Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Perception, 1995, volume 24, pages 287-298
Cortical representation of visual three-dimensional
space
Yves Trotter
Centre de Recherche Cerveau et Cognition, CNRS, Faculte de Medecine de Rangueil, Universite
Paul Sabatier, 133 route de Narbonne, 31062 Toulouse Cedex, France
Based on paper presented at the Conference in Binocular Stereopsis and Optic Flow, Toronto,
Canada, 22-26 June 1993
Abstract. Perception of real depth includes information on stereopsis and distance. How both
interact in the visual pathway was the subject of a study performed on the behaving monkey.
Neurons in the primary visual cortex (area VI) have their activity, visual and/or spontaneous,
modulated by the viewing distance. Disparity selectivity may be present or better expressed at a
given viewing distance. This modulation is independent of the visual pattern. The use of prisms
shows that vergence is implicated in this phenomenon. Consequently, extraretinal signals
related to ocular motility have access to area VI. Among them, proprioceptive signals from the
eye muscles have been shown to be involved in visual cortical function and in the development
of depth perception. It is possible that the same signals may also be involved in the distance
modulation shown in VI neurons, but this remains to be examined. A possible specialisation of
disparity-selective cells in different cortical areas is discussed.
1 The problem
The visual system must extract several kinds of information from the spatial environment to perform any accurate motor act. For instance, to grasp an object we need to
evaluate its thickness, its position relative to other objects, and the distance separating
the object from us. There are different types of information available such as colour,
orientation, and motion cues, familiar knowledge of size, and all monocular cues,
including texture, shadows, perspective, overlapping etc, which may considerably help
in deciding the adequate motor strategy. However the most direct and powerful cues
are probably related to binocular-vision geometry.
Stereoscopic depth perception is related to binocular vision and by this process the
image of the viewed object is projected on both retinas in slightly different positions
because of the interocular distance. It allows determination of the relative position of
an object to another. In 1838 Wheatstone invented the mirror stereoscope and thus
demonstrated for the first time that stereopsis is based on a retinal-disparity cue.
Subsequently, Julesz (1960) used psychophysical experiments with random-dot stereograms to prove that a difference in the horizontal position of corresponding elements
of left and right images was sufficient to reconstruct depth relative to a fixation plane.
However stereopsis alone is not sufficient to obtain complete three-dimensional (3-D)
perception of space in our immediate environment. We also need to evaluate the
distance to the fixation plane. This is particularly important and critical in programming the motor act required to grasp objects or to navigate. The final motor
strategy, of course, depends on how well absolute distances are evaluated. Indeed,
kinematic analysis performed by Servos et al (1992) revealed that prehensile movements carried out under monocular viewing differ substantially from those performed
under binocular conditions. Under monocular viewing, distances of objects are
underestimated and grasping movements take longer movement times and exhibit
lower peak velocities, longer deceleration phases, and smaller grip apertures than
movements performed under binocular viewing. Removal of binocular vision in a
situation which is otherwise rich in depth cues has a profound effect on the spatio-
288
Y Trotter
temporal organisation of prehensile movements whether in their initial programming
or in their efficient on-line control (Servos and Goodale 1994). Correct evaluation of
distances, which allows skilful manipulation of objects in the immediate environment,
requires binocular vision, including stereoscopic information along with other sources
from vergence and/or accommodation and/or vertical disparity (Foley 1980; Bishop
1989; Cumming et al 1991; Rogers and Bradshaw 1993).
Relative cues involve perceptual interactions between two or more stimulus objects,
whereas absolute cues are interactions between the observer and the stimulus object
that contribute to the perceived characteristics of an object. The perceived distance
between objects produced by a relative cue such as retinal disparity refers to perceived
exocentric distance. The perceived distance from the egocentre of the observer
produced by an absolute cue such as convergence refers to perceived egocentric
distance (Gogel 1977). Relative distances of objects can be determined from the
relative disparity between the images projected on both retinas, independently of the
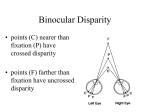
fixation point or eye movements. Disparity relative to the function plane refers to
absolute disparity (figure 1). This form of disparity (crossed or uncrossed) can be
used to evoke vergence movements (respectively convergence or divergence) that
bring a selected object into the fixation plane. In this case, the absolute distance of
the object can be directly calculated from the angle formed by the lines of sight or
angle of convergence. In contrast, absolute distance of an object located outside the
fixation plane requires a calculation that combines the angle of convergence with the
retinal disparity of the object (see reviews by Collewijn and Erkelens 1990; Collewijn
et al 1991).
