Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
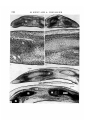
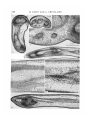
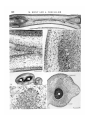
/ . Embryol. exp. Morph. Vol. 49, pp. 153-165,1979 Printed in Great Britain © Company of Biologists Limited 1979 153 Autonomy of tendon development in the embryonic chick wing By MADELEINE KIENY 1 AND ALAIN CHEVALLIER 1 From the Laboratoire de Zoologie et de Biologie animate, Equipe de recherche associee au CNRS n° 621: 'Morphogenese expe'rimentale\ Universite scientifique et medicale de Grenoble SUMMARY The aim of this study performed in the embryonic chick wing is to test the ability of the tendons to form and develop in the absence of the muscle bellies. The experiments were performed on 2-day chick embryos by destroying a portion of the somitic mesoderm by local X-irradiation. The irradiated part included the wing somite level 15-20 and extended three somites (or presumptive somites) in front and two to six presumptive somites in the rear of the wing somite level. The wings of the operated side were examined histologically 3-8 days after the X-irradiation. The radio-destruction of the somitic mesoderm totally inhibited or severely impaired the development of the forearm muscles. But, despite the absence of the flexor and extensor muscles the differentiation of the distal manus tendons could be observed. This differentiation occurred at the same time and in the same position as in controls. However, these tendons were transient structures. They disappeared within three days after their individuation. Two mechanisms that progressed in proximo-distal direction were involved in their resorption: cellular dislocation and cell death. We conclude that tendons start to develop autonomously from the muscle bulks, but for their maintenance and further development they require connexion to a muscle belly. INTRODUCTION Investigations using the chick/quail marker system and performed independently by two groups of authors (Christ, Jacob & Jacob (1974, 1911a, b), on one hand, and Chevallier, Kieny & Mauger (1976, 1977) and Chevallier, Kieny, Mauger & Sengel (1977) on the other hand), have shown that the limb musculature is of double mesodermal origin. Each anatomical muscle appears as a composite structure, the muscle cells being mostly if not totally of somitic origin, the tendons and the connective network (epimysium, perimysium, endomysium) being of somatopleural origin. Furthermore, our group (Chevallier et al. 1976, 1977), specified that the organogenesis of the limb musculature did not depend on the cephalo-caudal level 1 Authors' address: Laboratoire de Zoologie et Biologie animale, Univeisite scientifique et medicale de Grenoble, B.P. 53 X, 38041 Grenoble Cedex, France. 154 M. KIENY AND A. CHEVALLIER from where the somite-myogenic cells emigrated. Indeed, the somitic mesoderm adjacent to the presumptive wing territory could be replaced by the somitic mesoderm of any cephalo-caudal level (neck, flank, leg) without causing modifications of the muscular organogenesis at 12 days of incubation. Thus, it became apparent that the somatopleural component of the anatomical muscle played an active role in the muscular arrangement. Moreover, the destruction by local X-irradiation of a portion of somitic mesoderm, which comprised the part adjacent to the wing primordium and, in addition, extended in front and in the rear of it, prevented the wing mesoderm from being invaded by somitic cells (Chevallier et al. 1978). Thus, a high proportion of 13-day wings, on the irradiated side, contained neither muscles, nor tendons or connective network, myogenic and tendinous areas being occupied by loose unorganized mesenchyme. From this tentative conclusion regarding the active role of the somatopleural component, the question arose whether, during the development of the wings deprived of myogenic cells, the tendon blastemae did not form or whether they did form and disappeared afterwards. The latter possibility would then suggest that, at the very beginning of their development, tendons and muscles would be independent, or at least the tendon's development would not require the presence of myogenic