Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
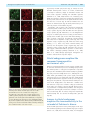
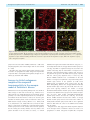
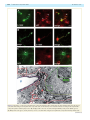
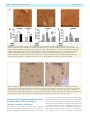
doi:10.1093/brain/awr166 Brain 2011: 134; 2321–2338 | 2321 BRAIN A JOURNAL OF NEUROLOGY Endogenous morphine-like compound immunoreactivity increases in parkinsonism Giselle Charron,1,2 Evelyne Doudnikoff,1,2 Alexis Laux,3 Amandine Berthet,1,2 Gregory Porras,1,2 Marie-Hélène Canron,1,2 Pedro Barroso-Chinea,1,2 Qin Li,4 Chuan Qin,4 Marika Nosten-Bertrand,5 Bruno Giros,5 François Delalande,6 Alain Van Dorsselaer,6 Anne Vital,1,2 Yannick Goumon3 and Erwan Bezard1,2,4 1 2 3 4 5 University of Bordeaux, Institut des Maladies Neurodégénératives, UMR 5293, F-33000 Bordeaux, France CNRS, Institut des Maladies Neurodégénératives, UMR 5293, F-33000 Bordeaux, France Institut National de la Santé et de la Recherche Médicale, INCI, CNRS UPR3212, Strasbourg, France Institute of Laboratory Animal Sciences, Chinese Academy of Medical Science and Peking Union Medical College, Beijing 100021, China Université Pierre et Marie Curie, Institut National de la Santé et de la Recherche Médicale – Centre National de la Recherche Scientifique, CNRS UMRS-952, UMR 7224, Paris, France 6 Laboratoire de spectrométrie de masse BioOrganique, IPHC-DSA, ULP, CNRS, UMR7178, Strasbourg 67087, France Correspondence to: Erwan Bezard, CNRS UMR 5293, 146 rue Léo Saignat, 33076 Bordeaux cedex, France E-mail: [email protected] Morphine is endogenously synthesized in the central nervous system and endogenous dopamine is thought to be necessary for endogenous morphine formation. As Parkinson’s disease results from the loss of dopamine and is associated with central pain, we considered how endogenous morphine is regulated in the untreated and L-DOPA-treated parkinsonian brain. However, as the cellular origin and overall distribution of endogenous morphine remains obscure in the pathological adult brain, we first characterized the distribution of endogenous morphine-like compound immunoreactive cells in the rat striatum. We then studied changes in the endogenous morphine-like compound immunoreactivity of medium spiny neurons in normal, Parkinson’s disease-like and L-DOPA-treated Parkinson’s disease-like conditions in experimental (rat and monkey) and human Parkinson’s disease. Our results reveal an unexpected dramatic upregulation of neuronal endogenous morphine-like compound immunoreactivity and levels in experimental and human Parkinson’s disease, only partially normalized by L-DOPA treatment. Our data suggest that endogenous morphine formation is more complex than originally proposed and that the parkinsonian brain experiences a dramatic upregulation of endogenous morphine immunoreactivity. The functional consequences of such endogenous morphine upregulation are as yet unknown, but based upon the current knowledge of morphine signalling, we hypothesize that it is involved in fatigue, depression and pain symptoms experienced by patients with Parkinson’s disease. Keywords: immunohistochemistry; medium spiny neuron; astrocyte; dopamine; mouse; rat; monkey; endogenous morphine; human Abbreviations: DAT = dopamine transporter; MPTP = 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine Received January 30, 2011. Revised June 9, 2011. Accepted June 12, 2011. Advance Access publication July 8, 2011 ß The Author (2011). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For Permissions, please email: [email protected] 2322 | Brain 2011: 134; 2321–2338 Introduction It is now soundly established that morphine is endogenously synthesized in the CNS of vertebrates (Gintzler et al., 1978; Stefano et al., 1996, 2000; Muller et al., 2008; Goumon et al., 2009). However, its biosynthesis was debated until a recent report using mice that are unable to synthesize dopamine due to a genetic deletion of tyrosine hydroxylase specifically in dopaminergic neurons, showed that endogenous dopamine is necessary for endogenous morphine formation in normal mammalian brain (Neri et al., 2008). Those dopamine-null mice displayed basal hyperalgesia, further supporting a role for dopamine in endogenous morphine biosynthesis (Hnasko et al., 2005). The neurological disorder classically associated with dopamine depletion is Parkinson’s disease (Hornykiewicz, 1966). Apart from parkinsonian motor symptoms, Parkinson’s disease is less widely appreciated as a disease that causes a large variety of pain syndromes, with a prevalence of 40% (Chaudhuri and Schapira, 2009). Pain syndromes are attenuated by the most effective symptomatic therapy represented by the dopamine precursor, L-3,4-dihydroxyphenylalanine (L-DOPA), suggesting that the dysfunction may occur in dopamine-dependent centres (Schestatsky et al., 2007; Ford, 2010). For instance, the pain threshold for cold is substantially lower in patients with Parkinson’s disease who were withdrawn from treatment compared with controls, and is then normalized after L-DOPA therapy (Brefel-Courbon et al., 2005). Patients with Parkinson’s disease with primary central pain displaying hyperalgesia and the absence of habituation of sympathetic sudomotor response to repetitive pain are improved by L-DOPA treatment (Schestatsky et al., 2007). Although endogenous morphine distribution has been studied in the normal brain (Gintzler et al., 1978; Bianchi et al., 1993; Muller et al., 2008; Laux et al., 2011), its cellular origin and distribution remains obscure in parkinsonism despite earlier findings reporting morphine level modulation in urine of patients with Parkinson’s disease OFF and ON medication (Matsubara et al., 1992). We therefore first characterized the endogenous morphine-like compound immunoreactive cell distribution in the rat striatum and second, studied the changes in the endogenous morphine-like compound immunoreactivity of the medium spiny neurons in normal, Parkinson’s disease-like and L-DOPA-treated Parkinson’s disease-like conditions in experimental and human Parkinson’s disease. Our results reveal an unexpected dramatic regulation of neuronal and glial endogenous morphine immunoreactivity in experimental and human Parkinson’s disease, somewhat normalized by L-DOPA treatment. Materials and methods Rodent experiments Experiments were performed in accordance with French (87-848, Ministère de l’Agriculture et de la Forêt) and European Communities Council Directive of 24 November 1986 (86/609/EEC) for care of laboratory animals and were approved by the Ethical Committee of Centre National de la Recherche Scientifique, Région Aquitaine. G. Charron et al. Wild-type rats Twelve adult male Sprague Dawley rats (Charles River Laboratories) weighing 175–200 g at the beginning of the experiment were used. On Day 0 of the protocol, unilateral dopamine deprivation of the striatum was obtained by 6-hydroxydopamine (3 mg/ml) injection in the right medial forebrain bundle (2.5 ml at anteroposterior = 3.7 mm, mediolaterial = + 1.7 mm and dorsoventral = 8 mm, relative to bregma) as previously described (Schuster et al., 2008, 2009; Berthet et al., 2009). Animals displaying a 495% loss of tyrosine hydroxylase-immunopositive fibres in the striatum (Schuster et al., 2008, 2009; Berthet et al., 2009), as assessed after completion of all experiments (not shown), were retained for final analysis. From Day 21 to Day 30 post-surgery, rats were treated once daily with benserazide [15 mg/kg, intraperitoneally (i.p.)] and either vehicle (n = 4) or L-DOPA (6 mg/kg, i.p.) (n = 4). The benserazide and L-DOPA treated mice were dyskinetic at the time of death. On Day 30, all animals received the last vehicle injection L-DOPA and were sacrificed 60 min later. Wild-type mice Experiments were performed on 21 45-day-old laboratory-bred adult male C57BL/6 mice weighting 30 3 g. Dopamine depletion was achieved by injecting either saline (controls) or a combination of 2.5 mg/kg reserpine i.p. (Sigma Aldrich) at 24 h and 100 mg/kg of -methyl-p-tyrosine (Sigma Aldrich) 24, 16, 4 and 1 h before sacrifice, as previously described (Thomas et al., 2008). Thirteen mice were used for neurochemistry and eight for immunohistochemistry. Dopamine transporter knock-out mice Dopamine transporter (DAT) mutant mice were generated by in vivo homologous recombination as previously described (Giros et al., 1996). Twelve female mice between the ages of 2 and 4 months were used for immunohistochemistry: four wild-type mice (DAT + / + ), four heterozygous mice (DAT / + ) and four homozygous mice (DAT / ). Tissue preparation Terminal procedure was identical for mice and rats as previously described (Berthet et al., 2009). Animals were deeply anaesthetized with chloral hydrate (VWR), 150 mg/kg (i.p.). Brains of saline and reserpine-treated mice for neurochemical experiments were collected fresh, freeze–thawed in isopentane and immediately stored at 80 C until further use. All other animals were transcardially perfused with a mixture of 2% paraformaldehyde and 0.2% glutaradehyde in 0.1 M phosphate-buffer at pH 7.4. Brains were removed and post-fixed overnight in 2% paraformaldehyde at 4 C. Coronal sections were cut at 60 mm on a Vibratome (Leica, VT 1000 S) and collected in phosphatebuffered saline (pH 7.4). To enhance the penetration of immunoreagents, the sections were equilibrated in a cryoprotectant solution, phosphate-buffered saline with 25% saccharose, freeze–thawed in isopentane and stored in phosphate-buffered saline with 0.03% sodium azide at 4 C until use. Non-human primate experiments All experiments were carried out in accordance with the European Communities Council Directive of 24 November 1986 (86/609/EEC) for care of laboratory animals in an AAALAC accredited facility and were approved by the Institute of Lab Animal Science IACUC. Animals were housed in individual primate cages under