Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
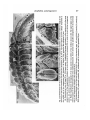
/. Embryol. exp. Morph. Vol. 64, pp. 23-43, 1981 Printed in Great Britain © Company of Biologists Limited 1981 23 Somitogenesis in the amphibian Xenopus laevis: scanning electron microscopic analysis of intrasomitic cellular arrangements during somite rotation By B. WOO YOUN 1 AND GEORGE M. MALACINSKI 2 From the Program in Cellular, Molecular, and Developmental Biology, Department of Biology, Indiana University SUMMARY The intrasomitic changes in cell arrangement which accompany somite rotation during somitogenesis in Xenopus laevis were analysed with the scanning electron microscope (SEM). Longitudinal, horizontal fractures of whole embryos were examined at various dorsoventral levels of stage-22 to -24 embryos. Observations of the gross morphological features of somitogenesis, and the cellular changes which accompany somite segmentation and somite rotation were made. Several of these observations lead to modifications of previous models for the cellular basis of somitogenesis in Xenopus. Individual cellular rearrangements, rather than simultaneous block rotation of a whole somite, appear to be responsible for the 90° rotation of myotomal cells within a single somite. Cellular arrangments in fused somites were also examined. Some ultraviolet-irradiated embryos displayed a complete lack of a notochord. The somites in those embryos were fused across the midline beneath the neural tube. The dorsal and ventral arms of the somites are not fused. Normal rotation occurs only in the dorsal and ventral arms while, in the majority of cases, cells in the fused region fail to rotate normally. In some cases, individual cells in the fused region undergo partial rearrangement. Those observations support the notion that individual cellular rearrangements account for the rotation of the whole somite. INTRODUCTION Analysis of somite formation during amphibian embryogenesis provides various opportunities for gaining insights into both the cellular basis of morphogenesis (Deuchar & Burgess, 1967) and the theoretical aspects of pattern formation (Cooke & Zeeman, 1976). For example, attempts at understanding the manner in which the paraxial mesoderm becomes segmented into somites has been the object of numerous investigations. This issue has been approached in various ways (reviewed by Bellairs, 1979), one of which is the study of the morphological changes which occur during somite segmentation. From obser1 Authors' present address: Department of Biology, Princeton University, Princeton, NJ 08540, U.S.A. 2 Authors' address for all correspondence: Department of Biology, Indiana University, Jordan Hall 138 Bloomington, Tndiana 47405, U.S.A. 24 B. W. YOUN AND G. M. MALACINSKI vations made with the light microscope, Hamilton (1969) developed a model to describe the process of somite segmentation in Xenopus laevis. The paraxial mesodermal cells of Xenopus embryos first elongate perpendicularly to the notochord. The process of somite segmentation then occurs as transverse fissures isolate successive blocks of spindle-shaped cells. Segmentation progresses in a cranial to caudal direction. Each somite then proceeds to rotate through 90 ° with its medial edge moving forward. Individual somites thus become single bundles of spindle-shaped cells (myoblasts) that lie parallel to the notochord. In other amphibia, however, the pattern of somite formation is quite different. In urodeles, for example, the somites form before cell elongation, in a manner comparable to many other vertebrates (e.g. chick, see Bellairs, 1979; Meier, 1979) where each somite forms initially as a rosette. During subsequent differentiation, the myotome cells alter their orientation, become elongated anteroposteriorly, and fuse to form multinucleate muscle fibres (Loeffier, 1968). In the case of Xenopus, several questions remain concerning the mechanical details of somite rotation: Does a whole somite block rotate simultaneously, or do the cells within a somite rotate as individuals? Does a centre of rotation exist around which the other parts of the somite rotate in a manner analagous to an axle and wheel? Do the neural tube, notochord, lateral mesoderm, or dermatome facilitate myotome rotation? What, if any, locomotory processes are involved as individual myotomal cells change position? In a recent study (Youn, Keller & Malacinski, 1980), we provided scanning electron microscopic descriptions of somitogenesis in Xenopus. Those data demonstrated that distinctive changes in cellular morphology and arrangement occur among the paraxial mesoderm cells, as predicted by the earlier descriptions of Hamilton (1969). The present studies were undertaken in order to provide further insights into the mechanical basis of somite rotation. We have examined the cellular arrangement of myotomal cells in the rotating somite itself. When frontal fractures were made through a rotating somite at various dorsoventral levels, groups of myotomal cells were observed to exhibit a gradient of direction in cellular orientation. Moreover, we have extended our discovery that ultraviolet irradiation (U.V.) of the vegetal hemisphere of the fertilized, uncleaved Xenopus egg produces embryos which lack a notochord and display fusion of somites along the midline beneath the neural tube (Youn & Malacinski, 1980a). According to the model for somite morphogenesis proposed by Hamilton (1969), fusion of somites across the midline of the embryo should prevent the rotation of individual somites. Our results, however demonstrate that myotomal cells in both the dorsal and ventral portions of the somite orient in the same direction as those of normal, unfused somites. Those in the fused region do not rotate normally. These observations contradict the idea that simultaneous block rotations occur along the dorsoventral length of the whole somite. Rather, individual cellular rearrangements accompanied by the active cell movements within the somite appear to occur. Amphibian somitogenesis 25 MATERIALS AND METHODS Xenopus laevis embryos were obtained by either gonadotrophic hormoneinduced mating of adults (Gurdon, 1967), or artificial insemination (Wolf & Hendrick, 1971; Youn & Malacinski, 19806). Embryos collected from natural spawnings were chemically dejellied in a 2 % cysteine-HCL solution (pH 7-4 with Tris buffer), allowed to develop in dechlorinated tap water (DTW), and fixed in 2 % glutaraldehyde (pH 7-6 with 0-1 M-cacodylate buffer) at Nieuwkoop & Faber (1967) stage 22-24. Artificially inseminated eggs were U.V.