cells. In order to test this developmental independency, we undertook a new series of radio-destruction of the somitic mesoderm at the wing level; the wings on the operated side were examined histologically 3-8 days after the X-irradiation. MATERIALS AND METHODS The experiments were carried out on 2-day chick embryos (Wyandotte x Rhode Island Red) that reached, at the time of operation, the stages of 12-20, usually 16-18, pairs of somites. A portion of somitic mesoderm on the right side was destroyed by local Xirradiation, which was made under the same conditions as reported in the previous paper (Chevallier et al. 1978), namely under 20 kV and 30 mA, during 10 min, the embryo being placed at a distance of 37 cm from the anticathode. The irradiation (ca. 10000 rad.) was performed through a rectangular (1-6 mm long x 0-1 mm wide) slot cut through an 0-1 mm thick tantalum screen that protected the rest of the embryo and part of the extra-embryonic area. The slotted shield was placed in such a way that a 1-6 mm long portion of partially segmented or unsegmented somitic mesoderm behind the 11th somite was irradiated on the right side. Thus, the irradiated part always included wing somite levels 15-20 and the somite levels 12-14 in front of it, while the extent in the rear of the wing level varied, according to the stage of the embryo, from two to six presumptive somites. All experiments were performed before the stage of 21 pairs of somites, i.e. before the normally invading somitic cells had started their immigration into the wing primordium (Chevallier, 1978). Tendon development in embryonic chick wing 155 The irradiated embryos were sacrificed daily after 3-8 days reincubation (to stages 25-37 of Hamburger & Hamilton (1951)). Thirty-seven right experimental wings were examined together with 19 control left wings. They were fixed in Bouin-Hollande's fixative, paraffin-embedded, longitudinally (for the forearm but not necessarily also for the hand) sectioned at 7-7-5 fim. and stained with haemal um-eosin. The sections were viewed and photographed every 70-75/tm with a Zeiss photomicroscope. RESULTS As in previous papers we confined our study to the (intrinsic) muscles that develop entirely within the limb itself, namely to the dorsal extensor and ventral flexor muscles contained in the lower arm and to their distal tendons that insert onto the cartilaginous digits. Our purpose is not to give a detailed study of normal muscle and tendon differentiation, as this is being done by Shellswell and coworkers (reported in Shellswell & Wolpert (1977)), but only to use normalcy as the control. We know (Mauger, 1970) that within the irradiated portion of somitic mesoderm, the dermatome fails to differentiate, which leads to plumage deficiencies in the corresponding portion of the spinal feather tract. Moreover, we have shown (Chevallier et al. 1978) that muscleless wings are always associated with these plumage deficiencies. In this new series of local X-irradiation only the morphology of the spinal tract of those irradiated embryos that had reached at least 7 days incubation when sacrificed could be used as a test for the efficacy of the X-irradiation, the plumage development being then far enough advanced. In addition, it must be emphasized that the local X-irradiation of the somitic mesoderm did not prevent any of the 37 experimental wings from being innervated (Figs 3, 5, 7, 8, 10, 11, 12, 13). (1) 3-4 days after irradiation. Before 6 days incubation, there were no discernible differences in the histological aspect, in particular in the pattern of compactness of the mesoderm, in the experimental (five cases) versus the control wings. From 6 days incubation on, some differences became apparent. In controls, two dense condensations surrounded the radius and ulna anlagen dorsally and ventrally. In the experimental wings (four stage-27 embryos) the corresponding regions appeared less compact than in the controls, but nevertheless were distinctly denser than the subcutaneous mesenchyme (Figs 1-3). In slightly older cases (stages 28-29), the differences between control and experimental wings were more conspicuous. Four out of the six experimental wings were characterized by an absence of muscular blocks around the cartilages and by an evident loosening of the mesenchyme, which, however, remained more compact than the subcutaneous mesenchyme. The two remaining cases had the same aspect as the controls in which dorsal and ventral muscle masses had split into blocks. Neither in the experimental nor in the control wings were the tendons well defined. 