Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
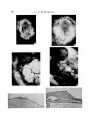
/ . Embryol. exp. Morph. Vol. 25, l,pp. 85-96, 1971 Printed in Great Britain g5 The origin and movement of the limb-bud epithelium and mesenchyme in the chick embryo as determined by radioautographic mapping By GLENN C. ROSENQUIST 1 From the Department of Pediatrics, The Johns Hopkins Hospital SUMMARY The origin of the limb-bud cells was determined by tracing the movements of [3H]thymidinelabelled grafts excised from late medium-streak to 5-somite stage chick embryos and transplanted to the epiblast, streak, and endoderm-mesoderm of similarly staged recipient embryos. Although exact definition of the prelimb areas was not possible because of the small number of grafts placed at each developmental stage, the study showed in general that at the late medium-streak stage the future limb-bud epithelium is in the epiblast (dorsal) layer near the lateral margin of the area pellucida. It moves medially toward the embryonic axis, just lateral to the premesoderm cells which will be invaginated at the primitive streak. With regression of the streak, the limb-bud epithelium moves relatively anteriorly into a position dorsal to the limb-bud mesoderm, beginning at least as early as the early head-fold stage. At the definitive-streak stage, the future limb-bud mesoderm is in the epiblast layer about halfway from the streak to the lateral margin of the area pellucida, at a level about halfway between the anterior and posterior ends of the streak. From this position the prelimb mesoderm migrates medially to the streak, and is invaginated into the mesoderm layer at a position about halfway between the anterior and posterior ends of the streak; after the head-process stage, it migrates anteriorly and laterally into the somatic layer of the lateral plate, ventral to the limb-bud epithelium. Mesoderm which will form the anterior limb-bud migrates anterior to mesoderm which will form the posterior limb-bud; mesoderm which will form the ventral portion of each limb-bud migrates posterolateral to mesoderm which will form the dorsal portion of each limb-bud. INTRODUCTION The limb-buds of the chick embryo begin to form at about the 30-somite stage, when the somatic mesoderm along the right and left flanks begins to accumulate into mounds of mesenchyme, covered by a layer of epidermal ectoderm. Although previous investigators (Wolff, 1936; Rudnick, 1945; Chaube, 1959) have identified areas of limb-bud-forming tissue in the right and left flanks of embryos prior to the 30-somite stage, the position of this tissue prior to the head-fold stage has not been investigated. 1 Author's address: Department of Pediatrics, The Johns Hopkins Hospital, Baltimore, Maryland 21205, U.S.A. 86 G. C. ROSENQUIST Using radioautographic analysis the present investigation traces the movement of [3H]thymidine-labelled transplants from their original positions in the epiblast, streak and endoderm-mesoderm layer of recipient embryos into the limbbuds of these embryos. Although the small number of grafts placed prevents exact definition of the prelimb areas, these regions are more precisely defined than they were previously, and the mesoderm and ectoderm layers are treated separately. MATERIALS AND METHODS The methods of preparation of recipient and [3H]thymidine-labelled donor embryos and of transplantation and radioautographic analysis of the grafts were identical to those described in previous publications (Rosenquist, 1966, 19706) and the description will not be repeated here. The late medium-streak to head-process stages of both donor and host embryos have been described previously (Rosenquist, 1970a) and are shown diagrammatically in Fig. 1A-C. Early head-fold (EHF) stage embryos had an elongated head process and a depression at the site of the future head fold, which had not as yet formed a pocket (Fig. 1D). Host embryos fixed prior to the 7-somite stage were normal when examined with the naked eye or microscopically. In those host embryos which survived to the 26-somite to early limb-bud stage the heart was beating, but extra-embryonic circulation had ceased and the development of the embryos was retarded compared to that of similar embryos incubated in ovo. The number of pairs of somites could not always be determined accurately (Fig. 2B, E). In the older host embryos which had not as yet developed limb-buds, the anterior and posterior limb regions, and their dorsal and ventral portions, were designated in cross-sections on the basis of their distance from the anterior intestinal portal, primitive streak, midline axis and nephrotome in comparison with embryos which had already