-irradiated according to the method outlined by Malacinski, Brothers & Chung (1977). Irradiated eggs were kept in DTW until the embryos were classified at stage 22-24 according to their defects in neural structures (Malacinski, Benford & Ching, 1977; Youn & Malacinski, 1980#). Embryos which displayed extents of neural defects in the 0- to +2 range ( 0 - , near normal neural morphology; + 2, extreme microcephaly; see Youn & Malacinski, 1980#) were selected and fixed in the 2 % glutaraldehyde solution. Fixed embryos were washed in 0-1 M-cacodylate buffer (pH 7-6). Then the epidermis and neural tube (sometimes together with the notochord) were removed using a fine steel knife and forceps. In order to expose the intrasomitic cellular morphology and arrangement, longitudinal, horizontal fractures were made to each embryo at various dorsoventral levels (see Results for details of the fracture methods). The fractured embryos were dehydrated in increasing concentrations of ethanol and critical-point dried using liquid CO2. All the specimens were mounted on aluminium stubs with conducting silver paint and coated with gold-palladium in a sputter coater E5100 (Polaron Equipment, Ltd). These were observed with an Etec Autoscan U-l SEM and photographed on Polaroid Type 55 positive-negative film. RESULTS I. Morphological features of somitogenesis in Xenopus laevis The cellular arrangement in the paraxial mesoderm is shown diagrammatically in Fig. 1. SEM observations of transverse sections of the paraxial mesoderm at the early neurulation period have been made in the previous paper (Youn et al. 1980). Such observations are summarized here in order to provide some basic information on how changes in cellular morphology and arrangement occur among the paraxial mesodermal cells before somite segmentation begins. At approximately stage 12 f , the paraxial mesoderm consists of two layers of cells. These cells are polyhedral-shaped and are perhaps slightly greater in height than width (Fig. \a). At about stage 13 f the paraxial mesoderm continues to maintain a double-layered configuration, but the cells become more elongated dorsoventrally (Fig. 1 b). Cells near the notochord, where the upper and lower layers meet, have a rather irregular shape but are oriented medio- 26 B. W. YOUN AND G. M. MALACINSKI (a) W (O N Fig. 1. Diagrammatic representation of cellular arrangements (mid-transverse view) in the paraxial mesoderm of Xenopus laevis at about stage 12+ (a), stage 13+ (/?), and stage 18 (c). Pointers and arrow in (c) indicate the prospective dermatomal cells and the pre-myocoel, respectively. N, notochord. (Redrawn from Youn etal., 1980). laterally. When the neural folds rise, changes in arrangement among the paraxial mesoderm cells occur. By stage 18, cells in the upper layer come to lie mediolaterally whereas those in the lower layer lie dorsoventrally (Fig. lc). Cells near the notochord assume a radial arrangment and become elongated further mediolaterally. This arrangement is also observed in the unsegmented posterior portion of stage-22 to -24 embryos. All the prospective myotomal cells become more spindle-shaped than at earlier stages. The prospective dermatomal cells (see pointers in Fig. lc) form a sheet over the lateral surface of the prospective somite. A small pre-myocoel is visible in the unsegmented mesoderm (see arrow in Fig. 1 c). By about stage 18, three to four somites have formed (Youn et ah 1980). As the elevated neural folds fuse and the whole embryo increases in length, more somites are added posteriorly. Figure 2a displays a dissection from a stage-22 to -24 embryo which shows the mediolateral view of the right side of the somite files and the rest of the unsegmented mesoderm. The somite mesoderm can be divided into three regions: (1) Region A, to which the neural tube attaches; (2) region B, where the notochord resides; and (3) region C, where the mesoderm contacts the endodermal roof of the gut. Cells in regions A, B, and C originate at earlier stages from the upper, middle, and lower layers of the paraxial mesoderm, respectively. Those three regions are sharply (pointers in Fig. Id) delineated in the posterior portion of the unsegmented mesoderm. That sculpturing, however, becomes less distinctive in the anterior portion of the unsegmented plate. In the segmented mesoderm, almost no sculpturing is observed. This is probably due to the fact that regions A and C become stretched dorsoventrally in the more anterior unsegmented mesoderm. It can also be seen in Fig. 2a that somite segmentation in the most anterior part of the stage-22 to -24 embryo occurred by the formation of transverse fissures. As a result, the shape of the anterior-most somites is 'stump-like'. More posteriorly, somites are 'bow-shaped'. Further posteriorly - in the last few segmented somites-they display an 'arrowhead-like' shape. The manner in the which the various shapes are acquired at different developmental times is not known. It is probable, however, that differential movements between the neural ectoderm (or epidermis) and the trunk mesoderm during the periods of Fig. 2 (a) Mediolateral view of the right side of the somitefilesand the unsegmented mesoderm of a Xenopus embryo at stage 22-24, showing how the somite mesoderm was divided into three regions, A, B, and C. The three regions are most sharply sculptured in the caudal end of the unsegmented plate (pointers). Ant, anterior; Post, posterior. Bar represents 01 mm. (b) Posteriodorsal view of stage-22 to -24 Xenopus embryo. Longitudinal fractures were made in a few somites and in the most anterior portion of the unsegmented mesoderm at or above the level of the notochord (N), in order to reveal cellular airangement pattern. Ant, anterior; Post, posterior. (c) Higher magnification micrograph of the most posterior two somites of the embryo shown in (b). The upper region of the somites appears to be twisted backward in the mediolateral direction. Arrow points to the parallel arrangement of the myotomal cells in the unsegmented mesoderm. Bar represents 50 fim. (d) Posterotransverse view of the most posterior