156 M. KIENY AND A. CHEVALLIER Tendon development in embryonic chick wing 157 (2) 5 days after irradiation. With increasing age (stages 30-31) the abovementioned features became more accentuated. Whereas in the controls the subdivision into muscular blocks or even individual muscles had taken place (Fig. 6), in four out of the seven experimental wings the muscular areas remained devoid of any individuated muscular masses, although the cellular density of the area appeared high as compared to that of the adjacent subcutaneous mesenchyme, but low compared to that of the subdivided muscle masses in the controls. In one experimental wing, however, two of the 12 forearm muscles were developing within the otherwise unorganized mesenchyme, dorsally to the radius. According to their location, one of them was interpreted as the extensor metacarpi radialis (EMR) and the other as an interosseus muscle, probably the extensor medius longus (EML). Finally the two remaining experimental wings did not differ from the controls. Tendons. In the controls and in the two above-mentioned experimental wings from normally feathered embryos, tendons developed towards the humerus epiphysis, and dorsally and ventrally to the digits. In the other five experimental wings the proximal tendons were not discernible, but the differentiation of the distal ones occurred more or less normally (compare Figs 5 and 6). Four tendons running dorsally and ventrally to the digital skeleton and to the wrist (and extending for some of them into the distal zeugopod) had differentiated (Figs 4, 5). They can be tentatively identified as the EML (extensor medius longus) (or EDC (extensor digitorum communis)), EMU (extensor metacarpi ulnari) tendons, on the dorsal side; and the FDP (flexor digitorum profundus), FDS (flexor digitorum superficialis) tendons, on the ventral side. These wings were from embryos whose spinal tract presented an indentation of bare skin on the right side. FIGURES 1-6 Histological aspect of wings in 6-day (Figs 1, 3) and 7-day (Figs 4, 5) embryos whose somitic mesoderm has been X-irradiated respectively at stages of 16 and 18 somite pairs. Figs 1, 2. Longitudinal sections through the interosseous zone of the forearm and part of the hand in experimental (Fig. 1) and control (Fig. 2) wings 4 days after irradiation. The muscle forming area (MA) is less compact in the experimental than in the control wing, but nevertheless denser than the subcutaneous mesenchyme. DM, Dorsal muscle mass; VM, ventral muscle mass. Fig. 3. Detail of Fig. 1. N, Nerve. Figs 4-6. Histological sections of experimental (Figs 4, 5) and control (Fig. 6) wings 5 days after irradiation passing approximately at the same longitudinal level. Despite the absence of muscle bellies (Mb) in the experimental wing (compare Figs 5 and 6), tendons developed dorsally and ventrally to the skeleton of the hand (Figs 4, 5). H, Humerus; Mclll, metacarp of digit III; TV, nerve; R, radius; I. EDC, tendon of extensor digitorum communis muscle; T. FDP, tendon of flexor digitorum profundus muscle; U, ulna. (Scale bar: 100/tm for Figs 3, 4 and 200 /tm for Figs 1, 2, 5 and 6.) II EMB49 158 M. KIENY AND A. CHEVALLIER Tendon development in embryonic chick wing 159 (3) 6 days after irradiation. The spinal tract of all seven irradiated embryos sacrificed at 8 days incubation (stages 32-33) presented a glabrous indentation at the right wing level. Whereas in the controls the forearm muscle organogenesis was proceeding and the individual muscles showed a well defined epimysium, in the experimental wings the mesenchyme was characterized by the absence (four cases) or severe deficiency (three cases) of the muscle bulks. It appeared as a loose unorganized tissue within which part of the muscle-forming area remained discernible due to a denser region of