It has long been proposed that oculomotor cues such as vergence and accommodation could be implicated in distance perception (see reviews by Ogle 1962a; Foley
1980). There has been controversy over the last century as to whether vergence is
implicated. Nevertheless, recent psychophysical experiments have provided evidence
that the visual system computes distance (at least for short distances of up to 2 m) by
taking into account the convergence parameter only, rather than that of accommodation or both (von Hofsten 1976; Ritter 1977; Foley 1980; Morrisson and Whiteside
1984; Collett et al 1991; Cumming et al 1991). Theoretically, there is another retinal
cue which may be involved in the calculation of true depth, ie vertical disparity
Relative disparity of A/B: a - b = 61- 62
Absolute disparity of A: a2 - ax = x - a
Absolute disparity of B: f$x - f$2 = x - b
Figure 1. Geometrical aspects of relative/absolute disparity (adapted from Blakemore 1969;
Collewijn and Erkelens 1990).
Cortical representation of 3-D space
289
(Mayhew 1982; Mayhew and Longuet-Higgins 1982; Bishop 1989). This could not
be verified in psychophysical experiments (Cumming et al 1991; Sobel and Collett
1991). However, in a subsequent study Rogers and Bradshaw (1993) reported that
vertical disparity can provide a cue to viewing distance, but only when the field of
view is sufficiently large.
When we look at different objects or parts of them located in different directions
and at different distances, we fixate them successively and therefore their retinal
representations are changed. However, the perceived position of the objects as well
as the spatial relations among them remain relatively unchanged. An object perceived
at different distances has the same aspect of thickness and shape. A book, for example,
appears as thick when viewed from 20 cm or 80 cm despite the fact that disparities
are smaller by a factor of 16. Theoretically, it should appear to flatten when moved
away from the observer because the size of a retinal image changes almost in inverse
proportion to the viewing distance, whereas retinal disparities change in inverse proportion to the square of the viewing distance (for reviews see Ogle 1962a, 1962b). To
achieve this depth-constancy phenomenon, wherein the depth being judged remains
constant while viewing distance varies, the decrease in retinal disparity as a function
of the viewing distance has to be internally compensated. This transformation process
requires egocentric (oculomotor) cues such as vergence and/or accommodation
(Wallach and Zuckerman 1963; Ritter 1977; for reviews see Gogel 1977; Ono and
Comerford 1977; Foley 1980).
2 Physiology
So far, nearly all studies on neural mechanisms of depth perception have concentrated
on stereopsis. But little is known concerning the neural mechanisms related to
binocular distance perception. Disparity-sensitive cells were first discovered in the
primary visual cortex of anaesthetised paralysed cat. Area 17 is the first cortical area
where inputs originating from each eye are integrated. Left and right receptive fields
of a binocular neuron may be in exact topographic correspondence in the two eyes or
may have slightly disparate locations. Consequently, two identical stimuli falling in
the same position in both receptive fields or in slightly different positions will
produce binocular facilitation, suppression, and/or summation (Barlow et al 1967;
Nikara et al 1968; Joshua and Bishop 1970; Ferster 1981). This property of disparity selectivity is generally considered to be the neural basis for stereopsis (for a
review see Poggio and Poggio 1984). In normal binocular vision conditions, distinct
categories of cells have been described in area VI of behaving monkeys: some preferentially respond to stimuli in front of the fixation plane ('near' cells) whereas others
prefer stimuli behind the fixation plane ('far' cells), or around the fixation plane
('tuned' cells) (Poggio and Fischer 1977). The existence of disparity-sensitive neurons
was more recently confirmed by means of random-dot stereograms, and the classification of disparity cells expanded (Poggio et al 1985, 1988; Trotter et al 1992).
It is thus essential to know how relative and absolute cues interact in the visual
pathway. We thus addressed this question in area V I of behaving monkeys (Trotter
etal 1992). Three predictions could be made. First, disparity sensitivity remains
unchanged regardless of fixation distance; this would be the neural basis of a pure
stereoscopic mechanism. Second, a given neuron may qualitatively change its disparity selectivity as a function of the distance. For example, a 'far'-type neuron for
close fixation would become a 'near' type for distant fixation. This would be a neural
basis for the absolute-depth mechanisms. Last, disparity selectivity could be
modulated by the fixation distance. This would be an intermediate step in coding
absolute depth. This third possibility was actually observed in area VI of the behaving
monkey.
290
Y Trotter
To test these predictions, a classical fixation task was used to present static
random-dot stereograms, generated through colour filters at six different horizontal
disparity values. T h e monkeys were trained to fixate at three distances (20 cm,
40 cm, or 80 cm). T h e same dot patterns subtending the same visual angle on the
retina and the same disparity values were projected at the three distances. Figure 2
shows a sample neuron which is sensitive to the viewing distance. It is a 'near' type at
20 cm but is poorly visually responsive at 40 cm and 80 cm distance. A second
example is shown in figure 3 where the neuron is almost silent at 40 cm but is clearly a
'tuned excitatory' type at 80 cm. In fact, most cells (more than 80%) have their visual
responses modulated by the viewing distance so that disparity sensitivity emerges or is
better expressed at a certain distance with no preference for any particular one, nor
for a particular 'near', 'far', or 'tuned' cell type. In addition, non-disparity-selective
cells can also show some modulation of responsiveness (about 15%).