somite of Xenopus at late neurula stage. NF, neural fold; S, somite; N, notochord; DM, dermatome; EP, epidermal ectoderm; AR, endodermal roof of archenteron. Bar represents 50/*m. t^- Post * Ptost to 1- 28 B. W. YOUN AND G. M. MALACINSKI neurulation and post-neurulation may, in part, contribute to the different shapes of the somites (see Discussion). Viewed dorsally, the upper regions of the arrowhead-like somites appear to be twisted backwards (Fig. 2b-d). Therefore, the initial process of somite segmentation in the upper region of the arrowhead-like somites occurs by the formation of diagonal rather than transverse fissures. Since cells in the unsegmented mesoderm are oriented perpendicularly to the notochord (arrow in Fig. 2 c), the development of transverse fissures should be mechanically more simple. Questions concerning (1) the manner in which the embryo makes such diagonal fissures which are directed against this cellular arrangement, and (2) the role of the somite segmentation pattern in the overall process of somite rotation are, however, beyond the scope of this study. It should be noted here that the dermatome also segments together with the somites (Figs. 2b, c). Kordylewski (1978) previously reported similar observations in Xenopus. These observations, however contradict the earlier description made by Hamilton (1969) that the dermatome remains as a curtain during somite segmentation. The fact that the dermatome also segments raises an important question concerning the problem of somite rotation in Xenopus: Does the dermatome also rotate? Since the medial edge of the rotating somite always goes forward the dermatome in the lateral side moves backward to be positioned in the intersomitic space. This process does not, however, occur in vivo. The prospective myotomal cells should be able to rotate freely and independently of the segmented dermatome (see below). IT. Intrasomitic cellular arrangment in the rotating somite Neither mediolateral nor transverse views (Figs. 2a, d) yielded information about intrasomitic cellular arrangements in the rotating somite. Longitudinal fractures along the whole length of the somite file at three different dorsoventral levels were, therefore, made. An embryo in which a fracture was made through region A is shown in Fig. 3a; through region B in Fig. 3b; and through region C in Fig. 3 c). In all those micrographs, cells in the unsegment?d mesoderm lie perpendicular to the notochord. In the most posterior somite, cells shown in Fig. 3 a as well as in Fig. 3 c are oriented at about 45° to the notochord. This observation confirms the earlier findings of Hamilton (1969). When a fracture is made through region B, the oblique orientation of the myotomal cells in the most posterior somite is rarely seen. As shown in Fig. 3 b, a portion of the cells appear to have completed their rotation. Those cells located in the medial edge are, however, actually in the process of completing rotation. In the more anterior somites the myotomal cells seen in Fig. 3 b are oriented parallel to the long axis of the embryo. Two main conclusions can be drawn from observing the intrasomitic cellular arrangements shown in Fig. 3. Firstly, there exists a dorsoventral gradient in the myotomal cell orientation of the rotating somite. Cells in the upper (A) and Amphibian somitogenesis ;/\ - 29 ivn Fig. 3. Dorsal view of longitudinally fractured somite mesoderm olXenopus at stage 22-24 showing intrasomitic cellular arrangements. Fractures were made through regions A(o), B(b), and C(c). Arrows indicate the last segmented somites. Ant, anterior; Post, posterior. Bars represent 01 mm. EMB 64 30 B. W. YOUN AND G. M. MALACINSKI lower (C) regions of the rotating somite are at the same stage of cellular orientation - i.e. approximately 45° to the notochord. Most of the cells which occupy the middle (B) region have completed rotation through 90° and lie parallel to the notochord. This observation indicates that the cells within an individual somite do not rotate simultaneously along the entire dorsal/ventral length of the somite. Since cells in region B rotate first, there may actually be an intrasomitic sequential wave of a rotational signal. That signal might arise in the middle region first and then be propagated toward both ends of the somite. Secondly, a gradient in the myotomal cell orientation exists within a single fracture plane of a rotating somite (see Fig. 3 b). Observations of these various cell orientations suggest that individual cellular 'rearrangements' rather than the simultaneous block rotation of a whole somite is responsible for the rotation of the myotomal cells within an individual somite by 90°. Further evidence to support the first conclusion (re. dorsal/ventral gradient) was obtained from observations of the type shown in Fig. 4. The two most posterior somites were photographed ventrally or mediolaterally at various tilt angles. In the most posterior somite (viewed ventrally), myotomal cells located in the most ventral region are orientated almost perpendicularly to the notochord (long arrow in Fig. 4 a). However, cells which lie more dorsally approach an orientation which is parallel to the notochord (short arrow in Fig. 4a). These differences in cellular arrangement in the ventrodorsal direction can be observed more clearly at higher tilt angles which reveal both the ventral and the mediolateral aspects of the rotating somite (long and short arrows, in Figs 4b, c). When viewed mediolaterally (Fig. 4d), both the upper and lower parts of the somite are twisted backward in the mediolateral direction. This twisted configuration of the most posterior somite in the upper region can also be seen in Fig. 2 c. A dorsal view of the twisted ventral portion of the somite is shown in Fig. 5. The second conclusion (re. intrasomitic cell patterns) was further strengthened by the results of additional examinations of myotomal cellular arrangements of the type displayed in Fig. 6. In these cases the longitudinal fracture was made between regions A and B (see Fig. 6 a for the lateral view of the fractured whole embryo). A higher magnification dorsal view of the myotomal cellular arrangement is shown in Fig. 6b. As mentioned previously, cells in the unsegmented plate are oriented perpendicularly to the long axis of the embryo (Fig. 3). Cells in the most