controlled conditions of humidity (50% 5%), temperature (24 1 C) and light Endogenous morphine increases in Parkinsonism Brain 2011: 134; 2321–2338 | 2323 Table 1 List of antibodies Antibody Species Dilution Source Morphine D1 dopamine receptor D2 dopamine receptor Glutamate acid decarboxylase 65-67 Dopamine and cAMP-regulated phosphoprotein, 32 kDa S100 NG2 Glial fibrillary acidic protein CYP2D1 Tyrosine hydroxylase Choline acetyltransferase Parvalbumin Mouse Rat Rabbit Rabbit Rabbit Rabbit Rabbit Mouse Rabbit Rabbit Goat Guinea pig 1:2000 1:500 1:500 1:1000 1:500 1:1000 1:500 1:1000 1:1000 1:2000 1:200 1:1000 Aviva Systems Biology Sigma Santa Cruz Millipore Cell Signalling Abnova Millipore Millipore ENZO Life Sciences Incstar Millipore Synaptic System (12 h light/12 h dark cycle, time lights on 08:00 a.m.), food and water were available ad libitum and animal care supervised by veterinarians skilled in the healthcare and maintenance of non-human primates. The six female cynomolgus monkeys (Macaca fascicularis, Xierxin) involved in this study have been previously presented (Guigoni et al., 2005, 2007; Nadjar et al., 2006). Briefly, two macaques were left untreated (control group); four were administered with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) hydrochloride. Once a bilateral parkinsonian syndrome had stabilized (i.e. unchanged disability score over several weeks), two macaques were kept without any dopaminergic supplementation (MPTP-lesioned group) and two macaques were treated chronically with twice daily administration of levodopa (Modopar; L-DOPA/carbidopa, ratio 4:1) for 6–8 months at a tailored dose designed to produce a full reversal of parkinsonian condition (L-DOPA-treated MPTP-lesioned group) as previously described (Guigoni et al., 2005, 2007; Nadjar et al., 2006). The L-DOPA-treated MPTP-lesioned group were dyskinetic at the time of death. Animals were deeply anaesthetized with sodium chloral hydrate (150 mg/kg) 1 h after the last vehicle/L-DOPA dose and perfused transcardially with a mixture of 2% paraformaldehyde and 0.2% glutaradehyde in 0.1 M phosphate buffer. Brains were removed, bisected along the midline, stored in 2% paraformaldehyde overnight and cut into 60 mm frontal sections with a vibratome (Leica, VT1000S). Sections were collected in phosphate-buffered saline, cryoprotected in phosphate-buffered saline with 25% saccharose, freeze–thawed in isopentane and stored in phosphate-buffered saline with 0.03% sodium azide until use. The clinical assessments and the characterization of the extent of nigrostriatal denervation have been previously published (Guigoni et al., 2005, 2007; Nadjar et al., 2006) in order to ensure that all the MPTP-treated animals displayed comparable lesion of the nigrostriatal pathway. Indeed, the number of tyrosinehydroxylase immunopositive neurons in the substantia nigra pars compacta and the striatal tyrosine-hydroxylase immunostaining were identical between the two MPTP-treated groups and dramatically low compared with control animals (loss of terminals 495%) as previously shown (Guigoni et al., 2005). Post-mortem human samples The observations on human tissue were based on the analysis of formalin-fixed and paraffin-embedded human specimens from an archival collection (Comite Protection des Personnes N CEBH 2009/ 03; Ministere Enseignement Superieur et Recherche: DC-2008-337) declared and approved by the ethics committee (Comité de Protection des Personnes Sud-Ouest et Outre Mer III) of Bordeaux University Hospital. Striatum material was available for both neuropathologically confirmed cases of Parkinson’s disease (n = 3; two males, one female; 62 7 years old at death) and controls with no detectable CNS disease (n = 3; two males, one female; 63 8.5 years old at death). The post-mortem delay was similar for all cases (524 h) but the delay for the female Parkinson’s disease case was 60 h. The striatum was studied within a coronal 5-mm thick section demonstrating connections between the head of the caudate and the putamen across the internal capsule. Immunocytochemical procedures Immunofluorescent single labelling Vibratome-cut, free-floating sections were rinsed in 0.1 M phosphatebuffered saline (pH 7.4) and incubated in 0.3% H2O2/phosphatebuffered saline to inhibit endogenous peroxidases. After preincubation in 3% normal goat serum, 3% bovine serum albumin in phosphatebuffered saline to minimize non-specific labelling, the tissue sections were incubated with the primary antibody (diluted in phosphatebuffered saline 0.1 M, 3% bovine serum albumin, 3% normal goat serum) overnight at room temperature. The nature, origin and dilution of the primary antibodies are listed in Table 1. A goat anti-rabbit IgG conjugated to alexa 488 fluorochrome (Molecular Probes) or a goat anti-mouse IgG conjugated to alexa 568 fluorochrome (Molecular Probes) were used as secondary antibodies (1:400). Sections were then washed in phosphate-buffered saline and mounted in Vectashield mounting medium with DAPI (Vector Laboratories) and examined using a Zeiss Axioplan 2 fluorescent microscope. Fluorescence was visualized at 488 nm (green), 568 nm (red) and 350 nm (blue). The specificity of the anti-endogenous morphine-like compound antibody has been reported elsewhere (Muller et al., 2008; Glattard et al., 2010; Laux et al., 2011). When sections were incubated without the primary antibody, no labelling was observed. Similarly, incubation of endogenous morphine primary antibody with an excess of morphine, completely abolished staining, confirming the specificity for endogenous morphine (Fig. 1). Immunofluorescent double-labelling To further characterize the striatal endogenous morphine-like compound positive cells, we performed a double immunohistochemistry 2324 | Brain 2011: 134; 2321–2338 G. Charron et al. Figure 1 Distribution of endogenous morphine-like compound immunoreactive structures. (A) Sagittal section of rat brain stained with anti-endogenous morphine antibody showing main endogenous morphine-like compound positive midbrain and forebrain regions. Scale bar = 3 mm. (B) Absence of staining when anti-endogenous morphine antibody was preadsorbed with the immunizing alkaloid before incubation. (C–J) Magnifications of selected endogenous morphine-like compound positive regions boxed in A: cortex (C), caude-putamen (D), Calleja islets (E), substantia nigra pars reticulata (F), pons (G), interposed cerebellar nuclei (H), cerebellum (I), inferior colliculus (J); scale bar = 50 mm. (K–L) Immunocytochemical localization of endogenous morphine-like compounds on striatal coronal section. Inset in L shows that both weakly (open circle) and strongly (asterisk) immunoreactive neurons are present as well as glial cells (arrow). (M) Control immunohistochemistry with omission of primary antibody. Scale bar = 2 mm (K), 25 mm (L–M), 10 mm (L inset). procedure against endogenous morphine and against: (i) markers of medium spiny neurons (dopamine and cAMP-regulated phosphoprotein, 32 kDa, D1 dopamine receptor, D2 dopamine receptor and glutamate acid decarboxylase 65/67); (ii) the rate-limiting enzyme for endogenous morphine synthesis, CYP2D1; and (iii) markers of interneurons, (choline acetyltransferase and parvalbumin). Immunohistochemical procedure was as described above except that two primary antibodies derived from different animal species were applied together. Similarly, two secondary antibodies raised in different species and labelled with two different fluorochromes, alexa 488 conjugated against rabbit and alexa 568 against mouse, were applied. Immunoperoxidase labelling in experimental models The number of striatal endogenous morphine-like compound immunopositive neurons was defined after immunolabelling with the peroxidase technique. Brain sections were incubated overnight in endogenous Endogenous morphine increases in Parkinsonism morphine primary antibody, as described above. Sections were first incubated in biotinylated secondary antibody and then in avidin– biotin complex (VectastainÕ Elite ABC kit) prepared according to the manufacturer’s recommendations. Morphine immunoreactivity was revealed with NovaREDTM kit (Vector Laboratories). After washing with phosphate-buffered saline, sections were mounted on microscope slides in Eukitt (Merck). Immunoperoxidase labelling on post-mortem human samples Four micrometre-thick paraffin-embedded striatal sections were processed as previously described (Vital et al., 2009). Briefly sections were dewaxed and pressure cooked in sodium citrate buffer (0.01 M, pH 6.0) for antigen retrieval. The sections were washed, blocked with a universal blocking buffer (Biogenex) and incubated with endogenous morphine antibody overnight. Control sections were incubated with blocked antiserum. After transfer in 0.3% H2O2/phosphate-buffered saline to block the endogenous peroxidase reaction, the sections were treated with biotinylated anti-rabbit IgG (diluted 1:200 in phosphate-buffered saline) and incubated with VectastainÕ Elite ABC reagent. The colour was developed using NovaREDTM kit. Sections were counterstained with Mayer’s Hemalun and mounted with an aqueous agent for microscopy AquatexÕ (Merck). Immunogold labelling Subcellular endogenous morphine-like compound distribution was analysed at electron microscopic level in the rat striatum. The endogenous morphine was detected by the pre-embedding immunogold technique as previously described (Dumartin et al., 1998, 2000; Guigoni et al., 2007; Berthet et al., 2009). Sections were incubated in 4% normal donkey serum for 45 min and then in a mixture of endogenous morphine antibody (1:2000 in phosphate-buffered saline supplemented with 1% normal donkey serum) overnight at room temperature. After washing in phosphate-buffered saline and PBS-BSAcTM (Aurion), the sections were incubated for 3 h at room temperature in donkey