We checked that the modulations of visual responsiveness were not artifactual,
resulting for instance from subtle changes in positions of the dots in the receptive
fields that might have occurred when the position of the video monitor was physically
changed. If this were the case, one would expect different kinds of modulations when
20 cm
80 cm
40 cm
80
- 60
J3 40
vr
20
S-.*-"*^
-0.3
0.0
0.3
0.6
-0.3 0.0 0.3 0.6
Disparity/deg
-0.3 0.0
- f t «
0.3
0.6
Figure 2. Effects of viewing distance on disparity selectivity in area VI of the behaving monkey.
Static random-dot stereograms were flashed for 300 ms on a colour video screen under constant
angular size conditions at two or three distances, 20 cm, 40 cm, and 80 cm. The whole stereogram appeared as a large flat surface (width 18 deg, height 14 deg) floating in front of or behind
a fixation point in the centre of the screen. Retinal-disparity-tuning curves (continuous lines)
and spontaneous activity (dotted curves) are shown at the three distances. Vertical bars indicate
standard errors. This cell shows a modulation of visual responsiveness with a visual responsiveness and a disparity selectivity present at a distance of 20 cm but not at 40 cm and 80 cm.
80
80 cm
40 cm
60
aa
40
20
0
-0.3 0.0
0.3
0.6
Disparity/deg
-0.3 0.0
0.3
0.6
Figure 3. Retinal-disparity-tuning curves (continuous lines) and spontaneous activity (dotted
lines) for distances of 40 cm and 80 cm. An example of 'tuned-excitatory' type cell at 80 cm; it
is almost not visually responsive at 40 cm.
291
Cortical representation of 3-D space
using two different dot patterns. For some cells, we tested disparity selectivity with
two random-dot-stereogram patterns, one with a 1 0 % dot density and the second with
a density of 20%, at different distances, and showed that the qualitative effect of
changing the distance was always constant. This indicates that the effect on visual
responsiveness has a nonretinal origin, as supported by the effects of fixation distance
on the level of spontaneous activity (figure 4). This cell, as many other cells, has a
level of spontaneous activity which increases as the distance becomes shorter. But as
observed for this cell, visual responsiveness and spontaneous activity modulations are
not necessarily related. This cell shows suppressed neural activity around 0.15 deg
disparity at both distances. However, the excitatory visual response is the highest
when the level of spontaneous activity is lowest, with the emergence of a clear
disparity selectivity of 'near' type. Consequently, modulation of visual responsiveness
does not appear to be dependent on neuron excitability level. Modulation of spontaneous activity appears to be of extraretinal origin as if it is related to muscular effort
at short distances.
20 cm
80 cm
40
30
20
10
-0.3
0.0
0.3
0.6
-0.3
0.0
0.3
0.6
Disparity/deg
Figure 4. Effect of viewing distance (20 cm or 80 cm) on visual responsiveness (solid curve) and
spontaneous activity (dotted curve). The spontaneous activity is higher by a factor of about
three at a short distance than at a longer distance. Meanwhile the visual reponsiveness with
clear disparity sensitivity is present at the longer distance, but not at the shorter one, where it is
suppressed around the plane of fixation.
20 cm
80 cm + 10 dioptres
80 cm
100
^
S
80
60
'% 40
20l
0
=*
-0.3 o!o
0.3
0.6
-0.3 0.0 0.3 0.6
Disparity/deg
o-tf
'Hi_
-0.3 0.0
0.3
0.6
Figure 5. Effect of vergence, produced by using wedge prisms (base out) on disparity sensitivity.
The cell is clearly sensitive to negative disparity values ('near' type) when the monkey fixates at
20 cm distance. This disparity sensitivity is much lower at 80 cm distance. At this distance, two
wedge prisms of 5 dioptres each were applied in front of each eye of the monkey. The power of
the prisms was chosen by calculation so as to reproduce an angle of vergence similar to that
occuring at 20 cm viewing distance in this animal. The visual responsiveness and disparity
sensitivity thus reappear and are similar to that at 20 cm distance. The higher variability of
visual response is due to adaptation to prisms.
292
Y Trotter
These neural-activity modulations of cells in area VI may originate from vergence
and/or accommodation. To investigate a possible effect of vergence, wedge prisms
were used to produce an angle of vergence similar to that normally present at a
certain fixation distance. If vergence angle is responsible for the distance effect, the
use of prisms should reproduce the same effects as provoked by changing the distance
of fixation. Figure 5 shows an example where the cell is not very disparity sensitive at
80 cm, but is clearly disparity selective at 20 cm fixation. However when prisms are
added at 80 cm to reproduce an angle of convergence similar to that normally found
at a distance of 20 cm in this monkey, the disparity sensitivity becomes similar to that
occurring at this particular distance. Study of other cells revealed that when the
distance effect was present, it could be reproduced by using prisms. Sustained
vergence activity thus appears to modulate disparity sensitivity in area V I .