central mesenchyme (Figs 7,11). The three poorly muscled wings each contained one complete anatomical muscle, that could be recognized as the extensor metacarpi radialis (EMR) in two cases and the flexor carpi ulnaris (FCU) in one case, originating respectively from the initial dorsal and ventral muscular masses. Regarding the organogenesis of the hand muscles, in the controls the muscle masses were individuated, but in the experimental wings no muscle bulk had formed. Despite the complete or nearly complete absence of the muscle bellies in the forearm, the four already mentioned tendons were present. But their proximal ending contained numerous necrotic cells; and the cellular debris seemed to be mainly eliminated into the blood circulation, rarely through the scavenger action of phagocytes. However, dorsal and ventral tendons did not always end at the same proximo-distal level of the wing (Figs 8, 9). Usually they ended both in the distal zeugopod or wrist region, but sometimes the dorsal tendons failed in this region and they ended in the middle of the metacarpal region. In addition to these four tendons belonging to digit III and IV, one or two tendons belonging to digit II had differentiated (Fig. 8). The dorsal tendon might FIGURES 7-12 Histological aspect of wings in two 8-day (Figs 7, 8 and Figs 9-11) and one 9-day (Fig. 12) embryos whose somitic mesoderm has been X-irradiated respectively at stages of 12, 21 and 18 somite pairs. Figs 7, 8. Wing 6 days after irradiation. Despite the absence of muscle bellies in the forearm (longitudinal section in Fig. 7), tendons developed in the hand (transverse section in Fig. 8). The dorsal EMV tendon, present at the proximo-distal level illustrated in Fig. 8, is vanishing. Note the absence of the hand muscles. Figs 9-11. Longitudinal section through forearm and hand of another experimental muscleless wing 6 days after irradiation, with details of the tendons contained in the distal metacarpal region (Fig. 10) and of the loosening of the mesenchyme in the midst forearm region (Fig. 11). Fig. 12. Illustration of one of the two complete muscles that were present in one wing 7 days after irradiation, probably the extensor medius' longus (EML) muscle. H, Humerus; N, nerve; McII, McIII, metacarpal of digit II, III; R, radius; T. EDC, tendon of extensor digitorum communis muscle; T. EMU, tendon of extensor metacarpi ulnari muscle; T. FDP, tendon of flexor digitorum profundus muscle; T.FDS, tendon of flexor digitorum superficialis muscle; T. UMV, tendon of ulnametacarpalis ventralis muscle; U, ulna. (Scale bar: 200/tm in each case.) 160 M. KIENY AND A. CHEVALLIER r r""i—BiniTiiri— T.FDP Tendon development in embryonic chick wing 161 be interpreted as the joint EMR/EIL tendon (extensor metacarpi radialis/ extensor indicis longus) and the ventral one as the UMV (ulnimetacarpalis ventralis) tendon. Contrary to the earlier developing tendons, no degeneration was observed at their proximal endings. (4) 7 days after irradiation. Five of the six embryos sacrificed at 8 days incubation presented a dorsal apterium that extended over five to nine feather rows. The remaining embryo was normally feathered and its right wing was also normally muscled. In the other five cases the right wings were either completely (four cases) or nearly completely (one case containing an unidentifiable interosseus muscle and the FCU muscle) devoid of muscles, the forearms containing bones, nerves, and rarely muscles within a homogeneously loose mesenchyme Figs 12-14). In the autopods of experimental wings in which a loosening of the subcutaneous mesenchyme was under way, we observed a proximo-distal resorption of the tendons, whose presence was restricted to the phalangeal and metacarpal regions. In their distal half, the tendons remained as healthy compact organs, but in their proximal half, their contours were more and more blurred and the intercellular spaces more and more augmented, in such a way that the tendinous tissues passed progressively into the loose unorganized mesenchyme of the zeugopod. The cellular degeneration (Figs 15-18) that one day earlier was mainly observed at the wrist and zeugopodial levels, had always shifted distally