anterolateral portion of the unsegmented mesoderm, however, assume an oblique orientation (short arrows in Fig. 66). A similar cellular arrangement can also be observed in the comparable region of the unsegmented mesoderm displayed in Fig. 3 b. This indicates that those cells are already in the process of rotating prior to segmentation. The cellular arrangement in the fracture plane of the posterior-most somite is particularly interesting. Cells in the lateral and medial edges are oriented parallel to the notochord (long arrows in Fig. 6 b). Conversely, those cells in the centre portion maintain the parallel Amphibian somitogenesis Fig. 4. Scanning electron micrographs of the two posterior-most (right side) somites of Xenopus at stage 22-24. A fracture was made through region C, and the somite file was inserted to reveal the ventral aspect of cellular arrangement pattern. These somites were then photographed ventrally or mediolaterally at various tilt angles: (a), 0°; (b), 20°; (c), 40°; (d), 60°. Long arrow points to the group of cells which lie perpendicular to the long axisof the embryo. Short arrow points to the group of cells which lie more dorsally than those indicated by the long arrow and which also appear to assume an oblique orientation at higher tilt angles. Ant, anterior; Post, posterior. Bar in (d) represents 50/tm for all the figures shown in Fig. 4. 31 32 B. W. YOUN AND G. M. MALACINSKI Fig. 5. Dorsal view of the posterior-most somite of Xenopus at stage 22-24 showing the parallel cellular arrangement and twisted configuration in the most ventral portion of the somite. A fracture was made through region C. Bar represents 20 /*m. orientation up to a certain point (pointer in Fig. 6b) where they turn sharply toward the notochord. These observations support the second conclusion that individual cellular rearrangements, rather than block rotation of a whole somite, is involved in the turning of the myotomal cells by 90°. These observations also yield another important conclusion - that the rotating myotomal cells have elastic properties. That is, cells bend while in the rearrangement process. Moreover, cells in the medial edge of the most posterior somite which lay parallel to the notochord appear to reside deeper (more ventral) than those other regions of the somite (thick arrow in Fig. 6 a). This parallel arrangement of the myotomal cells can also be observed if a fracture is made more ventrally through this region (Fig. 3 b). This observation also supports the conclusion that the turning of the cells in the middle region (B) occurs prior to rotation of cells in regions A and C. It is also worthwhile to examine the myoblast cellular morphology and the cell-to-cell contacts in both the unsegmented mesoderm and the segmented somites. Most cells in the caudal portion of the unsegmented mesoderm are 'rod-like' rather than'spindle-like'. They are in contact with each other through small protrusions (Fig. 6c). In this region the lateral ends of cells are round and smooth, and display only a few protrusions. Unfortunately, it is difficult to analyse the detailed cellular morphology and contacts in the medial side. The presence of large amounts of extracellular fibrillar materials obscure those contacts. Occasionally it was observed that many cells send out small protrusions which, in turn, appear to connect to the notochord by a network of extracellular fibrils (short arrow in Fig. 6 c). In the most anterior region where the myotomal cells are beginning to change their orientation, cells also show changes Amphibian somitogenesis 33 in morphology - especially at their ends. Their anterior or medial ends become flat and broad (pointers in Fig. 6c), whereas their lateral ends become narrow and spindle-shaped (long arrows in Fig. 6 c). There appears to be less extracellular material in this medial space compared with the posterior portion. This is perhaps mainly due to the fact that the fracture was made at slightly different dorsoventral levels (arrow in Fig. 6a). In the most posterior somite, parallel and oriented cells display broadened ends at the posterior side (pointers in Fig. 6d). Cells which are in the process of rotating and bending are narrow and sharply pointed at both ends (long arrows in Fig. 6d). These observations imply that the extensive changes in cellular morphology occur during myotomal cellular rearrangement and rotation (see Discussion). In the more anteriorly located somites cells become spindle-shaped. Among those a few cells may maintain blunt ends at the posterior side (short arrows in Fig. 6d). In this region extracellular fibrillar materials are abundant only in the region between the notochord and the intersomitic space. III. Intrasomitic cellular arrangement in fused somites Previous studies by Malacinski et al. (1975, 1977) have demonstrated that ultraviolet irradiation (254 nm) of the vegetal hemisphere of the fertilized, uncleaved egg results in abnormal axial structure development. The range of defects in the formation of the axial structures is very broad. Recently, Youn & Malacinski (1980a) employed the SEM to examine various types of defective embryos. In that study the following notations were employed to categorize the types of abnormal embryos employed: 0, no apparent irradiation-induced damage to axial structures; 0~, slightly microcephalic; + 1 , microcephalic; +2, extremely microcephalic; + 3 , poor head morphology and shortened axial structures. In the present study U.V.