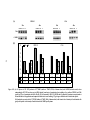
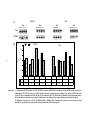
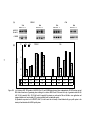
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
CHAPTER I INTRODUCTION Plant growth regulators or phytohormones, either naturally occurring or synthetic, can increase yields of a target plant. These include auxins, cytokinins, gibberellins, ethylene and abscisic acid (ABA) (Farooq et al 2009). The phytohormone abscisic acid (ABA) controls many important aspects of plant growth and development including seed development, desiccation tolerance of seeds, seed dormancy etc. Besides these, it plays crucial role in the plant’s response to abiotic and biotic stresses (Chinnusamy et al 2004). Plants accumulate increased amounts of ABA under drought, cold or salt stress conditions where drought stress is having the most prominent effect (Finkelstein et al 2002). Two ABA-dependent regulatory pathways have been proposed. One is bZip/ABRE system and other is MYC/MYB system (Agarwal et al 2006), however there are several ABA-independent pathways are also proposed to be working in plants where one pathway includes DREB/CBF system (Fowler and Thomashow 2002). Ultimate effectors of these all regulatory pathways are stress responsive genes which have been categorized into different groups like genes encoding transcription factors, protein kinases (Kaur and Gupta 2005), genes encoding proteins involved in the protection of membranes and proteins of the cell like LEA proteins, HSPs, chaperons etc (Rodriguez et al 2005), genes encoding osmoregulatory proteins (Bohnert et al 1995), genes encoding antioxidant and detoxifying proteins (Bartosz 1997), genes encoding ion transporters/channels (Bohnert et al 1995). ABA-responsive genes are either containing ABRE (ABS Response Element) in their promoters region recognized by bZip/ABF transcription factors or MYBRS/MYCRS (MYC Recognition site/ MYB Recognition Site) recognized by MYC/MYB transcription factors. Many ABA-responsive genes have been identified like genes encoding seed storage proteins, LEA and various other proteins. Examples of ABA-responsive abiotic-stress related genes are Em in wheat, Em and Rab16 in rice, Hva1, Hva2 and dehydrins in barley, Em and Rd29 in Arabidopsis thaliana (Bray 1993, Zhang et al 2005). LEA-proteins are Late Embryogenesis Abundant-proteins which are accumulated in seeds during seed maturation and play roles in protection of cell membranes and enzymes during dessication period after seed maturation. LEA-proteins have been classified into nine groups based on their sequence similarities and conserved domains (Hundertmark and Hincha 2008). Among nine groups, roles of only group 1, 2 and 3 have been investigated in imparting abiotic stress tolerance. ABA is found to induce expressions of these genes during seed development and abiotic stresses (Finkelstein et al 2002, Dalal et al 2009). Many LEA genes are reported to be responsive to ABA but some are non-responsive to ABA also. As LEA-proteins play important roles in protecting cells’ structures under stresses, hence these represent a group of stress-responsive genes. These genes are the downstream genes of both ABA-dependent and –independent signal pathways (Hong-Bo et al 2005, Kobayashi et al 2008a, 2008b). In wheat, reports related to such type of studies like elucidating the ABAdependency or –independency of LEA genes are very few except few reports like LEA genes (belonging to group 2 and group 3) were studied in wheat seedlings under low temperature, drought and ABA stresses, four of them were found to be responsive to ABA (Kobayashi et al 2004, 2006) and later found to contain ABRE in their promoter regions (Kobayashi et al 2008a, 2008b). One mode of ABA action is related to oxidative stress in plant cells. Role of ABA in the induction of antioxidant defence has been the subject of extensive research. It has been documented that ABA resulted in increased generation of ROS in maize (Guan et al 2000, Jiang and Zhang 2001, 2002a, 2002b, 2002c, Hu et al 2005), Vicia faba (Zhang et al 2001); induced the expression of antioxidant genes encoding SOD, CAT in maize (Guan et al 2000), rice (Kaminaka et al 1999); enhanced the activities of antioxidant enzymes (SOD, CAT, APX, GR) and increased contents of antioxidant metabolites (ascorbic acid, glutathione, tocopherol, carotenoids) in maize (Jiang and Zhang 2001), tobacco (Bueno et al 1998), cotton (Bellaire et al 2000 ). However, all antioxidant genes are not responsive to ABA as both ABA-dependent and ABA-independent signalling pathways are found to be involved in expression of antioxidant genes under abiotic stresses (Bellaire et al 2000, Guan et al 2000). LEA proteins and antioxidant mechanism play important roles in stress resistance mechanism of plant. As ABA is involved in regulation of such enzymes/genes during abiotic streses, elucidation of ABA-dependency/independency of these proteins/genes under stresses will contribute to understanding of stress tolerance mechanism of wheat plant. Comparing such responses in two wheat cultivars contrasting in drought tolerance for these enzymes/genes will generate better understanding of such mechanisms. The present study was aimed, to study such responses under water stress and to compare same in two wheat cultivars PBW343 (drought susceptible) and C306 (drought resistant) with following objectives: 1. To understand antioxidation mechanism of wheat seedlings at molecular level under drought in the presence and absence of exogenous ABA. 2. To study role of LEA genes of wheat seedlings under drought and studying for ABA-dependent or independent induction of these genes. 2 CHAPTER II REVIEW OF LITERATURE 2.1 Late embryogenesis abundant (LEA) proteins 2.1.1 Late embryogenesis abundant proteins 2.1.2 Desiccation induced folding of LEA 2.1.3 Classification 2.1.3.1 Group 1 2.1.3.2 Group 2 2.1.3.3 Group 3 2.1.3.4 Group 4 2.1.4 Localization of dehydrin in plants treated by different stresses 2.1.5 Association in expression of stress responsive genes 2.1.6 Expression of LEA genes in different plants at different developmental stages 2.1.7 Induction of expression of Lea genes 2.1.8 Promoter analysis of stress inducible genes 2.2 Antioxidant metabolism 2.2.1 Reactive oxygen species and their production sites 2.2.2 ROS scavenging enzymatic antioxidants 2.2.2.1 Superoxide dismutase 2.2.2.2 Catalase 2.2.2.3 Ascorabte peroxidase 2.2.2.4 Guaicol peroxidase 2.2.2.5 Glutathione reductase 2.2.3 Non-enzymatic antioxidants 2.2.3.1 Ascorbic acid 2.2.3.2 Reduced glutathione 2.2.3.3 Proline 2.2.4 Hydrogenperoxide (H2O2) 2.2.4.1 Cellular and extracellular sources of H2O2 2.2.4.1.1 Chloroplast 2.2.4.1.2 Peroxisomes 2.2.4.1.3 Mitochondria 2.2.4.1.4 Other sources 2.2.4.2 H2O2 in stress conditions and as signaling molecule 2.2.4.3 H2O2 signaling during growth and development 2.2.5 Malondialdehyde 2.3 Abscisic acid 2.3.1 ABA as a long distance signal mediating whole plant response to drought and salt stress 2.3.2 Cell signaling from stress perception to ABA accumulation 2.3.3 ABA and antioxidant defense in plant cell 2.3.3.1 ABA –induced ROS generation 2.3.3.2 Expression of genes encoding antioxidant enzymes induced by ABA 2.3.3.3 ABA induced modulation of metabolic and redox control pathways Plants have evolved two major mechanisms to cope up with water deficit: stress avoidance and stress tolerance. Stress avoidance is achieved by specialized morphological adaptations in plant architecture e.g. the development of specialized leaf structures to decrease the rate of transpiration, the reduction of leaf area, sunken stomata or increase in root length and density to use water more efficiently. Water stress tolerance appears to be the result of co-ordination of physiological and biochemical alteration at cellular and molecular level: i.e. the accumulation of various osmolytes and late embrogenesis abundant proteins coupled with efficient antioxidant system (Ramanjulu and Bartels 2002). 2.1.1 Late Embryogenesis Abundant Proteins (LEA) LEA proteins are the family of hydrophilic proteins that forms an integral part of dessication tolerance of seeds. LEA proteins were first identified by Leon Dure III about 29 yrs ago when he observed the proteins accumulated in high level during maturation phase of cotton (Gossypium hirsutum) embryogenesis. Subsequently LEA like proteins or their genes were found in seeds of many other plants as well as in vegetative organs when plant exposed to stress conditions such as cold, drought, high salinity or ABA. They are mainly localized in cytoplasm and nuclear region. Their common structural features include a high hydrophilicity, a lack or low proportion of Cys and Trp residues and a preponderance of certain amino acid residues such as Gly, Ala, Glu, Lys/Arg and Thr which later led them to be considered as a subset of hydrophilins (Garay-Arroyo et al 2000). LEA proteins are class of hydrophilic proteins expressed in late seed development and have been consistently associated with dessication tolerance because of their expression profile and their hydrophilicity. LEA proteins are prominent in plants (there are > 50 lea 4 genes in Arabidopsis Thaliana) but have also been found and related to dessication tolerance in anhydrobiotes belonging to other kingdoms, such as bacteria and nematodes (Battista et al 2001, Browne et al 2002). A number of mechanisms have been proposed to justify their role in dessiaction tolerance: water replacement, ion sequestering, macromolecules and membrane stabilization. Experimentally, several lea proteins were shown to behave invitro as cryoprotectants (Bravo et al 2003), stabilizes glassy states (Wolkers et al 2001), prevent protein aggregation (Goyal et al 2005) and bind to lipid vesicles (Koag et al 2003) or actin filaments (Abu-Abied et al 2006). The molecular mechanism of their action is rather enigmatic. One likely reason for the slow progress in this area of structure-function characterization of lea protein is that they are highly hydrophilic and lack well defined structures (Goyal et al 2003, Mouillon et al 2006). 2.1.2 Dessication induced folding of LEA proteins Lea proteins seem to be natively unfolded, at least in the hydrated state. Structural disorder confers advantages onto the protein such as increased speed of interactions, the combination of specificity with weak and reversible binding and able to carry out more than one function (Tompa et al 2004). Recent experimental data showed conformational shift on dehydration of group3 lea protein from nematode AavLEA1. Fourier transform infrared spectronic analysis shows that protein become more folded, developing a significant alphahelical component when dried. This was extremely unusual observation because protein dehydration is more often associated with a loss of structure, aggregation rather than increase in structure, folding or subunit assembly. 2.1.3 Classification Initially, LEA proteins were referred to according to their molecular weights-D-7, D- 11, D-19, D-29, D-34, D95, D113. Later LEA proteins were classified according to the appearance of different sequence motifs/patterns or biased amino acid composition, into seven distinctive groups or families. Groups 1, 2, 3, 4, 6 and 7 correspond to the hydrophilic or ‘‘typical’’ LEA proteins, whereas those LEA proteins that show hydrophobic characteristics (‘‘atypical’’) have been kept in group 5, where they could be subclassified according to their homology A computational method called protein or oligonucleotide probability profile (POPP) analysis has recently been developed in peptide composition rather than in sequence (Wise 2003). 2.1.3.1 Group 1 Sequence motifs and peptide profiles This set of LEA proteins, originally represented by the D-19 and D-132 proteins from developing cotton seeds, were recognized by an internal 20-mer sequence (TRKEQ[L/M]G[T/ E]EGY[Q/K]EMGRKGG[L/E]). They contain a very large proportion 5 of charged residues, which contributes to their high hydrophilicity and a high content of Gly residues (approximately 18%). Secondary structure They exist largely as random coils or unstructured in aqueous solution. Structural analyses using circular dichroism (CD) strongly indicate that group1 members exhibit a high percentage (70%–82.5% of their residues) of random coil configuration in aqueous solution, with a small percentage of the protein exhibiting a left-handed extended helical or poly-(LPro)-type (PII) conformation (Soulages et al 2002). Functions Their possible role in the adaptation of different organisms to water scarcity is supported by the fact that the transcripts of bacterial group 1 LEA-like proteins also accumulate under stressful conditions such as stationary growth phase, Glc or phosphate starvation, high osmolarity, high temperature, and hyperoxidant conditions. Further evidence comes from the presence of these proteins in organisms with extreme habitats, such as archaeons (uncultured like methanogenic RC1), as well as in some primordial saltwater crustaceans such as Artemia (Wang et al 2007). Group 1 LEA-like proteins are particularly abundant in the thick-shelled eggs of Artemia, whose encysted form can survive in a dried, metabolically inactive state for 10 or more years while retaining the ability to endure severe environmental conditions (Macrae 2005). Direct evidence showing a function for group 1 LEA proteins is scarce. In vitro experiments using recombinant versions of wheat (Triticum aestivum) Em protein suggested their ability to protect citrate synthase or LDH from aggregation and/or inactivation due to desiccation or freezing (Goyal et al 2005, Gilles et al 2007). A mutation in the predicted ahelical domains in the N terminus of the rEm protein suggested a role for this region in providing protection from drying (Gilles et al 2007). Tolerance to stress conditions induced by the constitutive expression of genes from this group has not been reported in plants; however, the expression of wheat TaEm in Saccharomyces cerevisiae seems to attenuate the growth inhibition of yeast cultures normally observed in high-osmolarity media. Also, the absence of one of two group 1 members in Arabidopsis plants led to a subtle phenotype of premature seed dehydration and maturation, suggesting a role during seed development (Manfre et al 2006). The expression in vegetative tissues from plants grown under optimal growth conditions of some of the group 1 LEA proteins implies that they may also have a role during normal seed development. 