anti-mouse IgGs conjugated to ultra-small gold particles (0.8 nm; Aurion) diluted 1:100 in PBS-BSAcTM gel. After post-fixation in 1% glutaraldehyde in phosphate-buffered saline and several washes, the immunogold signal was intensified using a silver enhancement kit (HQ silverTM; Nanoprobes) for 6 min at room temperature in the dark. The sections were then post-fixed in 0.5% osmium tetroxide and dehydrated in ascending series of ethanol dilutions that also included 70% ethanol containing 1% uranyl acetate. The sections were post-fixed, dehydrated and embedded in resin (Durcupan ACM; Fluka). Serial ultra-thin sections were cut with a Reichert Ultracut S, contrasted with lead citrate and examined with Hitachi H-7650 microscope. Image analysis Fluorescent images were captured on a confocal Leica microscope DM 2500 TCS SPE. Digital images were acquired separately for each wavelength [488 nm (green) and 568 nm (red)] and then merged. Adobe Photoshop CS3 was used to further process digital images. Any adjustments to brightness and contrast were made uniformly to all parts of the image. Cell counting was performed using a computerized image analysis system (Mercator V6.50, Explora Nova) linked to a Leica microscope type DM 6000B ( 20 objective). The number of immunoreactive medium spiny neurons (round or oval shape with a cell body diameter of 10–15 mm) was counted for two populations and classed according Brain 2011: 134; 2321–2338 | 2325 to labelling intensity, from weak ( + ) to strong labelling ( + + + ) (Fig. 1L). Quantitative analysis was carried out on the whole striatum area and results were expressed as an average density of endogenous morphine cells per mm2. To minimize the inherent variability in the immunochemical procedures, comparisons were made in tissue sections that were processed simultaneously. Only those neurons that were on the surface of the sections, where antibody penetration was guaranteed, were counted. Neurochemical procedures Dopamine quantification by reverse phase high-performance liquid chromatography Mouse brains were homogenized in 1 ml of 1 M ClHO4 and centrifuged for 30 min at 14 000 g. Supernatants were analysed using an Äkta PurifierTM high-performance liquid chromatography system (GE Healthcare Bioscience) with reverse phase column (250/4 Nucleosil 100-5-C18HD; Macherey Nagel). Buffer A was 5 mM H3PO4, 5 mM hexanesulphonic acid in water. Buffer B was 100% acetonitrile. Detection of dopamine was monitored with a fluorescence detector (RF-10AXL; Shimadzu; excitation at 270 nm and emission at 320 nm) as previously described (Muzzi et al., 2008). Morphine-specific enzyme-linked immunosorbent assay Homogenized brains were sonicated at 4 C (3 10 s) in phosphatebuffered saline. Extracts were centrifuged (30 min, 10 000 g at 4 C). Forty microlitres of supernatants (tested in duplicates) were used for enzyme-linked immunosorbent assay analysis (Glattard et al., 2010). The morphine-specific enzyme-linked immunosorbent assay kit from Immunalysis Corporation (provided by AgriYork 400 Limited) was used for the quantification of morphine present in brain tissue extracts (control mice, n = 4; -methyl-p-tyrosine–reserpine mice, n = 5) (Muller et al., 2008). The specificity of the test for morphine was confirmed by testing different amounts of dopamine, adrenaline, noradrenaline, norlaudanosoline/tetrahydropapaveroline, morphine, morphine-6-glucuronide, morphine-3-glucuronide and codeine (0.01– 25 ng/ml, data not shown) (Muller et al., 2008; Glattard et al., 2010). For all tests, morphine standards were diluted in the appropriate buffer. Samples with a higher coefficient of variation value were retested to obtain a coefficient of variation 48%. The calculated methodological detection limit of the batch of enzyme-linked immunosorbent assay kit used for the study was 0.01 ng/ml of morphine. Morphine-like compound extraction from mouse brain and striatum Mouse brain and striatum were homogenized in 1 ml of methanol. After centrifugation (14 000 g, 20 min at 4 C), the supernatant was recovered and dried with a SpeedVac evaporator. The sample was suspended in 300 ml of urea buffer (1 M, pH 8.4) and submitted to heat treatment (100 C, 30 min). After a centrifugation (14 000 g, 20 min at 4 C), alkaloids present in the supernatant were purified into OasisTM HLB 96-well plates (60 mm, 60 mg, Waters) that had been preconditioned with 500 ml of methanol, followed by 500 ml (NH4)2CO3 (10 mM, pH 8.8). Samples were loaded onto the OasisTM HLB plate and the SPE was rinsed with 500 ml (NH4)2CO3 (10 mM, pH 8.8). Alkaloids were eluted with 300 ml of acetonitrile/ isopropanol/formic acid (40/59/1 v:v:v). Extracted samples were dried prior to liquid chromatography-tandem mass spectrometry analysis. 2326 | Brain 2011: 134; 2321–2338 High-performance liquid chromatography-tandem mass spectrometry instrumentation and analytical conditions Liquid chromatography separations were carried out with an Agilent LC 1100 binary pump, autosampler, vacuum degasser and column oven coupled with an Agilent 6410 Triple Quad LC/MS (Agilent Technologies). The dry samples were dissolved in 10 ml acetonitrile (70%), vortex-mixed for 1 min and injected onto an acrylamido-type column (TSKgel Amide-80, TOSOH) at 25 C. The solvent system consisted of 100% water, 0.15% formic acid and 5 mM ammonium acetate (solvent A) and 100% acetonitrile (solvent B). Elution was performed at a flow rate of 220 ml/min with a 70–40% linear gradient (solvent B) over the first 8 min, followed by an 80% stage (solvent B) over 2 min before the reconditioning of the column at 70% of solvent B. The system was fully controlled by MassHunter software (Agilent Technologies). Electrospray ionization was achieved in the positive mode with the spray voltage set at 4000 V. Nitrogen was used as nebulizer gas and nebulizer pressure was set at 20 psi with a source temperature of 100 C. Desolvation gas (nitrogen) was heated to 350 C and delivered at a flow rate of 10 l/min. Qualification was performed in selected reaction monitoring mode with the following transitions: morphine, m/z (mass-to-charge ratio) 286.1–165.1, and codeine, m/z 300.2–214.8, and m/z 300.2–164.8. G. Charron et al. Table 2 List of endogenous morphine-like compound immunoreactive brain areas Brain area Immunoreactivity Cerebral cortex Caudate putamen Accumbens nucleus Islands of Calleja Substantia nigra pars reticulata Subthalamus nucleus Hippocampus Red nucleus Interstitial nucleus of Cajal Darkschwitsh nucleus Inferior colliculus Superior colliculus Pontine nuclei Cerebellum (Purkinje layer) Cerebellum (molecular layer) Interposed cerebellar nuclei Vestibular nucleus (lateral) + + + + + + + + + + + + + + + + + + + + ++ + + ++ ++ ++ ++ ++ + Immunostaining intensity was visually scored as follows: + weak; + + moderate; + + + strong. Statistical analysis Comparisons between groups were carried out using either one- or two-factor (ANOVA) followed by Bonferroni post hoc test. Unpaired t-test was used for monkey experiments in which a low n was used or for comparisons between two groups. The level of statistical significance was set at P 5 0.05. Results Endogenous morphine-like compound immunoreactivity in the rat brain We first characterized the distribution of the endogenous morphine-like compound immunoreactivity (morphine, morphine-6-glucuronide, morphine-3-glucuronide and codeine) on sagittal rat brain sections (Fig. 1A and B). The endogenous morphine-like compound staining was heterogeneously distributed between brain areas (Fig. 1C–J; Table 2). The cortex (Fig. 1C), the caude-putamen (Fig. 1D) and the substantia nigra pars reticulata (Fig. 1F) are clearly stained, whereas the islands of Calleja (Fig. 1E), the pons (Fig. 1G), the interposed cerebellar nuclei (Fig. 1H), the cerebellum (Fig. 1I) and the inferior colliculus (Fig. 1J) were strongly stained. Interestingly, most of these nuclei play a role in motor control. Phenotype of striatal endogenous morphine-like compound immunopositive neurons The striatum plays a major role in the pathophysiology of Parkinson’s disease, being the primary target of nigrostriatal dopamine neurons (Parent et al., 1995; Prensa and Parent, 2001). We therefore focused our attention on the striatum that displays endogenous morphine-like compound immunopositivity (Fig. 1D and K) in both neuronal and non-neuronal labelling (Fig. 1L). Two types of endogenous morphine-like compound positive neurons were observed in the rat striatum: the vast majority is constituted by the oval-shaped medium spiny neurons (15 mm in size) while a few larger neurons (20 mm) were also labelled. In the medium spiny neurons, endogenous morphine-like compound immunostaining was present in the cytoplasm and in proximal processes (Fig. 2A–D). Endogenous morphine-like compound immunolabelling was observed in 45% of dopamine and cAMPregulated phosphoprotein, 32 kDa-immunopositive medium spiny neurons (Fig. 2A). Those endogenous morphine-like compound immunoreactive medium spiny neuron-like neurons were glutamate acid decarboxylase positive (Fig. 2B) and either D1 dopamine receptor positive (Fig. 2C) or D2 dopamine receptor positive (Fig. 2D). We investigated if these medium spiny neurons also expressed CYP2D1, the rat enzyme homologue of human CYP2D6 that is considered a key enzyme in endogenous morphine synthesis (Boettcher et al., 2005; Stefano et al., 2008). Varying degrees of CYP2D1 immunoreactivity were found but of the majority of endogenous morphine-like compound positive, medium spiny neurons were also CYP2D1-positive (Fig. 2E). Finally, we defined the phenotype of the larger neurons using either choline acetyltransferase or parvalbumin immunohistochemistry (Parent et al., 1995). None of the choline acetyltransferase-immunopositive neurons were endogenous morphine positive (Fig. 2F), while the parvalbumin-immunopositive