-irradiated embryos which displayed those neural defects were examined after the epidermis was peeled was observed. It was discovered, for example, that 0~ to +3 embryos lacked a notochord. One result of notochordlessness is the fusion of somites across the midline underneath the neural tube (Fig. 7). The process of somite segmentation in irradiated embryos appeared, however, to occur normally. It was observed that somite counts of 0 to + 2 embryos were approximately the same as those of normal, unirradiated control embryos. If each block of segmented somites on either side must rotate through 90° with its medial edge moving forward, the process of rotation should be prevented by the fusion of somites which occurs in irradiated embryos. Several observations on irradiated embryos were made. Embryos showing 0~ to +2 U. V. defects were selected at stage 22-24. Longitudinal fractures were made through the upper (A), middle (B), and lower (C) regions as shown in Fig. 7. The intrasomitic cellular arrangement was then examined. In Fig. 8a a typical embryo in which a fracture was made through region A is shown. Examination of the intrasomitic cellular arrangement reveals that the prospective myotomal cells 34 — B. W. YOUN AND G. M. MALACINSKI <aj (b) N i 1 Fig. 6. Dynamic changes in myotomal cell shape during rotation. Somite rotation is a dynamic process which produces strikingly different cell shapes at different developmental times and at various dorsoventral levels. The process of rotation must occur very rapidly at the time of somite segmentation. Therefore, the probability of obtaining a fracture/section in which cells in the last segmenting somite show a bent configuration and different morphologies at their ends during the process of rotation is very low (we estimate that probability to be less than 1 %). Fig. 6. (a) Lateral view of the posterior half of Xenopus embryo at stage 22-24 showing how the fracture was made. Arrow points to a deeply fractured region in the unsegmented plate. Ant, anterior; Post, posterior. Bar represents 0-1 fim. (b) Higher magnification of dorsal view of the longitudinally fractured somite mesoderm of the embryo shown in (a) displaying myotomal cellular arrangement of the last five somites and the unsegmented plate. The oblique orientation of the cells in the most anterior portion of the unsegmented plate is indicated by the short arrows. Long arrows point to cells arranged in parallel in the lateral and medial edges of the most posterior somite. A cell with a bent configuration is indicated by the pointer. N. notochord. Bar represents 0-1 mm. (c) Higher magnification micrograph of the unsegmented mesoderm (dorsal view) taken from the region indicated in (b). Additional information on the cellular morphology arrangement, and contacts can be obtained from this micrograph. Short arrow points to an example of a cell process which might have been connected to the notochord through a network of extracellular fibrils. Long arrows indicate sharply pointed lateral (prospective caudal) ends of the turning myotomal cells. Pointers 35 Amphibian somitogenesis Fracture levels Fig. 7. Schematic diagram of the cross-sectional view of the fused somite. The three regions A, B, and C (also see Fig. Id) are indicated. Approximate levels of fractures made in each region for observation are indicated by solid lines. undergo the same rotation processes seen in normal embryos (Fig. 3 and 6). ]n the unsegmented plate, cells lie perpendicularly to the long axis of the embryo. Cells in the most posterior somites rotate, while those in the rest of the somites become oriented parallel to the long embryonic axis. In Fig. 8Z>, a fracture through region A is displayed. A few mistakes in the process of somite segmentation can be observed. The somites on either side are not matched symmetrically (the figure legend contains a detailed description of this abnormal, asymmetric segmentation). The same cellular arrangement can, however, be seen along the long axis of the embryo. When a fracture was made through region C (Fig. 8 c), a similar rotation process was also observed. The perpendicular arrangement of cells in the unsegmented mesoderm, the oblique orientation of the turning cells in the most posterior somite, and the parallel arrangement in the more anteriorly located somites were all observed. The turning myotomal cells in the most posterior somite reside, however, in a reverse orientation compared with those in Figs. 8#, b. This is because in Fig. 8c the fractured embryo was inverted to reveal the ventral aspects of the intrasomitic cellular arrangement. The intracellular arrangement in the middle (fused) region is complex (Fig. 9). Cells in this region are oriented in several directions. Cells in the unsegmented plate are elongated in the mediolateral direction. The unsegmented plate is a indicate broad, flattened protrusions at the prospective anterior ends of the cell which are about to turn. (d) Higher magnification micrograph of the last three somites taken from the region indicated in (b). In the most caudal somite some cells are already arrayed parallel to the notochord with their posterior ends being broad (pointers), while others still show a perpendicular orientation. Cells which are in the process of rotating and bending appear to have narrow and sharply pointed ends (long arrows). Short arrows indicate cells with blunt ends at the posterior side. Bar represents 20/<m for (c) and (cl). 36 B. W. YOUN AND G. M. MALACINSKI Fig. 8. (a) Dorsal views of the myotomal cellular arrangement in the upper (A) region of the fused somites in a U.V.-irradiated Xenopus embryo a tstage 22-24. Arrow indicates the most posterior somite. Ant, anterior; Post, posterior for all the embryos shown in Fig. 8. Bar represents 01 mm. (6) Dorsal view of myotomal cellular arrangement in the upper (A) region of the fused somites. U.V. irradiation sometimes causes mismatched, asymmetric fusion. An extra somite which forms as a result of the mismatch is indicated by thin, long arrow. However, the same perpendicular orientation in the unsegmented mesoderm and parallel arrangement in the somites are obtained in this abnormal embryo. Thick, short arrow points to the most posterior somite which contains the cells with oblique orientation. Bar represents 01 mm. (c) Ventral view of myotomal cellular arrangement in the lower region (C) of the fused somites. After a fracture was made, the embryo was inverted to reveal the ventral aspect of the cellular arrangement pattern. Arrow indicates the most posterior somite. Bar represents 01 mm. Amphibian somitogenesis lb) Fig. 9. Dorsal views of the myotomal cellular arrangement in the middle (B) fused region of the fused somites in U.V.