6 2.1.3.2 Group 2 Sequence motif and peptide profile A distinctive feature of group 2 LEA proteins is a conserved, Lys-rich 15-residue motif, EKKGIMDKIKEKLPG, named the K -segment which can be found in one to 11 copies within a single polypeptide. An additional motif also found in this group is the Y-segment, whose conserved consensus sequence is [V/T]D[E/Q]YGNP, usually found in one to 35 tandem copies in the N terminus of the protein . Many proteins of this group also contain a tract of Ser residues, called the S-segment (Jiang and Wang 2004). The presence and arrangement of these different motifs in a single polypeptide allow the classification of group 2 LEA proteins into five subgroups. Proteins that only contain the K-segment are in the Ksubgroup, and those that include the S-segment followed by K-segment are in the SKsubgroup. In addition, there are the YSK-, YK-, and KS-subgroups. Only K -segment is present at least once in all dehydrins, making it the distinguishing feature of this group. Functional significance of S-segment is known. It is phosphorylated leading to calcium binding activity in some dehydrins. Secondary structure CD spectra of full length Arabidopsis dehydrins (COR47, LTI29, LTI30, RAB18) and isolated peptides (K-, Y- and K-rich segments) showed mostly unordered structures in solution, with a variable content of poly-Pro helices. However, neither temperature, metal ions, nor stabilizing salts could promote ordered structures in either the peptides or the fulllength proteins. The motif studied most in depth was the K-segment of dehydrin. It can form amphipathic hydrophilic alpha-helix structure which is probably involved in protein-protein interactions and protein-lipid interactions (Mouillon et al 2006). Experimental evidence showed that K-segment was important to stabilize other cellular components under stress conditions. S-segment and RRKK segment were considered to be involved in nucleus localization signal. Y-showed similarity to nucleic acid binding site of chaperon. Role Like group 1 LEA proteins, several studies of specific group 2 LEA proteins have confirmed that they accumulate during seed desiccation and in response to water deficit induced by drought, low temperature,or salinity (Nylander et al 2001).These proteins are also present in nearly all vegetative tissues during optimal growth conditions (Rorat et al 2004). A role in bud dormancy has also been attributed to group 2 LEA proteins (Karlson et al 2003a, 2003b). The ability to withstand freezing is highly developed in some trees, in which the buds, which are critical for reassuming growth after winter, can build up tolerance to temperatures as low as -196 ºC. Therefore, of particular interest is the fact that LEA proteins 7 from this group are expressed in birch (Betula spp.) apices during wintertime dehydration, a period in which buds become highly desiccated during endodormancy (Puhakainen et al 2004b). Similarly, the accumulation of chilling-responsive LEA proteins from this group was detected in floral buds of blueberry (Vaccinium myrtillus), a woody perennial. 2.1.3.3 Group 3 Group3 lea proteins are characterized by multiple copies of an 11-amino acid motif TAQAAKEKAGE (Battaglia et al 2008). The protein sequences have regularly spaced lysine repeats. Peptide composition also showed that alanine and glutamine are overexpressed, where as glycine is underrepresented (Wise 2003, Tunnacliffe and Wise 2007). Secondary structure Secondary structure prediction of group3 proteins by bioinformatics tool suggests that 11-mer repeating unit principally form amphipathic alpha-helices, in the presence of sucrose and glycerol after fast drying, which may dimerizes into an unusual right handed coiled-coil arrangement (Battaglia et al 2008). Slow drying reversibly leads to both alphahelical and intermolecular extended beta-sheet structure (Wolkers et al 2001). E.g. of group3 in different species D-7 protein from Typhalatifolia, AavLEA1 protein from nematode (Goyal et al 2003), GmPM16 protein from soyabean (Shih et al 2004) and PsLEAM protein from pea (Tolleter et al 2007 ). Functions The propensity of group3 protein for alpha-helix formation enable them to contribute to the formation of cytoplasmic glass or interact with membrane during dessication condition thus stabilizing the cellular structure or maintains the activity of enzymes (Tolleter et al 2007). Group3 proteins hiC6 and hiC12 from Chlorella vulgaris have cryoprotectant activity, through which hiC6 and hiC12 protect Lactate Dehydrogenase and Malate Dehydrogenase against freeze-inactivation. The relationship between cryoprotection and the unit number of 11-mer amino acid motif in hiC6 was investigated (Honjoh et al 2000). Cryoprotectant activity decreased with decrease in unit number of 11-mer motifs. 2.1.3.4 Group 4 Sequence motif and peptide profile The LEA4 proteins are characterized by a high content of A, T, and G amino acid residues, the latter highly represented in unstructured proteins. They have a conserved Nterminal domain of 70 to 80 residues, predicted to form amphipathic a-helices, and a less conserved C-terminal region with variable size and random coil structure. Like other LEA proteins, the LEA4 group is highly accumulated in all embryo tissues of dry seeds. LEA4 proteins are involved in the adaptive response of vascular plants to withstand water deficit. 8 Role These may act as protectors of macromolecules and some cellular structure during water deficit, by preferentially interacting with the available water molecules and providing a hydration shell to protect the target” integrity and function (Garay-Arroyo et al 2000, Hoekstra et al 2001). An invitro dehydration assay showed that plant hydrophilins (LEA proteins from group 2, 3, 4) were able to protect the activity of malate dehydrogenase and lactate dehydrogenase under low water availability conditions (Reyes et al. 2005). Similarly same invitro assay were used to assess the protective activity of group 4 LEA proteins under dehydrations and cold. 2.1.4 Localization of dehydrins in plants subjected to different stresses Cold-induced dehydrins were present in the epidermis and in the vascular cylinder around xylem and phloem in Deschampsia Antarctica.This report is in agreement with previous reports on dehydrins (group2) localization in other plant species. LTI30 was present in the vascular tissue in Arabidopsis thaliana treated by low temperature (Nylander et al 2001); P-80, a dehydrin from barley, accumulates in vascular cylinders and epidermis in response to low temperature; PCA60, a dehydrin from peach, was accumulated in epidermal, cortical, phloem and xylem tissue. Early ice nucleation in vascular cylinder and epidermis could induce a severe dehydration in adjacent cells. Therefore, the presence of cold-induced dehydrins in these tissues could be related to preventing protoplasmic dehydration stress associated with freezing and thus increasing freezing tolerance. In the cell surrounding the xylem vessels dehydrins might function as water attractants during the transport of water from xylem vessel to sink tissues. Accumulation of dehyrin constitutes an adaptive response in plants to cope with further stress imposed by freezing. Dehydrin localization is extensive. Dehyrins have also been identified in cytosol of dessicated leaf cells of Craterostigma plantagiineum Hochst and the euchromatin, nucleolus and cytosol of various cells in tomato seedlings and mature plant tissues (TAS 14). In Arabidopsis thaliana, tissue and cell type specific accumulation of dehydrins have been reported as ERD14 in vascular tissue, LT129 in roots and stems, RAB18 provascular tissue of embryo (Nylander et al 2001). Pea (Pisum sativum) seed mitochondria were indeed found to accumulate a LEA protein (LEAM) with repeated motifs predicted to form amphipathic helices (Grelet et al 2005). There is also e.g. of two closely related group3 LEA proteins located in endoplasmic reticulum. N -terminal signal peptide suggesting translocation of ER membrane. In case of maize gp2 proteins, DHN1/Rab17, distribution between nucleus and cytoplasm is controlled by phosphorylation of serine stutter: removal of this sequence results in lack of phosphorylation and retention in cytoplasm. 9 2.1.5 Association between expressions of stress responsive genes Based on Venn diagram analysis, differences and cross talk of gene expression among cold-, drought-, and high-salinity stress responses and ABA response in rice has been reported. As shown in Figure-(2.1) 36, 62, 57, and 43 genes are cold-, drought-, highsalinity-, and ABA-inducible genes identified by RNA gel- blot analysis, respectively. More than 98% of the high-salinity- and 100% of ABA-inducible genes were also induced by drought stress, which indicates a strong relationship not only between drought and highsalinity responses but also between drought and ABA responses. These results indicate the existence of a substantial common regulatory system or a greater cross talk between drought and high-salinity stress and between drought and the ABA signaling process than that between cold and high-salinity stress or between cold and the ABA signaling process. These results in rice are consistent with previous observations on the overlap of drought- and highsalinity- responsive gene expression in Arabidopsis (Shinozaki and Yamaguchi-Shinozaki 2000, Seki et al 2002a, 2002b) as well as in Brassica Napus (Chen et al 2010). However, these results are contradictory to the observations made by Kreps et al (2002) in cold- and high-salinity-stressed Arabidopsis and to those made by Ozturk et al (2002) in drought and high-salinity-stressed barley (Hordeum vulgare). In their studies, the majority of stressregulated changes appeared to be stress specific and not part of a general stress response common to cold, drought, and high-salinity stress. These discrepancies may be attributable to the difference in plant species used, stress treatment, plant growth condition, array strategy, or detection methodologies. During the identification of dehydrin gene fragment in Deschampsia Antarctica northern blot analysis of LEA gene showed the transcript level in osmotic stress was higher than in salt stress, suggesting that ionic effect of salt may reduces its expression. Salt stress induces osmotic adjustment of the cytoplasm by balancing the increased osmotic potential (lowered water potential) caused by sodium sequestration inside the vacuole, where it can accumulate to concentration that have marked effect on the osmotic balance of plant cell suggesting that salt and osmotic stress have a similar signal transduction pathway. Fig. 2.1 10 2.1.6 Expression of LEA genes in various plant tissues and at different developmental stages Expression patterns could provide insight into the function of LEA genes. Accumulation of LEA proteins is found to occur during the late stages of development. Some LEA genes were expressed in non-seed tissues under normal or stress conditions. Most peanut LEA genes showed high expression during seed development which is consistent with expression of LEA genes from other plant species. In this work, semi-quantitative RT-PCR was used for expression analysis of peanut LEA genes. The results showed that most AhLEA genes were highly expressed in seeds from 30 to 90 DAP (days after pegging) and were not detected in non-seed tissue, which was similar to the results observed from Arabidopsis (Bies-Ethève et al 2008). The expression of AhLEA1, AhLEA4, AhLEA5 and AhLEA8 genes was only detected in seeds. The expression of seven LEA protein genes (AhDHN1, AhLEA3-1, AhLEA3-2, AhLEA3-4, AhLEA3-6, AhLEA6-1 and AhLEA7-1) could be detected both in seed and non-seed tissues including root, stem, leaf and flower. The expression of AhLEA3-2 and AhLEA6-1 was observed in all the five non-seed tissues tested. AhDHN1 was expressed weakly in leaf, flower and stem. AhLEA3-1 and AhLEA7-1 were expressed in root, stem, leaf and flower. AhLEA3-6 was expressed in root, leaf and flower. Interestingly, AhLEA3-4 was highly expressed in flower but not in other non-seed tissues. In 10 DAP seeds the expression of most LEA genes was undetectable during seed development. However, the expression of AhLEA6-1, AhLEA6-2 and AhLEA7-1 was clearly detected in 10 DAP seeds. The expression of 17 LEA genes could be detected in 30 DAP seeds. The results showed that different groups of LEA genes exhibited variable expression patterns during seed development, for example, the genes of LEA1, LEA5, LEA6, LEA7 and LEA8 groups. Even different members from one group of LEA showed distinct expression patterns, for instance, LEA6-1 and LEA6-2 (Su et al 2011). 