neurons displayed a clear endogenous morphine immunopositivity (Fig. 2G). The electron microscopy analysis confirmed the light microscopy observations, i.e. that endogenous morphine-like compounds are Endogenous morphine increases in Parkinsonism Brain 2011: 134; 2321–2338 | 2327 expressed by medium spiny neurons (Fig. 3). Indeed, the most abundant immunopositive neurons are circular, medium-sized (10–15 mm in diameter), display an unindented nucleus and scant somatic cytoplasm containing few subcellular organelles (Dumartin et al., 1998, 2000; Berthet et al., 2009) (Fig. 3A). Within the cytoplasm, endogenous morphine-like immunoreactivity was present at the inner face of the plasma membrane and at the outer face of vesicles (Fig. 3A and B). Dendrites were also found to be immunoreactive (Fig. 3C and D), with reaction products being located on microtubules (Fig. 3C). Interestingly, only one-third of the endogenous morphine-like compound positive medium spiny neurons were D1 dopamine receptor positive (Fig. 4A), while two-thirds were D2 dopamine receptor positive (Fig. 4B). Furthermore, one can distinguish two categories of medium spiny neurons according to their immunoreactivity. Most medium spiny neurons with weak labelling ( + ), where moderate endogenous morphine-like immunoreactivity was found, were D1 dopamine receptor positive (Fig. 4A) or D2 dopamine receptor negative (Fig. 4B), while those with strong labelling ( + + + ), where very intense immunoreactivity was observed, were D1 dopamine receptor negative (Fig. 4A) or D2 dopamine receptor positive (Fig. 4B). Altogether, these observations suggest a differential influence upon the so-called ‘direct’ and ‘indirect’ striatofugal pathways (Alexander et al., 1986; Albin et al., 1989) although physiological investigations are needed to characterize the respective impact. Striatal endogenous morphine-like compound immunopositive non-neuronal cells Figure 2 Phenotype of endogenous morphine-like compound immunopositive striatal medium spiny neurons. Double immunofluorescence was conducted for endogenous morphine-like compounds (eM, red) and dopamine and cAMP-regulated phosphoprotein, 32 kDa (DARPP-32, A), glutamate acid decarboxylase 65-67 (GAD, B), D1 dopamine receptor (D1R, C), D2 dopamine receptor (D2R, D), CYP2D1 (E), choline acetyltransferase (ChAT, F) and parvalbumin (PV, G) (all in green). Arrows and asterisks indicate immunopositivity for the considered marker. Scale bars = 10 mm. Besides clear immunoreactivity in neurons, stellate cells with smaller cell bodies and numerous processes resembling astrocytes were also immunopositive for endogenous morphine-like compounds. Astrocytes were identified by their immunoreactivity for glial fibrillary acidic protein and S100b (Kacem et al., 1998; Raponi et al., 2007). While glial fibrillary acidic protein signal preferentially stained astrocyte processes rather than cell bodies, S100b staining strongly labelled both the cytoplasm and the processes (Fig. 5A). Endogenous morphine-like compound immunoreactivity was detected in the S100b-positive cells, which have a ramified form, their numerous cell processes sometimes coiling up around blood vessels and in close vicinity of neurons (Fig. 5B). Electron microscopy localization of S100b (immunoperoxidase) and endogenous morphine-like (immunogold) revealed clear examples of such triadic associations (Fig. 5D). In contrast, endogenous morphine-like immunoreactivity was not observed in NG2-positive glial cells (Fig. 5C). Increase in striatal endogenous morphine-like immunoreactivity in the rat model of Parkinson’s disease As the medium spiny neurons are the primary targets of the dopamine terminals originating from the substantia nigra pars compacta (Parent et al., 1995; Prensa and Parent, 2001), we investigated 2328 | Brain 2011: 134; 2321–2338 G. Charron et al. Figure 3 Immunogold detection of endogenous morphine-like compound at electron microscopic level in the rat striatal medium spiny neurons. (A) An endogenous morphine-like compound immunoreactive soma with the size and cytological features of a medium spiny neuron. Gold particles are associated with the outer face of vesicles and with the inner face of the plasma membrane. (B) Detail from a Golgi apparatus. Gold particles are associated with vesicles at the outer face. (C) An endogenous morphine-like compound immunoreactive dendrite. The inset shows a higher magnification with gold particles located on microtubules. (D) An axodendritic synapse (arrow) between an endogenous morphine-like compound labelled dendrite (D2) and an unlabeled axon terminal (A1). Scale bar = 1 mm (A); 0.5 mm (B and C); 0.2 mm (D); 0.2 mm (C inset). the change in endogenous morphine-like compound immunoreactivity of these neurons in the three experimental conditions. The pattern of distribution of endogenous morphine-like compound immunopositive elements varied in number and in staining intensity (Fig. 6A). We distinguished two categories of medium spiny neurons according to their immunoreactivity as defined in Fig. 4: those with weak labelling ( + ), where moderate endogenous morphine-like compound immunoreactivity was found, and those with a strong labelling ( + + + ), where very intense immunoreactivity was observed. A qualitative analysis reveals that in the striatum of sham animals, weakly labelled ( + ) medium spiny neurons were observed among a very lightly immunoreactive neuropil (Fig. 6A). Lesioning the nigrostriatal pathway in 6-hydroxydopamine rats led to an increased immunostaining with an obvious increase in the number of strongly labelled ( + + + ) medium spiny neurons (Fig. 6A) and of the neuropil. While such an increase was more pronounced on the lesioned side, it was also present on the unlesioned side (Fig. 6A). L-DOPA treatment of 6-hydroxydopaminelesioned rats led to a decrease in intensity of neuronal and neuropil immunostaining on both sides (Fig. 6A), without however, returning to the immunostaining observed in sham animals. Quantitative analysis shows that the total number of immunopositive medium spiny neurons is not statistically different after unilateral 6-hydroxydopamine lesion and/or further L-DOPA treatment (Fig. 6B). This result suggests that (i) the overall number of endogenous morphine-like compound immunopositive medium spiny neurons does not change across experimental conditions; and (ii) lesioning one hemisphere affects endogenous morphinelike compound immunoreactivity in both hemispheres. When distinguishing between the strongly labelled and weakly labelled medium spiny neurons (Fig. 6C), these two categories show opposing variations. The number of strongly labelled medium spiny neurons dramatically increases after 6-hydroxydopamine lesion (P 5 0.05), on both sides of the brain, although the magnitude is greater on the lesioned side (Fig. 6C). The number of weakly labelled medium spiny neurons decreases on the lesioned side of 6-hydroxydopamine-lesioned rats (P 5 0.05; Fig. 6C). On the lesioned side, the number of strongly labelled medium Endogenous morphine increases in Parkinsonism Brain 2011: 134; 2321–2338 | 2329 Figure 4 Differential endogenous morphine-like compound labelling of striatal medium spiny neurons based upon their dopamine receptor immunoreactivity. (A) D1 dopamine receptor immunoreactivity (arrows) is located on weakly endogenous morphine-positive medium spiny neurons ( + ). (B) Only strongly endogenous morphine-like compound positive medium spiny neurons ( + + + ) express D2 dopamine receptor immunoreactivity (arrowheads). Scale bar = 10 mm. D1R = D1 dopamine receptor; D2R = D2 dopamine receptor; eM = endogenous morphine. spiny neurons decreased after L-DOPA treatment (P 5 0.05 versus 6-hydroxydopamine) but remained higher than in sham animals (P 5 0.05). Altogether these data suggest that dopamine depletion in the rat leads to an upregulation of striatal endogenous morphine-like compounds that is subsequently downregulated, though not normalized, by treatment with L-DOPA. Increase in striatal endogenous morphine-like compound immunoreactivity in the macaque model of Parkinson’s disease Unilateral lesion of the nigrostriatal pathway in the rat is known to induce changes on both sides of the brain (e.g. Johansson et al., 2001; Konradi et al., 2004) although the most prominent changes generally occur on the dopamine-depleted side. As a clinically relevant bilateral model was therefore needed, we used the goldstandard non-human primate model of Parkinson’s disease, the MPTP-treated macaque monkey (Bezard et al., 2003a; Gold et al., 2007; Berton et al., 2009; Ahmed et al., 2010). The distribution of endogenous morphine-like compound immunoreactive medium spiny neurons in the caudate nucleus and the putamen, the two forming the monkey striatum, was very similar to the pattern described in the rat model (see above), i.e. uniform distribution throughout the striatum and with two categories, i.e. the weakly labelled and the strongly labelled medium spiny neurons (Fig. 7A). While no significant change was observed in the caudate nucleus (Fig. 7B), the total number of endogenous morphine-like compound immunopositive medium spiny neurons in the putamen was greater in the MPTP-lesioned group compared with the control group (unpaired t-test, P 5 0.01; Fig. 7B). Chronic L-DOPA treatment led to a normalization of the total number of endogenous morphine-like compound immunopositive medium spiny neurons in the putamen (P 5 0.05 versus MPTP-lesioned; non-significant versus control group; Fig. 7B). When distinguishing between the strongly labelled and weakly labelled medium spiny neurons (Fig. 7C and D), these two categories show opposing variations. The number of strongly labelled-like-immunolabelled medium spiny neurons dramatically increased in the MPTP-lesioned group in both caudate and putamen (P 5 0.01), while