3 Extraretinal sources
Modulation of the visual response and spontaneous activity by vergence indicates that
an extraretinal source related to ocular motility has access to the visual pathway,
directly in the visual cortex or even at the level of the dorsolateral geniculate nucleus
(LGNd). It should be pointed out that spontaneous-activity modulation of about one
third of the LGNd cells has been obtained by manipulating the vergence angle with
prisms in the awake monkey (Richards 1968). Modulation at the cortical level may
therefore result from extraretinal influences as early as the LGNd. What are their
possible origins? The two standard possible sources are efference copy and extraocular muscle proprioception (EMP).
Indeed, the latter source was shown to be strongly implicated during postnatal
development of depth perception in cat. Eye muscles in this species, as opposed to
human primates and most nonhuman primates, do not contain classical spindles, but
rather contain palisade endings, located at the musculotendinous junction, which play
the same role (Alvarado-Mallart and Pingon-Raymond 1979). Moreover, responses
to eye-muscle stretching have been shown in numerous parts of the brain including
the primary visual cortex (Buisseret and Maffei 1977; see a review by Steinbach
1987). Most proprioceptive afferents coming from extraocular muscles run through
the ophthalmic branch of the trigeminal nerve (Vj nerve) in the cranial part (Batini
and Buisseret 1974; Batini et al 1975), so one way to eliminate them is to cut this
nerve at the entrance to the Gasserian ganglion. By means of this paradigm, it was
shown that an imbalance of proprioceptive inputs originating from extraocular
muscles was responsible for disruption of cortical binocularity in the primary visual
cortex during a critical period of postnatal development (Trotter et al 1987). This
deficit was accompanied by strong impairment of binocular depth perception, which
remained permanently throughout the life of the animal (Graves et al 1987; Trotter
et al 1991). Physiological studies on the same animals revealed a drastic loss of
disparity sensitivity in the primary visual cortex, which was mainly due to a reduction
of binocular suppression (but not of binocular facilitation) combined with a specific
increase in the variability of the binocular responses (but not of monocular responses)
(Trotter et al 1993). During growth, there must be constant adjustment between
sensory-input maps and motor-output maps to maintain spatial constancy. During
postnatal development, for instance, the interocular distance changes and thus the
amount of eye convergence required to fixate an object at a certain distance will
increase. Clearly, the visual system must account for these progressive peripheral
constraints to allow constant depth perception (see von Hofsten 1977; Ritter 1977).
Therefore, there must be a continuous process of calibration to specify veridical
Cortical representation of 3-D space
293
properties of space. The visual system probably uses balanced EMP signals as reference
signals for interocular alignment during the development of orbital tissues to calibrate
binocular disparities.
EMP is fundamental to the development of depth perception, but its role is still
unclear in the adult cat (Fiorentini et al 1985, 1986; Graves et al 1987; Trotter et al
1991). This may be due to variations in magnitude and often a lack of vergence
movements to near objects in this species (Hughes 1972). Conversely in primates,
which have highly precise control of vergence movements, EMP deafferentations
severely impair vergence tasks and maintenance of binocular alignment during binocular or monocular target fixation (Guthrie et al 1982). This indicates a functional role
of EMP in controlling ocular vergence movements in the adult. This has been
corroborated by experiments performed in human subjects which showed that passive
monocular deviation of one eye results in phoria changes, suggesting that EMP may
be used at some central level to ensure static control of eye-axis alignment and movement conjugacy (Gauthier and Vercher 1992). Clinical observations also strongly
support an important role of EMP in the etiology of strabismus and in the efficacy of
its surgical correction (Lennerstrand et al 1992; Steinbach 1992). If EMP plays a
significant functional role in oculomotility in the adult, it is probably in the control of
eye fixation alignment, vergence control, adaptation mechanisms, and perhaps in
pursuit eye movements, ie in all oculomotor controls directly implicated in 3-D depth
perception that require slow but fine adjustment of images on both retinas. EMP also
probably plays little or no role in the control of saccadic eye movements (Guthrie
et al 1983; Gauthier and Vercher 1992). It is likely that in combination with other
signals such as corollary discharge it contributes to localisation of objects in space
(Gauthier et al 1990; Bridgeman and Stark 1991).