2.1.7 Induction and regulation of expression of LEA genes One characteristic feature of these proteins is that the expression of the genes that encode them is transcriptionally induced by the plant growth regulator abscisic acid and this induction is occur in highly seed specific manner. The LEA polypeptide is proteolytically degraded rapidly following seed germination, and although its synthesis can be re-induced in germinating embryos subjected to osmotic stress or ABA treatment, such stress- and hormone induction does not occur in mature vegetative tissues. This highly seed specific pattern of expression sets of group1 LEA proteins apart from members of LEA group2 and group3, which additionally accumulate in ABA- and stress- induced vegetative tissues, and likewise have an association with desiccation tolerance (Ramunjulu and Bartels 2002, Wise and Tunnacliffe 2004). 11 In higher plants, the Group 1 LEA genes are expressed in a strictly seed-specific manner and transcription is induced in response to the plant growth regulator ABA and also to the imposition of osmotic stress in the maturation and early germination phases of seed development. As a bryophyte, Physcomitrella does not produce seeds. However, it is known that the promoter of the wheat Em gene does exhibit an ABA- and stress-responsive pattern of expression, when introduced as a transgene to Physcomitrella. It was therefore clearly of interest to determine whether the bryophyte counterpart of this gene exhibited a similar pattern of expression. Northern blot hybridization of RNA isolated from Physcomitrella protonemal tissue showed unambiguously that PpLEA-1 transcript is induced to accumulate rapidly to a high concentration upon incubation of tissue with ABA or following the imposition of an osmotic stress (10% mannitol) sufficient to induce plasmolysis of the cells. Additionally, the degree of accumulation of the PpLEA-1 transcript appears correlated with the concentration of ABA applied to the tissue. During seed maturation, the LEA gene expression is regulated by the coordinated action of transcription factors (TFs), such as ABI3/VP1, ABI4, ABI5, LEC1, LEC2, and FUS3. LEC1, LEC2, and FUS3 primarily regulate the transition from embryogenesis to germinative growth. ABI3, ABI4, and ABI5 may regulate the ABA response, possibly forming a regulatory complex that mediates seed -specific and/or ABA-inducible expression (e.g., LEA synthesis). LEC1 and FUS3 also partly participate in these processes (Finkelstein et al 2002). With abiotic stresses, such as salt, drought stress, the biosynthesis and accumulation of ABA are increased in plant. The increased ABA interacts with the ABA receptors PYR/PYL/RCAR of START proteins, which in turn bind to and inhibit the protein phosphatase types 2C (PP2C) proteins (e.g., ABI1, ABI2, and HAB1) and activate the SNF1related kinase 2 (SnRK2 kinase). The activated SnRK2s phosphorylate downstream effectors, such as the basic leucine-zipper (bZIP) TFs called ABFs/AREBs and ABI5, which bind to ABA-responsive element (ABRE) in the promoters of many ABA-induced genes (e.g., LEA genes), thereby switching on the stress-response programs (Fujii and Zhu 2009, Ma et al 2009, Melcher et al 2009, Nakashima et al 2009, Park et al 2009). 2.1.8 Promoter analysis of stress-inducible genes Promoter region of abiotic stress-responsive Cor/LEA genes contain several conserved motifs as cis-acting elements functioning in stress-responsive transcription, such regulatory system include both ABA –dependent and –independent signaling pathways (Yamaguchi- Shinozaki and Shinozaki 2005). A conserved motif, PyACGTGGC, was first identified as a Cis-acting element named ABRE (ABA- responsive element) in promoter 12 region of ABA responsive genes such as wheat Em gene and rice Rab16 gene. A number of ABRE-binding proteins are members of bZIP (basic-domain leucine zipper) -type DNAbinding proteins, such as Arabidopsis bZIP-type ABF (ABRE-binding factor)/ AREB (ABAresponsive element binding) proteins, which binds to ABRE and activate ABA-dependent stress responsive gene expression (Choi et al 2000, Uno et al 2000). Another functional cisacting element of the Cor genes, i.e. the CCGAC motif known as CRT (C-repeat)/DRE (dehydration responsive element) sequences play a critical role in promoter function of Arabidopsis COR15A and RD29A. Arabidopsis CBF (CRT binding factor)/ DREB1 (DRE binding protein1) and DREB2 recognize CRT/DRE. The transcription factor’s CBF/DREB1 and DREB2 gene expression can be induced either by drought stress or by low temperature and both activate the expression of gene possessing a CRT/DRE cis-element. Overexpression of Arabidopsis DREB2A results in significant drought stress tolerance but only slight freezing tolerance in transgenic plants (Sakuma et al 2006a). DREB2A positively regulates expression of many abiotic stress-responsive genes possessing DRE sequences in their 5’ upstream regions. Many cereals DREB2 homologs have been identified in rice, maize, pearl millet, barley and wheat (Dubouzet et al 2003, Shen et al 2003, Aggarwal et al 2004, Xue and Loveridge 2004, Egawa et al 2006, Qin et al 2007,). Although among these proteins OsDREB2A, PgDREB2A and TaDREB1 bind to DRE sequences. The barley HvDRF1 gene also belongs to the CBF/DREB family (Xue and Loveridge 2004). The HvDRF1 transcription factor binds preferentially to a CT-rich element called a DRF1E motif (T(T/A)ACCGCCTT) rather than to CRT/DRE. A wheat DREB2 homolog Wdreb2, shows quite a high similarity to HvDRF1 and its transcription is enhanced by low temperature, drought, salt and ABA (Egawa et al 2006). Wheat ABA-responsive Cor/LEA genes have been characterized. Expression pattern of four Cor/LEA Wdhn13, Wrab17, Wrab 18 and Wrab 19 correspond well to the expression profile of Wdreb2 under low temperature, drought, salt and ABA (Egawa et al 2006). Recently Kobayashi et al (2008) analyzed the direct relationship between WDREB2 and the four Cor/LEA genes in development of abiotic stress tolerance. Recent approaches, especially the use of ABA-deficient and ABA-insensitive mutants, have shown that signaling of water stress may be understood in two major pathways: the ABA -dependent and -independent pathways. The ABA -dependent pathway may also have two different routes: i.e. requiring new protein synthesis or not. In the route where new protein synthesis is not required, the promoter domain in all ABA-responsive genes has an ABA-responsive element ABRE (PyACGTGGC where Py indicates a pyrimidine base, C or T). When ABRE is bound with its corresponding bzip family of 13 transcription factors such as EmBP-1, it can lead to ABA-induced gene expression. An example is the ABA mediated induction in Arabidopsis of the dehydration-responsive rd29B gene, which possesses two ABREs essential for its expression. Two bzip transcription factors, AREB1 and AREB2, are involved in the ABA-mediated expression of rd29B probably via phosphorylation mediated by an ABA activated protein kinase (Uno et al 2000). In the route where new protein synthesis is required for the ABA induced gene expression, de novo production of new proteins is the prerequisite. Such genes have no ABREs and its ABA-responsive elements combine with the MYC family transcription factors. The activation of the genes for MYC family transcription factors and the synthesis of the transcription factors should precede the activation of any ABA-initiated genes. Some genes may be induced by both the ABA-dependent and the ABA-independent pathways. Gene rd29A is an example; its promoter has two types of regulatory cis- elements, one ABAresponsive and the other independent of ABA. The independent cis -element has a conserved 9-basepair motif, TACCGACAT, also known as DRE/C-repeat element. 2.2 Antioxidant metabolism 2.2.1 Reactive oxygen species and their production sites Exposure of plant to unfavorable environmental conditions such as vicissitude of temperature, high light intensity, water availability, salt stress can increase the production of reactive oxygen species such as singlet oxygen, superoxide radical, hydrogen peroxide and hydroxyl radical. Plant posses enzymatic and non –enzymatic mechanisms for scavenging ROS. The enzymatic mechanism design to minimize the production of O2-. and production of H2O2.The enzymes overproduced so far include superoxide dismutase (SOD), Ascorbate peroxidase (APX), Glutathione reductase (GR). Superoxide radical is regularly synthesized in chloroplast and mitochondria though small quantity is also reported to produced in microbodies. Scavenging of superoxide by SOD results in the production of H2O2 which is removed by APX and CAT. However both O2-.and H2O2 are not as toxic as OH- which is formed by combination of O2-.and H2O2 in the presence of trace amount of Fe3+ by HaberWeiss reaction. Hydroxyl radical can damage chlorophyll, protein, DNA, lipids and other macromolecules that fatally affecting plant metabolism and ultimately growth and yield. A schematic presentation of production and scavenging of O2-. ,H2O2 and OH mediated lipid peroxidation and glutathione mediated stabilization of lipids are present in figure 2.2. 14 Fig. 2.2: Production and scavenging of ROS. 2.2.2 ROS scavenging enzymatic antioxidants 2.2.2.1 Superoxide dismutase (SOD) SOD has been first line of defense against the toxic effects of elevated levels of ROS. The SOD removes superoxide anion O2-. by catalyzing its dismutation, one O2-. being reduced to H2O2 and another oxidized to O2 . It removes O2.- and hence decrease the risk of OH. formation via metal catalyzed reaction. This reaction is 10,000 fold faster than spontaneous dismutation (Mittler 2002). SODs are classified by their metal cofactors into three known types, which are localized in different cellular compartments: The Mn-SOD is found in mitochondria, Cu/Zn-SOD found in cytosolic fraction and also in chloroplast of higher plants (Rio et al 2003). The Fe-SOD isozymes, often not detected in plants, are usually associated with chloroplast compartments when present. There has been reported the significant variation in the level of SOD under abiotic stress. Significant increase in SOD activity under salt stress has been observed in various plants viz. mulberry (Harinasut et al 2003) Cicer arietinum (Kukreja et al 2005) and Lycopersicon esculentum (Gapinska et al 2008). Eyidogan and Oz (2005) noted three SOD activity bands (MnSOD, FeSOD and Cu/ZnSOD) in Cicer arietinum under salt stress. Wang and Li (2008) studied the effect of water stress on the activities of total leaf SOD and chloroplast SOD in Trifolium repens L. and reported significant higher increase in SOD activity under water stress. O2-. + O2-. + 2H+ O2 + H2O2 2.2.2.2 Catalase (CAT) Catalase is a tetrameric heme containing enzyme with the potential to directly dismutate H2O2 into H2O and O2 and is indispensable for ROS detoxification during stressed 15 conditions (Garg et al 2009). CAT has one of the highest turnover rates for all enzymes: one molecule of CAT can convert 26 million molecules of H2O2 into H2O and O2 per minute. CAT is important in the removal of H2O2 generated in peroxisomes by oxidases involved in ß-oxidation of fatty acids, photorespiration and purine catabolism. The CAT isozymes have been studied extensively in higher plants. Maize has 3 isoforms (CAT1, CAT2 and CAT3), found on separate chromosomes. CAT1 and CAT2 are localised in peroxisomes and the cytosol, whereas, CAT3 is mitochondrial. It has also been reported that apart from reaction with H2O2, CAT also react with some hydroperoxides such as methyl hydrogen peroxide (MeOOH). Pretreatment of rice seedlings with H2O2 under non-heat shock conditions resulted in an increase in CAT activity. Similarly, increase in CAT activity in Cicer arietinum roots following salinity stress was noted. Stoilova et al (2010) reported increased CAT activity in wheat under drought stress but it was higher especially in sensitive varieties. In another study, Sharma and Dubey (2005) reported a decrease in CAT activity in rice seedlings under drought stress. 