developed limb-buds. This stage is referred to as the 26-30-somite stage in the Table and text, and is equivalent to stage 1 (Hamburger, 1938) and stage 15 (Hamburger & Hamilton, 1951). Host embryos which had developed limb-buds (stages 2-3 of Hamburger, 1938; stages 16-17 of Hamburger & Hamilton, 1951) were said to be at the early limb-bud (ELB) stage. In these embryos the portion of the ectoderm and somatic mesoderm lateral to the crest of the ectodermal cone is referred to in the text and Table as the future ventral portion of the limb, while the portion of the ectoderm and somatic mesoderm medial to the ectodermal cone is called the future dorsal portion of the limb. Although a part of each of the labelled transplants lay in the limb-budforming region of its host embryo, the number of embryos investigated was relatively small, and each transplant contained cells other than those destined for the limb-buds (as indicated in Table 1). Therefore the positions of the transplants at each stage in Fig. 1 suggest the location of the limb-bud cells at that stage, but do not define it precisely. Origin of limb-bud 87 The mapping of the limb-bud regions is based upon the following assumptions: (1) that since previous studies have established the general position of the premesoderm and pre-ectoderm cells in the epiblast layer of the chick blastoderm without mapping every part of that layer at each stage (Rosenquist, 1966), a small number of transplants carefully placed can demonstrate the position of more specific portions of the mesoderm and ectoderm, such as the limb-buds. (2) That similar graft positions in different embryos of the same stage are homologous even if the embryos were incubated for different lengths of time, and that the migration pathways followed by more than one accurately placed graft can be combined to follow movements of a group of cells through several overlapping stages of development. (3) That maps of presumptive organ-forming regions of the embryo are valid even if structures in the recipient embryos other than the organ to be mapped contain labelled cells. Throughout the text and figures, an asterisk (*) after the embryo number indicates that the position shown is that of the graft after its migration in the host embryo. RESULTS Limb-bud epithelium At the late medium-streak stage the cells which would form the epithelium of the limb-buds were in the epiblast layer at the lateral edge of the area pellucida, lateral to the primitive streak; in some cases they may have been on or outside the boundary between the area pellucida and the area opaca (embryos 1-5, Table 1, Fig. 1A). At the early head-fold stage, the cells which would form the epithelium of the anterior limb-bud were in the epiblast layer near the anterior half of the streak, about halfway between the streak and the lateral margin of the area pellucida (embryos 6, 7, Table 1, Fig. ID). By the 4-7-somite stage, the transplant in embryo 6* had migrated anteriorly and laterally to a position between the anterior end of the streak and the somite region (Table 1, Fig. 1E). If embryo 6 had been allowed to develop to the limbbud stage, labelled cells would have migrated into the anterior limb-bud, as did those in embryo 7* (Fig. ID, G). Therefore, the transplants in embryos l*-4*, which had reached positions similar to that of the graft in embryo 6* (Fig. 1E), also were considered to be destined for the anterior limb-bud. The transplant in embryo 5* and the posterior end of the transplant in embryo 3* were posterior to the transplants in embryos l*-4* and 6* at this stage; although there is no evidence from the present study that they would have contributed to the posterior limb-bud, they lay in the presumptive posterior limb-bud region as it was defined by Rudnick (1945) and Chaube (1959) (Fig. 3C), and were therefore considered to be destined for the epithelium of the posterior limb-bud. At the early limb-bud stage (equivalent to stages 2-3 of Hamburger, 1938 and 16-17 of Hamburger & Hamilton, 1951), labelled cells from the transplant in 88 G. C. ROSENQUIST embryo 7* had formed portions of the epithelium of the right anterior limb-bud, extending along the dorsal side of the ectodermal cone; other labelled cells from this transplant contributed to the epithelial layer of the embryo between the anterior limb-bud and the neural crest, but not to the ectoderm between the anterior and posterior limb-buds, nor to the posterior limb-bud (Table 1, Figs. 1G, 2). Table 1. Position of labelled cells in recipient embryos carrying tritiated thymidine-labelled grafts* Embryo no. Incubated f (h) Stage fixed Position of labelled cells 1 LMS 2 3 LMS LMS 22 23 5S 7S Ec: ALBJ Ec: ALBJ 22 5S Ec: ALBJ 4 5 6 7 LMS LMS EHF EHF DS HP HP EHF EHF EHF EHF EHF EHF EHF EHF EHF EHF 4S 4S 5S 15 19 7 66 5 4 10 48 108 66 108 68 66 68 68 48 92 56 66 60 6S Ec: ALBJ Ec: PLBJ Ec: ALBJ Ec: ALB; Px M St M: PLBJ LP M: ALBJ, PLBJ; YS En LP M: ALBJ, PLBJ; YS En LPM: ALBJ (dv); YSEn LPM: ALBJ(v); YSEn LPM: ALBJ (d); YSEn LP M: ALBJ (dv), PLBJ (dv); YS En LP M: ALB (v), PLB (v); YS En LP M: ALBJ (d), PLBJ (d); YS En LP M: ALBJ (d), PLBJ (d); YS En LP M: ALB (dv), PLB (dv); YS En LP M: ALBJ (d), PLBJ (d); YS En LP M: ALB (v), PLB (v); YS En LPM: ALBJ (dv); YSEn LPM: ALB (v); YSEn Px, LPM: PLB (d); YSM 8 / Stage grafted 9 110 11 12 13 14 I115 16 -17 Us19 20 21 22 23 6S 7S ELB EHF EHF 4S 26-30S 26-30S 26-30S 26-30S ELB 26-30S 26-30S ELB 26-30S ELB 26-30S ELB ELB * Key to abbreviations: ALB, anterior limb-bud; d, dorsal; DS, definitive-streak stage; Ec, ectoderm; EHF, early head-fold stage; ELB, early limb-bud stage; En, endoderm; HP, head-process stage; LMS, late medium-streak stage; LP, lateral plate; M, mesoderm; Px, paraxial; PLB, posterior limb-bud; S, somite; St, streak; v, ventral; YS, yolk sac. t Hours of incubation after placement of the graft. J Presumptive limb-bud region. Limb-bud mesoderm At the definitive streak stage, the cells which would form the mesoderm of the limb-buds were in the epiblast layer on each side of the primitive streak, at a level about halfway between the anterior and posterior ends of the streak, and about halfway from the streak to the lateral margin of the area pellucida. The transplant in embryo 8 (Table 1, Fig. IB) was within the area destined to be Origin of limb-bud 89 invaginated at the streak, which was identified in a previous mapping study (Rosenquist, 1966). By the head-process stage the mesoderm which would form portions of the limb-buds had been invaginated at the primitive streak and had migrated into the lateral plate mesoderm on each side of the streak, at a level about 40-50 % of the distance from the anterior to the posterior end of the streak (embryos 9, 10, Table 1, Fig. 1C). At the early head-fold stage, a portion of the mesoderm which would form D (ehf) A (Ims) \ a E (4-7s) F (26-30s) 12*. 21" 11*, 14*, 16*-18* Fig. 1. Drawings (ventral views) illustrating the position of thepresumptivelimb-bud material in the epiblast or mesoderm layer. The positions of [3H]thymidine-labelled grafts (shaded areas) which contributed to the limb-bud epithelium are shown on the left side of each figure. The positions of labelled grafts (shaded areas) which contributed to the limb-bud mesoderm are shown on the right side of each figure. Abbreviations: Ims, late medium-streak stage; ds, definitive-streak stage; hp, head-process stage; ehf, early head-fold stage; s, somite; elb, early limb-bud stage. As in the text, an asterisk (*) after the embryo number indicates that the position shown is that of the graft after its migration in the host embryo. Each graft is therefore shown in two positions—immediately after it was placed, and after incubation. 90 G. C. ROSENQUIST ec Origin of limb-bud 91 the anterior limb-bud (embryos 9*, 11-13, Table 1, Fig. ID) had migrated anteriorly and laterally from the streak into the strip of somatic mesoderm ventral to the ectoderm which would form the anterior limb-bud (Rudnick, 1945 and embryos 6 and 7, Fig. ID). Additional mesoderm which would form portions of the anterior limb-bud was located closer to the streak (embryos 14-20, Table 1; Fig. 1D). Mesoderm which would form the posterior limb-bud was located in the streak or in the nearby mesoderm (embryos 14-20). Mesoderm which would form the ventral portion of each limb was positioned more posteriorly in the streak and more posterolaterally in the lateral plate mesoderm (embryos 12, 19, 20, Table 1, Fig. ID) than was mesoderm which would form the dorsal portions of the limbs (embryos 13, 16-18, Table 1, Fig. ID). The transplants in embryos 11,14 and 15 were placed in an intermediate position and contributed to both dorsal and ventral portions of the somatic layer of mesoderm or the limbs (Table 1, Fig. ID). At the 4-7-somite stage, the limb-bud mesoderm had migrated further into more lateral and anterior portions of the lateral plate. The transplants in embryos 10*, 21 and 22 indicated the position of the mesoderm which would form the anterior limb-bud, which was in the portion of the embryo identified as destined for the anterior limb-bud by Rudnick (1945), Chaube (1959), and by the ectodermal grafts in the present study (embryos l*-4*, 6*, 7*, Table 1, Fig. 1D, E, G). The anterior end of this zone was about at the level of the last somite which had formed; the posterior end of this zone was anterior