to the metacarpal level, where it extended over a length of ca. 600 jum in a tendon belonging to digits III and IV and over a length of ca. 100 /im in a tendon belonging to digit II (Figs 16, 18). Nevertheless, the distal portion of the tendon that was still in continuity with the overlying integument appeared healthy, and the more the tendon had approached the skeleton proximally, the more its FIGURES 13-18 General view and details of wings in two (Figs 13-15, Figs 16-18) 9-day embryos whose somitic mesoderm has been X-irradiated respectively at stages of 16 and 18 somite pairs. Fig. 13. General view of the interosseus zone of the forearm at the same level than that represented in Fig. 12. The subcutaneous mesenchyme is homogeneously loose. Fig. 14. Detail of the distal forearm region at a distance of 45 /wi from the preceding figure. Note the presence of the dorsal radialis profundus (Nrp) and ventral metacarpalis (Nm) nerves within the loose unorganized mesenchyme. Fig. 15. Cellular degeneration in the proximal portion of a flexor tendon. No phagocytes, but direct elimination of cellular debris (CD) into the blood vessel (BV). Figs 16-18. Slightly oblique section of the hand (Fig. 16) with details of digit II. Cellular degeneration (CD) and some phagocytes (P) are visible in the dorsal joint EMR/EIL tendon (T. EMR/EIL). ANZ, Normal anterior necrotic zone; N, nerve; Me, metacarpal; R, radius; T. FDP, T. FDS and T. UMV, see legend Figs 7-12. U, ulna. (Scale bar: 50 /tm for Figs 15, 18; 100/tm for Figs 14, 17 and 200/tm for Figs 13, 16.) 162 M. KIENY AND A. CHEVALLIER degeneration had intensified. These degenerative processes were never observed in the control tendons. (5) 8 days after irradiation. The resorption of the tendons was nearly completed. One or two tendons belonging to the long digits were still recognizable at the phalangeal and distal metacarpal levels as narrow condensations with indistinct contours. They were not necrotic. The other tendinous tissues had already vanished, so that, at 10 days incubation, the subcutaneous mesenchyme of the whole experimental wing, except the acropod, appeared as homogeneously loose. These vestiges of the tendons will disappear also; previous experiments showed a total absence of tendinous organogenesis 2 days later (Chevallier etal. 1978). DISCUSSION The results presented in this paper offer a clear demonstration of the ability of tendinous blastemae to form and evolve into tendinous structures independently from, and in the absence of, muscle blocks. As in previous experiments (Chevallier et at. 1978), the great reduction of somite-originated myogenic cells affected the development of skeletal muscles in such a way that, in the majority of the cases, no muscle bellies formed or, in a few cases, only one or two of the 12 forearm muscles were present. In the latter condition, the muscle bellies were slightly reduced but normally prolonged by their proximal and distal tendons. When and where the muscles were absent a loose connective tissue extended throughout the zeugopod. Nevertheless, despite the absence of muscle bellies, the differentiation and terminal distribution of six long manus tendons occurred normally at schedule and in space. These tendons, however, were transient structures. Within 3 days after their individuation, they were transformed into loose connective tissue. This process, characterized by an enlargement of intercellular spaces and by cell death, started at the proximal endings of all six tendons, and later shifted distally throughout their length. As concerns the connective network of the muscle belly, it is clear that the mesenchyme of the muscle area segregated from the subcutaneous loose mesenchyme. But, because of the lack of myocytes, the organization of the connective tissue did not take place in the muscle-forming areas, i.e. splitting into distinct masses did not occur and, hence, differentiation of the epimysial envelopes failed. In the rare cases in which some muscular organogenesis occurred, only one or two slightly reduced muscle bellies (with their corresponding tendons) differentiated. According to Shellswell & Wolpert's diagram (1977), these sporadic muscles were