-irradiated Xenopus embryos. Approximately 60 notochordless embryos from thtee separate irradiations were examined. Each of the three irradiations represented spawnings of different females. Three typical examples are shown in this figure. Pointers in (a) and (c) indicate cells which exhibit oblique orientations. Thin, short arrows in (a), (b), and (c) point to a group of cells in the lateral side of somites which assume parallel orientation. Sometimes fractures were made through the level slightly higher (more dorsally) than other regions of the same embryo, such that all the myotomal cells indicated by long arrows in (/?) are perfectly arranged parallel to the long axis of the embryo. Thick short arrows indicate the most posterior somites. Ant, anterior; Post, posterior. Bars represent 01 mm. 37 38 B. W. YOUN AND G. M. MALACINSKI few cells wide rather than just one cell wide. In some of the posterior-most somites, cells either do not display any definite orientation (Fig. 9 a), show the correct parallel arrangement pattern (Fig. 9 b), or display the perpendicular orientation (Fig. 9 c). The parallel arrangement shown in Fig. 9 b (long arrows) may be due to the fact that the fracture level is slightly higher (more dorsal) than other regions of the same embryo, or the same, comparable regions of other embryos shown in Figs. 9 a, c. Therefore, the parallel-cell arrangement appears to reflect the cell orientation pattern only in the upper region (Figs. 9 a, 9b). Further anteriorly, cells become more spindle-shaped and tend to maintain the perpendicular arrangement. Again, the parallel arrangement of the cells seen frequently in the lateral side (short arrows in Figs 9a-c) can be regarded as a continuation of the cell orientation pattern from the upper region. The oblique orientation of the cells (pointers in Figs. 9 a, c) can be observed if they are in a transitory zone, changing from the parallel to the perpendicular arrangement. It is possible that this represents independent rotation of cells originally derived from the left and right sides of the embryos. The turning of prospective myotomal cells by 90° in the upper and lower regions (Fig. 8) even under the condition of somite fusion provides additional support for the conclusion drawn in the previous section of this report: Individual cellular rearrangement, rather than simultaneous block rotation of a whole somite, plays an important role in the rotation process. In the previous section of this report it was also suggested that since the myotomal cells in the middle region turn first, the motive force for the cells in the upper and lower regions to turn might be exerted from the middle region of the somite and propagated towards both ends. That suggestion probably does not pertain to irradiated embryos since cells in the middle region tend to be oriented perpendicularly to the long axis of the embryo (Fig. 9). These observations imply that the prospective myotomal cells in each dorsoventral level have an intrinsic capacity for cellular rearrangement and, therefore, turn independently of each other. The fusion of somites somehow prevents the cells in the middle region from behaving in the normal fashion (see Discussion). DISCUSSION The first point for discussion is the relative roles that morphogenetic movements of the mesodermal mantle, the superficial epidermis, and the prospective neural area play in the shaping of Xenopus somites. It was shown in Fig. 2a that in a mediolateral view the shape of the somites displays a progressive change in the anteroposterior direction from a stump-like to an arrowhead-like morphology. The somite reversal experiments of Deuchar and Burgess (1967) have also shown that grafted, reversed pieces take on a new orientation to correspond with the rest of the host axis. They suggested from those results that some overall influence from surrounding tissues governed the anteroposterior orientation of Amphibian somitogenesis 39 the somites, and that the orientation was not irrevocably fixed until after the end of the neurulation. The manner in which the surrounding tissue might affect the overall shaping of the somites at different developmental times could be explained in part by the results of vital-dye mapping experiments. Keller (1976) placed dye marks through the double-layered prospective neural area and the underlying prospective dorsal mesoderm at stage 13 and studied the relative movements of these layers during neurulation. Examination of the locations of the dye marks in cross sections at either stage 18 or 22 revealed that relative anteroposterior movements occur between the dorsal epidermis and the lateral somite mesoderm (i.e. Fig. 2a, region C) and also between the neural plate and the somite mesoderm (i.e. Fig. 2a, region A) further medially. The neural and dorsal epidermal ectoderm moves posteriorly relative to the somite mesoderm. This movement increases in the more posterior regions. The medial register of the somite mesoderm and the overlying neural or epidermal ectoderm (i.e. Fig. 2a, region B), is however, essentially preserved throughout neurulation. In the regions where the relative anteroposterior movement takes place, the upper (A) and lower (C) portions of the somites come to lie further posteriorly with respect to the middle portion (B). The extent of the contribution of relative movements varies in different regions of the embryo. Somites which form in the posterior region of the embryo acquire an arrowhead-like shape, since most of the movement occurs in this region. Those which develop more anteriorly lose the arrowhead-like shape and assume a bow-like configuration. The shape of the first few somites located in the most anterior region of the embryo is stump-like, because they are least affected by that relative anteroposterior movement. The second point for discussion concerns the rotation of somites. Is somite rotation a property of the individual myotomal cells or the whole somite? Cooke (1977) observed with light microscopic examination of horizontal sections through whole embryos that cells of each somite block maintain a close contact, yet move partly as individuals. Our SEM findings support that observation. Individual cellular rearrangements, rather than the simultaneous block rotation of a whole somite, appear to be responsible for the rotation of myotomal cells by 90° (Figs. 4 and 6). Individual cell rearrangements do not, however, alone account for the orientation pattern of somite cells in U.V.irradiated embryos. Cells in the fused region tend to be oriented perpendicularly to the long axis of the embryo (see Fig. 9). There are at least two possible explanations for the abnormal behaviour of cells in the fused somite region of U.V.