2H2O2 2H2O + O2 2.2.2.3 Ascorbate peroxidase (APX) APX is thought to play the most essential role in scavenging ROS and protecting cells in higher plants, algae, euglena and other organisms. APX is involved in scavenging of H2O2 in water- water and ASH-GSH cycles and utilizes ascorbate (ASH) as the electron donor. The APX family consists of at least five different isoforms including thylakoid (tAPX) and glyoxisome membrane forms (gmAPX), as well as chloroplast stromal soluble form (sAPX), cytosolic form (cAPX). APX has a higher affinity for H2O2 (mM). Significant increase in APX activity was noted under water stress in three cultivars of Proteus vulgaris (Zlatev et al 2006) and Proteus asperata (Yang et al 2008). Sharma and Dubey (2005) found that mild drought stressed plants had higher chloroplastic- APX activity than control grown plants but the activity declined at the higher level of drought stress. H2O2 + Asc D-Asc + 2 H2O 2.2.2.4 Guaiacol peroxidase (POD(GPX)) APX can be distinguished from plant-isolated guaiacol peroxidase (POD(GPX)) in terms of differences in sequences and physiological functions. (POD(GPX))decomposes indole-3-acetic acid (IAA) and has a role in the biosynthesis of lignin and defense against biotic stresses by consuming H2O2. GPOX prefers aromatic electron donors such as guaiacol and pyragallol usually oxidizing ascorbate at the rate of around 1% that of guaiacol. The activity of GPOX varies considerably depending upon plant species and stresses conditions. It increased in Cd exposed plants of Triticum aestivum (Milone 2003), Arabidopsis thaliana 16 (Cho et al 2005) and Ceratophyllun demersum (Arvind et al 2003). Radotic et al (2000) noted an initial increase in GPOX activity in spruce needles subjected to Cd stress and subsequent Cd-treatments caused a decline in the activity. A concomitant increase in GPOX activity in both the leaf and root tissues of Vigna radiate. H2O2 + 2GSH GSSG + 2 H2O ROOH + 2 GSH GSSG + ROH + H2O 2.2.2.5 Glutathione reductase (GR) GR is a flavo-protein oxidoreductase, found in both prokaryotes and eukaryotes (Romero-Puertas et al 2006). It is a potential enzyme of the ASH-GSH cycle and plays an essential role in defense system against ROS by sustaining the reduced status of GSH. It is localized predominantly in chloroplasts, but small amount of this enzyme has also been found in mitochondria and cytosol. GR catalyzes the reduction of GSH, a molecule involved in many metabolic regulatory and antioxidative processes in plants where GR catalyses the NADPH dependent reduction of disulphide bond of GSSG and is thus important for maintaining the GSH pool (Reddy et al 2006, Chalapathi et al 2008). Actually, GSSG consists of two GSH linked by a disulphide bridge which can be converted back to GSH by GR. GR is involved in defense against oxidative stress, whereas, GSH plays an important role within the cell system, which includes participation in the ASH-GSH cycle, maintenance of the sulfhydryl (eSH) group and a substrate for GSTs (Reddy et al 2006). GR and GSH play a crucial role in determining the tolerance of a plant under various stresses (Chalapathi et al 2008). GSSG + NADPH NADP+ + 2 GSH 2.2.3 Non-enzymatic antioxidants Different abiotic stress factors may provoke osmotic stress, oxidative stress and protein denaturation in plants, which lead to cellular adaptive responses such as accumulation of compatible solutes, induction of stress proteins, and acceleration of reactive oxygen species scavenging systems (Zhu 2002). One of the most common stress responses in plants is overproduction of different types of compatible organic solutes (Serraj and Sinclair 2002). Compatible solutes are low molecular weight, highly soluble compounds that are usually nontoxic at high cellular concentrations. Generally, they protect plants from stress through different courses, including contribution to cellular osmotic adjustment, detoxification of reactive oxygen species, protection of membrane integrity and stabilization of enzymes/proteins. Furthermore, because some of these solutes also protect cellular components from dehydration injury, they are commonly referred to as osmoprotectants. These solutes include proline, sucrose, polyols, trehalose and quaternary ammonium 17 compounds (QACs) such as glycine betaine, alaninebetaine, prolinebetaine, choline Osulfate, hydroxyprolinebetaine, and pipecolatebetaine. 2.2.3.1 Ascorbic acid The small antioxidant molecule vitamin C (L-ascorbic acid, AsA) fulfils essential metabolic functions in plants such as protection against oxidative damage (Smirnoff 2000), maintain the photosynthetic apparatus (Chen and Murata 2002) delay premature senescence of leaves and protect the chlorophylls. Several researchers provided that ascorbic acid plays an important role as plant growth regulator, stimulates the synthesis of mRNA, involved in flowering, and encourages the emergence of lateral buds. Recently it was shown that ascorbic acid is very important for the regulation of photosynthesis, flowering and senescence (Davey et al 2000, Barth et al 2006). It also fights against the negative effects of salt stress on tomato (Shalata and Neumann 2001) and wheat (Al- Hakimi and Hamada 2001, Athara et al 2008). In several studies conducted in the same way, it was found that only tolerant plants best respond to the application of exogenous AsA (Athara et al 2008) suggesting that the action of AsA to alleviate the harmful effects of salt stress is a variety dependent. However, the positive effect of AsA on the growth of wheat plants stress is caused by its action on the division and cell expansion (Smirnoff 2000). It seems that ascorbic acid increased mitotic activity of cells including the transition from G1 to S phases. Indeed several research conducted on the ascorbic acid, reported that they promote the cell elongation and cell proliferation (Blokhina et al 2002). Due to the fact that AsA serves as an important cofactor in the biosynthesis of many plant hormones, including ethylene, gibberellic acid (GA), and abscisic acid (ABA), one has to assume that the endogenous level of AsA will affect not only the biosynthesis, but also the levels and therefore the signaling of these molecules (Barth et al 2006). So it is likely to say that modulation of growth by AsA is may be induced by restoring the hormone equilibrium which is disturbed in salt stress conditions. The effect of AsA on the content of proline and soluble sugars can suggest that AsA probably improves growth of stressed plants, further to its antioxidant action, by intensification of their potential for osmotic adjustment and activities of growth (cell division and expansion). Indeed, several studies have shown that AsA plays an important role in improving plant tolerance to abiotic stress (Al-Hakimi and Hamada 2001, Shalata and Neumann 2001, Athara et al 2008). Moreover, the antioxidant effect exerted by the AsA on treated plants showed through the determination of the content of H2O2 and carotenoids, confirms the importance of antioxidant defence systems in plant tolerance to salinity (Agarwal and Pandey 2004, Sairam et al 2005). Finally, the study of the effect of NaCl on the leaf H2O2 content and analysis of correlations between the leaf H2O2 content and the growth of leaves, in the presence and absence of vitamin C, can suggest that the AsA effect 18 on promoting the growth under stress is probably due to its role in improving the antioxidant capacity (decrease of the H2O2 content). From the results mentioned above, it can be concluded that ascorbic acid counteracts with salinity by enhancing antioxidant capacity, improving growth and might also be by restoring the hormone balance which have to be verified in the future study. 2.2.3.2 Reduced glutathione Glutathione is a tripeptide (γ-glutamyl cysteinylglycine), which has been detected virtually in all cell compartments such as cytosol, chloroplasts, endoplasmic reticulum, vacuoles and mitochondria (Millar et al 2003) Glutathione is the major source of non-protein thiols in most plant cells. The chemical reactivity of the thiol group of glutathione makes it particularly suitable to serve a broad range of biochemical functions in all organisms. The nucleophilic nature of the thiol group also is important in the formation of mercaptide bonds with metals and for reacting with selected electrophiles. This reactivity along with the relative stability and high water solubility of GSH makes it an ideal biochemical to protect plants against stress including oxidative stress, heavy metals and certain exogenous and endogenous organic chemicals (Millar et al 2003, Foyer et al 2005). Glutathione takes part in the control of H2O2 levels (Shao et al 2006). The change in the ratio of its reduced (GSH) to oxidized (GSSG) form during the degradation of H2O2 is important in certain redox signaling pathways. It has been suggested that the GSH/GSSG ratio, indicative of the cellular redox balance, may be involved in ROS perception. Reduced glutathione (GSH) acts as an antioxidant and is involved directly in the reduction of most active oxygen radicals generated due to stress. There was a report reporting that glutathione, an antioxidant helped to withstand oxidative stress in transgenic lines of tobacco (Rio et al 2006). 2.2.3.3 Proline Amino acid proline is known to occur widely in higher plants and normally accumulates in large quantities in response to environmental stresses (Ozturk and Demir 2002, Hsu et al 2003, Kavi Kishore et al 2005). In addition to its role as an osmolyte for osmotic adjustment, proline contributes to stabilizing sub-cellular structures (e.g. membranes and proteins), scavenging free radicals, and buffering cellular redox potential under stress conditions. It may also function as a protein compatible hydrotrope alleviating cytoplasmic acidosis, and maintaining appropriate NADP+/NADPH ratios compatible with metabolism. Also, rapid breakdown of proline upon relief of stress may provide sufficient reducing agents that support mitochondrial oxidative phosphorylation and generation of ATP for recovery from stress and repairing of stress-induced damages. 19 Furthermore, proline is known to induce expression of salt stress responsive genes, which possess proline responsive elements (e.g. PRE, ACTCAT) in their promoters (Satoh et al 2002, Ohno et al 2003, Chinnusamy et al 2005). In response to drought or salinity stress in plants, proline accumulation normally occurs in the cytosol where it contributes substantially to the cytoplasmic osmotic adjustment. Furthermore, It was determined that, in response to water deficit, increased concentration of proline in maize root apical meristem was paralleled with increased concentration of abscisic acid. In plants, the precursor for proline biosynthesis is l-glutamic acid. Two enzymes, pyrroline-5-carboxylate synthetase (P5CS) and pyrroline-5-carboxylate reductase (P5CR) play major roles in proline biosynthetic pathway. Transgenic tobacco plants over-expressing P5CS have shown increased concentration of proline and resistance to both drought and salinity stresses. Fig. 2.3: Biosynthetic pathway of proline in higher plants Accumulation of proline under stress in many plant species has been correlated with stress tolerance and its concentration has been shown to be generally higher in stress-tolerant than in stress-sensitive plants In wheat, an assessment of the effects of drought stress on proline accumulation in a drought-tolerant and a drought-sensitive cultivar revealed that the rate of proline accumulation and utilization was significantly higher in the drought-tolerant cultivar (Nayyar and Walia 2003). In light of the available information, however, it seems that proline accumulation in plants is mediated by both ABA-dependent and ABA -independent signaling pathways (Zhu 2001b, 2002). ABA is known to mediate signals in plant cells subjected to environmental stresses. These signals can bring about expression of stress-related genes followed by synthesis of compatible osmolytes such as proline (Kavi Kishore et al 2005). Furthermore, ABA accumulation in plants in response to osmotic stress has been determined to regulate expression of P5CS gene, which is involved in proline biosynthesis (Xiong et al 2001). Despite the presence of a strong correlation between stress tolerance and accumulation of proline in higher plants, this relationship may not be universal. For example, in rice plants grown under salt stress, accumulation of proline in the leaf was deemed to be a symptom of 20 salt injury rather than an indication of salt tolerance. Similarly, assessment of proline accumulation and distribution during shoot and leaf development in two sorghum genotypes contrasting in salt tolerance suggested that proline accumulation was a reaction to salt stress and not a plant response associated with tolerance (de-Lacerda et al 2003) and yet in another study, under salt stress, sensitive rice cultivars accumulated greater amounts of proline than did the tolerant genotypes. However, further studies are needed to determine whether the relationship between stress tolerance and accumulation of proline is species-specific or if it can be altered by experimental conditions. 