to the anterior Fig. 2. (A) [3H]thymidine-labelled transplant (arrow) was placed in the epiblast layer of an early head-fold stage recipient embryo (embryo 7). (B) After 66 additional h of incubation, the recipient embryo had a well-developed heart. Cellsfromthe transplant formed the dorsal epithelium of theanterior limb-bud. The plane of the cross-section in (C) is shown by the dotted line at the lower right. s — sinoatrium; v = ventricle. (C) Cross-section (x200) through embryo 7 at the plane indicated in (B), illustrating how the radioautographed cells from the transplant (black grains over nuclei, arrows) formed the epithelium of the trunk of the embryo, extending into the medial (dorsal) portion of the anterior limb-bud, d — dorsal; ec = ectodermal cone; v = ventral. (D) A [3H]thymidine-labelled transplant (arrow) was placed in the endodermmesoderm layer and primitive streak of an early head-fold stage recipient embryo (embryo 20). (E) After 92 additional h of incubation, the recipient embryo had a well-developed heart (Ji) and both anterior (alb) and posterior limb-buds (plb), but was otherwise poorly developed compared to embryos incubated in ovo. Cells from the transplant contributed to the mesenchyme of both anterior and posterior limb-buds. The plane of the cross-section in (F) is indicated by the dotted line through the posterior limbbud. (F) Cross-section (x200) through embryo 20 at the level shown in (E). The radioautographed cells from the transplant (black grains over nuclei, arrows) contributed to the posterior limb-bud mesoderm, which at this level had formed from the somatic layer of the lateral plate, d = dorsal; ec = ectodermal cone; v = ventral. 92 G. C. ROSENQUIST end of the streak. The transplant which later formed the mesoderm of the posterior limb-bud was more posteriorly positioned; it was adjacent to the primitive streak, and extended anteriorly and laterally into the lateral plate mesoderm (embryo 23, Table 1, Fig. IE, G). This position corresponded with the areas previously identified as destined for the posterior limb-bud by Rudnick (1945), Chaube (1959), and an ectodermal graft of the present investigation (embryo 5*, Table 1, Fig. IE). The transplant in embryo 22 contributed to the ventral portion of the anterior limb-bud mesoderm and was positioned slightly lateral to the transplant in embryo 21, which would have contributed to both dorsal and ventral portions of the limb-bud mesoderm. It was in a migration pathway lateral to the pathway which is believed would be followed by the transplant which contributed to the dorsal portion of the posterior limb-bud mesoderm (embryo 23). At the 26-30-somite stage (equivalent to stage 1 of Hamburger, 1938, and 15 of Hamburger & Hamilton, 1951), the cells which would form the mesoderm of the anterior limb-bud were located in the lateral portion of the somatic layer of the lateral plate, posterior to the anterior intestinal portal (embryos 12*, 21*, Table 1, Fig. IF). Cells which would form the mesoderm of the posterior limbbud were even more posteriorly located in the lateral part of the somatic part of the mesoderm (embryos 11*, 14*, 16*—18*, Table 1, Fig. IF). At the early limb-bud stage, the transplants in embryos 13*, 15*, 19*, 20* and 22* all had formed portions of the anterior limb-bud mesoderm (Table 1, Fig. 1G). Of these only the transplants in embryos 22* and 13* did not also form portions of the posterior limb-bud mesoderm; they were among the more anteriorly positioned transplants at the early head-fold and 4-7-somite stages, respectively (Fig. ID, E). The transplant in embryo 23* formed the dorsal portion of the posterior limb-bud mesoderm; it was the most posteriorly positioned transplant at the 4-7-somite stage (Fig. 1E). DISCUSSION Previous attempts to learn the origin and movements of the cells which form the limbs have utilized several different experimental techniques. Wolff (1936) produced localized injury withX-irradiation to embryos asyoungas9-13-somites, and proposed that the anterior limb-bud regions were at the level of the anterior end of the rhomboid sinus (i.e. posterior to the last somite), while the posterior limb-bud regions were close to the midline and posterior to the anterior end of the streak. Rudnick (1945) divided chick blastoderms into full-thickness pieces of various sizes, and transplanted them into the right coelomic space of host embryos incubated 2\ days; after 8-10 additional days of incubation in this environment, the fragments differentiated into morphologically recognizable limbs. Using this method, Rudnick demonstrated that the anterior limb-buds would differentiate from a strip of tissue which extended laterally