-irradiated embryos. Firstly, cells in that region may not be truly myotomal, in the sense that they could have originated from the group of cells which would normally have become the notochord. Those cells may still have some of the characteristics of notochordal cells at these relatively early stages (stage 22-24). Such cells conceivably could be unable to exhibit the rotational movements of authentic myotomal cells. Further detailed 40 B. W. YOUN AND G. M. MALACINSKI ultrastructural studies using transmission electron microscopy are needed in order to test this possibility. Secondly, if the cells in the fused region are indeed true myotomal cells, then the bilateral separation of somites caused by the presence of the notochord must be considered essential for the rotation of myotomal cells by 90°. In U.V.-irradiated embryos cells in the upper and lower (unfused) portions of the somite were shown to be arranged in a normal, parallel orientation (Fig. 8). The manner in which the fusion of somites hinders the rotational behaviour of the myotomal cells remains obscure. Furthermore, the possibility that the neural tube, notochord, lateral mesoderm, or dermatome might aid in myotome rotation was not tested in the present study. Such structures are known to be involved in the proper shaping of chick somites by exerting packing-pressure on them (Packard & Jacobson, 1979). The third and last point for discussion concerns the mechanisms of motile and adhesive control through which the myotomal cells as individuals might execute such rearrangements during somite rotation. Cell shape changes have been implicated in many developmental processes involving tissue folding, bending, invagination, etc. For example, cell shape changes are a primary cause of neural-fold formation (reviewed by Karfunkel, 1974) "and mesodermal migration following involution (Keller & Schoenwolf, 1977). Various mechanisms of motile and adhesive control are postulated to be involved in those processes (Trinkaus, 1976). In chick somitogenesis, Meier (1979) has reported that somite cell rearrangements precede the process of somite segmentation. Changes in cell contacts appear to be responsible for the formation of the 'somitomere-like structure', the segmentation process, and rosette formation (Bellairs, 1979; Bellairs, Curtis & Sanders, 1978). These changes may be due to changes in cell adhesion. The scanning electron micrographs in Fig. 6 also reveal that extensive changes in cellular morphology occur during the rotation of Xenopus myotomal cells by 90°. The turning process itself can be divided into several distinct steps in which the cells exhibit typical changes in morphology and orientation (Fig. 10). (a) Step I. During this initial period of rotation, myotomal cells in the most anterior region of the unsegmented plate begin to change their arrangements and assume oblique orientations. Meanwhile, those cells also display changes in morphology, especially at their ends. Their medial or prospective anterior ends are flat and broad, whereas their lateral or prospective posterior ends become narrow and spindle-shaped. Thus, the cells become polarized. The broad, flattened protrusions at the prospective anterior ends resemble the lamellipodia of cells which are propagated in culture. They are attached to neighbouring cells by short filiform protrusions. Such lamelliform protrusions may actually function in locomotion and, thus, affect the rearrangement of myotomal cells. At present, however, there is no direct evidence to support this speculation. Amphibian somitogenesis Unsegmented mesoderm Last segmented somite 41 Anterior somites Fig. 10. Schematic diagram of the myotomal cells in the somite mesoderm showing how changes in cell shape and the presence of cellular processes and contacts might be related to the rotation process. Cells exhibiting typical morphologies were selected from Figs 6 c and 6d and redrawn. (b) Step II. As the process of rotating and bending continues, cells become narrow and sharply pointed at both ends. It is important to determine whether component cells of the myotome retain their original neighbour relationships during rotation. Cell slippage and neighbour exchanges could cause relaxation of the tensions that produce cell bending. Our SEM observations (e.g., Fig. 6) show that individual myotomal cells are in contact with many of their neighbouring cells along their sides by many short finger-like protrusions. Each cell appears to remain firmly affixed to its neighbours during rotation. In such cases, bent cells are, most likely, under considerable tension. In order to endure such tension, the cells may need strong cell-to-cell adhesive interactions at both ends. The changes in shape that take place in fibroblasts and similar cells when they are migrating in culture (reviewed by Trinkaus, 1976) could provide insight into the mechanics of somite rotation. When these cells are attached to the substrate by both ends, they are put under tension. As mentioned above, both ends of the bent cells appear to be narrow and sharply pointed. Bent cells are bipolar and usually have an elastic property. Polarity and elasticity may depend on contractile changes in the cytoskeletal network of the cell. Contractile proteins are known to be active in generating cell shape changes (reviewed in Amer. Zool., vol. 13 (1973), 'Factors controlling cell shape during development'; see also Burnside, 1978). Those types of proteins are also involved in cell movements (reviewed in Ciba Symp., vol.14 (1973), 'Locomotion of tissue cells'). It is not known, however, how relationships between neighbouring cells affect cell shape changes which accompany the rotation of a whole somite. Since the myotomal cells in the middle region (B) start to rotate first, environmental and/or intrinsic clues may initially be given to those cells. Since the cells are thought to remain firmly affixed to their neighbours (see above), cell shape changes in those cells could act together to produce tensions which result in the sequential rotation of other cells in the upper (A) and the lower (C) regions. Thus, a whole somite can be regarded as a deformable solid. (c) Steps III and IV. In Step 111 the caudal ends of cells become round while 42 B. W. YOUN AND G. M. MALACINSKI the anterior ends continue to maintain a sharply pointed configuration. At the same time the cells become oriented parallel to one another. The caudal