Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Molecular mimicry wikipedia , lookup
DNA vaccination wikipedia , lookup
Adaptive immune system wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
12-Hydroxyeicosatetraenoic acid wikipedia , lookup
Immune system wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
5-oxo-eicosatetraenoic acid wikipedia , lookup
Immunosuppressive drug wikipedia , lookup
5-Hydroxyeicosatetraenoic acid wikipedia , lookup
Hygiene hypothesis wikipedia , lookup
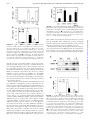
Pathogen-Induced Apoptotic Neutrophils Express Heat Shock Proteins and Elicit Activation of Human Macrophages This information is current as of June 18, 2017. Limin Zheng, Min He, Min Long, Robert Blomgran and Olle Stendahl J Immunol 2004; 173:6319-6326; ; doi: 10.4049/jimmunol.173.10.6319 http://www.jimmunol.org/content/173/10/6319 Subscription Permissions Email Alerts This article cites 50 articles, 33 of which you can access for free at: http://www.jimmunol.org/content/173/10/6319.full#ref-list-1 Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2004 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 References The Journal of Immunology Pathogen-Induced Apoptotic Neutrophils Express Heat Shock Proteins and Elicit Activation of Human Macrophages1 Limin Zheng,2*† Min He,*† Min Long,† Robert Blomgran,† and Olle Stendahl† N eutrophils are the first cells to be recruited to the site of bacterial infections, and they provide the first line of host defense against invading microorganisms. Upon encountering bacteria, neutrophils initially react by using their microbicidal mechanisms to kill the microbes and/or prevent them from spreading until the macrophages (M)3 can accumulate. Thereafter, the neutrophils are programmed to die by apoptosis and are rapidly removed by the M (1–3). There is emerging evidence that the clearance of apoptotic neutrophils not only prevents the release of potentially toxic intracellular intracellular substances, but also actively regulates the infection-induced inflammatory responses (4, 5). Uptake of apoptotic neutrophils has been shown to inhibit the production of proinflammatory mediators in M by secretion of anti-inflammatory cytokines such as TGF- (4 –10). This active anti-inflammatory response in M represents a mechanism for the safe clearance of apoptotic cells under physiological conditions and serves as a key step in the resolution phase of inflammation (4, 5). In contrast, such anti-inflammatory events can inhibit Ag presentation and promote the growth of intracellular *Key Laboratory of Gene Engineering of the Education Ministry, Department of Biochemistry, College of Life Sciences, Sun Yatsen (Zhongshan) University, Guangzhou, China; and †Division of Medical Microbiology, Faculty of Health Sciences, Linkoping University, Linkoping, Sweden Received for publication October 22, 2003. Accepted for publication September 10, 2004. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 This work was supported by the Natural Science Foundation of China and Guangdong Province (Projects 30371600 and 031620), the Swedish Medical Research Council (Projects 13026, 14689, and 5968), and the King Gustaf V Memorial Foundation, the “973” project (2004CB518801). 2 Address correspondence and reprint requests to Dr. Limin Zheng, Department of Biochemistry, College of Life Sciences, Zhongshan University, Guangzhou 510 275, People’s Republic of China. E-mail address: [email protected] Abbreviations used in this paper: M, macrophage; HSP, heat shock protein; MPO, myeloperoxidase; PI, propidium iodide. 3 Copyright © 2004 by The American Association of Immunologists, Inc. parasites within M (5, 11, 12), which implies that induction of apoptosis in neutrophils represents a pathogenic strategy for microbes to eliminate these key immune cells and use them as “Trojan horses” to enter M. However, in the cited studies, neutrophil apoptosis was induced by age or irradiation; thus, the results may not apply to the very early phase of infection, when many neutrophils undergo pathogen-induced apoptosis. Increasing numbers of pathogens have been found to modulate host cell apoptosis and thereby influence the progression of diseases (13–20). Recent studies have shown that induction of phagocyte apoptosis may not necessarily represent a pathogenic strategy to eliminate these key immune cells but rather a host defense mechanism against invading microorganisms. For example, apoptosis of M limits the intracellular growth of mycobacteria, and uptake of these infected apoptotic cells by neighboring M reduces the viability of intracellular bacteria and facilitates Ag presentation (20 –22). We have previously observed that proinflammatory responses in M are triggered by uptake of mycobacteria-induced apoptotic neutrophils, but not by ingestion of uninfected apoptotic neutrophils (16). Other investigators have shown that interactions between M and apoptotic inflammatory neutrophils prevent the growth of Leishmania major both in vitro and in vivo, whereas phagocytosis of uninfected apoptotic cells by M promotes the intracellular growth of Trypanosoma cruzi (11, 19). These findings imply that interaction with apoptotic neutrophils primes M so that they are not simply anti-inflammatory actors, as previously suggested, but they also respond to danger signals in a more complex way that may play a crucial role in host defense. It is not yet known how M distinguish between and react differently to uninfected or inflammatory apoptotic neutrophils. The role of apoptosis in modulating the pathogenesis of infectious diseases varies with the causative organisms, and the majority of pathogens that induce apoptosis are extracellular bacteria (13, 14). Therefore, we used both Gram-negative and Gram-positive extracellular bacteria as model systems to determine whether 0022-1767/04/$02.00 Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 Ingestion of aged or irradiated apoptotic neutrophils actively suppresses stimulation of macrophages (M). Many bacterial pathogens can also provoke apoptosis in neutrophils, but little is known about how such apoptotic cells influence M activation. We found that neutrophils undergoing apoptosis induced by UV irradiation, Escherichia coli, or