weakly labelled-like-immunolabelled medium spiny neurons decreased (P 5 0.05; Fig. 7C and D). Chronic L-DOPA treatment decreased the number of strongly labelled medium spiny neurons and increased the number of weakly labelled medium spiny neurons with no more significant difference with either the control or the MPTP-lesioned animals (Fig. 7C and D). Altogether these data suggest that striatal dopamine depletion in the macaque model of Parkinson’s disease also leads to an upregulation of striatal endogenous morphine-like compounds 2330 | Brain 2011: 134; 2321–2338 G. Charron et al. Figure 5 Phenotype of endogenous morphine-like compound immunopositive striatal glial cells. (A) S100b-positive glial cells (green) co-localize with endogenous morphine-like compounds (red) as indicated by arrow. Asterisk indicates an endogenous morphine-like compound positive medium spiny neuron. (B) S100b-positive cell (red) colocalizes with glial fibrillary acidic protein (GFAP, green). (C) NG2-postive cells (green, arrow), however, do not co-localize with endogenous morphine-like compound immunopositivity (red). (continued) Endogenous morphine increases in Parkinsonism Brain 2011: 134; 2321–2338 | 2331 Figure 6 Dopamine depletion increases the number of strongly labelled endogenous morphine-like compound immunopositive striatal medium spiny neurons in the rat. (A) (Top) Examples of endogenous morphine-like compounds immunostaining (brown) in the striatum of sham-operated, 6-hydroxydopamine-lesioned and L-DOPA-treated 6-hydroxydopamine-lesioned rats. (Middle) Examples of topographical counting maps. Weakly labelled medium spiny neurons are indicated by blue dots while strongly labelled medium spiny neurons are indicated by red dots. (B) Total number of endogenous morphine-like compound positive medium spiny neurons (MSN) per mm2 (mean SEM) on both lesioned (L) and unlesioned (UL) sides in sham-operated, 6-hydroxydopamine-lesioned and L-DOPA-treated 6-hydroxydopamine-lesioned rats. Two-way ANOVA; lesion effect: F(1,23) = 0.28; L-DOPA effect: F(1,23) = 0.03; interaction: F(1,23) = 1.52. (C) Number of weakly and strongly labelled medium spiny neurons per mm2 (mean SEM) on both lesioned (L) and unlesioned (UL) sides in sham-operated, 6-hydroxydopamine-lesioned and L-DOPA-treated 6-hydroxydopamine-lesioned rats. Two-way ANOVAs followed by Bonferroni. Strongly immunolabelled: lesion effect: F(1,23) = 7.65, P 5 0.05; L-DOPA effect: F(1,23) = 1.13; interaction: F(1,23) = 11.17, P 5 0.01. Weakly immunolabelled: lesion effect: F(1,23) = 11.15, P 5 0.01; L-DOPA effect: F(1,23) = 1.78; interaction: F(1,23) = 7.81, P 5 0.05. *P 5 0.05 with sham values on same side; **P 5 0.05 with 6-hydroxydopamine-lesioned values. Scale bar = 70 mm (A). eM = endogenous morphine. that are later downregulated, though not normalized, by L-DOPA treatment. Increased endogenous morphine-like compound immunoreactivity in the striatum of human Parkinson’s disease Endogenous morphine-like compound immunoreactivity is faint in post-mortem control samples (Fig. 8A) with only a few medium spiny neurons displaying a clear staining. In patients with Parkinson’s disease, however, endogenous morphine-like compound immunopositive medium spiny neurons were far more numerous with, as before, two populations: one with strongly labelled perikarya and the other with a weak staining (Fig. 8B). Endogenous morphine-like compound immunopositive astrocytes became observable in patients with Parkinson’s disease (Fig. 8B). Even though these patients have been exposed to dopaminergic drugs in the course of their disease, these observations support the experimental data collected in the rat and macaque models of Parkinson’s disease, i.e. that parkinsonism is associated with a dramatic increase in endogenous morphine-like compound immunoreactivity. Figure 5 Continued (D) Triadic association unravelled at electron microscopy level with combined endogenous morphine (immunogold particles) and S100b (black, peroxidase reaction) staining. An endogenous morphine-like compound positive medium spiny neuron (MSN; highlighted in green) is surrounded by an endogenous morphine-like compound and a S100b-positive astrocyte (A; highlighted in red) which process apposes to the basal membrane of endothelial cell lining a blood vessel (V; highlighted in blue). Green box is further magnified to better show endogenous morphine-like immunogold particles in the medium spiny neuron, while the red box shows that astrocyte stroma was S100b labelled with peroxidase and endogenous morphine-like labelled with immunogold particles. Scale bar = 10 mm (A–C), 1 mm (D). V = blood vessel. 2332 | Brain 2011: 134; 2321–2338 G. Charron et al. Figure 7 Increased number of endogenous morphine-like compound immunopositive striatal medium spiny neurons in the MPTP-lesioned macaque. (A) Examples of endogenous morphine-like compound immunostaining (brown) in the striatum of control, MPTP-lesioned and L-DOPA-treated MPTP-lesioned macaque monkeys. (B) Total number of endogenous morphine-like compound positive medium spiny neurons per mm2 (mean SEM) in the caudate nucleus and the putamen of control, MPTP-lesioned and 2 L-DOPA-treated MPTP-lesioned macaque monkeys. Number of weakly and strongly labelled medium spiny neurons per mm (mean SEM) in the caudate nucleus (C) and the putamen (D) of control, MPTP-lesioned and L-DOPA-treated MPTP-lesioned macaque monkeys. (B–D): *P 5 0.05 with control value; **P 5 0.05 with MPTP-lesioned value. Scale bar = 50 mm (A). Figure 8 Striatal endogenous morphine-like compound immunoreactivity increases in human Parkinson’s disease. Representative microphotographs of differential endogenous morphine-like compound immunostaining in the striatum of a control human subject (A) and a Parkinson’s disease subject (B). + and + + + refer to staining intensity classification. Inset in (B) shows that no endogenous morphine-like compounds immunostaining was observed in subjects with Parkinson’s disease when the primary antibody was preabsorbed with exogenous morphine. Asterisk indicates a medium spiny neuron and the arrow an astrocyte. Scale bar = 25 mm. Increase of endogenous morphine levels in mouse brain after a-methyl-ptyrosine–reserpine treatment Such consistency across models and human disease supports the proposal that dopamine depletion leads to an increased production of endogenous morphine-like compounds. These results are contrary to the expectation based on the demonstration that dopamine-null mice do not express endogenous morphine (Neri et al., 2008), i.e. that dopamine is required for the synthesis of endogenous morphine. Further neurochemical evidence supporting our anatomical data was therefore mandatory to demonstrate our case and we selected first the -methyl-p-tyrosine–reserpinetreated mouse model of Parkinson’s disease. As expected, there was a striking increase in the total number of endogenous Endogenous morphine increases in Parkinsonism Brain 2011: 134; 2321–2338 | 2333 Figure 9 Endogenous morphine levels increase in dopamine-depleted mice. (A) Number of weakly and strongly labelled medium spiny neurons per mm2 in the caudate-putamen of control (n = 4) and -methyl-p-tyrosine–reserpine (AMPT/Res) treated mice (n = 4). Results are presented as mean SEM relative to controls. Unpaired t-test: *P 5 0.05. (B and C) Amounts of dopamine (B) and morphine (C) present in brains of control (n = 4) and -methyl-p-tyrosine–reserpine treated mice (n = 5). Results are presented as nmol/g of wet tissue SEM. Unpaired t-test: *P 5 0.05. (D) Characterization of morphine in mouse brain extracts performed in selected reaction monitoring (SRM) mode with the following transitions: m/z 286.2 ! 165.1. Selected reaction monitoring trace of the specific transition for morphine in standard samples of morphine (Top: 50, 100, 500 and 1000 fmol) and mouse brain (Bottom). morphine-like compound positive medium spiny neurons in -methyl-p-tyrosine–reserpine-treated mice compared with saline-treated mice (299.3 17.0 versus 173.1 20.3, unpaired t-test, P 5 0.0001). Accordingly, there was a huge significant rise in the number of strongly labelled, associated with a significant decrease in weakly labelled medium spiny neurons (Fig. 9A). The extent of dopamine depletion induced in -methyl-p-tyrosine– reserpine-treated mice (P 5 0.05; Fig. 9B) correlated with a striking increase in endogenous morphine brain levels in -methyl-p-tyrosine–reserpine treated mice (morphine-specific enzyme-linked immunosorbent assay; P 5 0.05; Fig. 9C) compared with saline-treated animals. These neurochemical data suggest that dopamine depletion induced by -methyl-p-tyrosine– reserpine treatment in the mouse model induces an upregulation of true endogenous morphine. To confirm the presence of morphine in brain extracts of mice, liquid chromatography–tandem mass spectrometry analyses were performed in selected reaction monitoring mode. The specific transition m/z 286.2–165.1 was used to qualify the presence of morphine. The results obtained unambiguously confirm the presence of morphine in mouse brain (Fig. 9D for standard of morphine and for mouse brain). We then micro-dissected the whole striatum of two control and two -methyl-p-tyrosine–reserpine-treated mice and searched for endogenous morphine and codeine, its direct precursor (specific transition of codeine m/z 300.2–214.8, and m/z 300.2–164.8). In such low amounts of tissue, neither codeine nor endogenous morphine was detected in control mice, whereas codeine was present in -methyl-p-tyrosine–reserpine-treated mice (21.9 8.5 fmol/mg of tissue). The endogenous morphine level was still below sensitivity, reflecting the current limits of mass spectrometry detection of endogenous alkaloids in small, specific brain areas. However, the detectable level of the direct endogenous morphine precursor codeine in dopamine-depleted