stress.
Results:
Exposure to disadvantaged environments is associated
with reduced telomere length (TL) by age 9 years.
There is a significant associations between low income,
low maternal education, unstable family structure, harsh
parenting and TL.
These effects were moderated by genetic variants in
serotonergic and dopaminergic pathways.
Consistent with the differential susceptibility hypothesis,
subjects with the “vulnerable” genotype had the shortest
TL when exposed to disadvantaged social environments
and the longest TL when exposed to advantaged
environments
Other G x E interactions
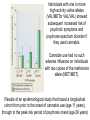
Evidence from studies around the world shows that cannabis use
is a statistical risk factor for the emergence of psychosis,
ranging from psychotic symptoms (such as hallucinations and
delusions) to clinically significant disorders.
However, most people who use cannabis do not develop
psychosis, which suggests that some individuals may be
genetically vulnerable to its effects.
Which is the genetic risk factor?
COMT gene polymorphism???
Individuals with one or more
high-activity valine alleles
(VAL/METor VAL/VAL) showed
subsequent increased risk of
psychotic symptoms and
psychosis-spectrum disorder if
they used cannabis.
Cannabis use had no such
adverse influence on individuals
with two copies of the methionine
allele (MET/MET).
Results of an epidemiological study that traced a longitudinal
cohort from prior to the onset of cannabis use (age 11 years),
through to the peak risk period of psychosis onset (age 26 years)
Cannabis affects in subjects with different genotype
Subjects were tested on two occasions, separated by 1 week, as
part of a double-blind, placebo controlled cross-over design.
In randomized order, they received either 0 g or 300 g -9tetrahydrocannabinol (the principal component of cannabis) per
kilogram bodyweight.
Cannabis affected cognition and state psychosis, but this
was conditional on COMT genotype.
Individuals carrying two
copies of the valine
allele exhibited more
cannabis-induced
memory and attention
impairments than
carriers of the
methionine allele, and
were the most sensitive
to cannabis-induced
psychotic experiences.
Vulnerabilità genetica verso la dipendenza da
sostanze: dati nell’uomo
In one experiment, the researchers investigated whether
a polymorphism in the D4 dopamine receptor gene
(DRD4) affected craving after priming doses and drug
cues.
Participants were tested on two occasions, randomly
assigned to receive three alcoholic drinks on the first
session and three control drinks on the second session,
or the reverse.
Individuals carrying the DRD4 long (L) allele reported
a stronger urge to drink in the alcohol condition than
in the placebo condition. By contrast, individuals with
two short DRD4 alleles (S) reported no differences in the
urge to drink between the two conditions.
These findings suggest that the DRD4
polymorphism moderates craving after
alcohol consumption, and indicate that
DRD4*L individuals may be more
susceptible to losing control over drinking
But the DRD4 polymorphism is not simply a genetic risk for alcohol abuse.
Individuals carrying the L allele also experience more craving and arousal after
exposure to tobacco smoking cues, whereas DRD4*S individuals do not.
This suggests that DRD4 may influence the incentive salience of
appetitive stimuli more generally, and offers a clue as to why
different addictive disorders tend to co-occur in the same
individuals.
Much genetic research has been guided by the
assumption that genes cause diseases, but the expectation
that direct paths will be found from gene to disease has not
proven fruitful for complex psychiatric disorders.
Findings of G x E interaction point to a different, evolutionary
model.
This model assumes that genetic variants maintained
at high prevalence in the population probably act to promote
organisms' resistance to environmental pathogens.
They extend the concept of environmental pathogens to
include traumatic, stressful life experiences and propose that the
effects of genes may be uncovered when such pathogens are
measured (in naturalistic studies) or manipulated (in experimental
studies).
Futura facenda
The characterization of subjects' genetic vulnerability
as opposed to their resilience needs to move beyond
single genetic polymorphisms.
New approaches will use information about biological
pathways to identify gene systems and study sets of
genetic polymorphisms that are active in the
pathophysiology of a disorder.
For example, in relation to depression, information
about the psychobiology of psycho-social stress can
be used as a first step to characterize a set of genes
that define a genotype that is vulnerable as opposed
to resilient to stressful life events.
Incorporating information about genetic pathways
into gene–environment interaction studies will
enhance explanatory power, but it will also
present unique statistical challenges related to
the use of data-mining tools and the pooling of
data across different studies.
If environmental risk exposure differs
between samples, candidate genes may
fail replication.
If environmental risk exposure differs
among participants within a sample,
genes may account for little variation in
the phenotype.
Hypothesis
Some multifactorial disorders, instead of
resulting from variations in many genes
of small effect, may result from
variations in fewer genes whose effects
are conditional on exposure to
environmental risks.
Necessity of implementing gene x
environment interaction research protocols
Necessity of accurate phenotyping
Interazioni geni ambiente nel Bucharest Early
Intervention Project
Fin qui 19 ottobre
Charles Nelson III
Cosa è il Bucharest Early Intervention project
Science, 2008
Modification of depression by COMT val158met polymorphism
in children exposed to early severe psychosocial deprivation.
Drury SS, Theall KP, Smyke AT, Keats BJ, Egger HL, Nelson CA,
Fox NA, Marshall PJ, Zeanah CH. (2010)
OBJECTIVE:
To examine the impact of the catechol-O-methyltransferase
(COMT) val(158)met allele on depressive symptoms in young
children exposed to early severe social deprivation as a result of
being raised in institutions.
METHODS:
One hundred thirty six children from the Bucharest Early Intervention Project
(BEIP) were randomized before 31 months of age to either care as usual (CAU)
in institutions or placement in newly created foster care (FCG). At 54 months of
age, a psychiatric assessment using the Preschool Age Psychiatric Assessment
(PAPA) was completed. DNA was collected and genotyped for the COMT
val(158)met polymorphism. Multivariate analysis examined the relationship
between COMT alleles and depressive symptoms.
RESULTS:
Mean level of depressive symptoms was lower among
participants with the met allele compared to those with two
copies of the val allele (P<0.05).
Controlling for group and gender, the rate of depressive symptoms
was significantly lower among participants with the met/met or the
met/val genotype [adjusted relative risk (aRR)=0.67, 95% CI=0.45,
0.99] compared to participants with the val/val genotype, indicating
an intermediate impact for heterozygotes consistent with the
biological impact of this polymorphism.
The impact of genotype within groups differed significantly.
There was a significant protective effect of the met allele on
depressive symptoms within the CAU group, however there
was no relationship seen within the FCG group.
.
CONCLUSIONS:
This is the first study to find evidence of a gene x
environment interaction in the setting of early social
deprivation.
These results support the hypothesis that individual
genetic differences may explain some of the variability in
recovery amongst children exposed to early severe
social deprivation