2.2.3.3.1 Exogenous application of proline Exogenous application of proline can play an important role in enhancing plant stress tolerance. This role can be in the form of either osmoprotection or cryoprotection. Proline can also protect cell membranes from salt-induced oxidative stress by enhancing activities of various antioxidants (Yan et al 2000). For example, growth of tobacco suspension cells under salt stress was promoted by exogenous application of 10mM proline, which was proposed to be due to proline action as a protectant of enzymes and membranes (Okuma et al 2000). In soybean cell cultures maintained under salt stress, exogenous application of proline increased activities of superoxide dismutase and peroxidase, which normally contribute to increased salt tolerance (Yan et al 2000, Hua and Guo 2002). In barley embryo cultures under saline conditions, exogenous application of proline resulted in a decrease in Na+ and Cl− accumulations and an increase in growth. In contrast to the above findings on beneficial effects of exogenous application of proline, there are a few reports cautioning its use. For example, foliar application of proline to rice plants growing under saline conditions did not change concentrations of either Na+ or Cl− in the leaves. Moreover, in Arabidopsis plants exogenous application of proline was suggested to cause damages to ultra-structures of chloroplast and mitochondria. 2.2.4 Hydrogen peroxide (H2O2) H2O2 as one of the major and most stable ROS that regulate basic acclimatory, defense and developmental processes in plants. The addition of single electron to the molecular oxygen reduces it to the superoxide anion radical. The superoxide is a free radical. It is relatively unstable, either converted back to O2 or in reaction with proton, to H2O2, either spontaneously or in a reaction catalyzed by Superoxide dismutase (SOD). Because of this and longer half life of H2O2 than that of superoxide anion radical, H2O2 is more likely to be a long –distance signaling molecule. 21 2.2.4.1 Cellular and extracellular sources of H2O2 Hydrogen peroxide is produced not only through the disproportionation of superoxide, but also due to the reduction of O2-. by reductant (X) such as ascorbate, thiols, ferredioxins and others. Thus hydrogen peroxide at cellular levels is strictly linked with the generation of superoxide. Nevertheless, other oxidases such as glycolate oxidase, glucose oxidase, amino-acid oxidases release H2O2 following oxidation of their respective substrate. 2.2.4.1.1 Chloroplast The photosynthetic electron transport (PET) chain in the chloroplast is responsible for H2O2 production. The electron transport components of (PET) are auto-oxidizable which cause limiting availability of NADP, superoxide anion radicals can be formed (Dat et al 2000, Foyer & Noctor 2003). ‘Mehler’ reaction’ is considered as primary and the most powerful source of H2O2/ROS in chloroplast. Secondly, the H2O2 production in chloroplasts is catalysed by SOD isoforms (Alsher et al 2002). The reduction of O2-. by ascorbate (Asc) and reduced glutathione (GSH) does not contribute much to the production of H2O2 in chloroplast. 2.2.4.1.2 Peroxisomes The main function of peroxisomes in plant cell is photorespiration, which is associated with generation of H2O2 (Dat et al 2000, Wingler et al 2000). Besides glycolate oxidation H2O2 can be generated via ß-oxidation of fatty acids and oxidation of other substrates (Dat et al 2000), but significance of this process in H2O2 production in comparision to glycolate oxidation in C3 plants appear minor (Foyer and Noctor 2003). 2.2.4.1.3 Mitochondria In plant mitochondria superoxide anion radical production occurs mainly at two sites of the electron transport chain: NAD(P)H dehydrogenases (complex I) and cytochrome bc1 complex (complex III) (Moller 2001). This process results in the formation of H2O2, primarily through the action of a mitochondrion-specific manganese SOD (Rhoads et al 2006). The amount of H2O2 produced in plant mitochondria is less than that of chloroplasts or peroxisomes when exposed to light (Foyer and Noctor 2003), but in dark or in non-green tissues, mitochondria can be major source of ROS (Rhoads et al 2006). 22 Fig. 2.4: Production of ROS in mitichondria 2.2.4.1.4 Other sources of H2O2 In the cytoplasm, the electron transport chain associated with endoplasmic reticulum is the main source of H2O2/ROS. The cytoplasm cannot be regarded as a major source of H2O2 in plant cell but may act as sink for hydrogen peroxide leaking from other cellular component. NADPH oxidase at the plasma membrane in the plant cell is the most intensively studied oxidase system. The system catalyzes the production of O2- by one electron reduction of O2 using NADPH as electron donor (Desikan et al 2003, Mahalingam & Federoff 2003, Apel & Hirt 2004). The superoxide anion radical is most likely located in the appoplastic space and is converted to H2O2 (Karpinska et al 2001, Bolwell et al 2002). Many enzymes of extracellular membrane, such as pH-dependent cell wall peroxidases, germins, germin like oxalate oxidases and amine oxidases have been proposed as a source of H2O2 in apoplast (Bolwell et al 2002, Kacperska 2004). 2.2.4.2 H2O2 in stress conditions and as a signaling molecule H2O2 play a central role in responses to both biotic and abiotic stresses in plants. Most studies have reported increase in concentration after exposure to stress. The rate of H2O2 depends upon the strength and exposure of imposed stress (Neill et al 2002a, Kacperska 2004). Experiments with plant material have demonstrated that plant tissues can tolerate high concentrations of H2O2 in the range 10–2 × 105 μM. Moreover, plants pre-treated with H2O2 were more resistant to excess light and chilling stresses (Karpinska et al 2000, Yu 23 et al 2003). In the literature, the endogenous concentration of H2O2 is reported to lie in a wide range, ranging from nanomoles to several hundred micromoles of H2O2 per gram fresh mass (Veljovic-Jovanovic et al 2002). The relative stability and higher concentrations of H2O2 in plant cells could point to the fact that H2O2 plays a key role as a signal transduction factor. However, signal molecules are usually present in cells in very low concentrations; the relatively high level of H2O2 in plant tissues supports the assumption that H2O2 is not only a signaling molecule, but also plays a key role in primary plant metabolism. Moreover, H2O2 regulates the expression of various genes, including those encoding antioxidant enzymes and modulators of H2O2 production (Neill et al 2002a, 2002b). Hydrogen peroxide has also been shown to be an intercellular signal mediating systematic acquired resistance and systematic acquired acclimation. 2.2.4.3 H2O2 signaling during growth and development Recently, information on the role of H2O2/ROS as signal molecules regulating growth and is not only a stress signal molecule, but may also be an intrinsic signal in plant growth and development. H2O2 has shown to be involved in differentiation of cellulose rich cell wall. Recently it has been demonstrated that diminished extracellular Cu/Zn SOD expression in poplar trees regulate extracellular H2O2 level and plant development. Fig. 2.5 Reactive oxygen species (ROS), including hydrogen peroxide (H2O2), are among the important second messengers in abscisic acid (ABA) signaling in guard cells. To investigate specific roles of H2O2 in ABA signaling in guard cells, the effects of mutations in the guard cell-expressed catalase (CAT) genes, CAT1 and CAT3, and of the CAT inhibitor 3- 24 aminotriazole (AT) on stomatal movement has been examined. The cat3 and cat1 cat3 mutations significantly reduced CAT activities, leading to higher basal level of H2O2 in guard cells, when assessed by 2_,7_-dichlorodihydrofluorescein, whereas they did not affect stomatal aperture size under non-stressed condition. In addition, AT-treatment at concentrations that abolish CAT activities, showed trivial affect on stomatal aperture size, while basal H2O2 level increased extensively. In contrast, cat mutations and AT-treatment potentiated ABA-induced stomatal closure. Inducible ROS production triggered by ABA was observed in these mutants and wild type as well as in AT-treated guard cells. These results suggest that ABA-inducible cytosolic H2O2 elevation functions in ABA-induced stomatal closure, while constitutive increase of H2O2 does not cause stomatal closure (Jannat et al 2011). Fig. 2.6 2.2.5 Malondialdehyde (MDA) Estimation of malondialdehyde (MDA) amount, which is a secondary end product of polyunsaturated fatty acid oxidation, is widely used to measure the extent of lipid peroxidation as indicator of oxidative stress. Many studies have shown greater accumulation of MDA and H2O2 caused by salinity and drought (Sairam and Srivastva 2001). 25 2.3 Abscisic acid The plant hormone abscisic acid (ABA) was first named as such (Addicott et al 1968) a few years after its isolation from cotton fruits (Ohkuma et al 1963) as an abscission accelerating factor, and from sycamore leaves (Cornforth et al 1965) during a search for endogenous substances that induce dormancy. Although the role of ABA in fruit abscission and dormancy of woody plants remains unclear (Schwartz et al 2003), a vast body of evidence supports the implication of this ubiquitous hormone in essential plant processes such as seed maturation, desiccation, dormancy and germination (Hoth et al 2002, Seki et al 2002, Leonhardt et al 2004). The plant hormone abscisic acid has long been known to be involved in the responsiveness of plants to various environmental stresses including drought, salinity, and low temperature. ABA production is increased in tissues during these stresses and this cause a variety of physiological effects, including stomata closure in leaves. Previous evidence showed that an elevation of cytisolic Ca, an increase in pH and a reduction in K and Cl and organic solute content in both guard cells surrounding the stoamtal pore, are downstream elements of ABA-induced stomatal closure. In addition, cADP-Rib, phospholipase C and phospholipase D have been identified as siganlling molecules in the ABA response and exerting their effects by regulating cytosolic Ca concentration and inward K channels. Furthermore, Ca channels and anion channels at the plasma membrane of stomatal guard cells are activated by hyperpolarization and ABA (Pei et al 2000) and increase in Ca concentration resulting from activation of Ca channels leading to Ca influx is known to inactivate inward-rectifying K channels, biasing the plasma membrane for solute efflux, which drives stomatal closure. 2.3.1 ABA as a long distance signal mediating whole plant response to drought and salt stresses Stress ABA produced in dehydrated roots in drying soil is transported to the xylem and regulates stomatal opening and leaf growth in roots and shoots. This mechanism is modified by ionic conditions and pH in xylem. Hartung et al (2002) have shown that pH changes play a central role in the ABA redistribution in leaf tissues and control the stomata at times when no significant changes in ABA concentration are detected in the xylem. It appears that plants have evolved two responses to soil drying. Initial soil drying may be sensed by part of the root system and a root-sourced ABA in the xylem may regulate the stomatal conductance such that water loss in the shoots may be reduced and leaf water deficit can be avoided. When the soil drying is prolonged and becomes more severe, the shoot water deficit becomes unavoidable and some older leaves may wilt, perhaps because of a weak 26 hydraulic link with the main stem or a weaker control of stomatal conductance. Such leaf wilting should account for the much accelerated ABA concentration in the xylem and much more severe stomatal inhibition that can be observed in the young leaves. The effect of ABA on the regulation of plant growth is rather complex. Early results showed that stress ABA is a regulator of shoot growth and development under water stress and shoot growth can be inhibited when xylem ABA is increased as a result of soil drying. More recent results have shown, however, differences in response exist between root and shoot. Sharp and his group have shown that maintenance of root growth under low water potential is a function of enhanced ABA accumulation in roots. They have shown that ABA’s role may involve the inhibition of ethylene production, which is a growth inhibitor under stress (Spollen et al 2000, Sharp and LeNoble 2002) even suggested that stress ABA may also be helpful to maintain some limited shoot growth as well as root growth under water deficit condition would enhance drought tolerance. More recent experiments have supported the notion that ABA has dual roles in its physiological regulation (Cheng et al 2002, Finkelstein et al 2002). Its inhibitive role functions when it is accumulated in large amount under stress to help plant survival through inhibition of processes such as stomatal opening and plant size expansion. Its promoting role, when it is at low concentration and at more ‘normal’ condition, has been shown essential for vegetative growth in several organs e.g., primary root growth (Spollen et al 2000) and post-germination seedling development. In addition to osmotic stress, salinity imposes on plants other stresses such as ion toxicity, as a result of ion entry in excess of appropriate compartmentation, and nutrient imbalances, as commonly seen in the displacement of potassium by sodium. The main damage to plants, however, could result from osmotic stress imposed externally due to high ion concentrations in the soil or internally when excess salt uptake resulted in high salt accumulation in the intercellular spaces. In this regard, the damage caused by salinity is mainly due to altered water relations; thus plant responses to salinity and water deficit are closely related with overlapping mechanisms. Exposure of plants to salinity is known to induce a proportional increase in ABA concentration that is in most cases correlated with leaf or soil water potential, suggesting that salt-induced endogenous ABA is due to water deficit rather than to a specific salt effects. 