4 A cortical specialisation of disparity-selective cells?
What is the meaning of modulation of disparity selectivity by the distance of fixation
as early as cortical area V I ? First, according to the predictions made earlier, one can
conclude that area VI is not the site of a pure neural stereoscopic mechanism, but
rather a step for a neural 3-D distance mechanism including stereopsis. Vergence
appears to control disparity sensitivity at the cortical level. Moreover, models have
been developed to represent distance by means of disparity-selective neurons whose
gain is modulated by vergence angle (Lehky et al 1990), suggesting that the egocentric
distance of objects is encoded according to a distributed representation (Pouget and
Sejnowski 1994). Generally, disparity is considered to be the stimulus which controls
vergence to bring or keep the binocular retinal stimuli within the fusional area
(Rashbass and Westheimer 1961; Westheimer and Mitchell 1969; for review see
Collewijn and Erkelens 1990). Vergence has rarely been proposed to control disparity, even though psychophysical studies have shown that binocular vergence
influences the choice of stereo matches in an ambiguous display, with matches in the
fixation plane being strongly preferred over matches with large disparities (McKee
and Mitchison 1988; Mallot and Bideau 1990). The matching process between the
two eyes is a critical problem in stereopsis from a computational point of view (Marr
and Poggio 1979). An algorithm was developed more recently which shows that
vergence can help provide the necessary constraints for the matching process (Geiger
and Yuille 1989).
It is thus possible that vergence may control the neural matching process at a
particular viewing distance by gating finely 'tuned' disparity-sensitive cells. Indeed,
disparity-selective cells, considered to be the neural basis for stereopsis, occur in
the monkey brain from the primary visual cortex to at least the lateral intraparietal
(LiP) area in the parietal cortex. However, their presence varies along this pathway.
294
Y Trotter
If we consider 'tuned excitatory' cells, those that respond in the plane of fixation at 0 deg
are particularly common in area VI but they decrease in proportion in area V2 and
they represent only 10% in areas V 3 - V 3 A where 'tuned near' and 'tuned far' (shifted
from 0 deg) predominate (Poggio et al 1988) (figure 6a). 'Tuned' cells also appear to
be present only in early steps of visual processing since they appear to be almost nonexistent beyond area MT (figure 6b). So it is possible that 'tuned vergence-gated' cells
are part of the neural mechanism that favours the stereo matching process at an early
step of the visual pathway to permit binocular single vision at a fixed viewing distance.
90
100
80
701
S 60
501
40
MT MST LiP
VI V2
V2
V3
fu\
Cortical area
Cortical area
Figure 6. Proportion of 'tuned' cells with peak tunings > 0 . 1 deg (which represent 'tuned near'
and 'tuned far' cells) as a function of the cortical area (adapted from Poggio et al 1988). Note
that they are much more represented in area V 3 than in area V I where 'tuned 0 deg' predominate, (b) Average percentage of 'tuned', 'near', and 'far' cells in different cortical areas (adapted
from Maunsell and van Essen 1983; Poggio et al 1988; Gnadt and Mays 1 9 9 1 ; Roy et al 1992;
Trotter et al 1992). It can be seen that 'tuned' cells are present only in the early processing of
visual 3-D information.
VI
VI V 2 V 3 - V 3 A
MT
MST
LIP
coarsely tuned.
" near/far
• fine
near/far/tuned
binocular
matching
*
motor control for
vergence initiation
direction
selectivity
fine 3-D
localisation
I
3-D motion
3-D direction of
self-motion
proprioceptive
vergence signal?
Figure 7. Scheme for a possible cortical specialisation of disparity cells in 3-D oculomotor
control and binocular matching. Disparity-sensitive cells may subserve different functions
depending on their quantitative and qualitative cortical representation. Fine-tuned cells are
found only at an early step of visual processing. 'Near' and 'far' cells beyond V 3 - V 3 A cortical
areas are coarsely tuned. In M T and MST areas, they may subserve motion and direction of
self-motion. In LiP area, they may initiate vergence movements to bring binocular images of
objects in correspondence under a feedback oculomotor-signal control. Tuned-disparity cells in
V I - V 2 - V 3 - V 3 A may subserve binocular matching with the help of a vergence signal, possibly
proprioceptive, to provide the fine localisation of objects in real depth.
Cortical representation of 3-D space
295
'Near' and 'far' cells may subserve several functions. They constitute about 50% of
disparity-selective cells in each cortical area, from V I to MT, and their proportion
increases beyond these areas: there are about 90% in MST area (Roy et al 1992) and
100% in LiP (Gnadt and Mays 1991) (figure 6b). 'Near' and 'far' cells, whose
functions are also gated by the vergence angle in areas VI and V2 (unpublished
observation) and perhaps in V 3 - V 3 A , may be neural substrates for the 3-D static
localisation of objects in depth at various distances during fixation and may also
participate in the neural matching process along with 'tuned' cells. Beyond these
cortical areas, 'near' and 'far' cells may subserve other functions. In MT and MST
areas, they may play a role in signalling motion in depth and also the direction of selfmotion of the observer (3-D dynamic detection) through the environment according
to the interdependence between disparity selectivity and direction selectivity, as
shown in the MST area, where neurons are much less frequently influenced by
vergence than in the earlier visual cortical processing (Roy et al 1992). Another role
of 'near' and 'far' cells in LiP, which is connected to MT and MST areas, would be to
initiate vergence movements (vergence initiation) to bring the object into the plane of
fixation (figure 7). Indeed, their activity is modulated by vergence (Gnadt and Mays
1991) and neurons in that area project to the superior colliculus, a region which is
usually assigned an oculomotor-control function (Sparks 1986). However, studies to
investigate a possible role of the colliculus in vergence control would now be required.