Staphylococcus aureus could either stimulate or inhibit M activation. In contrast to M that had ingested irradiated apoptotic neutrophils, M that had phagocytosed bacteria-induced apoptotic neutrophils exhibited markedly increased production of the proinflammatory cytokine TNF-␣, but not the anti-inflammatory cytokine TGF-. Moreover, ingestion of bacteria, but not UV-induced apoptotic neutrophils, caused increased expression of Fc␥RI on M, and this effect was not provoked directly by bacteria associated with the apoptotic neutrophils. Instead, we found that a link between pathogen-induced apoptotic neutrophils and up-regulation of the heat shock proteins HSP60 and HSP70, and we also observed that recombinant HSP60 and HSP70 potentiated LPS-stimulated production of TNF-␣ in M. The opposing macrophage responses to neutrophils undergoing apoptosis induced in different ways may represent a novel mechanism that regulates the extent of the immune response to invading microbes in two steps: first by aiding the functions of M at an early stage of infection, and subsequently by deactivating those cells through removal of uninfected apoptotic neutrophils. HSP induction in neutrophils may provide the danger signals required to generate a more effective macrophage response. The Journal of Immunology, 2004, 173: 6319 – 6326. 6320 UPTAKE OF BACTERIA-INDUCED APOPTOTIC NEUTROPHILS ACTIVATES M these pathogen-induced apoptotic neutrophils influence M activation, and if so, how they exert that effect. We found that uptake of neutrophils undergoing pathogen-induced apoptosis caused M to increase both production of the proinflammatory cytokine TNF-␣ and surface expression of Fc␥RI. This activation of M was not provoked by the bacteria associated with apoptotic neutrophils. Instead, the danger signals needed to trigger the activation of macrophages seemed to come from heat shock proteins (HSP) expressed on the inflammatory apoptotic neutrophils. Materials and Methods Reagents Preparation of bacteria Escherichia coli strain ATCC 25922 and Staphylococcus aureus strain Wood46 (both from American Type Culture Collection, Manassas, VA) were cultured for 18 h (15, 17) and then washed and resuspended in RPMI 1640 medium containing 5% FCS and 2 mM L-glutamine (RPMI medium). In some experiments the bacteria were fixed with 7.3% formalin for 30 min and washed before use. Cell isolation Human neutrophils were isolated from peripheral blood of healthy donors as previously reported (16). Neutrophils of ⬃96% purity were resuspended in RPMI medium. Human M were prepared from PBMC as described previously (8, 16). The cells in DMEM alone were plated at 4 ⫻ 106/well in 24-well plates for 1.5 h, washed, then cultured in DMEM containing 10% human AB serum. The medium was changed every 3 days and was replaced with DMEM without serum before use in the experiments (8, 16). For all experiments, M had been cultured for 6 –9 days before use. Induction of neutrophil apoptosis Neutrophils (2 ⫻ 106/ml) were incubated with E. coli or S. aureus (cell to bacteria ratio, 1:20) for 20 min at 37°C; when using fixed bacteria, the neutrophils were also exposed to UV irradiation for 8 min. Thereafter, the samples were washed twice (200 ⫻ g, 5 min each time) with PBS and incubated in RPMI medium containing 50 g/ml gentamicin at 37°C for 3 h in a humidified CO2 incubator. For UV-induced apoptosis, neutrophils were exposed to UV irradiation for 8 min, followed by culture for 3 h. In some experiments neutrophils were exposed to mild heat (43°C) for 30 min, then incubated at 37°C for 3 h (23). This methodology routinely yielded populations comprising 40 –70% cells positively stained with annexin V and ⬍4% positive for propidium iodide (PI). Where indicated, neutrophils were pretreated with 10 g/ml cycloheximide for 10 min before exposure to bacteria or heat stress. Assessment of neutrophil apoptosis Neutrophil apoptosis was quantified by flow cytometry using FITC-conjugated annexin V and by morphological examination (16, 17). Specific binding of annexin V was achieved by incubating 106 cells in 60 l of binding buffer saturated with annexin V for 15 min at 4°C in the dark. To discriminate between early apoptosis and necrosis, the cells were simultaneously stained with annexin V and PI before analysis. The binding of annexin V-FITC (FL1) and PI (FL2) to the cells was measured by flow cytometry (FACSCalibur, BD Biosciences) using CellQuest software (BD Biosciences, Mountain View, CA) (16). At least 10,000 cells were counted in each sample, and a gate based on forward and side scatters was set to exclude cell debris. Morphological assessment of apoptosis was performed on Giemsa- or Turck-stained cytocentrifuged neutrophils as previously described (16, 17). Neutrophils were lysed, and the genomic DNA was extracted according to the protocol for apoptotic DNA laddering kit (R&D Systems). Samples (2 g of DNA/lane) were analyzed by gel electrophoresis (1.8% agarose) and ethidium bromide staining. The gel was visually examined under 305 nm of UV illumination (16). Flow cytometric analysis of Fc␥RI expression M were left untreated or were incubated for 1 h with neutrophils undergoing apoptosis induced in different ways and were subsequently washed, then cultured for an additional 18 h. Fc␥RI expression on M was determined by flow cytometry using R-PE-conjugated anti-Fc␥RI and an isotype-matched control mAb (DakoCytomation, Glostrup, Denmark), essentially as previously described (24). Cytokine production in M after phagocytosis of apoptotic neutrophils Neutrophils undergoing apoptosis induced by exposure to UV irradiation, heat shock, S. aureus, or E. coli were washed twice with PBS and resuspended in DMEM without serum. Before use, M growing in each well were washed and replaced with DMEM without serum. Apoptotic neutrophils (4 ⫻ 106/well of M) were added, and the plates were incubated at 37°C for 1 h. Thereafter, the wells were washed vigorously to remove uningested neutrophils (8, 16, 25), fresh DMEM without serum was added, and the supernatants were collected 18 h later (8, 16). As a control, apoptotic neutrophils were cultured for 18 h in DMEM without serum. In some experiments, M were incubated with recombinant human HSP60, HSP70, or LPS (from E. coli 