2.3.2 Cell signaling from stress perception to ABA accumulation It has been proposed that for the cell signaling of ABA accumulation the perception mechanism may be different not only between water and salt stress but also between leaf and root tissues. The evidence for such a claim is that prevention of cell dehydration could completely diminish the osmotic stress-induced ABA accumulation in leaf cells, while such 27 treatment had no visible effect on root tissues. As roots usually contain a lower level of xanthophylls than leaves, ABA synthesis in roots is more likely to be limited by the size of the precursor pools than in leaves. In addition, the osmosensor may be more abundant in roots than in leaf cells (Jia et al 2002b), while the dehydration-induced production of ABA is usually observed in shoots. Seo et al (2000) found that one of the key control points for ABA accumulation, ABA-aldehyde oxidase, exists in different isoforms in the roots and leaves of Arabidopsis and is encoded by different genes in the two tissues. 2.3.3 Abscisic acid and antioxidant defense in plant cells 2.3.3.1 ABA- induced ROS generation In ABA-induced stomatal closure of guard cell of Arabidopsis, ABA treatments induced a rapid increase in the production of H2O2 (Pei et al 2000). Treatment with 1 micromole/L ABA increased the production of H2O2 by 36.8% and treatment with 50 micromole/L ABA increased by 49%. Similar results were also observed in ABA-induced stomatal closure of guard cell of Arabidopsis, Vicia faba (Zhang et al 2001) rice roots (Lin and Kao 2001) and in maize embryo cells (Guan et al 2000). Accumulation of apposplastic H2O2 is involved in the induction of the chloroplastic and cytosolic antioxidant enzymes. These results suggest that H2O2 produced at a specific cellular site could coordinate the activities of antioxidant enzymes in different subcellular compartments (Hu et al 2005). Although recent studies have shown that ABA-induced ROS production increase the total leaf activity of SOD, CAT, APX and GR in maize (Jiang and Zhang 2002a, b), it is not clear whether ABA induced ROS production co-ordinates the different subcellular activities of antioxidant enzymes. 2.3.3.2 Expression of Genes Encoding Antioxidant Enzymes Induced by ABA It has been documented that ABA can induce the expression of antioxidant genes encoding Cu/Zn-superoxide dismutase (SOD), Mn SOD and Fe SOD and catalase (CAT) (Guan et al 2000) in plants. The expressions of Sod and Cat genes in response to ABA depend on plant species or varieties. For example, in rice seedlings, ABA induced an increase at the transcript levels of SodA1, which encodes Mn-SOD, SodB, which encodes FeSOD, and SodCc1 and SodCc2, which encodes cytosolic Cu/Zn-SODs, in a dose-dependent manner. But in tobacco BY-2 cell suspensions, ABA treatments only increased the accumulation at the transcript level of Mn-SOD gene, and did not change the transcript level of Fe-SOD gene and reduced that of cytosolic Cu/Zn- SOD gene. In an inbred maize line, the transcript of Cat3 was induced 3.6-fold by ABA in coleoptiles and 3.3-fold in mesocotyls. In another inbred maize line, however, the Cat3 transcript decreased after 2h of ABA treatment 28 in leaves. Moreover, the expressions of Sod and Cat genes are different in response to ABA at different developmental stages in plants. For example, in maize, the transcripts of Sod4 and Cat1 accumulated in response to ABA in developing and germinating embryos, and in young leaves; the Sod4A transcript showed no increase in response to ABA in developing and germinating embryos, but increased in young leaves; Cat2 and Cat3 transcripts were upregulated only at very high ABA concentrations (10-3 mol/L) during late embryogenesis and in response to various concentrations of ABA in germinating embryos, and the Cat2 transcript increased in response to ABA and the Cat3 transcript decreased in young leaves. These results suggest that the Sod and Cat genes in maize are regulated by ABA in a multilayered fashion. ABA not only induces the increase in activities of these antioxidant enzymes but also induces the increase in contents of non-enzymatic antioxidants such as ascorbate, reduced glutathione, alpha-tocopherol and carotenoids (Jiang and Zhang 2001, 2002a). These results suggest that ABA can induce the capacity of whole antioxidant defense systems including enzymatic and non-enzymatic constituents in plants. Pretreatment with ABA enhances the capacity of antioxidant defense systems in plants exposed to environmental stress such as chilling, high temperature, NaCl stress and water stress (Jiang and Zhang 2002a). The increase in antioxidant defense system is closely related to stress tolerance. A recent study provides a genetic evidence for involvement of ABA in the protection against oxidative damage in Arabidopsis exposed to heat stress. ABA reduces the level of lipid peroxidation and enhanced survival in Arabidopsis. 2.3.3.3 ABA induced modulation of metabolic and redox control pathways Besides the up-regulation of pathways related to the biosynthesis of compatible solutes as a response to ABA treatment, metabolic profiling indicated that specific antioxidants, particularly alpha-tocopherol and L-ascorbic acid, were accumulated at high level in ABA-treated seedlings compared to appropriate controls. The transcriptions of genes involved in alpha-tocopherol biosynthesis were co-ordinately upregulated and appeared to be integrated into a network of reactions controlling the level of reactive oxygen species. The present study was aimed at providing evidence regarding metabolite levels that are affected by the treatment with ABA and to provide comparison with metabolite pool that are accumulated under cold and drought stress (which are possibly mediated by ABA). The observed increase in the level of antioxidants alpha-tocopherol (1.7 fold increase) and Lascorbic acid (3.1 fold increase) is consistent with an early study reporting a dose-dependent alpha-tocopherol formation in ABA treated maize seedlings (Jiang and Zhang 2001). 29 In guard cells of water-stressed seedlings, ABA causes the closing of stomata, mediated by the second messenger Ca2+, thus reducing transcriptional water loss. ABAinduced increases in Ca2+ levels are facilitated by influx through plasma membrane Ca2+ channel and release from internal stores (MacRobbie 2000) Ca2+ channels are activated by reactive oxygen species (Pie et al 2000 and Kohler et al 2003) the level of which increase as the part of ABA signaling cascade (Guan et al 2000, Jiang and Zhang 2001, Zhang et al 2001 and Kwak et al 2003). ABA treatment is known to induce the transcription of the gene encoding cytosolic ascorbate peroxidase (apx2), possibly to adjust the cytosolic redox poise (Chang et al 2004, Davletova et al 2005), this finding was confirmed by quantitative realtime PCR analysis (approx. 4.5- fold up-regulated in ABA-treated seedlings at 24h) there was up-regulation of genes encoding the cytosolic enzyme involved in ROS-scavenging ascorbate-glutathione cycle (APX, up 4.5 fold), monodehydro ascorbate (MDAR, up 1.8 fold) reductase, glutathione reductase (GR, up 2-fold) and dehydro ascorbate reductase (DHAR, up 1.6-fold). Based on gene expression observed in the experiments, the ABA treatment led to the activation of cytosolic transcriptional redox control network in Arabidosis seedling. 30 CHAPTER III MATERIAL AND METHODS 3.1 Plant material The experiments were conducted on roots and shoots of seedlings of two wheat cultivars PBW343 (drought sensitive) and C306 (drought tolerant), exposed to different treatments. Seeds were obtained from Department of Plant Breeding and Genetics, PAU, Ludhiana. Chemicals used for this study, were purchased from different companies like Bovine serum albumin (BSA), glutathione (oxidized), NADPH, ascorbic acid, ninhydrin, Nitro blue tetrazolium chloride (NBT), Diethylpyrocarbonate (DEPC), guaiacol, pyrogallol were purchased from SRL Pvt. Ltd. Mumbai (India); bipyridyl, Thiobarbituric acid (TBA) were purchased from Himedia Laboratories Pvt. Ltd. Mumbai (India); Hydrogen peroxide, mannitol, sodium chloride from Fisher-Scientific Mumbai (India); Oligo(dT)18 and agarose were purchased from G-Biosciences Pvt. Ltd. Noida (India). TRIsoln, M-MuLV Reverse Transcriptase, Human Placental RNase Inhibitor, Taq DNA Polymerase, DNase I were purchased from GeNei Pvt. Ltd. Banglore (India). Primers used in present study were obtained synthesized by Sigma Aldrich Chemicals Pvt. Ltd. Banglore (India). 3.2 Germination of seeds under different treatments For germination, seeds were washed by adding 2-3 drops of laboratory detergent and then seeds were disinfected by immersing in 0.1% mercuric chloride solution for 4-5 minutes. Seeds were then washed 4-5 times with sterilized distilled water. After sterilization, seeds were kept in incubator at 25 ºC for overnight for stratification (Tan et al 2008). On next day, seeds were placed on moistened filter paper spreaded in sterilized petri dishes (10 cm diameter). Autoclaved distilled water was used to moisten filter paper. Plates were kept in dark at 25 ºC for 4 days. During this period, seeds were rewatered regularly using autoclaved distilled water. Stress was applied on 4th day, where for ABA treatments, filter paper was moistened with 20 µM ABA, for WS treatments filter paper was moistened with 6% mannitol, for combined stress treatments filter paper was moistened with 20 µM ABA plus 6% mannitol. Control seedlings were kept growing on autoclaved distilled water. Plates were similarly incubated in dark at 25 ºC for another 1-3 days. Data was collected from 3 stages; 24h, 48h, 72h after stress treatment and compared with similarly old control seedlings grown on water as well as with 0h stage taken on 4th day before applying stress treatment. 3.3 Growth measurement Growth was measured as shoot length, fresh weight, dry weight and water contents in roots and shoots of germinating seedlings. 3.3.1 Measurement of shoot length Shoot length (cm) was measured with scale (of 50 seedlings) at 0h, 24h, 48h and 72h stages. 3.3.2 Measurement of fresh weight and dry weight Fresh roots and shoots of 25 seedlings were weighed separately in triplicates. Fresh tissues were subjected to drying for 24hours at 80 ºC in oven and their dry weights were recorded. 3.3.3 Measurement of water content (g H2O/g DW) Water contents of roots and shoots were determined in triplicates by using equation: WS=FW- DW/DW; where FW refers to fresh weight and DW refers to dry weight. 3.3.4 Determination of R/S ratio Root to shoot ratio was determined in triplicates by using root and shoot dry weights of 25 seedlings each. Statistical analysis Data was calculated as Mean ± S.D and analyzed by Duncan’s Multiple Test at P ≤ 0.05 to test for statistical differences among samples using DSAASTAT software version 1.101. 3.4 Enzymatic Assays 3.4.1 Common extraction procedure for all antioxidant enzymes All antioxidant enzymes studied, were extracted by using common extraction procedure (as in Yang et al 2008) and were assayed by their specific procedures. Fresh tissue (root and shoot) of 250 mg was homogenized in 1.5 ml of ice cold buffer containing 75 µmole of potassium phosphate buffer (pH 7.0), 1.5 µmole of EDTA, 2% PVP, 0.05% triton-X- 100 using pre-chilled pestle-mortar. Homogenate was passed through layers of cheese cloth and then centrifuged at 10000 x g at 4 ºC for 15 minutes. Supernatant after centrifugation was used as enzyme extract in following assays. 3.4.2 Assay for Glutathione reductase (GR) Glutathione reductase catalyses the reduction of oxidized glutathione at the expense of 2 molecule of NADPH+ H+. GSSG + 2NADPH + H+ 2GSH + 2NADP+ 32 GR was assayed (as in Keles and Oncel 2002) in spectrophotometric cuvette at 25ºC in a reaction volume of 3 ml containing 150 µmole of potassium phosphate buffer (pH 7.0), 1.75 µmole of glutathione oxidized (GSSG), 1.75 µmole of NADPH+H+. All contents were mixed and reaction was started by adding 50-100 µl of enzyme extract. Change in absorbance was recorded for 3 minutes at the interval of one minute at 340 nm. Decrease in nmole of NADPH+H + min-1 was calculated using molar extinction coefficient of NADPH (6.22 mM-1 cm-1). The glutathione reductase was expressed as nmole of NADPH+H+ disappeared min-1 g-1 DW and specific activity was expressed as nmole of NADPH+H+ disappeared min-1 mg-1 protein. 