striatum supports the dramatic increase in endogenous morphine-like compound immunoreactivity observed in our rat and monkey models of Parkinson’s disease. Lack of endogenous morphine-like compound regulation in striatum of hyperdopaminergic DAT / mice Neurochemical data unequivocally demonstrate opposing dopamine/endogenous morphine regulation in parkinsonism instead of a parallel regulation (Neri et al., 2008). What holds true in parkinsonism might be wrong in animals with an intact nigrostrial pathway and yet hyperdopaminergic. Would dopamine be required for the synthesis of endogenous morphine? One can expect that hyperdopaminergia in a non-dopamine-depleted striatum would be linked to an increased endogenous morphine-like immunoreactivity. Endogenous morphine-like compound 2334 | Brain 2011: 134; 2321–2338 G. Charron et al. Figure 10 Endogenous morphine-like compound immunoreactive medium spiny neurons in DAT / mice striatum. (A) Similarly to the rat and the monkey, endogenous morphine-like compound immunopositive medium spiny neurons were observed in DAT + / + , DAT + / and DAT / with either moderate or intense staining. (B) The total number of endogenous morphine-like compound immunopositive medium spiny neurons was not affected by genotype. One-way ANOVA: F(2,20) = 2.32. (C) Distinguishing strongly-stained from the weakly-stained medium spiny neurons in the medial (M) or lateral (L) part of the striatum did not reveal further difference. HO = homozygous; HT = heterozygous; WT = wild-type. Scale bar = 50 mm (A). immunoreactivity in the striatum was therefore measured in a mouse model of hyperdopaminergia without degeneration of the nigrostriatal pathway, namely the dopamine transporter knock-out mouse (DAT / ). Indeed DAT / mice are known to exhibit a 500% increase in dopamine extracellular content compared with DAT + / + mice (Jones et al., 1998; Benoit-Marand et al., 2000). As no change in endogenous morphine-like compound immunoreactivity was observed in the striatum of DAT / mice compared with both DAT + / + and DAT + / (Fig. 10A–C), and because of its presence in a dopamine-depleted state, we propose that endogenous morphine synthesis is not linked to dopamine content, suggesting that alternate synthesis pathways must exist. Discussion The present data demonstrate an upregulation of endogenous morphine-like immunoreactivity, both in number of immunopositive cells and intensity of immunostaining, in the dopaminedepleted striatum of the 6-hydroxydopamine-lesioned rat model, the -methyl-p-tyrosine–reserpine treated mouse, the MPTPtreated macaque model and of human Parkinson’s disease. Such consistency across models and human disease supports the proposal that dopamine depletion leads to an increased production of endogenous morphine. In addition, L-DOPA treatment in experimental parkinsonism consistently decreased the number of endogenous morphine-like compound immunopositive medium spiny neurons. These results are contrary to expectations based on the demonstration that dopamine-null mice do not express endogenous morphine-like compounds (Neri et al., 2008), i.e. that dopamine is required for the synthesis of endogenous morphine. Endogenous morphine-like immunoreactivity increase is likely to be due to an overproduction of dopamine metabolites in the brain, such as tetrahydropapaveroline that can be used by non-dopaminergic cells to synthesize morphine-like compounds (Boettcher et al., 2005; Goumon et al., 2009). The morphine precursor tetrahydropapaveroline represents a degradation product of dopamine that is found in extracellular brain spaces, CSF and urine. Interestingly, the presence of a high quantity of endogenous morphine is reported in the urine and serum of L-DOPA-treated patients with Parkinson’s disease (Matsubara et al., 1992; Arun et al., 1998) as well as in patients with alcohol addiction. The model is invalidated by the present data set as: (i) endogenous morphine-like compounds increase occurs in a Parkinson’s disease-like state; and (ii) partial normalization of endogenous morphine-like compound immunoreactivity occurs in animals treated with L-DOPA. It thus raises the questions of the cellular origin and synthesis pathway of endogenous morphine. In patients with Parkinson’s disease, endogenous morphine-like compounds are overproduced compared with non-pathological individuals. Tetrahydropapaveroline (or norlaudanosoline) is Endogenous morphine increases in Parkinsonism supposedly related to Parkinson’s disease development and also may be derived from its treatment with L-DOPA (Soto-Otero et al., 2006). Tetrahydropapaveroline is easily found in the plasma, urine and brain of parkinsonian patients treated with L-DOPA (Matsubara et al., 1992; Cashaw, 1993a, b). It was proposed that tetrahydropapaveroline is spontaneously synthesized by a condensation of dopamine and 3,4-dihydroxyphenylacetaldehyde (metabolic degradation of dopamine by monoamine oxidase), but recent data suggest that tetrahydropapaveroline is probably converted to reticuline (next intermediate in morphine biosynthesis) by a specific enzyme (Soto-Otero et al., 2004, 2006). While sinister-tetrahydropapaveroline is involved in morphine biosynthesis in plants, recent studies have demonstrated that only rectus-tetrahydropapaveroline is converted to reticuline by the amine N-methyltransferase into endogenous morphine in mammals (Grobe et al., 2011). This exclusive selectivity for the rectus-tetrahydropapaveroline enantiomer is of great importance since high amounts of the sole sinister-tetrahydropapaveroline have been found in the human brain (Sango et al., 2000). Because rectus-tetrahydropapaveroline was not detected and considering the recent discovery of its pivotal role in endogenous morphine synthesis, we hypothesize that the rectus-tetrahydropapaveroline enantiomer is present but rapidly processed into endogenous morphine. Accordingly, L-DOPA administration, which leads to sinister-tetrahydropapaveroline formation, does not induce endogenous morphine formation in mouse brain (Grobe et al., 2010). These data explain, at least in part, the apparent discrepancy of our results with literature as the enantiomer issue explains both the lack of further endogenous morphine increase after L-DOPA treatment in our models and the increase of tetrahydropapaveroline in the urine of L-DOPA-treated patients with Parkinson’s disease. Endogenous morphine is expressed in various cell types, i.e. in neuronal and non-neuronal cells (Laux et al., 2011). Two hypotheses may explain the presence of endogenous morphine or related compounds in neurons. One possibility is that endogenous morphine and/or endogenous morphine-like compounds (morphine glucuronides and/or codeine) could be taken up from the extracellular space by active or passive transport. Indeed, several reports suggest that morphine-like compounds could be synthesized by non-dopaminergic or non-catecholaminergic cells such as the pancreatic cells, hepatocytes, placenta-derived cells (Weitz et al., 1987; Molina et al., 1995; Poeaknapo et al., 2004). In our Parkinson’s disease models, biosynthesis of endogenous morphine is obviously not completed in dopaminergic neurons. Endogenous morphine-like compounds present in non-dopaminergic brain cells may thus be synthesized after uptake of precursors (e.g. dopamine and sinister-tetrahydropapaveroline). Such a hypothesis implies cooperation between different cell types to ensure complete biosynthesis of endogenous morphine. Our results show the presence of endogenous morphine-like compounds in a large number of astrocytes. Astrocytes do not express tyrosine hydroxylase (Jaeger, 1985) and thus are unable to de novo synthesize dopamine and thus endogenous morphine. Two hypotheses can be proposed to explain the presence of endogenous morphine-like compounds in these cells. Firstly, astrocytes may achieve morphine biosynthesis after precursor Brain 2011: 134; 2321–2338 | 2335 uptake. Secondly, astrocytes are known to be involved in reuptake and the catabolism of neurotransmitters (Iversen, 2006). Interestingly, astrocytes express multidrug-resistance like protein 1, 3, 4 and 5 as well as glutamate transporter, which correspond to transporters that are described to be potentially involved in morphine/morphine-6-glucuronide/morphine-3-glucuronide/ codeine uptake/efflux (Hirrlinger et al., 2002, 2005; Bourasset et al., 2003; Somogyi et al., 2007). In addition, presence of UDP-glucuronosyltransferase 1A6 enzyme (UGT1A6) in rat astrocytes (Suleman et al., 1998; Heurtaux et al., 2006) suggests that after morphine uptake, astrocytes may catabolize morphine in morphine-glucuronides. Moreover, the presence of morphine and/or morphine glucuronides in astrocytic foot processes around blood vessels may suggest a possible uptake from the blood or release into the blood (Brix-Christensen et al., 1997, 2000). The fact that high levels of endogenous morphine are found in urine of patients with Parkinson’s disease (Matsubara et al., 1992) strongly suggests that this endogenous morphine is mainly produced by peripheral cells as endogenous morphine levels in the brain, even increased in Parkinson’s disease, would hardly explain such a rise. It is likely that biosynthesis of endogenous morphine is different in brain and peripheral cells (Laux et al., 2011). While endogenous morphine-like compound (morphine-6glucuronide) is fully synthesized in catecholaminergic chromaffin cells (Goumon et al., 2006), CNS endogenous morphine accumulates in -aminobutyric acid-ergic neurons and astrocytes that may have taken up a specific precursor (Laux et al., 2011). This difference in the endogenous morphine synthesis pathway between brain and periphery is of importance since the peripheral endogenous morphine may be taken into account for urinary excretion independently of the brain. We cannot