055:B5) for 18 h. Where indicated, M were preincubated with 10 g/ml polymyxin B sulfate for 30 min. The supernatants were centrifuged to remove particular debris and were stored in aliquots at ⫺70°C. Cytokine concentrations in the culture supernatants were determined by ELISA, using Quantikine immunoassay kits according to the instructions provided by the manufacturer. We performed a phagocytic assay (8, 16, 25) to ensure that the M ingested equivalent numbers of neutrophils undergoing apoptosis induced in different ways. In short, the monolayers were fixed with 1% formalin overnight and stained for myeloperoxidase (MPO) as a marker of ingested neutrophils (8, 16); the M themselves were routinely negative for MPO staining. Phagocytosis of apoptotic neutrophils was quantified under an inverted phase contrast microscope essentially as previously described (8). M that showed discrete, round, MPO-positive inclusions were scored as having ingested one or more apoptotic neutrophils (8, 25). Western blotting The neutrophils were washed three times with PBS, and the pellets were resuspended in lysis buffer (16) for 20 min on ice. After centrifugation at 10,000 ⫻ g for 10 min, the supernatants were dissolved in Laemmli sample buffer (26) and heated at 95°C for 5 min. Equal amounts of cellular proteins (15 g/lane for HSP70; 35 g/lane for HSP60 and HSP90) were separated on 10% SDS-PAGE and electrotransferred to nitrocellulose membranes. The membranes were blocked with 5% milk, and the presence HSPs on the blots was detected with specific Ab and a commercial ECL kit (16). To confirm that each lane received the same amount of proteins, the blots were stripped and reprobed with anti-actin (C-2) Ab (Santa Cruz Biotechnology, Santa Cruz, CA). LPS detection To analyze the LPS content in M containing ingested apoptotic neutrophils, the monolayers were lysed by vigorous pipetting in H2O for 1 h and then collected and centrifuged (2500 ⫻ g, 10 min). LPS in the samples was determined using a Limulus amebocyte lysate endotoxin assay kit. Statistical analysis The data on cytokine concentrations are given as the mean ⫾ SEM, and the data on apoptotic rates are the mean ⫾ SD. Statistical significance was determined by Student’s t test. A value of p ⬍ 0.05 was considered statistically significant. Results Neutrophil apoptosis induced by viable E. coli and S. aureus In initial experiments, flow cytometry using FITC-conjugated annexin V revealed that neutrophils exposed to E. coli or S. aureus underwent rapid apoptosis. This effect was positively correlated Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 An annexin V apoptosis detection kit and Quantikine kits for immunoassays of TNF-␣, TGF-, and IL-10 were purchased from R&D Systems (Abingdon, U.K.). Anti-HSP60 (SPA-806), HSP70 (SPA-810, detects the inducible form of HSP70), and HPS90 (SPA-830) Ab and recombinant human HSP60 (ESP-540) and HSP70 (ESP-555, the inducible form of HSP70) were obtained from StressGen (Victoria, Canada). ESP-540 and ESP-555 were the low endotoxin preparations containing ⬍50 endotoxin U/mg proteins. The Limulus amebocyte lysate endotoxin assay kit (Charles River Endosafe, Charleston, SC) and cell isolation and tissue culture reagents were obtained from Invitrogen Life Technologies (Lidingo, Sweden), and electrophoresis and ECL reagents were purchased from Amersham Biosciences (Uppsala, Sweden). All other reagents were obtained from Sigma-Aldrich (St. Louis, MO) unless otherwise indicated in the text. DNA fragmentation assay The Journal of Immunology with exposure time and bacteria to cell ratio, and excessive apoptosis was associated with loss of membrane integrity in an increased portion of neutrophils, which indicates necrosis or late apoptosis (data not shown). Apoptotic and necrotic cells have different effects on activation of M (5, 25, 27); therefore, we exposed the neutrophils to bacteria at a ratio of 1:20 for 20 min, then washed and cultured them for 3 h. Under these conditions, ⬃50% of the neutrophils were in the early stage of apoptosis with preserved membrane integrity (lower right quadrants in Fig. 1A), and ⬍4% were positive for both annexin V and PI (upper right quadrants in Fig. 1A). Morphological examination showed typical ap- 6321 optotic changes in neutrophils, including decreased cell volume and chromatin condensation with loss of multilobular nuclear structure (Fig. 1B). Similar results were obtained in complementary experiments performed to assay DNA fragmentation (Fig. 1C). Only viable bacteria induced appreciable apoptosis, although low rates of such cell death were observed in uninfected neutrophils (6 ⫾ 3%) and in neutrophils exposed to fixed bacteria under identical conditions (7 ⫾ 4 and 6 ⫾ 2% for fixed E. coli and S. aureus, respectively). Ingestion of bacteria-induced apoptotic neutrophils stimulates TNF-␣ production in M Uptake of bacteria-induced apoptotic neutrophils increases the expression of Fc␥RI on M To further test the hypothesis that M are activated by bacteriainduced apoptotic neutrophils, we examined another response exhibited by M, namely expression of Fc␥RI (CD64). Flow cytometry revealed that Fc␥RI was expressed constitutively on the surface of human M, and the level of expression was not affected by ingestion of uninfected apoptotic neutrophils (mean fluorescence intensity, 433 ⫾ 36 and 420 ⫾ 28, respectively). In contrast, FIGURE 1. Neutrophil apoptosis induced by exposure to UV irradiation, S. aureus, or E. coli. Neutrophils were left untreated (1) or were exposed to UV irradiation (2), S. aureus (3), or E. coli (4). Thereafter, the neutrophils were cultured for 3 h, washed, and then analyzed as indicated. A, Flow cytometric analysis of binding of annexin V-FITC (FL1) and PI (FL2) in neutrophils. The relative distribution of cells manifesting early (annexin⫹PI⫺) and late (annexin⫹PI⫹) apoptosis is illustrated (percentage); for clarity, the fluorescence profiles of 10% of the analyzed events are shown. B, Morphological analysis of apoptosis. Cytospin preparations of neutrophils were stained with Turck’s reagent (gentian violet) and examined under a light microscope; the micrograph shows neutrophils with condensed nuclei characteristic of apoptosis. C, Agarose gel electrophoresis of internucleosomal DNA fragmentation in neutrophils. The molecular markers are indicated to the left (M). The results shown are representative of at least five independent experiments. The proportions of apoptotic cells (mean ⫾ SD) are indicated in Table I. FIGURE 2. Photomicrograph of human monocyte-derived M that have ingested apoptotic neutrophils. The neutrophils are stained for MPO and appear as condensed black bodies within the MPO-negative macrophages. Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 Uptake of uninfected apoptotic neutrophils is known to suppress M activation (4 –9). To determine whether neutrophils undergoing apoptosis provoked by bacteria have the same impact on M, we compared the influence of bacteria- and UV-induced apoptotic neutrophils on the inflammatory responses of M. Approximately 40% of the M stained positively for neutrophils, and each positive M had ingested ⬃1.2 neutrophils (Fig. 2 and Table I). In agreement with previous reports (4 –9), M that ingested UV-induced apoptotic neutrophils in our experiments displayed inhibited production of the proinflammatory cytokine TNF-␣, whereas they produced increased levels of anti-inflammatory TGF-1. In contrast, M that phagocytosed bacteria-induced apoptotic neutrophils showed a markedly increased level of TNF-␣ and a slightly decreased amount of TGF-1 (Fig. 3). Ingestion of apoptotic neutrophils elicited by E. coli appeared to augment IL-10 production in M, but this increase was not statistically significant, because the IL-10 level produced by unstimulated macrophages was low (Fig. 3). Moreover, apoptotic neutrophils incubated in the absence of M did not generate any measurable levels of cytokines (data not shown); thus, the cytokines we detected must have been produced by the M. UPTAKE OF BACTERIA-INDUCED APOPTOTIC NEUTROPHILS ACTIVATES M 6322 Table I. Induction and rates of neutrophil apoptosis and phagocytosis of apoptotic neutrophils by Ma Treatment of Neutrophils UV E. coli Fixed E. coli/UV S. aureus Fixed S. aureus/UV Neutrophil Apoptosis (%) M with Ingested Neutrophils (%) 54 ⫾ 8.9 51 ⫾ 13.9 42 ⫾ 7.9 44 ⫾ 8.2 42 ⫾ 8.1 41 ⫾ 3.0 42 ⫾ 0.7 40 ⫾ 2.5 40 ⫾ 2.8 39 ⫾ 4.5 a Apoptosis was induced in neutrophils by UV irradiation, exposure to viable bacteria, or both fixed bacteria exposure and UV irradiation as described in Materials and Methods. Rates of apoptosis were measured by flow cytometry using annexin V. Phagocytosis of apoptotic neutrophils by M was determined by MPO staining. Each value represents the mean ⫾ SD of results from at least five paired experiments. Activation of M by inflammatory apoptotic neutrophils is not caused by bacteria associated with the neutrophils Intact bacteria or bacterial components can elicit proinflammatory responses in M. Accordingly, we performed three sets of experiments to determine whether activation of M by inflammatory neutrophils undergoing apoptosis is actually caused by the bacteria that are associated with the neutrophils. First, neutrophils were incubated with fixed E. coli or S. aureus under the same conditions as those used with viable bacteria, and this did not trigger apoptosis in the neutrophils (Table I). M that were allowed to interact with the neutrophils exposed to fixed bacteria produced only a moderate amount of TNF-␣ (Fig. 4). Only dead neutrophils are ingested by M; thus, the divergent effects on M activation exerted by neutrophils exposed to fixed vs viable bacteria may have been due to differences in the rates of apoptosis exhibited by the neutrophils. To address that possibility, we also irradiated neutrophils exposed to fixed bacteria to increase the apoptosis (Table I), but the M that ingested those neutrophils still displayed much lower production of TNF-␣ than M incubated with neutrophils undergoing apoptosis induced by viable bacteria (Fig. 4). Even though the neutrophils were washed extensively before incubation with M, they still may have carried a small number of bacteria that could have activated the M. Therefore, we performed a second set of experiments in which a CFU assay was used to determine the number of bacteria associated with neutrophils (28). No viable bacteria were detected in the suspension of apoptotic neutrophils before it was added to M, and ⬍5 ⫻ 104 CFU were found in 106 neutrophils immediately after exposure to viable E. coli or S. aureus. Each well on the culture plate contained ⬃106 M (8, 16), and ⬃40% of the M contained neutrophils; thus, we estimated that, at most, 2 ⫻ 104 bacteria had interacted with the M in each well. As shown in Fig. 4, TNF-␣ production was only marginally increased in M exposed to 5 ⫻ 105 viable bacteria. In the third set of experiments conducted to gain further evidence that neutrophil-associated bacteria did not interfere with activation of M, we measured LPS present in M that had been exposed to apoptotic neutrophils. M were incubated with neutrophils undergoing apoptosis induced by viable E. coli or by fixed E. coli and UV irradiation, which led to LPS levels of 0.19 ⫾ 0.11 and 0.11 ⫾ 0.03 ng/well, respectively (n ⫽ 5). However, adding 0.2 ng of LPS/well caused the M to produce 300 ⫾ 31 pg of FIGURE 3. Phagocytosis of bacteria-induced apoptotic neutrophils stimulates production of TNF-␣ by human M. M were incubated for 1 h with neutrophils undergoing apoptosis induced by exposure to UV irradiation, S. aureus, or E. coli. Thereafter, the preparations were washed, the supernatants were collected 18 h later and cytokine concentrations were determined by ELISA. M incubated with medium alone served as controls. The illustrated data represent the mean ⫾ SE of results from 12 separate experiments. ⴱ, p ⬍ 0.05; ⴱⴱ, p ⬍ 0.01 (compared with controls). TNF-␣/ml, which is only ⬃10% of the level observed in M exposed to viable E. coli-induced apoptotic neutrophils. These results clearly indicate that very little of the activation of M caused by inflamed apoptotic neutrophils could have been induced by the bacteria associated with the neutrophils. Thus, these apoptotic neutrophils must generate some other signals that can be sensed by M and consequently elicit active immune responses. Increased expression of HSP in inflammatory apoptotic neutrophils It has been shown that certain members of the HSP family (e.g., HSP60, HSP70, and HSP90) can directly stimulate cells of the innate immune system and thereby act as danger-signaling molecules (29 –35). Therefore, we performed Western blot analysis to determine the amounts of HSP60, HSP70, and HSP90 expressed in apoptotic neutrophils. The results revealed that freshly isolated neutrophils contained all three of these proteins, and the level of expression was not altered in neutrophils undergoing UV-induced apoptosis (Fig. 5). In contrast, the expression of HSP60 and HSP70, but not HSP90, was significantly increased in pathogeninduced apoptotic neutrophils (Fig. 5). Reprobing the blots with anti-actin Ab confirmed that equal amounts of the proteins had been loaded in each lane (data not shown). Effect of heat-stressed apoptotic neutrophils and recombinant HSP on activation of M We used both heat-stressed apoptotic neutrophils and recombinant HSP to determine whether the increased expression of HSP60 and HSP70 in bacteria-induced apoptotic neutrophils played a role in activation of M. Neutrophils that were exposed to mild heat exhibited increased apoptosis (55 ⫾ 6% cells stained positively with Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 Fc␥RI expression was increased by ⬃60% in M that had phagocytosed apoptotic neutrophils induced by E. coli or S. aureus (mean fluorescence intensity, 716 ⫾ 52 and 696 ⫾ 30, respectively; n ⫽ 5). The Journal of Immunology annexin V and ⬍3% stained positively for PI; n ⫽ 5) and upregulated expression of HSP60 and HSP70 (Fig. 5), and M that ingested these neutrophils showed markedly increased production of TNF-␣ (Fig. 6A). Moreover, such generation of TNF-␣ was only marginally affected by the addition of polymyxin B, whereas in parallel experiments the activity of LPS was completely blocked by polymyxin B (Fig. 6B). These results indicate that heat-stressed apoptotic neutrophils and LPS stimulate M activation in different ways. Numerous investigations have shown that both HSP60 and HSP70 potently provoke proinflammatory responses (29, 33–36), but two recent studies of murine M have suggested that the TNF␣-inducing activities of recombinant HSP preparations are due entirely to contaminating LPS (37, 38). Accordingly, to clarify the role of HSP in activation of human M, we used highly purified recombinant HSP60 and HSP70 preparations (ESP-540 and ESP555; StressGen Biotechnologies, Victoria, Canada), which contain low levels of endotoxin and do not induce release of TNF-␣ from murine M (37, 38). In our experiments, both recombinant HSP60 and HSP70, at concentrations up to 5 g/ml, caused only a slight increase in TNF-␣ production by human M (Fig. 7). Interestingly, recombinant HSP60 synergistically increased the LPS-stimulated release of TNF-␣ from M, and recombinant HSP70 had the same effect, albeit not as pronounced (Fig. 7). HSP60 and HSP70 had no additive effect in the presence or the absence of LPS (Fig. 7). We used the protein synthesis inhibitor cycloheximide to further confirm that HSPs from apoptotic neutrophils are involved in activation of M. Pretreatment of neutrophils with cycloheximide almost completely blocked the increased expression of HSP60 and HSP70 induced by exposure to viable E. coli or heat stress (Fig. 8A). Furthermore, TNF-␣ production was significantly lower in M that had ingested the cycloheximide-treated apoptotic neutrophils (Fig. 8B). FIGURE 5. Increased expression of HSP60 and HSP70 in neutrophils undergoing bacteria-induced apoptosis. Neutrophils were left untreated or were exposed to heat stress, UV irradiation, E. coli, or S. aureus. Thereafter, the neutrophils were cultured for 3 h and lysed. Equal amounts of cellular proteins (15 g/lane for HSP70; 35 g/lane for HSP60 and HSP90) were separated on 10% SDS-PAGE and blotted with Ab specific against HSP60, HSP70, and HSP90. Each of the illustrated blots is representative of four separate experiments. Discussion Neutrophils are the first cells attracted to the site of bacterial invasion. These leukocytes are responsible for direct bactericidal activities, such as phagocytosis and generation of the respiratory burst, and they can also regulate inflammatory responses by undergoing apoptosis (2). In our experiments, apoptotic neutrophils induced by exposure to UV irradiation, E. coli, or S. aureus either stimulated or inhibited the activation of M. We found that phagocytosis of bacteria-induced apoptotic neutrophils, but not uptake of UV-induced apoptotic neutrophils, caused M to increase both production of the proinflammatory cytokine TNF-␣ and surface expression of Fc␥RI. These findings indicate that bacteria-induced neutrophil apoptosis represents a novel host defense mechanism that aids the activities of local M and links innate and adaptive immune responses to achieve control of infection. Moreover, we found that HSPs expressed by inflammatory apoptotic neutrophils may constitute the signals required to generate a more effective immune response. Our results give important new insights into the significance of neutrophil-macrophage interactions during inflammation. The dynamics of inflammation and changes in the inflammatory environment over time may have a substantial impact on the way that apoptosis is induced in neutrophils, which, in turn, can lead to either pro- or anti-inflammatory responses in M. In agreement with previous reports (4 – 8, 25, 27), we noted that uptake of UVinduced apoptotic neutrophils actively suppressed stimulation of M. However, as mentioned above, we also found that ingestion of bacteria-induced apoptotic neutrophils caused M to increase production of TNF-␣ and expression of Fc␥RI. Our finding that M were activated by neutrophils undergoing apoptosis induced by either extracellular (present study) or intracellular (16) pathogens clearly indicates that apoptosis that occurs in neutrophils after exposure to bacteria represents a general host defense mechanism, but not a strategy that invading pathogens use to kill immune cells. Inasmuch as the short-lived neutrophils are the first cells to temporarily infiltrate the site of microbial invasion, infection of M can occur indirectly through uptake of bacteria-laden apoptotic neutrophils, instead of directly (i.e., by entry of bacteria themselves into the M), which