3.4.3 Assay for Superoxide dismutase (SOD) SOD causes the conversion of superoxide anion to H2O2 in the presence of molecular O2 hence scavenges toxic superoxide anion. O2-. + O2-. + 2H+ O2 + H2O2 SOD was assayed (as in Gapinska et al 2008) at 25 ºC in spectrophotometric cuvette containing 2.5 ml reaction volume of 1.42% Triton X-100, 125 µmole of Tris HCl buffer pH 8.5, 62.5 nmole of NBT to which, 0.01 µmole of pyrogallol was added. Reduction of NBT was observed as change in absorbance at 550 nm for 2.5minutes at the interval of 30 seconds. This was taken as blank. For enzymatic assay, 100-200 µl enzyme extract was added in another similar above mentioned reaction after the addition of pyrogallol and change in absorbance was recorded similarly at 550 nm. Enzyme activity was expressed in SOD units where one SOD unit was defined as the amount of enzyme required to cause 50% inhibition of NBT reduction as opposed to 0% observed in blank. SOD activity was expressed as SOD units g-1 DW. SOD specific activity was expressed as SOD units mg-1 protein. 3.4.4 Assay for Ascorbate peroxidase (APX) APX detoxifies H2O2 to H2O by using reducing power of ascorbate. Ascorbate + H2O2 Monodehydroascorbate + H2O APX was assayed (as in Yang et al 2008) in reaction volume of 2.5 ml containing 125 µmole of potassium phosphate buffer of pH 7.0, 0.25 µmole of EDTA, 0.75 µmole of ascorbate, 2.91 µmole of H2O2 to which 50-100 µl of enzyme extract was added. Change in absorbance was recorded at 290 nm in spectrophotometric cuvette at 25 ºC for 2.5 minutes at the interval of 15 seconds. Decrease in µmole of ascorbate per minute was calculated using molar extinction coefficient of ascorbate of 2.8 mM-1 cm-1. APX activity was expressed as µmole of ascorbate disappeared min-1 g-1 DW and specific activity was expressed as µmole of ascorbate disappeared min-1 mg-1 protein. 33 3.4.5 Assay for Guaiacol peroxidase (GOPX) Guaiacol peroxidase detoxifies H2O2 by using reduction power of plant phenol guaiacol. Guaiacol + H2O2 tetraguaiacol + H2O GOPX was assayed (as in Simova-Stilova et al 2008) in reaction volume of 2.5 ml containing 250 µmole of potassium phosphate buffer of pH 6.5, 125 µmole of guaiacol, 80 µmole of H2O2, to which enzyme extract was added. Change in absorbace at 470 nm was recorded in spectrophotometric cuvette at 25 ºC for 2.5 minutes at the interval of 30 seconds. Change in µmole of tetraguaiacol per minute was calculated using molar extinction coefficient of tetraguaiacol of 26.6 mM-1 cm-1. Activity was expressed in µmole of tetraguaiacol appeared min-1 g-1 DW and specific activity was expressed as µmole of tetraguaiacol appeared min-1 mg-1 protein. 3.4.6 Assay for Catalase Catalase detoxifies H2O2 to H2O and O2 by following equation 2 H2O 2 H2O + O2 Catalase assay was performed (as in Yang et al 2008) in spectrophotometric cuvette at 25 ºC in reaction volume of 2.5 ml containing 125 µmole of potassium phosphate buffer of pH 7.0 and 62 µmole of H2O2, to which 50 µl of enzyme extract was added. Change in absorbance was recorded at 240 nm for 2.5 minutes at the interval of 15 seconds. Change in µmole of H2O2 per minute was calculated using molar extinction coefficient of H2O2 of 0.0394 mM-1 cm-1. Activity was expressed in µmole of H2O2 disappeared min-1 g-1 DW and specific activity was expressed as µmole of H2O2 disappeared min-1 mg-1 protein. Statistical analysis on enzymatic data: All enzymes were extracted in triplicates. Mean ± SD was calculated. Data was analyzed by Duncan’s Multiple Test at P ≤ 0.05 to test for statistical differences among samples using DSAASTAT software version 1.101. 3.5 Measurement of metabolites involved in antioxidant response of plants Ascorbate, H2O2, dehydroascorbate, proline, MDA contents were measured where ascorbate and proline were important non-enzymatic antioxidants in plant under stress. Ascorbate/ dehydroascorbate ratio, H2O2 level and MDA content were indicators of oxidative levels of plant tissues under stresses. 3.5.1 Extraction and estimation of H2O2 For extraction, 250 mg of fresh tissue was homogenized in 1.5 ml of ice cold 0.1% TCA. Homogenate was passed through layers of cheese cloth and then centrifuged at 10000 x g at 4 ºC for 15 minutes. Supernatant after centrifugation was used for estimation. 34 H2O2 was estimated (as in Alexieva et al 2001) by adding 0.5-1 ml of supernatant to 2 ml of reaction mixture containing 4 mmole of potassium iodide and 0.1 mmole of potassium phosphate buffer of pH 7.0. Test tubes were incubated at room temperature in dark for 1 hour. Absorbance was read at 390 nm against blank which carried distilled water in place of supernatant. The amount of H2O2 was calculated by preparing standard curve of 50200 nmole of H2O2. H2O2 was expressed as nmole of H2O2 g-1 DW. 3.5.2 Extraction and estimation of ascorbic acid 250 mg of fresh tissue was homogenized in 1.5 ml of ice cold 5% TCA. Homogenate was passed through layers of cheese cloth and then centrifuged at 10000 x g at 4 ºC for 15 minutes. Supernatant after centrifugation was used for estimation (as in Nobuhiko et al 1981). 500 µl of appropriately diluted supernatant was added to reaction volume of 2 ml containing 1% H3PO4, 0.05% FeCl3, 0.25% bipyridyl in ethanol. Blank was prepared with 500 µl of 5% TCA in place of supernatant. The reaction mixture was incubated at 37 ºC for 40 minutes. Absorbance was measured at 525 nm against blank. The quantity of ascorbic was determined by making standard curve of 5-25 µg of ascorbic acid. Ascorbic acid content was expressed as µmole of ascorbate g-1 DW. 3.5.3 Extraction and estimation of MDA 1g of tissue was homogenized in 2.5ml of ice cold 0.1% TCA. Homogenate was passed through layers of cheese cloth and then centrifuged at 10000 x g at 4 ºC for 15 minutes. Supernatant after centrifugation was used for estimation (as in Heath and Packer 1968). For estimation, 1 ml of appropriately diluted supernatant was mixed with 4 ml of solution containing 0.5% thiobarbituric acid and 20% TCA. Blank was prepared by taking 1 ml of 0.1% TCA in place of supernatant. Test tubes were kept at 100 ºC for 30 minutes then cooled down to room temperature. Contents were centrifuged at high speed and then read at 532 nm and 600 nm against blank. Absorbance at 600 nm was subtracted from absorbance at 532 nm. The MDA content was calculated by using molar extinction coefficient of malondialdehyde of 155 mM-1 cm1 and expressed as nmole of MDA g-1 DW. 3.5.4 Extraction and estimation of Dehydroascorbate 250 mg tissue was homogenized in 1.5 ml of 5% metaphosphoric acid and 1% thiourea. Homogenate was passed through layers of cheese cloth and then centrifuged at 10000 x g at 4 ºC for 15 minutes. Supernatant after centrifugation was used for estimation (as in Roe and Oesterling 1943). 35 For estimation 1 ml of appropriately diluted supernatant was mixed with 1 ml of reagent (containing 2% 2,4-dinitrophenyl hydrazine (DNPH), 0.4% thiourea. 0.05% CuSO4.5H2O in 9N H2SO4) and incubated at 37 ºC for 3 hours. After incubation the test tubes were transferred immediately on ice bath and 5 ml of 85% H2SO4 (cold) was added and test tubes were incubated at room temperature for another 30 minutes. The content was read at 530 nm against blank containing 5% meatphosphoric acid in place of supernatant. The content of dehydroascorbate was calculated by using standard curve of 20-80 µg of dehydroascorbate and expressed as µg of dehydroascorbate g-1 DW. 3.5.5 Extraction and estimation of Proline 250 mg tissue was homogenized in 1.5 ml of ice cold 3% sulphosalicylic acid. Homogenate was centrifuged at 10000 x g for 15 min at 4 ºC. Supernatant was collected and used for estimation (Bates et al 1973). Estimation was done by taking 1 ml of appropriately diluted supernatant with 3% sulphosalicylic acid and then adding 1 ml of acid ninhydrin reagent (0.31 g ninhydrin, 7.5 ml acetic acid and 5 ml 6M phosphoric acid) and 1 ml of glacial acetic acid. Test tubes were incubated at 100 ºC for 1 hour. After 1 hour the reaction was terminated on ice bath. Reaction mixtures were reacted with 4 ml toluene and mixed vigorously over vortex for 15-20 seconds. Test tubes were placed at room temperature for phase separation. The pink-red upper layer of chromophore containing toluene was read at 520 nm using toluene for blank. The quantity of proline was determined by standard curve prepared by taking different concentrations of proline ranging from 0.05 µM to 0.5 µM. Proline contents were expressed as µmole of proline g-1 DW. 3.5.6 Estimation of Protein Proteins were estimated in all samples extracted for enzymatic assays. Estimation was done by using Lowry’s method (Lowry et al 1951).To 0.5 ml of appropriately diluted sample in test tubes, 2.5 ml of reagent C (reagent A and reagent B in 50:1, mixed just before use, where reagent A was 2% Na2CO3 in 0.1 N NaOH and reagent B was 0.5% CuSO4 in 1% sodium potassium tartarate) was added. Contents of test tubes were mixed and allowed to stand for 10 minutes at room temperature. 0.25 ml of reagent D (Folin ciocalteu phenol reagent: Water in 1:1, mixed just before use) was added and mixed rapidly. The intensity of blue colour developed was read after 30 minutes at 530 nm. The amount of protein was calculated from standard curve of different concentrations of bovine serum albumin (20-100 µg). 3.6 Gene Expression Analysis Gene expression analysis was done by semi-quantitative RT-PCR. In the present study, LEA genes and genes involved in antioxidant response of the plants were analyzed for such 36 assays. Assays were done in shoots of both cultivars at 24h and 48h stages after applying stress on 4th day after germination. Stress treatments were 20 µM ABA for ABA treatment, 6% mannitol for WS treatment, 20 µM ABA plus 6% mannitol for combined stress treatment. The results of stress treated samples were compared with similarly old grown control seedlings grown on autoclaved distilled water. The protocol for analysis is mentioned below: 3.6.1 Database search It was done for the availability of the gene sequences for above mentioned genes in wheat plant. Nucleotide, mRNA, EST and protein databases were searched at NCBI (www.ncbi.nlm.nih.gov/) for these genes in wheat. The results of this search were summarized in Table 3.1. Primers were designed manually as well as with Primer-BLAST software available at NCBI. Primer sequences corresponding to each gene are given in Table 3.1. 3.6.2 Total RNA isolation Total RNA was isolated from shoots of all the four treatments in both cultivars at 24h and 48h stages. Total RNA was isolated by using TRIsoln (GeNei Pvt. Ltd.) where tissue was crushed in TRIsoln and then chloroform was added to separate the aqueous phase from organic phase. RNA was precipitated from aqueous phase using isopropanol. Pellet obtained was washed with 70% ethanol and then dissolved in DEPC-treated autoclaved distilled water. Isolated RNA was treated with DNase I to remove contaminating DNA. The RNA was specifically precipitated from DNase I treated RNA sample in about 3.5 M LiCl at -20 ºC. The pellet obtained was washed in 70% ethanol and dissolved in DEPC-treated autoclaved distilled water. RNA content and quality were estimated by UV spectrophotometer at 260/280 nm. 3.6.3 RT-PCR (Reverse Transcription-Polymerase chain reaction) Approximately 1 µg of total RNA was used as a template to make cDNA using reverse transcriptase. Approximately 1 µg of total RNA was mixed with 5-100 pmole of oligo-(dT)18 and denatured at 72 ºC for 5 minutes. The denatured mixture was mixed with 500 µM of each dNTP, 20U of Recombinant Human Placental Ribonuclease inhibitor (GeNei Pvt. Ltd.), 70U of M-MuLV Reverse Transcriptase (GeNei Pvt. Ltd.) in 1X transcription buffer in a total reaction volume of 20 µl. The reaction mixture was incubated at 42 ºC for 1 hour and stored at -20 ºC. PCR was performed in a total reaction volume of 25 µl with 1X buffer containing 1.5 mM MgCl2, 200 µM of each dNTP, 0.4% PVP, 25 pmole of each primer and 5 Units of Taq Polymerase (GeNei Pvt. Ltd.). PCR was performed under standard conditions (35 cycles of 40 sec at 94 ºC/ 40sec at 55 ºC/ 1-2 min at 72 ºC/ final extension of 10 min). RT-PCR of cytosolic glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was also run on the same samples to normalize the amount of template in each sample. 37 3.6.4 Agarose Gel Electrophoresis PCR products were run on 1.5% agarose gel and then visualized in Gel DocTMXR (BioRad Laboratories, Inc). Intensity of bands were measured by using Quantity One 1-D Analysis software (BioRad Laboratories, Inc). Ratio was calculated by dividing intensity of band of gene by the intensity of the band of GAPDH (constitutive gene used as internal control). 