derived from the dorsal preaxial radius mass (EMR, interosseus EIL or EML muscles) and from the ventral postaxial ulnar mass (FCU muscle). This hypomorphosis could be interpreted as reflecting a tentative compensation either from the somitic mesoderm situated respectively anteriorly and Tendon development in embryonic chick wing 163 posteriorly to the destroyed portion of somitic mesoderm or from the somatopleural mesoderm. In conclusion, our results are consistent with the view that the development . of the tendons and tendon pattern occurs independently from the development of the muscle bellies. They are in line with Milaire's observation in mice and mole (1963), where tendinous and muscle blastemae start to develop autonomously from one another and join up secondarily. They are also in accord with the findings of Shellswell & Wolpert (1977) which indicate that tendons start to develop in grafted presumptive hands, despite the absence of the forearm. However, for their maintenance and further development tendons require connexion to a muscle belly, seemingly to any one of them, provided it is orientated correctly within the anteroposterior axis, as it was shown in experiments in which the dorso-ventral axis of a wing's distal tip had been inverted (Shellswell & Wolpert, 1977). Under such conditions, ventral tendons matched with dorsal muscles and vice versa. Likewise, experiments that investigated the effect of the dorso-ventral polarity inversion of 3- to 4-day limb-bud ectoderm showed that not only was the differentiation of the distal integument of the autopod in conformity with the polarity of the ectoderm (Pautou & Kieny, 1973; MacCabe, Errick & Saunders, 1974; Pautou, 1917a, b), but also the differentiation of the distal skeletal (Pautou, 1911b) and tendinous (Pautou, unpublished data) tissues were often in conformity with the dorso-ventrality of the ectoderm. Here the curvature of toes and claws suggested that the tip of the tendon was of flexor (ventral) nature and that it was attached to the extensor (dorsal) muscle by the proximal portion of an extensor tendon. Our results thus demonstrate the temporary autonomous development of tendons in the absence of muscle bellies. The reverse situation, namely the persistence of muscle bellies in the absence of distal tendons has been described in oligosyndactylism (Os/ + ) (Kadam, 1962; Milaire, 1962, 1967) or brachypodism (bp/bp) (Griineberg & Lee, 1973). Moreover, histological re-examination of several cases experimentally described by one of us (Kieny, 1964#, b; Kieny & Pautou, 1977, ex. figs 16 and 21) showed that, in the absence of the hand or the foot, the forearm or lower leg muscles were still present and healthy at 10-11 days incubation, although they lacked their distal tendons. Consequently muscle bellies without distal tendons may persist until 11 days at least, whereas distal tendons without muscle bellies have already disappeared. It may be concluded that the absence of connexion between tendon and muscle belly precludes the tendon's maintenance and causes it to vanish. The muscle belly, which comprises a mixed connective and muscular cell population, seemingly is not affected in its development when deprived of its distal tendon. Therefore three cellular compartments may be involved in muscular organogenesis: (1) cells of somatopleural origin which give rise to the connective network of the muscle and become segregated from the subcutaneous mesenchyme in the areas of prospective muscle formation; (2) cells of somatopleural origin which 164 M. KIENY AND A. CHEVALLIER differentiate into tendon blastemae at the extremities of prospective muscle areas and finally form the tendons; (3) cells ofsomitic origin which make up the third compartment; these are the myoblasts which will fuse into myotubes and finally give rise to muscle fibres within the connective network. Muscle bellies may develop independently from tendons, and conversely tendons may, at least temporarily, develop independently from muscle bellies. Whether muscle fibres may become functionally organized in situ in the absence of the connective network is unknown. Finally, since prospective muscle cells are devoid of any