might lead to different immune responses. For example, it has been shown that direct infection of M with mycobacteria blocks the responses of these phagocytes to stimulation with IFN-␥ (24, 39), but we have previously shown that M produce much more TNF-␣ after ingesting mycobacteriainduced apoptotic neutrophils than after being directly infected Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 FIGURE 4. Activation of M by neutrophils undergoing bacteria-induced apoptosis is not caused by the bacteria associated with the neutrophils. The neutrophils were incubated with viable or fixed E. coli (A) or S. aureus (B), or they were exposed to fixed bacteria and UV irradiation as described in Materials and Methods. M were subsequently coincubated with the neutrophils for 1 h, after which the cultures were washed, and the supernatants were collected 18 h later. M incubated in medium alone or only with bacteria (5 ⫻ 105/well) served as controls. TNF-␣ concentrations in the culture supernatants were determined by ELISA. Values represent the mean ⫾ SE from 10 separate experiments. ⴱⴱ, p ⬍ 0.01 (compared with viable E. coli-induced apoptotic neutrophils). 6323 6324 UPTAKE OF BACTERIA-INDUCED APOPTOTIC NEUTROPHILS ACTIVATES M FIGURE 7. Recombinant HSP60 and HSP70 synergistically increase LPS-stimulated release of TNF-␣ from M. Human M were incubated in medium alone or in medium containing 3 g/ml HSP60, HSP70, or HSP60 and HSP70 in the presence (f) or the absence (䡺) of 0.2 ng/ml LPS for 18 h. Thereafter, the TNF-␣ concentration in the culture supernatants was determined. Values represent the mean ⫾ SE from four separate experiments. ⴱⴱ, p ⬍ 0.01 (compared with LPS alone). with M. tuberculosis (16). These findings suggest that bacteriainduced neutrophil apoptosis represents a novel link between innate and adaptive immune responses. This conclusion is supported by recent studies in which it was observed that interactions between M and inflammatory apoptotic neutrophils promoted the killing of Leishmania major both in vitro and in vivo, whereas uninfected apoptotic lymphocytes amplified the intracellular growth of L. major and T. cruzi within M (11, 19). In addition, it has been found that depletion of neutrophils is associated with a decreased Th1 immune response and increased bacteria growth in various tissues (40 – 42). It is not yet known exactly how M can distinguish between and react differently to inflammatory apoptotic neutrophils and UVinduced apoptotic neutrophils. It has been speculated that the pattern recognition receptors on M enable the phagocytic synapse in these cells to perceive and interpret molecular patterns on apoptotic cells, which determine how the M respond (4, 5, 43). However, the pattern recognition receptors involved in binding of apoptotic cells overlap with those that recognize pathogens or necrotic cells (4, 5, 11, 43). This is illustrated by the findings that activation of M is inhibited by binding of the CD14 or phosphatidylserine receptor to apoptotic cells, whereas M are actually activated by binding of CD14 to bacteria or by linking phosphatidylserine receptors to necrotic cells (11, 27, 43– 45). Moreover, we noted that in the absence of serum, stimulation with LPS or intact bacteria alone or with neutrophils that had been exposed to fixed bacteria caused M to produce only moderate amounts of TNF-␣ that corresponded to ⬃10 –20% of the levels seen in M stimulated by incubation with neutrophils undergoing apoptosis induced by viable bacteria (Fig. 4). These results indicate that very little of the activation of M by inflammatory apoptotic neutro- FIGURE 8. Cycloheximide inhibits the expression of HSPs in neutrophils undergoing bacteria- or heat-stress-induced apoptosis and reduces the ability of those cells to stimulate production of TNF in M. Neutrophils were preincubated in medium alone or in medium containing 10 g/ml cycloheximide (CHX) for 10 min and were subsequently exposed to viable E. coli or heat stress, then cultured for 3 h. A, Expression of HSPs in these neutrophils was determined by Western blot analysis as described in Fig. 5, and each of the illustrated blots is representative of four separate experiments. B, M were incubated for 1 h with apoptotic neutrophils that had (f) or had not (䡺) been pretreated with cycloheximide, after which the cultures were washed, and supernatants were collected 18 h later. Values represent the mean ⫾ SE from five separate experiments. ⴱⴱ, p ⬍ 0.01 (compared with the M exposed to cycloheximide-treated neutrophils). Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 FIGURE 6. Uptake of heat-stressed apoptotic neutrophils stimulates production of TNF-␣ in M. A, Neutrophils were left untreated or were exposed to mild heat (43°C) for 30 min, then cultured for 3 h. Those neutrophils were subsequently coincubated with M for 1 h, after which the cultures were washed, and supernatants were collected 18 h later. M incubated in medium alone or with LPS (0.2 ng/ml) served as controls. B, M were preincubated with (f) or without (䡺) 10 g/ml polymyxin B (PB) for 10 min, after which LPS or neutrophils were added. Values represent the mean ⫾ SE from four separate experiments. ⴱⴱ, p ⬍ 0.01 (compared with control M in A and with M treated with polymyxin in B). phils could have been induced by the bacteria that were associated with the neutrophils. Thus, inflammatory apoptotic neutrophils themselves must generate additional signals that can be sensed by M and consequently provoke active immune responses. The deficiency of self/non-self paradigm has led to new hypotheses proposed by Janeway and Matzinger (46, 47), in that the immune system is more concerned with damage than with foreignness. According to this paradigm, the immune system is thought to The Journal of Immunology References 1. Malech, H. D., and J. I. Gallin. 1987. Neutrophils in human diseases. N. Engl. J. Med. 317:687. 2. Haslett, C. 1997. Granulocyte apoptosis and inflammatory diseases. Br. Med. Bull. 53:669. 3. Aderem, A., and D. M. Underhill. 1999. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 17:593. 4. Fadok, V. A., D. L. Bratton, and P. M. Henson. 2001. Phagocyte receptors for apoptotic cells: recognition, uptake, and consequences. J. Clin. Invest. 108:957. 