5 General conclusions
We are far from having a complete understanding of how the brain computes visual
3-D information and transforms it into an appropriate motor act in space. Vision and
action cannot be dissociated in this question. Early in the visual pathway, in area V I ,
retinal signals interact with oculomotor signals to process real depth. Neural coding
of retinal disparity is under control of the viewing distance and vergence appears to
be a determining cue. The extraretinal signals implicated in visual responsiveness and
spontaneous-activity modulations remain to be determined. Among them, EMP is a
possible candidate involved in this process because it was shown to be strongly
involved in development of depth perception and also in the underlying neural
mechanism. Such an extraretinal signal may be used to scale disparities both during
postnatal development and in the adult to yield space and depth constancies.
There is still much to know on how viewing distance influences disparity coding in
other cortical areas, particularly in V 3 - V 3 A , MT, PO, 7a, and beyond. So far, we
know that neurons in some of them are disparity selective [V3-V3A (Poggio et al
1988), MT (Maunsell and van Essen 1983)] or are sensitive to the angle of gaze in the
two-dimensional domain [V3A (Galletti and Bataglini 1989), PO (Galletti et al 1991),
7a (Andersen and Mountcastle 1983; Andersen et al 1990)]. This indicates that all of
these areas should be explored systematically in terms of disparity coding, eye position, and vergence to obtain further knowledge on the way 3-D information proceeds
and then is transformed in a motor act (such as ocular or arm movement).
Acknowledgements. I would like to thank Dr S Celebrini and Professor M Imbert for helpful
comments on the manuscript. I am also grateful to my colleagues who collaborated on the
experimental work. It was supported by grants from the Centre National de la Recherche Scientifique (CNRS) and the European Community, Esprit Basic Research Action (MUCOM).
References
Alvarado-Mallart R M, Pincon-Raymond M, 1979 "The palisade endings of cat extraocular
muscles: a light and electron microscope study" Tissue and Cell 1 1 5 6 7 - 5 8 4
Andersen R A, Mountcastle V B, 1983 "The influence of the angle of gaze upon the excitability
of the light-sensitive neurons of the posterior parietal cortex" Journal of Neuroscience 3
532-548
296
Y Trotter
Andersen R A, Bracewell R M, Barash S, Gnadt J W, FogassiL, 1990 "Eye position effects on
visual, memory and saccade-related activity in areas LIP and 7A of macaque" Journal of
Neuroscience 101176-1196
Barlow H B, Blakemore C, Pettigrew J D, 1967 "The neural mechanism of binocular depth
discrimination" Journal of Physiology (London) 193 321-342
BatiniC, Buisseret P, 1974 "Sensory peripheral pathway from extrinsic eye muscles" Archives
Italiennes de Biologie 112 18-32
BatiniC, Buisseret P, Buisseret-Delmas C, 1975 "Trigeminal pathway of the extrinsic eye
afferents in cat" Brain Research 85 74 - 78
Bishop P O, 1989 "Vertical disparity, egocentric distance and stereoscopic depth constancy: a
new interpretation" Proceedings of the Royal Society ofLondon, Series B 237 445 - 46 9
Blakemore C, 1969 "Binocular depth discrimination and the nasotemporal division" Journal of
Physiology (London) 205 471-497
Bridgeman B, Stark L, 1991 "Ocular proprioception and efference copy in registering visual
direction" Vision Research 3 1 1 9 0 3 - 1 9 1 3
Buisseret P, Maffei L, 1977 "Extraocular proprioceptive projections to the visual cortex" Experimental Brain Research 28 421-425
Collett T S, Schwartz U, Sobel E, 1991 "The interaction of oculomotor cues and stimulus size
in stereoscopic depth constancy" Perception 207 r 33 -754
CollewijnH, Erkelens C J, 1990 "Binocular eye movements and the perception of depth", in
Eye Movements and Their Role in Visual and Cognitive Processes Ed. E Kowler (Amsterdam:
Elsevier)pp 2 1 3 - 2 6 1
CollewijnH, Steinman R M, Erkelens C J, Regan D, 1991 "Binocular fusion, stereopsis and
stereoacuity with a moving head" in Binocular Vision Ed. D Regan (Boca Raton: CRC)
pp.121-136
Cumming B G, Johnston E B, Parker A J, 1991 "Vertical disparities and perception in threedimensional shape" Nature (London) 349 4 1 1 - 4 1 3