end becomes round in shape, perhaps because the cell has been released from the tension at that end. Finally, in Step IV, the anterior end acquires a round shape. In these later stages it seems that changes in cell shape and orientation could be related to the changes of tension applied to various portions of the myotomal cells during rotation. Conversely, the initial process of rotation (Steps I and II) may require both cellular motile activities and the generation of tension. Although the present SEM observations on the myotomal cellular morphology and the distribution of cell processes and contacts suggest that individual cellular rearrangement accounts for the process of somite rotation, it is still difficult to reconstruct the process with certainty from the static images obtainable with SEM. In order to understand how the turning of the myotomal cells by 90° occurs in vivo, it would be necessary to examine behaviour properties of the cells in vitro under various conditions. In vitro studies with different cell types have contributed to an understanding of the machinery used in cell shape changes and motile behaviour. For example, Bellairs et al. (1978, 1980) have shown that unsegmented mesoderm, newly segmented mesoderm, and differentiating somites exhibit striking differences in behaviour when explanted in vitro. Analysis of the dense cultures of human lung fibroblasts by Elsdale & Wasoff (1976) has revealed that cells in closely packed arrays do not behave independently but are influenced in both their shape and orientation by global constraints arising out of their interactions. These studies may serve as a guide to what factors might affect the behavioural pattern of the myotomal cells of Xenopus in vivo. Thus, the use of both cell culture and organ culture techniques probably represent the next logical step in any attempts to establish the casual relationships which actively promote amphibian somitogenesis. We wish to thank Dr Ray Keller for this advice and encouragement during the course of this investigation. This work was initiated with financial support from NSF PCM 77-04457, and completed with NSF PCM 80-06343 and NASA NAGW-60 support. REFERENCES BELLAIRS, R. (1979). The mechanism of somite segmentation in the chick embryo. / . Embryol. exp. Morph. 51, 227-243. BELLAIRS, R., CURTIS, A. S. G. & SANDERS, E. J. (1978). Cell adhesiveness and embryonic differentiation. / . Embryol. exp. Morph. 46, 207-213. BELLAIRS, R., SANDORS, E. J. & PORTCH, P. A. (1980). Behavioural properties of chick somitic mesoderm and lateral plate when explanted in vitro. J. Embryol. exp. Morph. 56, 41-58. BURNSIDE, B. (1978). Thin (actin) and thick (myosin like) filaments in cone contraction in the teleost retina. / . Cell Biol. 78, 227-246. COOKE, J. (1977). The control of somite number during amphibian development: Models and experiments. In Vertebrate Limb and Somite Morphogenesis (ed. D. A. Ede, J. R. Hinchliffe & M. Balls). Cambridge: Cambridge University Press. COOKE, J. & ZEEMAN, E. C. (1976). A clock and wavefront model for control of the number of repeated structures during animal morphogenesis. / . theor. Biol. 58, 455-476. Amphibian somitogenesis 43 M. & BURGESS, A. M. C. (1967). Somite segmentation in amphibian embryos: Is there a transmitted control mechanism? /. Embryol. exp. Morph. 17, 349-358. ELSDALE, T. & WASOFF, F. (1976). Fibroblast cultures and dermatoglyphics: The topology of two planar patterns. Wilhelm Roux' Archiv. devl. Biol. 180, 121-147. GURDON, J. B. (1967). African clawed frogs. In Methods in Developmental Biology (eds. Fred H. Wilt & Thomas Y. Wessels), pp. 75-84. New York: Crowell. HAMILTON, L. (1969). The formation of somites in Xenopus. J. Embryol. exp. Morph. 22, 253-264. KARFUNKEL, P. (1974). The mechanisms of neural tube formation. Int. Rev. Cytol. 38, 245-271. KELLER, R. E. (1976). Vital dye mapping of the gastrula and neurula of Xenopus laevis II. Prospective areas and morphogenetic movements of the deep layer. Devi Biol. 51, 118-137. KELLER, R. E. & SCHOENWOLF, G. C. (1977). An SEM study of cellular morphology, contact, and arrangement, as related to gastrulation in Xenopus laevis. Wilhelm Roux1 Archiv devl. Biol. 183, 165-186. KORDYLEWSKI, L. (1978). Scanning electron microscopic observations of the development of the somites and their innervation in anuran larvae. / . Embryol. exp. Morph. 45, 215-227. LOEFFLER, C. A. (1968). Evidence for the fusion of myoblasts in amphibian embryos. I. Homoplastic transplantations of somitic material labeled with tritiated thymidine. /. Morph. 128, 403-426. MALACINSKI, G. M., BENFORD, H. & CHUNG, H. M. (1975). Association of an ultraviolet irradiation sensitive cytoplasmic localization with the future dorsal side of the amphibian egg. /. exp. Zool. 191, 97-110. MALACINSKI, G. M., BROTHERS, A. J. & CHUNG, H. M. (1977). Destiuction of components of the neural induction system of the amphibian egg with ultraviolet irradiation. Devl Biol. 56, 24-39. MEIER, S. (1979). Development of the chick embryo mesoblast: Formation of the embryonic axis and establishment of metameric pattern. Devl Biol. 73, 25-45. NIEUWKOOP, P. D. & FABER, J. (1967). Normal Table of Xenopus laevis (Daudin), 2nd ed. Amsterdam: North-Holland. PACKARD, D. S., JR. & JACOBSON, A. G. (1979). Analysis of the physical forces that influence the shape of chick somites. /. exp. Zool. 207, 81-92. TRINKAUS, J. P. (1976). On the mechanism of metazoan cell movements. In The Cell Surface in Animal Embryogenesis and Development (ed. G.Poste & G. L. Nicholson), pp. 225-329. Amsterdam: North-Holland. WOLF, D. P. & HENDRICK, J. L. (1971). A molecular approach to fertilization. II. Variability and artificial fertilization of Xenopus laevis gametes. Devl Biol. 25, 348-359. YOUN, B. W., KELLER, R. E. & MALACINSKI, G. M. (1980). An atlas of notochord and somite morphogenesis in several anuran and urodelean amphibians. / . Embryol. exp. Morph. 59, 223-247. YOUN, B. W. & MALACINSKI, G. M. (1980a). Axial structure development in UV-irradiated (notochord-defective) amphibian embryos. Devl Biol. (in press). YOUN, B. W. & MALACINSKI, G. M. (19806). Action spectrum for ultraviolet irradiation inactivation of a cytoplasmic component(s) required for neural induction in amphibian egg. /. exp. Zool. 211, 369-377. DEUCHAR,E. (Received 3 October 1980, revised 10 February 1981)