5. Savill, J., I. Dransfield, C. Gregory, and C. Haslett. 2002. A blast from the past: clearance of apoptotic cells regulates immune responses. Nat. Rev. Immunol. 2:965. 6. Ren, Y., L. Stuart, F. P. Lindberg, A. R. Rosenkranz, Y. Chen, T. N. Mayadas, and J. Savill. 2001. Nonphlogistic clearance of late apoptotic neutrophils by macrophages: efficient phagocytosis independent of 2 integrins. J. Immunol. 166:4743. 7. Huynh, M. L., V. A. Fadok, and P. M. Henson. 2002. Phosphatidylserine-dependent ingestion of apoptotic cells promotes TGF-1 secretion and the resolution of inflammation. J. Clin. Invest. 109:41. 8. Fadok, V. A., D. L. Bratton, A. Konowal, P. W. Freed, J. Y. Westcott, and P. M. Henson. 1998. Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokines production through autocrine/paracrine mechanisms involving TGF-, PGE2, and PAF. J. Clin. Invest. 101:890. 9. Byrne, A., and D. J. Reen. 2002. Lipopolysaccharide induces rapid production of IL10 by monocytes in the presence of apoptotic neutrophils. J. Immunol. 168:1968. 10. Mevorach, D., J. O. Mascarenhas, D. Gershov, and K. B. Elkon. 1998. Complement-dependent clearance of apoptotic cells by human macrophages. J. Exp. Med. 188:2313. 11. Lopes, M. F., C. G. Freire-de-Lima, and G. A. DosReis. 2000. The macrophages haunted by cell ghosts: a pathogen grows. Immunol. Today 10:489. 12. Laskay, T., G. van Zandbergen, and W. Solbach. 2003. Neutrophil granulocytes: Trojan horses for Leishmania major and other intracellular microbes? Trend Microbiol. 11:210. 13. Gao, L. Y., and Y. A. Kwaik. 2000. The modulation of host cell apoptosis by intracellular bacterial pathogens. Trends Microbiol. 8:306. 14. Weinrauch, Y., and A. Zychlinsky. 1999. The induction of apoptosis by bacterial pathogens. Annu. Rev. Microbiol. 53:155. 15. Watson, R. W., H. P. Redmond, J. H. Wang, C. Condron, and D. Bouchier-Hayes. 1996. Neutrophils undergo apoptosis following ingestion of E. coli. J. Immunol. 156:3986. 16. Perskvist, N., M. Long, O. Stendahl, and L. Zheng. 2002. Mycobacterium tuberculosis promotes apoptosis in human neutrophils by activating caspase-3 and altering expression of Bax/Bcl-xL via an oxygen-dependent pathway. J. Immunol. 168:6358. 17. Lundqvist-Gustafsson, H., S. Norrman, J. Nilsson, and Å. Wilsson. 2001. Involvement of p38-mitogen-activated protein kinase in staphylococcus aureusinduced neutrophil apoptosis. J. Leukocyte Biol. 70:642. 18. Nash, P. B., M. B. Purner, R. P. Leon, P. Clarke, R. C. Duke, and T. J. Curiel. 1998. Toxoplasma gondii-infected cells are resistant to multiple inducers of apoptosis. J. Immunol. 160:1824. 19. Ribeiro-Gomes, F. L., A. C. Otero, N. A. Gomes, M. C. A. Moniz-de-Souza, L. Cysne-Finkelstein, A. C. Arnholdt, V. L. Calich, S. G. Coutinho, M. F. Lopes, and G. A. DosReis. 2004. Macrophage interactions with neutrophils regulate Leishmania major infection. J. Immunol. 172:4454. 20. Fratazzi, C., R. D. Arbeit, C. Carini, and H. G. Remold. 1997. Programmed cell death of Mycobacterium avium serovar 4-infected macrophages prevents the mycobacteria from spreading and induces mycobacterial growth inhibition by freshly added, uninfected macrophages, J. Immunol. 158:4320. 21. Schaible, U. E., F. Winau, P. A. Sieling, K. Fischer, H. L. Collins, K. Hagens, R. L. Modlin, V. Brinkmann, and S. H. Kaufmann. 2003. Apoptosis facilitates antigen presentation to T lymphocytes through MHC-I and CD1 in tuberculosis. Nat. Med. 9:1039. 22. Yrlid, U., and M. J. Wick. 2000. Salmonella-induced apoptosis of infected macrophages results in presentation of a bacteria-encoded antigen after uptake by bystander dendritic cells. J. Exp. Med. 191:613. 23. Eid, N. S., R. E. Kravath, and K. W. Lank. 1987. Heat-shock protein synthesis by human polymorphonuclear cells. J. Exp. Med. 165:1448. 24. Ting, L. M., A. C. Kim, A. Cattamanchi, and J. D. Ernst. 1999. Mycobacterium tuberculosis inhibits IFN-␥ transcriptional responses without inhibiting activation of STAT1. J. Immunol. 163:3898. 25. Fadok, V. A., D. L. Bratton, L. Guthrie, and P. M. Henson. 2001. Differential effects of apoptotic versus lysed cells on macrophage production of cytokines: role of proteases. J. Immunol. 166: 6847. 26. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage. Nature 227:680. 27. Cocco, R. E., and D. S. Ucker. 2001. Distinct modes of macrophage recognition for apoptotic and necrotic cells are not specified exclusively by phosphatidylserine exposure. Mol. Biol. Cell 12:919. 28. Zheng, L., T. P. L. Zomerdijk, C. Aarnoudse, R. van Furth, and P. H. Nibbering. 1995. Role of protein kinase C isozymes in Fc␥ receptor-mediated intracellular killing of S. aureus by human monocytes. J. Immunol. 155:776. 29. Asea, A., S. K. Kraeft, E. A. Kurt-Jones, M. A. Stevenson, L. B. Chen, G. C. Koo, and S. K. Calderwood. 2000. HSP70 stimulates cytokine production through a CD14-dependent pathway, demonstrating its dual role as a chaperone and cytokine. Nat. Med. 6:435. 30. Radsak, M. P., N. Hilf, H. Singh-Jasuja, S. Braedel, P. Brossart, H. G. Rammensee, and H. Schild. 2003. The heat shock protein Gp96 binds to human neutrophils and monocytes and stimulates effector functions. Blood 101:2810. 31. Kumaraguru, U., C. D. Pack, and B. T. Rouse. 2003. Toll-like receptor ligand links innate and adaptive immune responses by the production of heat-shock proteins. J. Leukocyte Biol. 73:574. 32. Srivastava, P. 2002. Roles of heat-shock proteins in innate and adaptive immunity. Nat. Rev. Immunol. 2:185. 33. Vabulas, R. M., P. Ahmad-Nejad, S. Ghose, C. J. Kirschning, R. D. Issels, and H. Wagner. 2002. HSP70 as endogenous stimulus of the Toll/interleukin-1 receptor signal pathway. J. Biol. Chem. 277:15107. Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017 call into action and initiate protective immune responses by “danger molecules” released from tissue during infection or stress (46). There is accumulating evidence that HSPs, which serve as chaperone molecules, are the key danger signals sent to the immune system (29, 32, 36). An M-stimulating role for these proteins is supported by the observations that pathogen-induced apoptotic neutrophils are associated with up-regulation of HSP60 and HSP70, and heat-stressed apoptotic cells activate M. This conclusion is further supported by our findings that recombinant HSP60 and HSP70 potentiated LPS-stimulated production of TNF-␣ in M, and inhibition of HSP expression in E. coli- or heat stress-induced apoptotic neutrophils also suppressed the activation of M elicited by these apoptotic neutrophils. In contrast to several previous reports (29, 33, 35), we found that recombinant HSP per se did not induce a significant amount of TNF-␣ by M, and this discrepancy can probably be explained by the use of different HSP preparations. Two recent investigations of murine macrophages (37, 38) have shown that the TNF-␣-inducing effects of recombinant HSP60 and HSP70 preparations are due entirely to the presence of contaminating LPS, which concurs with the fact that the highly purified HSP preparations used in our study did not induce the release of TNF-␣ from M. In all of our experiments, pathogen-induced apoptotic neutrophils had a greater capacity to stimulate M than did either heatstressed apoptotic neutrophils or a combination of purified HSP and LPS. This indicates that additional danger signals from apoptotic cells and/or other bacterial components are required to activate M. Perhaps pathogen-induced apoptotic neutrophils contain multiple stress proteins and bacterial components that can bind to a distinct set of receptors on M and thereby synergistically trigger different signaling pathways in M. This idea is supported by a recent study showing that stressed apoptotic tumor cells or a tumor cell-derived chaperone-rich cell lysate is generally superior to individual HSPs in terms of inducing antitumor immunity (34). In the course of infectious diseases, the immune system is permanently challenged to eliminate, or at least control, the infectious agents and to minimize the destruction of tissue architecture. Our results demonstrating that M react differently after ingesting neutrophils undergoing apoptosis induced in different ways might reflect a novel mechanism by which neutrophil apoptosis regulates the extent of the immune response to bacterial infections. More specifically, this means that during the initial stage of inflammation, pathogen-induced neutrophil apoptosis not only will lead to selective removal of inflamed cells from the site of infection, but will also help local M gain control over the microbes. Once the bacteria are cleared from the site of infection, phagocytosis of excess uninfected apoptotic neutrophils will inhibit M activation, resulting in rapid resolution of inflammatory responses, thus avoiding undesirable tissue damage. We also found that HSP60 and HSP70 potentiate the effect of LPS on M activation, and that function together with the previously reported receptor-mediated uptake of HSPs by APCs (48 –50) imply that HSPs are powerful natural adjuvants, which may have important implications for the development of vaccines against tumors and infectious diseases. 6325 6326 UPTAKE OF BACTERIA-INDUCED APOPTOTIC NEUTROPHILS ACTIVATES M 34. Feng, H., Y. Zeng, M. W. Graner, A. Likhacheva, and E. Katsanis. 2003. Exogenous stress proteins enhance the immunogenicity of apoptotic tumor cells and stimulate antitumor immunity. Blood 101:245. 35. Bethke, K., F. Staib, M. Distler, U. Schmitt, H. Jonuleit, A. H. Enk, P. R. Galle, and M. Heike. 2002. Different efficiency of heat shock proteins (HSP) to activate human monocytes and dendritic cells: superiority of HSP60. J. Immunol. 169:6141. 36. Wallin, R. P. A., A. Lundqvist, S. H. More, A. Bonin, R. Kiessling, and H.-G, Ljunggren. 2002. Heat-shock proteins as activators of the innate immune system. Trends Immunol. 23:130. 37. Gao, B., and M. F. Tsan. 2003. Endotoxin contamination in recombinant human heat shock protein 70 (Hsp70) preparation is responsible for the induction of tumor necrosis factor ␣ release by murine macrophages. J. Biol. Chem. 278:174. 38. Gao, B., and M. F. Tsan. 2003. Recombinant human heat shock protein 60 does not induce the release of tumor necrosis factor ␣ from murine macrophages. J. Biol. Chem. 278:22523. 39. Fortune, S. M., A. Solache, A. Jaeger, P. J. Hill, J. T. Belisle, B. R. Bloom, E. J. Rubin, and J. D. Ernst. 2004. Mycobacterium tuberculosis inhibits macrophage responses to IFN-␥ through myeloid differentiation factor 88-dependent and -independent mechanisms. J. Immunol. 172:6272. 40. Ismail, H. F., P. Fick, J. Zhang, R. G. Lynch, and D. J. Berg. 2003. Depletion of neutrophils in IL-10⫺/⫺ mice delays clearance of gastric Helicobacter infection and decreases the Th1 immune response to Helicobacter. J. Immunol. 170:3782. 41. Fulton, S. A., S. M. Reba, T. D. Martin, and W. H. Boom. 2002. Neutrophilmediated mycobacteriocidal immunity in the lung during Mycobacterium bovis BCG infection in C57BL/6 mice. Infect. Immun. 70:5322. 42. Gregory, S. H., and E. J. Wing. 2002. Neutrophil-Kupffer cell interaction: a critical component of host defenses to systemic bacterial infections. J. Leukocyte Biol. 72:239. 43. Gregory, C. D. 2000. CD14-dependent clearance of apoptotic cells: relevance to the immune system. Curr. Opin. Immunol. 12:27. 44. Fadok, V. A., D. L. Bratton, D. M. Rose, A. Pearson A, R. A. Ezekewitz, and P. M. Henson. 2000. A receptor for phosphatidylserine-specific clearance of apoptotic cells. Nature 405:85. 45. Lucas, M., L. M. Stuart, J. Savill, and A. Lacy-Hulbert. 2003. Apoptotic cells and innate immune stimuli combine to regulate macrophage cytokine secretion. J. Immunol. 171:2610. 46. Matzinger, P. 2002. The danger model: a renewed sense of self. Science 296:301. 47. Medzhitov, R., and C. A. Janeway, Jr. 2002. Decoding the patterns of self and nonself by the innate immune system. Science 296:298. 48. Becker, T., F. U. Hartl, and F. Wieland. 2002. CD40, an extracellular receptor for binding and uptake of Hsp70-peptide complexes. J. Cell Biol. 158:1277. 49. Long, K. H., F. J. Gomez, R. E. Morris, and S. L. Newman. 2003. Identification of heat shock protein 60 as the ligand on Histoplasma capsulatum that mediates binding to CD18 receptors on human macrophages. J. Immunol. 170:487. 50. Habich, C., K. Baumgart, H. Kolb, and V. Burkart. 2002. The receptor for heat shock protein 60 on macrophages is saturable, specific, and distinct from receptors for other heat shock proteins. J. Immunol. 168:569. Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017