Ferster D A, 1981 "A comparison of binocular depth mechanisms in area 17 and 18 of the cat
visual cortex" Journal ofPhysiology (London) 3 1 1 6 2 3 - 6 5 5
Fiorentini A, Maffei L, Cenni M C, Tacchi A, 1985 "Deafferentation of oculomotor proprioception affects depth discrimination in adult cats" Experimental Brain Research 59 2 8 6 - 3 0 1
Fiorentini A, Cenni M C, Maffei L, 1986 "Impairment of stereoacuity in cats with oculomotor
proprioceptive deafferentation" Experimental Brain Research 63 364-368
Foley J M, 1980 "Binocular distance perception" Psychological Review 87 411 - 434
Galletti C, Battaglini P P, 1989 "Gaze-dependent visual neurons in area V3A of monkey
prestriate cortex" Journal of Neuroscience 9 1 1 1 2 - 1 1 2 5
Galletti C, Battaglini P P, Fattori P, 1991 "Functional properties of neurons in the anterior bank
of the parieto-occipital sulcus of the macaque monkey" European Journal of Neuroscience 3
452-461
GauthierGM, Vercher J L, 1992 "Ocular muscle proprioception and ocular motor control", in
Muscle Afferents and Spinal Control of Movement IBRO series, Eds L Jami, E PierrotDeseilligny, D Zytnicki (Oxford: PergamonPress)pp 277-285
GauthierGM, Nommay D, Vercher J L, 1990 "The role of ocular muscle proprioception in
visual localization of targets" Science 249 5 8 - 6 1
Geiger D, Yuille A, 1989 "Stereo and eye movement" Biological Cybernetics 6 2 1 1 7 - 1 2 8
Gnadt J W, Mays L E, 1991 "Depth-tuning in area LIP by disparity and accommodative cues"
Society Neuroscience Abstract 1 7 4 4 3 - 5 1 1
Gogel W, 1977 "The metric of visual space", in Stability and Constancy in Visual Perception:
Mechanisms and Processes Ed. W Epstein (New York: John Wiley) pp 129 - 1 8 1
Graves A L, Trotter Y, Fregnac Y, 1987 "Role of extraocular muscle proprioception in the
development of depth perception in cats" Journal of Neurophysiology 58 816-831
Guthrie B L , Porter J D, Sparks D L, 1982 "Role of extraocular muscle proprioception in eye
movements studied by chronic deafferentation of intra-orbital structures" Society Neuroscience
Abstract^ 156
Guthrie B L, Porter J D, Sparks D L, 1983 "Corollary discharge provides accurate eye position
information to the oculomotor system" Science 221 1193-1195
HofstenCvon, 1976 "The role of convergence in visual space perception" Vision Research 16
193-198
Hofsten C von, 1977 "Binocular convergence as a determinant of reaching behavior in infancy"
Perception 6 139-144
Hughes A, 1972"Vergenceinthecat" Vision Research 12 1961-1994
Cortical representation of 3-D space
297
Joshua D E, Bishop P O, 1970 "Binocular single vision and depth discrimination. Receptive
field disparities for central and peripheral vision and binocular interaction on peripheral
units in cat striate cortex" Experimental Brain Research 1 0 3 8 9 - 4 1 6
Julesz B, 1960 "Binocular depth perception of computer-generated patterns" Bell Systems
Technology Journal 39 1125-1162
Lehky S R, Pouget A, Sejnowski T J, 1990 "Neural models of binocular depth perception" Cold
Spring Harbor Symposium on Quantitative Biology 5 5 7 6 5 - 7 7 7
Lennerstrand G, Tian S, Han Y, 1992 "Eye muscle proprioception in strabismus and related
disorders: sensory and motor aspects", in Muscle Afferents and Spinal Control of Movement
IBRO series, Eds L Jami, E Pierrot-Deseilligny, D Zytnicki (Oxford: Pergamon Press)
pp 225-237
McKee S P, Mitchison G J, 1988 "The role of retinal correspondence in stereoscopic matching"
Vision Research 9 1 0 0 1 - 1 0 1 2
Mallot H A, Bideau H, 1990 "Binocular vergence influences the assignment of stereo
correspondences" Vision Research 30 1521 -1523
Marr D, Poggio T, 1979 "A theory of human stereo vision" Proceedings of the Royal Society of
London, Series B 204 301 - 328
Maunsell H R, Essen D C van, 1983 "Functional properties of neurons in middle temporal
visual area of the macaque monkey. II. Binocular interactions and sensitivity to binocular
disparity" Journal of Neurophysiology 49 1148-1167
Mayhew J, 1982 "The interpretation of stereo-disparity information: the computation of surface
orientation and depth" Perception 11 3 8 7 - 4 0 3
Mayhew J, Longuet-Higgins H C, 1982 "A computational model of binocular depth perception"
Nature (London) 297 376 - 378
Morrison J D, Whiteside T C D, 1984 "Binocular cues in the perception of distance of a point
of source of light" Perception 13555-566