and posteriorly Origin of limb-bud 93 from the anterior end of the streak at the head-fold stage (Fig. 3 A), and from a strip of tissue which extended laterally from the anterior end of the streak toward the lateral margin of the area pellucida at the 6-7-somite stage (Fig. 3C). Rudnick did not find posterior limb-bud material at the head-fold stage; however, in 6-7-somite stage embryos, she did find such material slightly posterior to the prospective anterior limb-bud material, lateral to the anterior end of the streak (Fig. 3C). Rudnick and Wolff both noted a gradual anterior movement of both anterior and posterior limb-bud regions with additional development, but the presumptive anterior limb-bud remained anterior to the presumptive A 0>f) Fig. 3. Diagrams illustrating the position of the limb-bud material as determined by previous investigators. Left side of each figure: Rudnick, 1945, using coelomic grafting techniques. Right side: Chaube, 1959, using chalk particles in ovo. Hatched area = anterior limb-bud material; shaded area = posterior limb-bud material. /;/= head-fold stage; s = somite. posterior limb-bud in all their preparations. In no instance did limbs differentiate from material which originated at a level anterior to the last somite which had developed. One disadvantage of the studies with grafted blastoderm fragments in the embryonic coelom was that the mesoderm and ectoderm layers contained in the fragments could no longer move independently of each other as they might have done in the uncut blastoderm. The in ovo studies of Chaube (1959) overcame this difficulty. With chalk particles on glass needles introduced from the dorsal surface, Chaube marked adjacent points in the ectoderm and mesoderm layers, beginning at the 2-somite stage. The movement of the chalk particles during the subsequent development of the embryos showed that although both layers 94 G. C. ROSENQUIST expanded in an anteroposterior direction and moved anteriorly in relation to the streak, the points in each layer which were marked with chalk remained adjacent to each other. Chaube thus agreed with Rudnick that the prospective limb mesoderm had completed its gastrulation movements and had reached its definitive position in relation to the ectoderm by the 2-somite stage. Chaube also noted stable right-to-left relationships: laterally placed marks assumed ventral positions in the limb-buds while medially placed marks assumed dorsal positions in the limb-buds. Both Rudnick and Chaube attempted to study the limb regions of younger embryos. Although Rudnick (1945) transplanted portions of embryos as young as the primitive-streak stage into coelomic cultures, limbs did not form from fragments excised before the early head-fold stage. The present study indicates that at the early stages studied by Rudnick, the prelimb ectoderm is in a stable position in the epiblast (ectoderm); however, the prelimb mesoderm which has been invaginated at this stage has not as yet reached a position directly ventral to the prelimb ectoderm, so that the stable relationship between the two which Rudnick found at all later stages had not as yet developed. Chaube marked a total of thirty-five head-fold stage embryos; a large number died soon after they were marked and the remainder 'gave such bizarre results' that they could not be satisfactorily interpreted. From the present study it is clear that at the head-fold stage some of the prelimb mesoderm cells may still be in the epiblast layer, others are in the process of invaginating at the streak, and still others are migrating anteriorly and laterally from the streak toward the prelimb ectoderm regions. Chalk marks placed in such an environment would indeed give bizarre results since only some of the prelimb ectoderm and mesoderm cells have reached a stable position in relation to each other. The present study indicates that the migration of the prelimb mesoderm is as orderly and highly organized as that noted for mesoderm destined for the splanchnic layer (Rosenquist, 1970a). After its imagination at the streak, mesoderm destined for the dorsal portion of each limb is anterior to mesoderm destined for the ventral portion of each limb. This suggests (but does not prove) that equally organized relationships exist in the epiblast prior to invagination at the streak. Premesoderm destined for the dorsal portion of each limb may be anteromedial to premesoderm destined for the ventral portion of each limb. Since the ectoderm which will form the limbs is probably similarly arranged, we may consider the limb regions of the epiblast as consisting of two parallel bands: the more anteromedial band contains the ectoderm and