exclude, however, that endogenous morphine synthesis could occur in non-dopaminedepleted brain regions, thus acting as a compensatory mechanism (Bezard and Gross, 1998). Altogether our results suggest that astrocytes may be responsible for morphine catabolism in the CNS, a mechanism that now requires our attention because of the extensive use of morphine in clinics and as a drug of abuse. A central unresolved question is the role of endogenous morphine and of its regulation in the striatum. Although the vast majority of the striatal neurons express endogenous morphine-like compounds, there is a striking pattern with moderate endogenous morphine-like immunoreactivity in D1 dopamine receptor-positive medium spiny neurons and intense immunoreactivity in the D2 dopamine receptor-positive medium spiny neurons. This result suggests a differential influence upon the so-called ‘direct’ and ‘indirect’ striatofugal pathways (Alexander et al., 1986; Albin et al., 1989), although physiological investigations would be needed to characterize the respective impact. It remains that the large increase in immunoreactivity after dopamine depletion should thus mostly occur in the D2 dopamine receptor-positive medium spiny neurons of the indirect pathway. These neurons co-express encephalin with -aminobutyric acid as neurotransmitters. Encephalin levels are tightly regulated by dopamine tone as the rise in encephalin expression occurs even before the symptom appearance in progressive models of Parkinson’s disease (Bezard et al., 2003b). This spatial and temporal analogy would support the hypothesis of a compensatory role for the rise in endogenous 2336 | Brain 2011: 134; 2321–2338 morphine-like compounds in Parkinson’s disease. A similar rationale can be proposed for interneurons. While no choline acetyltransferase-immunopositive neurons were found to be endogenous morphine positive, the parvalbumin-immunopositive neurons (also called the fast-spiking interneurons), displayed clear endogenous morphine immunopositivity. Those fast-spiking -aminobutyric acid interneurons provide potent feed-forward inhibition of striatal projection neurons (Mallet et al., 2005). Endogenous morphine might thus play a role in the regulation of their physiological output towards medium spiny neurons (Mallet et al., 2006). Since the functional consequences of endogenous morphine increase have not been investigated in the present study, we should limit ourselves to highlighting a few functional possibilities. A rise in morphine levels is classically associated with an analgesic effect but effects could be far more widespread. Endogenous morphine has a high affinity for the m-opiate receptor that displays high-affinity binding for morphine-related morphinan alkaloids such as morphine-6-glucuronide, endomorphins, endorphins and dynorphins, as well as the clinically established antagonists naloxone and naltrexone (Stefano et al., 1996). Different splicing variants of the m-opiate receptor are expressed in neural tissues and mediate the inhibition of adenylate cylcase and induce nitric oxide production and release (Ferreira et al., 1991; Pan, 2005; Pasternak, 2007). Nitric oxide, as a morphine second messenger, has been associated with antinociception (Przewlocki et al., 1993) as well as tolerance and dependence (Majeed et al., 1994). In relation to the immune system, morphine, but not the other opioid peptides, stimulates constitutive nitric oxide release in macrophages, granulocytes, various types of human and rat endothelial cells, invertebrate neurons and immunocytes, and in rat median eminence fragments, all in a naloxone antagonizable manner (Stefano et al., 1996, 2008). Effects of opiate alkaloid signalling upon the immune system could be seen as moderately immunosuppressive. However, one can predict that such a phenomenon should be of low magnitude in Parkinson’s disease based upon the local paracrine mode of transmission, but could be put in parallel with the immunodepressed profile of patients with Parkinson’s disease that has been suggested to be associated with fatigue (Katsarou et al., 2007) and depression (Kummer and Teixeira, 2008). Pain is a possibly related symptom but, as morphine is antinociceptive, one can posit that endogenous morphine rise is a non-motor compensatory mechanism that fails over time as do the motor compensatory mechanisms (Bezard and Gross, 1998; Bezard et al., 2003b), at least in a subpopulation of patients. We thus have a variety of non-motor symptoms (Chaudhuri and Odin, 2010) whose pathophysiology could be related to enhanced endogenous morphine levels, paving the way for experimental and clinical investigations and target identification for future treatments. Acknowledgements We thank A. Estager and M.L. Thiolat for excellent technical assistance. E.B. designed and organized the experiments; G.C., E.D., A.B., G.P., A.L., F.D., A.V.D., M.-H.C., P.B.-C., Q.L., C.Q., G. Charron et al. M.N.-B and A.V. performed experiments; G.C., B.G., Y.G. and E.B. analysed the data and wrote the article. Funding The Université Victor-Segalen Bordeaux 2, the Université de Strasbourg and the Centre National de la Recherche Scientifique provided the infrastructural support. The present work was funded by MAPKinDYSK (to E.B.) and TRAFinLID (to E.B.) Agence Nationale de la Recherche grants, a Conseil Regional d’Aquitaine fellowship (to A.B.) a French Ministère délégué à la Recherche et à l’Enseignement Supérieur fellowship ( to A.L.) and a Fondation pour la Recherche Médicale grant (to Y.G.).; The funders had no role in study design, data collection and analysis, decision to publish or preparation of the article. References Ahmed MR, Berthet A, Bychkov E, Porras G, Li Q, Bioulac BH, et al. Lentiviral overexpression of GRK6 alleviates L-dopa-induced dyskinesia in experimental Parkinson’s disease. Sci Transl Med 2010; 2: 28ra28. Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci 1989; 12: 366–75. Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 1986; 9: 357–81. Arun P, Ravikumar A, Leelamma S, Kurup PA. Endogenous alkaloids in the brain of rats loaded with tyrosine/tryptophan & in the serum of patients of neurodegenerative & psychiatric disorders. Indian J Med Res 1998; 107: 231–8. Benoit-Marand M, Jaber M, Gonon F. Release and elimination of dopamine in vivo in mice lacking the dopamine transporter: functional consequences. Eur J Neurosci 2000; 12: 2985–92. Berthet A, Porras G, Doudnikoff E, Stark H, Cador M, Bezard E, et al. Pharmacological analysis demonstrates dramatic alteration of D1 dopamine receptor neuronal distribution in the rat analog of L-DOPA-induced dyskinesia. J Neurosci 2009; 29: 4829–35. Berton O, Guigoni C, Li Q, Bioulac BH, Aubert I, Gross CE, et al. Striatal overexpression of DeltaJunD resets L-DOPA-induced dyskinesia in a primate model of Parkinson disease. Biol Psychiatry 2009; 66: 554–61. Bezard E, Ferry S, Mach U, Stark H, Leriche L, Boraud T, et al. Attenuation of levodopa-induced dyskinesia by normalizing dopamine D3 receptor function. Nature Med 2003a; 9: 762–67. Bezard E, Gross CE. Compensatory mechanisms in experimental and human parkinsonism : towards a dynamic approach. Prog Neurobiol 1998; 55: 93–116. Bezard E, Gross CE, Brotchie JM. Presymptomatic compensation in Parkinson’s disease is not dopamine-mediated. Trends Neurosci 2003b; 26: 215–21. Bianchi E, Alessandrini C, Guarna M, Tagliamonte A. Endogenous codeine and morphine are stored in specific brain neurons. Brain Res 1993; 627: 210–15. Boettcher C, Fellermeier M, Drager B, Zenk MH. How human neuroblastoma cells make morphine. Proc Natl Acad Sci USA 2005; 102: 8495–500. Bourasset F, Cisternino S, Temsamani J, Scherrmann JM. Evidence for an active transport of morphine-6-beta-d-glucuronide but not P-glycoprotein-mediated at the blood-brain barrier. J Neurochem 2003; 86: 1564–7. Brefel-Courbon C, Payoux P, Thalamas C, Ory F, Quelven I, Chollet F, et al. Effect of levodopa on pain threshold in Parkinson’s disease: a Endogenous morphine increases in Parkinsonism clinical and positron emission tomography study. Mov Disord 2005; 20: 1557–63. Brix-Christensen V, Goumon Y, Tonnesen E, Chew M, Bilfinger T, Stefano GB. Endogenous morphine is produced in response to cardiopulmonary bypass in neonatal pigs. Acta Anaesthesiol Scand 2000; 44: 1204–8. Brix-Christensen V, Tonnesen E, Sanchez RG, Bilfinger TV, Stefano GB. Endogenous morphine levels increase following cardiac surgery as part of the antiinflammatory response? Int J Cardiol 1997; 62: 191–7. Cashaw JL. Determination of tetrahydropapaveroline in the urine of parkinsonian-patients receiving L-Dopa carbidopa (sinemet) therapy by high-performance liquid-chromatography. J Chromatogr Biomed Appl 1993a; 613: 267–73. Cashaw JL. Tetrahydropapaveroline in brain-regions of rats after acute ethanol administration. Alcohol 1993b; 10: 133–8. Chaudhuri KR, Odin P. The challenge of non-motor symptoms in Parkinson’s disease. Prog Brain Res 2010; 184: 325–41. Chaudhuri KR, Schapira AH. Non-motor symptoms of Parkinson’s disease: dopaminergic pathophysiology and treatment. Lancet Neurol 2009; 8: 464–74. Dumartin B, Caillé I, Gonon F, Bloch B. Internalization of D1 dopamine receptor in striatal neurons in vivo as evidence of activation by dopamine agonists. J Neurosci 1998; 18: 1650–1. Dumartin B, Jaber M, Gonon F, Caron M, Giros B, Bloch B. Dopamine tone regulates D1 receptor trafficking and delivery in striatal neurons in dopamine trasnporter deficient mice. Proc Natl Acad Sci USA 2000; 97: 1879–84. Ferreira SH, Duarte ID, Lorenzetti BB. The molecular mechanism of action of peripheral morphine analgesia: stimulation of the cGMP system via nitric oxide release. Eur J Pharmacol 1991; 201: 121–2. Ford B. Pain in Parkinson’s disease. Mov Disord 2010; 25 (Suppl 1): S98–S103. Gintzler AR, Gershon MD, Spector S. A nonpeptide morphine-like compound: immunocytochemical localization in the mouse brain. Science 1978; 199: 447–8. Giros B, Jaber M, Jones SR, Wightman RM, Caron MG. Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature 1996; 379: 606–12. Glattard E, Welters ID, Lavaux T, Muller AH, Laux A, Zhang D, et al. Endogenous morphine levels are increased in sepsis: a partial implication of neutrophils. PLoS One 2010; 5: e8791. Gold SJ, Hoang CV, Potts BW, Porras G, Pioli E, Kim KW, et al. RGS9-2 negatively modulates L-3,4-dihydroxyphenylalanine-induced dyskinesia in experimental Parkinson’s disease. J Neurosci 2007; 27: 14338–48. Goumon Y, Laux A, Muller A, Aunis D. Central and peripheral endogenous morphine. An R Acad Nac Farm 2009; 75: 389–418. Goumon Y, Muller A, Glattard E, Marban C, Gasnier C, Strub JM, et al. Identification of morphine-6-glucuronide in chromaffin cell secretory granules. J Biol Chem 2006; 281: 8082–9. Grobe N, Lamshoft M, Orth RG, Drager B, Kutchan TM, Zenk MH, et al. Urinary excretion of morphine and biosynthetic precursors in mice. Proc Natl Acad Sci USA 2010; 107: 8147–52. Grobe N, Ren X, Kutchan TM, Zenk MH. An (R)-specific N-methyltransferase involved in human morphine biosynthesis. Arch Biochem Biophys 2011; 506: 42–7. Guigoni C, Doudnikoff E, Li Q, Bloch B, Bezard E. Altered D1 dopamine receptor trafficking in parkinsonian and dyskinetic non-human primates. Neurobiol Dis 2007; 26: 452–63. Guigoni C, Dovero S, Aubert I, Qin L, Bioulac BH, Bloch B, et al. Levodopa-induced dyskinesia in MPTP-treated macaque is not dependent of the extent and pattern of the nigrostrial lesion. Eur J Neurosci 2005; 22: 283–7. Heurtaux T, Benani A, Moulin D, Muller N, Netter P, Minn A. Induction of UGT1A6 isoform by inflammatory conditions in rat astrocytes. Neuropharmacology 2006; 50: 317–28. Brain 2011: 134; 2321–2338 | 2337 Hirrlinger J, Konig J, Dringen R. Expression of mRNAs of multidrug resistance proteins (Mrps) in cultured rat astrocytes, oligodendrocytes, microglial cells and neurones. J Neurochem 2002; 82: 716–19. Hirrlinger J, Moeller H, Kirchhoff F, Dringen R. Expression of multidrug resistance proteins (Mrps) in astrocytes of the mouse brain: a single cell RT-PCR study. Neurochem Res 2005; 30: 1237–44. Hnasko TS, Sotak BN, Palmiter RD. Morphine reward in dopaminedeficient mice. Nature 2005; 438: 854–7. Hornykiewicz O. Dopamine and brain function. Pharmacol Rev 1966; 18: 925–64. Iversen L. Neurotransmitter transporters and their impact on the development of psychopharmacology. Br J Pharmacol 2006; 147 (Suppl 1): S82–88. Jaeger CB. Cytoarchitectonics of substantia nigra grafts: a light and electron microscopic study of immunocytochemically identified dopaminergic neurons and fibrous astrocytes. J Comp Neurol 1985; 231: 121–35. Johansson PA, Andersson M, Andersson KE, Cenci MA. Alterations in cortical and basal ganglia levels of opioid receptor binding in a rat model of l-DOPA-induced dyskinesia. Neurobiol Dis 2001; 8: 220–39. Jones SR, Gainetdinov RR, Jaber M, Giros B, Wightman RM, Caron MG. Profound neuronal plasticity in response to inactivation of the dopamine transporter. Proc Natl Acad Sci USA 1998; 95: 4029–34. Kacem K, Lacombe P, Seylaz J, Bonvento G. Structural organization of the perivascular astrocyte endfeet and their relationship with the endothelial glucose transporter: a confocal microscopy study. Glia 1998; 23: 1–10. Katsarou Z, Bostantjopoulou S, Hatzizisi O, Giza E, Soler-Cardona A, Kyriazis G. Immune factors or depression? Fatigue correlates in Parkinson’s disease. Rev Neurol 2007; 45: 725–8. Konradi C, Westin JE, Carta M, Eaton ME, Kuter K, Dekundy A, et al. Transcriptome analysis in a rat model of L-DOPA-induced dyskinesia. Neurobiol Dis 2004; 17: 219–36. Kummer A, Teixeira AL. Depressive disorders in Parkinson’s disease may be due to a shared immune-mediated neurodegenerative process. Med Hypotheses 2008; 70: 201–2. Laux A, Muller AH, Miehe M, Dirrig-Grosch S, Deloulme JC, Delalande F, et al. Mapping of endogenous morphine-like compounds in the adult mouse brain: evidence of their localization in astrocytes and GABAergic cells. J Comp Neurol 2011; 519: 2390–416. Majeed NH, Przewlocka B, Machelska H, Przewlocki R. Inhibition of nitric oxide synthase attenuates the development of morphine tolerance and dependence in mice. Neuropharmacology 1994; 33: 189–92. Mallet N, Ballion B, Le Moine C, Gonon F. Cortical inputs and GABA interneurons imbalance projection neurons in the striatum of parkinsonian rats. J Neurosci 2006; 26: 3875–84. Mallet N, Le Moine C, Charpier S, Gonon F. Feedforward inhibition of projection neurons by fast-spiking GABA interneurons in the rat striatum in vivo. J Neurosci 2005; 25: 3857–69. Matsubara K, Fukushima S, Akane A, Kobayashi S, Shiono H. Increased urinary morphine, codeine and tetrahydropapaveroline in parkinsonian patient undergoing L-3,4-dihydroxyphenylalanine therapy: a possible biosynthetic pathway of morphine from L-3,4-dihydroxyphenylalanine in humans. J Pharmacol Exp Ther 1992; 260: 974–8. Molina PE, Hashiguchi Y, Meijerink WJ, Naukam RJ, Boxer R, Abumrad NN. Modulation of endogenous opiate production: effect of fasting. Biochem Biophys Res Commun 1995; 207: 312–7. Muller A, Glattard E, Taleb O, Kemmel V, Laux A, Miehe M, et al. Endogenous morphine in SH-SY5Y cells and the mouse cerebellum. PLoS One 2008; 3: e1641. Muzzi C, Bertocci E, Terzuoli L, Porcelli B, Ciari I, Pagani R, et al. Simultaneous determination of serum concentrations of levodopa, dopamine, 3-O-methyldopa and alpha-methyldopa by HPLC. Biomed Pharmacother 2008; 62: 253–8. Nadjar A, Brotchie JM, Guigoni C, Li Q, Zhou SB, Wang GJ, et al. Phenotype of striatofugal medium spiny neurons in parkinsonian and dyskinetic nonhuman primates: a call for a reappraisal of the functional organization of the basal ganglia. J Neurosci 2006; 26: 8653–61. 2338 | Brain 2011: 134; 2321–2338 Neri C, Ghelardini C, Sotak B, Palmiter RD, Guarna M, Stefano G, et al. Dopamine is necessary to endogenous morphine formation in mammalian brain in vivo. J Neurochem 2008; 106: 2337–44. Pan YX. Diversity and complexity of the mu opioid receptor gene: alternative pre-mRNA splicing and promoters. DNA Cell Biol 2005; 24: 736–50. Parent A, Cote PY, Lavoie B. Chemical anatomy of primate basal ganglia. Prog Neurobiol 1995; 46: 131–97. Pasternak GW. When it comes to opiates, just say NO. J Clin Invest 2007; 117: 3185–7. Poeaknapo C, Schmidt J, Brandsch M, Drager B, Zenk MH. Endogenous formation of morphine in human cells. Proc Natl Acad Sci USA 2004; 101: 14091–6. Prensa L, Parent A. The Nigrostriatal Pathway in the Rat: A single-axon study of the relationship between dorsal and ventral tier nigral neurons and the striosome/matrix striatal compartments. J Neurosci 2001; 21: 7247–60. Przewlocki R, Machelska H, Przewlocka B. Inhibition of nitric oxide synthase enhances morphine antinociception in the rat spinal cord. Life Sci 1993; 53: PL1–PL5. Raponi E, Agenes F, Delphin C, Assard N, Baudier J, Legraverend C, et al. S100B expression defines a state in which GFAP-expressing cells lose their neural stem cell potential and acquire a more mature developmental stage. Glia 2007; 55: 165–77. Sango K, Maruyama W, Matsubara K, Dostert P, Minami C, Kawai M, et al. Enantio-selective occurrence of (S)-tetrahydropapaveroline in human brain. Neurosci Lett 2000; 283: 224–6. Schestatsky P, Kumru H, Valls-Sole J, Valldeoriola F, Marti MJ, Tolosa E, et al. Neurophysiologic study of central pain in patients with Parkinson disease. Neurology 2007; 69: 2162–9. Schuster S, Doudnikoff E, Rylander D, Berthet A, Aubert I, Ittrich C, et al. Antagonizing L-type Ca2 + channel reduces development of abnormal involuntary movement in the rat model of L-3,4-dihydroxyphenylalanine-induced dyskinesia. Biol Psychiatry 2009; 65: 518–26. G. Charron et al. Schuster S, Nadjar A, Guo JT, Li Q, Ittrich C, Hengerer B, et al. The 3-hydroxy-3-methylglutaryl-CoA reductase inhibitor lovastatin reduces severity of L-DOPA-induced abnormal involuntary movements in experimental Parkinson’s disease. J Neurosci 2008; 28: 4311–6. Somogyi AA, Barratt DT, Coller JK. Pharmacogenetics of opioids. Clin Pharmacol Ther 2007; 81: 429–44. Soto-Otero R, Sanmartin-Suarez C, Sanchez-Iglesias S, HermidaAmeijeiras A, Sanchez-Sellero I, Mendez-Alvarez E. The ability of 1,2,3,4-Tetrahydropapaveroline to cause oxidative stress: potential implications in relation to Parkinson’s disease. J Neurochem 2004; 90: 94–94. Soto-Otero R, Sanmartin-Suarez C, Sanchez-Iglesias S, HermidaAmeijeiras A, Sanchez-Sellero I, Mendez-Alvarez E. Study on the ability of 1,2,3,4-tetrahydropapaveroline to cause oxidative stress: Mechanisms and potential implications in relation to Parkinson’s disease. J Biochem Mol Toxicol 2006; 20: 209–20. Stefano GB, Cadet P, Kream RM, Zhu W. The presence of endogenous morphine signaling in animals. Neurochem Res 2008; 33: 1933–9. Stefano GB, Goumon Y, Casares F, Cadet P, Fricchione GL, Rialas C, et al. Endogenous morphine. Trends Neurosci 2000; 23: 436–42. Stefano GB, Scharrer B, Smith EM, Hughes TK Jr, Magazine HI, Bilfinger TV, et al. Opioid and opiate immunoregulatory processes. Crit Rev Immunol 1996; 16: 109–44. Suleman FG, Abid A, Gradinaru D, Daval JL, Magdalou J, Minn A. Identification of the uridine diphosphate glucuronosyltransferase isoform UGT1A6 in rat brain and in primary cultures of neurons and astrocytes. Arch Biochem Biophys 1998; 358: 63–7. Thomas DM, Francescutti-Verbeem DM, Kuhn DM. The newly synthesized pool of dopamine determines the severity of methamphetamineinduced neurotoxicity. J Neurochem 2008; 105: 605–16. Vital A, Fernagut PO, Canron MH, Joux J, Bezard E, Martin-Negrier ML, et al. The nigrostriatal pathway in Creutzfeldt-Jakob disease. J Neuropathol Exp Neurol 2009; 68: 809–15. Weitz CJ, Faull KF, Goldstein A. Synthesis of the skeleton of the morphine molecule by mammalian liver. Nature 1987; 330: 674–77.