region-specific determination (Chevallier et ah 1976, 1977), one can assume that the presumptive connective tissue of the muscular area is responsible for the region-specific muscular pattern formation according to the axes of polarity of the limb. REFERENCES CHEVALLIER, A. (1978). Etude de la migration des cellules somitiques dans le mesoderme somatopleural de l'ebauche de Paile. Wilhelm Roux's Arch, devl Biol. 184, 57-73. CHEVALLTER, A., KIENY, M. & MAUGER, A. (1976). Sur 1'origine de la musculature de 1'aile chez les oiseaux. C. r. hebd. Seanc Acacl. Sci., Paris D 282, 309-311. CHEVALLIER, A., KIENY, M. & MAUGER, A. (1977). Limb-somite relationship: origin of the limb musculature. /. Embryol. exp. Morph. 41, 245-258. CHEVALLIER, A., KIENY, M. & MAUGER, A. (1978). Limb-somite relationship: effect of removal of somitic mesoderm on the wing musculature. /. Embryol. exp. Morph. 43, 263-278. CHEVALLIER, A., KIENY, M. & MAUGER, A. & SENGEL, P. (1977). Developmental fate of the somitic mesoderm in the chick embryo. Symposium on Vertebrate Limb and Somite Morphogenesis, Glasgow, 1st to 4th September 1976, pp. 421-432. Cambridge University Press. CHRIST, B., JACOB, H. J. & JACOB, M. (1974). Ubet den Ursprung der Fliigelmuskulatur. Experimentelle Untersuchungen mit Wachtel- und Huhnerembryonen. Experientia 30, 1446-1449. CHRIST, B., JACOB, H. J. & JACOB, M. (1977a). Experimental analysis of the origin of the wing musculature in avian embryos. Anat. Embryol. 150, 171-186. CHRIST, B., JACOB, H. J. & JACOB, M. (19776). Experimentelle Befunde zur Muskelentwick- lung in den Extremitaten von Huhnerembryonen. Verh. anat. Ges., Jena 71, 1231-1237. GRUNEBERG, H. & LEE, A. J. (1973). The anatomy and development of brachypodism in the mouse. / . Embryol. exp. Morph. 30, 119-141. HAMBURGER, V. & HAMILTON, H. L. (1951). A series cf normal stages in the development of the chick embryo. /. Morph. 88, 49-92. KADAM, K. M. (1962). Genetical studies on the skeleton of the mouse. XXXI. The muscular anatomy of syndactylism and oligosyndactylism. Genet. Res., Camb. 6, 263-283. KIENY, M. (\964a). Regulation des excedents et des deficiences du bourgeon d'aile de l'embryon de Poulet. Archs Anat. microsc. Morph. exp. 53, 29-44. KIENY, M. (1964/?). Etude du mecanisme de la regulation des deficiences dans les chimeres 'aile-patte' et 'patte-aile' 7. Embryol. exp. Morph. 12, 357-371. KIENY, M. &PAUTOU, M. P. (1977). Proximo-distal pattern regulation in deficient avian limb buds. Wilhelm Roux's Arch, devl Biol. 183, 177-191. MACCABE, J. A., ERRICK, J. & SAUNDERS, J. W. Jr. (1974). Ectodermal control of the dorsoventral axis in the leg bud in the chick embryo. Devl Biol. 39, 69-82. MAUGER, A. (1970). Le developpement du plumage dorsal de l'embryon de Poulet etudie a l'aide d'irradiations aux rayons X. Devl Biol. 22, 412-432. MILAIRE, J. (1962). Detection histochimique de modifications des ebauches dan les membres en formation chez la souris oligo-syndactyle. Bull. Acad. r. Med. Belg. 48, 505-528. Tendon development in embryonic chick wing 165 J. (1963). Etude morphologique et cytochimique du developpement des membres chez la souris et chez la taupe. Archs Bioi, Liege 74, 129-317. MILAIRE, J. (1967). Histochemical observations on the developing foot of normal, oligosyndactylous (Os/ + ) and syndactylous (sm/sm) mouse embryos. Archs BioL, Liege 78, 223-288. PAUTOU, M. P. (1977o). Dorso-ventral axis determination of chick limb bud development. Symposium on Vertebrate Limb and Somite Morphogenesis, Glasgow, 1st to 4th September 1976, pp. 257-266. Cambridge University Press. PAUTOU, M. P. (19776). Etablissement de l'axe dorso-ventral dans le pied de l'embryon de poulet. /. Embryol. exp. Morph. 42, 177-194. PAUTOU, M. P. & KIENY, M. (1973). Interaction ecto-mesodermique dans Petablissement de la polarite dorso-ventrale du pied de l'embryon de poulet. C. r. hebd. Seanc. Acad. Sci., Paris D 227, 1225-1228. SHELLSWELL, G. B. & WDLPERT, L. (1977). The pattern of muscle and tendon development in the chick wing. Symposium on Vertebrate Limb and Somite Morphogenesis, Glasgow, 1st to 4th September 1976, pp. 71-86. Cambridge University Press. MILAIRE, (Received 12 July 1978, revised 20 September, 1978)