mesoderm cells which will form the dorsal body wall, including the dorsal portion of each limb, while the more posterolateral band contains the ectoderm and mesoderm cells which will form the ventral body wall, including the ventral portion of each limb. The premesoderm is nearer to the streak than is the pre-ectoderm; it migrates to a position below the ectoderm, and then the ectoderm and mesoderm migrate together in their respective bands. Such a concept may also apply to the Origin of limb-bud 95 portions of the epiblast destined to form ectoderm and mesoderm of more proximal and distal portions of the embryo. In future research it should be possible to place smaller transplants into the epiblast layer at these early stages, and incubate the host embryos to later stages, so that further details of these movements may be learned. RESUME Uorigine et les mouvements de Vepithelium et du mesenchyme dans le bourgeon de membre du Poulet demontres par le marquage radioautographique L'origine des cellules du bourgeon de membre a ete mise en evidence en suivant les mouvements de greffes marquees par la [3H]thymidine, excisees d'embryons de Poulet aux stades depuis la ligne primitive moyenne et avanceejusqu'a celui de 5 somites et transplants dans l'epiblaste, la ligne primitive et l'endo-mesoderme d'embryons notes d'age correspondant. Malgre qu'une definition exacte des aires presomptives des membres n'ait pas ete possible vu le nombre limite de greffes executees a chacun des stades, l'etude a pu demontrer de facon generate qu'au stade de la ligne primitive moyenne et avancee l'epithelium du futur bourgeon de membre se place dans la couche epiblastique (dorsale) proche de la marge laterale de 1'area pellucida. II se meut en dedans en direction de l'axe embryonnaire, pour se placer tout juste en dehors des cellules pre-mesodermiques qui vont s'invaginer dans la ligne primitive. Au cours de la regression de la ligne primitive, l'epithelium du membre se deplace relativement vers l'avant, atteignant une position dorsale vis-a-vis du mesoderme de ce membre, et ceci au moins depuis le stade du premier repli cephalique. Au stade de la ligne primitive a son terme, le mesoderme presomptif du bourgeon de membre se trouve dans la couche epiblastique a peu pres a mi-chemin entre la ligne primitive et la marge laterale de l'area pellucida, et a mi-distance entre les extremites anterieure et posterieure de la ligne primitive. A partir de cette position le mesoderme presomptif du membre migre vers le dedans et s'invagine dans la ligne primitive a mi-distance de ses deux extremites (anterieure et posterieure); apres le stade du prolongement cephalique, il migre vers l'avant et le dehors dans la couche parietopleurale de la lame laterale, ventralement par rapport a l'epithelium correspondant. Le mesoderme qui formera le membre anterieur migre en avant de celui qui formera le membre posterieur; le mesoderme qui formera la partie ventrale de chaque membre migre en arriere et en dehors de celui qui formera la partie dorsale. This investigation was supported by USPHS research grants HE 10191 and K3 HE 20074 from the National Heart Institute. The author wishes to thank James D. Ebert for his continued interest in this research, Soame D. Christianson for help in the preparation of the manuscript, and Dorothea Rudnick for a critical reading of the manuscript. REFERENCES S. (1959). On axiation and symmetry in transplanted wing of the chick. /. exp. Zool. 140, 29-77. HAMBURGER, V. (1938). Morphogenetic and axial self-differentiation of transplanted limb primordia of 2-day chick embryos. J. exp. Zool. 77, 379-399. HAMBURGER, V. & HAMILTON, H. L. (1951). A series of normal stages in the development of the chick embryo. /. Morph. 88, 49-92. ROSENQUIST, G. C. (1966). A radioautographic study of labeled grafts in the chick blastoderm. Development from primitive streak stages to stage 12. Carnegie Instn. Wash. Publ. No. 625, Contr. Embryol. 38, 71-110. ROSENQUIST, G. C. (1970a). Cardiogenesis in the chick embryo: topology of the precardiac region from early streak stages until heart formation. Devi Biol. 22, 461-475. CHAUBE, 96 G. C. ROSENQUIST G. C. (19706). The origin and movement of nephrogenic cells in the chick embryo as determined by radioautographic mapping. /. Embryol. exp. Morph. 24, 367-380. RUDNICK, D. (1945). Limb-forming potencies of the chick blastoderm: including notes on associated trunk structures. Trans. Conn. Acad. Arts Sci. 36, 353-377. WOLFF, E. (1936). Les bases de la teratogenese experimentale des Vertebres amniotes, d'apres les resultats de methodes directes. Arc/is Anat. Histol. Embryol. 22, 1-382. ROSENQUIST, (Manuscript received 26 June 1970)