Nikara T, Bishop P O, Pettigrew J D, 1968 "Analysis of retinal correspondence by studying
receptive fields of binocular signal units in cat striate cortex" Experimental Brain Research 6
353-372
Ogle K N, 1962a "Perception of distance and of size", in The Eye, Visual Optics and the Optical
Space Sense Ed. H Davson (New York: Academic Press) pp 247 - 269
OgleKN, 1962b "Spatial localization through binocular vision", in The Eye, Visual Optics and
the Optical Space Sense Ed. H Davson (New York: Academic Press) pp 271 - 324
Ono H, Comerford J, 1977 "Stereoscopic depth constancy", in Stability and Constancy in Visual
Perception: Mechanisms and Processes Ed. W Epstein (New York: John Wiley) pp 9 1 - 1 2 8
Poggio G F , Fischer B, 1977 "Binocular interaction and depth sensitivity in striate and prestriate cortex of behaving rhesus monkey" Journal of Neurophysiology 40 1392-1405
Poggio G F, Poggio T, 1984 "The analysis of stereopsis" Annual Review of Neurosciences 1
379-412
Poggio GF, Gonzales F, Krause F, 1988 "Stereoscopic mechanisms in monkey visual cortex:
Binocular correlation and disparity selectivity" Journal of Neuroscience 8 4531 -4550
Poggio G F , Motter B C, Squatrito S, Trotter Y, 1985 "Responses of neurons in visual cortex
(VI and V2) of the alert macaque to dynamic random-dot stereograms" Vision Research 25
397-406
Pouget A, Sejnowski T J, 1994 "A neural model of the cortical representation of egocentric
distance" Cerebral Cortex 4 314-329
Rashbass C, Westheimer G, 1961 "Disjunctive eye movements" Journal of Physiology (London)
159 339-360
Richards W, 1968 "Spatial remapping in the primate visual system" Kybernetik 4 146 -156
Ritter M, 1977 "Effect of disparity and viewing distance on perceived depth" Perception &
Psychophysics 22 400 - 407
Rogers B J, Bradshaw M F, 1993 "Vertical disparities, differential perspective and binocular
stereopsis" Nature (London) 361 253-255
Roy J P, Komatsu H, Wurtz R, 1992 "Disparity sensitivity of neurons in monkey extrastriate
area MST" Journal of Neuroscience 12 2478 - 2492
Servos P, Goodale M A, 1994 "Binocular vision and the on-line control of human prehension"
Experimental Brain Research 9 8 1 1 9 - 1 2 7
Servos P, Goodale M A, Jakobson L S, 1992 "The role of binocular vision in prehension: a
kinematic analysis" Vision Research 3 2 1 5 1 3 - 1 5 2 1
Sobel E C, Collett T S, 1991 "Does vertical disparity scale the perception of stereoscopic
depth?" Proceedings of the Royal Society of London, Series B 244 8 7 - 9 0
298
Y Trotter
Sparks D L, 1986 "Translation of sensory signals into commands for control of saccadic eye
movements: role of primate superior colliculus" Physiological Review 66 118-171
Steinbach M J, 1987 "Proprioceptive knowledge of eye position" Vision Research 27 1737' -1744
SteinbachM J, 1992 "The need for eye muscle proprioception", in Muscle Afferents and Spinal
Control of Movement IBRO series, Eds L Jami, E Pierrot-Deseilligny, D Zytnicki (Oxford:
Pergamon Press) pp 239 - 244
Trotter Y, Beaux J C, Pouget A, Imbert M, 1991 "Temporal limits of the susceptibility of depth
perception to proprioceptive deafferentations of extraocular muscles" Developmental Brain
Research 59 2 3 - 2 9
Trotter Y, Celebrini C, Beaux J C, GrandjeanB, Imbert M, 1993 "Long term dysfunctions of
neural stereoscopic mechanisms following unilateral extraocular muscle proprioceptive
deafferentation" Journal of Neurophysiology 69 1513-1529
Trotter Y, Celebrini C, Stricanne B, Thorpe S, Imbert M, 1992 "Modulation of neural stereoscopic processing in primate area VI by the viewing distance" Science 257 1279 -1281
Trotter Y, Fregnac Y, Buisseret P, 1987 "The period of susceptibility of visual cortical binocularity to unilateral proprioceptive deafferentation of extraocular muscles" Journal of
Neurophysiology 58 795-815
Wallach H, Zuckerman C, 1963 "The constancy of stereoscopic depth" American Journal of
Psychology 76 404 - 412
Westheimer G, Mitchell D E, 1969 "The sensory stimulus for disjunctive eye movements" Vision
Research 9 749-755
Wheatstone C, 1838 "Contributions to the physiology of vision. Part the first. On some
remarkable and hitherto unobserved phenomena of binocular vision" Philosophical Transactions of the Royal Society of London 128 371 - 394
p
© 1995 a Pion publication printed in Great Britain