38 Table 3.1: List of genes along with their primer sequences used in the present study Antioxidant Genes Gene ID Primer sequence (5’ to 3’) Gene name EF555121 Peroxisomal APX F- TAGGTCGTCCGCGATGGCGG R- CCCCTTACTTGCTCCTCTTGG X94352 CAT-2 F- GCTTTCTTCCTTCTTCCTCGCC R- CAACTACTCTCCAGTTCTCCT D86327 CAT-1 F- GCTGACCGTTCCTCCGTTCGC R- TTGACGTAGTGCGCCTTGCC Late Embryogenesis Abundant (LEA) Genes Group Gp2 Group Gp3 Gene ID Primer sequence (5’ to 3’) Gene name CV762802 Similar to X78429 (Td 16) of durum wheat F- GCCAAGTGAGCAAGACAACA R- ATGACCTTGCTGTCCGTAGG DR739608 Similar to X78430 (Td 25a) of durum wheat F- CGAGAAGAAGAGCCTCATGG R- GTTTTCCCAGTCACGACGTT AL815683 Similar to X78431 (Td 27e) of durum wheat F- AAAGCCACAACCAAGTCCAG R- GTAGGCTCCACCAGTTCCAG U73211 WCOR410 (similar to AJ890140, Td 11 of durum wheat) F- CGAGGAGGAGAAGAAAGGCT R- CTCCCACCTTGACACCAACT AB076807 Wdhn 13 F- TGAGGGCAAGATGGAGCACC R- ATACCATGCACCGGTTGAACC Gene ID Primer sequence (5’ to 3’) Gene name AF255052 Wrab 19 F- GAGCAATACTAGCAGTGAGATTTAC R- GTACTGTAGAAGGCTCGTGAAC AB115913 Wrab 15 F- CGCGTCTCACGTCAGTCGGT 39 R- TAGGCGAGGGTATGCGTGG Gp4 AF255053 Wrab 17 F- TCAACTTCAAAAATGTCTGGTTGGT R- ATAGCGAAACAGAAGGAGGG AB115914 Wrab 18 F- CTTCGTGTTTGTTGGTGAGAGAG R- GCGAACGACCAAACGAGTAAAGG AY148490 Ta LEA-3 like F- AGGTCGTGTTCCAAGAAACC R- TCGAAGGCAAACTTTTACACCA GH729039 Similar to AJ890139 (Td 29) of durum wheat F- ATTATTACGCCGTGCACACA R- CTCGACATACCGGTGAAGGT Constitutive Gene Gene ID EF592180 Primer sequence (5’ to 3’) Gene name Cytosolic Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 40 F- GGATGCTCCCATGTTTGTC R- CCTGTTGTCACCCTGAAAGT CHAPTER IV RESULTS AND DISCUSSION 4.1 Effect on growth The seedlings of both cultivars were grown for four days in distilled water, stress was applied on 4th day by 20 µM ABA for ABA treatment, 6% mannitol for WS treatment, 20 µM ABA plus 6% mannitol for combined stress treatment. Measurement of growth was done at 24h, 48h, 72h after applying stress and was compared with similarly old control seedlings growing on distilled water as well with 0h seedlings taken on 4 th day before applying stress. Growth data was collected from root and shoot separately and different growth parameters- shoot length (Table 4.1, Figure 4.1), water content (Table 4.2, Figure 4.2), fresh weight (Table 4.3, Figure 4.3), dry weight (Table 4.4, Figure 4.4) and root (dw)/ shoot (dw) ratio (Table 4.5, Figure 4.5) were studied. Shoot length was determined with the help of ruler for 50 seedlings. Mean ± S.D was calculated and data was analyzed by DSAASTAT by applying Duncan’s Multiple Test (p> 0.5). Decrease in shoot length was found under all stress treatments in both cultivars (Table 4.1; Figure 4.1). However, no significant variation for this parameter was found between cultivars under all stress treatments. Water contents were determined using FW-DW/DW formula. Water contents were found to be affected in roots and shoots of both cultivars under water stress and combined treatments. However water contents were not affected under ABA treatment in both cultivars, except in PBW343 shoots at 24h stage where these were decreased (Table 4.2; Figure 4.2). Comparing cultivars, water contents were decreased throughout the stress period in C306 however water contents were regained after 24h of stress in PBW343. It can be concluded from this study that shoot length but not water contents were decreased by ABA which is also documented in literature that ABA inhibits shoot growth in drying soil under conditions when shoot water deficit does not occur (Sharp and Le Noble 2002). Moreover low water content in C306 can also be correlated to its dehydration tolerance as dehydration tolerance includes the ability of plant to maintain cell functions under low water contents and hence a mechanism for dehydration tolerance (Passiosure et al 1993). Lower contents of RWC (Relative water content) were observed in leaves of drought tolerant wheat cultivars than drought susceptible cultivars at different days after sowing under field conditions (Lascano et al 2001). Table 4.1: Shoot length (cm) of germinating wheat seedlings of two wheat cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 C306 Shoot length 42 CT ABA WS ABA+WS CT ABA WS ABA+WS 0h 2.79 ± 0.48a 2.79 ± 0.48a 2.79 ± 0.48a 2.79 ± 0.48a 2.40 ± 0.47a 2.40 ± 0.47a 2.40 ± 0.47a 2.40 ± 0.47a 24h 5.60 ± 1.26g 4.35 ± 1.03cdef 3.47 ± 0.89b 3.92 ± 0.97c 4.34 ± 1.21ef 4.13 ± 1.26def 2.93 ± 0.94ab 3.23 ± 0.92bc 48h 7.30 ± 1.24h 5.33 ± 1.27g 4.18 ± 0.95c 4.27 ± 1.15cd 7.18 ± 2.58h 4.77 ± 1.42f 3.58 ± 1.36cd 3.87 ± 0.98cde 72h 9.68 ± 1.45i 5.71 ± 1.24g 4.75 ± 1.11df 4.27 ± 1.35cde 9.58 ± 2.89i 5.92 ± 1.94g 4.30 ± 1.14ef 3.62 ± 1.00cd *Shoot length was measured for 50 seedlings. Mean ± SD was calculated. *Data in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 14 72h 48h 24h 0h i shoot length (cm) 12 i h 10 h 8 gg g 6 43 4 g df cdef b c a a a a ef cd cde c f cde cd a a a ef def ab bc cd a 2 0 CT Fig. 4.1: ABA PBW343 WS ABA+WS CT ABA WS C306 ABA+WS Shoot length of germinating wheat seedlings of two wheat cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Shoot length was measured for 50 seedlings. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. Table 4.2: Water contents of shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 C306 WC CT ABA WS ABA+WS CT ABA WS ABA+WS Shoot 44 0h 10.21 ± 0.01de 10.21 ± 0.01de 10.21 ± 0.01de 10.21 ± 0.01de 12.31 ± 0.25h 12.31 ± 0.25h 12.31 ± 0.25h 12.31 ± 0.25h 24h 12.62 ± 4.73e 8.02 ± 1.21bcd 7.03 ± 0.40abc 7.06 ± 0.42abc 9.67 ± 0.01fg 10.13 ± 0.50g 7.17 ± 0.37cd 6.99 ± 0.33c 48h 8.74 ± 0.34bcd 9.44 ± 1.19cd 6.66 ± 0.50abc 6.39 ± 0.69ab 8.05 ± 0.10e 9.21 ± 0.29f 5.58 ± 0.06ab 6.01 ± 0.78b 72h 7.98 ± 0.19bcd 7.62 ± 0.043bcd 5.75 ± 0.47ab 4.63 ± 2.19a 7.77 ± 0.16de 7.64 ± 0.67cde 4.94 ± 0.14a 5.10 ± 0.47a Root 0h 15.22 ± 0.91f 15.22 ± 0.91f 15.22± 0.91f 15.22 ± 0.91f 14.98 ± 3.03e 14.98 ± 3.03e 14.98 ± 3.03e 14.98 ± 3.03e 24h 13.62 ± 1.94ef 10.47 ± 1.33cde 8.67 ± 3.4abcd 6.44 ± 0.80c 13.91 ± 0.93de 11.68 ± 1.44cd 8.67 ± 1.28ab 7.54 ± 1.63a 48h 14.14 ± 0.14f 11.82 ± 1.84def 8.08 ± 2.56abc 5.72 ± 0.30b 11.32 ± 0.20c 10.24 ± 1.28bc 6.40 ± 1.51a 6.29 ± 0.88a 72h 10.23 ± 3.63cd 9.61 ± 1.12bcd 9.46 ± 0.87bcd 6.50 ± 1.59a 12.12 ± 0.42cd 10.65 ± 1.44bc 8.40 ± 0.52ab 8.26 ± 0.10ab * Water contents were recorded in triplicates. Mean ± SD was calculated. *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 0h A 20 48h 72h e 18 Water content in shoot (g H2O/g DW) 24h 16 14 12 h h de bcd bcd 10 de cd bcd abc abc ab bcd 8 fg abcab e f cde de c cd a b ab 6 a a 4 2 0 CT ABA WS ABA+WS CT ABA PBW343 B 20 18 16 Water content in root (g H2O/g DW) h g de de h 14 0h 24h 48h f f cd f 72h e e e f de def cde 12 ABA+WS C306 e f ef WS cd abcd bcd 10 c abc bcd cd bcbc ab ab ab 8 a ab a ab a a 6 4 2 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.2: Water contents of shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Water contents were recorded in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 45 Table 4.3: Fresh weights (g) of shoot (upper panel) and root (lower panel) of 25 germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 C306 FW CT ABA WS ABA+WS CT ABA WS ABA+WS Shoot 46 0h 1.07 ± 0.07abcd 1.07 ± 0.07 abcd 1.07 ± 0.07 abcd 1.07 ± 0.07 abcd 1.13 ± 0.06ab 1.13 ± 0.06ab 1.13 ± 0.06ab 1.13 ± 0.06ab 24h 1.31 ± 0.04ef 1.24 ± 0.01de 1.08 ± 0.05abcd 1.10 ± 0.11bcde 1.44 ± 0.03de 1.40 ± 0.10de 0.93 ± 0.24a 1.19 ± 0.08bc 48h 1.48 ± 0.09f 1.19 ± 0.14cde 0.91 ± 0.18ab 1.01 ± 0.09abc 1.66 ± 0.02f 1.34 ± 0.14d 1.06 ± 0.12ab 1.00 ± 0.15ab 72h 1.72 ± 0.01g 1.17 ± 0.09cde 1.00 ± 0.09abc 0.89 ± 0.23a 1.74 ± 0.002f 1.55 ± 0.016ef 1.10 ± 0.16ab 1.17 ± 0.00bc Root 0h 0.60 ± 0.10abc 0.60 ± 0.10 abc 0.60 ± 0.10 abc 0.60 ± 0.10 abc 0.73 ± 0.04abc 0.73 ± 0.04 abc 0.73 ± 0.04 abc 0.73 ± 0.04 abc 24h 0.72 ± 0.15cde 0.66 ± 0.20 abcd 0.64 ± 0.05 abcd 0.62 ± 0.02 abcd 0.83 ± 0.07cd 0.69 ± 0.18 abc 0.54 ± 0.07a 0.56 ± 0.08 ab 48h 0.94 ± 0.06e 0.62 ± 0.03 abcd 0.52 ± 0.14 abc 0.46 ± 0.05ab 0.99 ± 0.02d 0.71 ± 0.19 abc 0.73 ± 0.15 abc 0.51 ± 0.01a 72h 0.84 ± 0.15de 0.54 ± 0.20 abc 0.68 ± 0.07bcd 0.45 ± 0.10a 0.84 ± 0.15cd 0.76 ± 0.18 abcd 0.60 ± 0.09 abc 0.80 ± 0.20bcd * Fresh weights of 25 seedlings were recorded in triplicates. Mean ± SD was calculated. *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. shoot fresh weight (g) per 25 seedlings A 2 0h 48h 72h g 1.8 f f 1.6 1.4 24h ef decd de cde de cde abcd bcde abcd abcd ababc abcd abca 1.2 abcd f ab ef ab ab ab aab ab bc bc ab WS ABA+WS 1 0.8 0.6 0.4 0.2 0 CT ABA WS ABA+WS CT ABA PBW343 B 1.2 0h 24h C306 48h 72h root fresh weight (g) per 25 seedlings e de d cd 1 cde 0.8 abc cd abcd abc abcd bcd abc abc abcd abc abcd abc abc 0.6 ab bcd abcd abc abc abc abc abc abc abc ab a a a 0.4 0.2 0 CT ABA WS ABA+WS PBW343 Fig. 4.3: CT ABA WS ABA+WS C306 Fresh weights of shoot (A) and of root (B) of 25 germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Fresh weights of 25 seedlings were recorded in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 47 Table 4.4: Dry weights (g) of shoot (upper panel) and root (lower panel) of 25 germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. DW PBW343 CT ABA C306 WS ABA+WS CT ABA WS ABA+WS Shoot a 0.10 ± 0.006 a 0.10 ± 0.006 a 0.10 ± 0.006a 0.09 ± 0.006a 0.09 ± 0.006a 0.09 ± 0.006a 0.09 ± 0.006a 48 0h 0.10 ± 0.006 24h 0.10 ± 0.03a 0.14 ± 0.02bcd 0.13 ± 0.01bcd 0.14 ± 0.02bcd 0.14 ± 0.002bc 0.13 ± 0.015b 0.11 ± 0.04ab 0.15 ± 0.003bcd 48h 0.15 ± 0.01cd 0.12 ± 0.03ab 0.12 ± 0.016abc 0.14 ± 0.001bcd 0.18 ± 0.004ef 0.13 ± 0.01bc 0.16 ± 0.02cde 0.15 ± 0.04bc 72h 0.19 ± 0.005e 0.14 ± 0.009bcd 0.15 ± 0.02bcd 0.16 ± 0.02cd 0.20 ± 0.003f 0.18 ± 0.016def 0.19 ± 0.02ef 0.19 ± 0.014ef Root 0h 0.04 ± 0.004a 0.04 ± 0.004a 0.04 ± 0.004a 0.04 ± 0.004a 0.05 ± 0.006a 0.05 ± 0.006a 0.05 ±0.006a 0.05 ± 0.006a 24h 0.05 ± 0.004ab 0.06 ± 0.010abc 0.07 ± 0.018bcd 0.08 ± 0.012d 0.06 ± 0.009ab 0.05 ± 0.008ab 0.06 ± 0.00ab 0.07 ± 0.004abc 48h 0.06 ± 0.004bcd 0.05 ± 0.004ab 0.06 ± 0.000abc 0.07 ± 0.004bcd 0.08 ± 0.003bcd 0.06 ± 0.010abc 0.10 ± 0.04d 0.07 ± 0.010abc 72h 0.08 ± 0.011cd 0.05 ± 0.013ab 0.06 ± 0.012bcd 0.06 ± 0.026bcd 0.06 ± 0.009abc 0.06 ± 0.007abc 0.06±0.006abc 0.09 ± 0.022cd * Dry weights of 25 seedlings were recorded in triplicates. Mean ± SD was calculated. *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. A 0.25 0h 48h 72h a a ef def ef cd bcd cd 0.15 ef f e 0.2 shoot dry weight (g) per 25 seedlings 24h bc cde bcd bcd bcd ab bcd abc a a ab b bc bc bcd bcd a a 0.1 a a a f 0.05 0 CT ABA WS ABA+WS CT ABA PBW343 B 0.16 0h 24h WS ABA+WS C306 48h 72h d root dry weight (g) per 25 seedlings 0.14 0.12 cd 0.1 d bcd cd bcd 0.08 bcd abc 0.06 a a bcd abc ab bcd abc ab ab ab bcd a a a abcabc ab abc aab abc abc a a 0.04 0.02 0 CT ABA WS ABA+WS PBW343 Fig. 4.4: CT ABA WS ABA+WS C306 Dry weight of shoot (A) and of root (B) of 25 germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Dry weights of 25 seedlings were recorded in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 49 Table 4.5: Root (dw)/shoot (dw) ratio of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 C306 50 CT ABA WS ABA+WS CT ABA WS ABA+WS 0h 0.39 ± 0.07a 0.39 ± 0.07a 0.39 ± 0.07a 0.39 ± 0.07a 0.55 ± 0.12bc 0.55 ± 0.12bc 0.55 ± 0.12bc 0.55 ± 0.12bc 24h 0.50 ± 0.12ab 0.42 ± 0.13a 0.51 ± 0.084ab 0.61 ± 0.009b 0.41 ± 0.06ab 0.42 ± 0.02ab 0.51 ± 0.15abc 0.45 ± 0.03abc 48h 0.41 ± 0.063a 0.43 ± 0.14a 0.49 ± 0.067ab 0.50 ± 0.029ab 0.44 ± 0.003abc 0.48 ± 0.04abc 0.65 ± 0.32c 0.49 ± 0.06abc 72h 0.40 ± 0.07a 0.37 ± 0.07a 0.44 ± 0.009a 0.38 ± 0.11a 0.32 ± 0.05a 0.36 ± 0.07ab 0.34 ± 0.01a 0.45 ± 0.08abc *Ratios were recorded in triplicates. Mean ± SD was calculated. *Data in each cultivar was analyzed by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. Root dry weight/shoot dry weight ratio 1.2 0h 24h 48h 72h 1 c 0.8 bc ab 0.6 a a a a ab a a bc abc bc abc ab ab a bc b a a a a abc ab abc 0.4 ab abc abc ab a a 51 0.2 0 CT ABA PBW343 WS ABA+WS CT ABA WS ABA+WS C306 Fig. 4.5: Root (dw)/shoot (dw) ratios of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Ratios were recorded in triplicates. Mean ± SD was calculated. Data was analyzed separately in each cultivar by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. Fresh weights (Table 4.3, Figure 4.3) and Dry weights (Table 4.4, Figure 4.4) of roots and shoots of 25 seedlings were measured in triplicates. Mean ± S.D. was calculated and data was similarly analyzed by DMT. Both fresh weight and dry weight were affected under all stress treatments in both roots and shoots of PBW343 only but not in C306. In C306 shoots, dry weights were maintained at all three stages under all stress treatments, except there was decrease in dry weight at 48h stage under ABA and combined treatment. In C306 roots, dry weights were not affected under any treatment. In PBW343, dry weights of shoots were affected more than roots. There was increase in the dry weights of shoots of PBW343 at 24h under all stress treatments but this increase was followed by decrease during later stages. In PBW343 roots, dry weights were decreased under ABA treatment only otherwise these were maintained under other stress treatments. Comparing ABA response and WS response in both cultivars, dry weights were affected more by ABA than by WS. Moreover these effects of ABA were more pronounced in shoots than in roots, which could be related to known function of ABA i.e. to maintain more root growth under stresses hence to maintain root/shoot growth ratio (Vysotskaya et al 2008). From above data, conclusion can be made that ABA affects dry weight accumulation in plants but does not affect water content. ABA treatment has also been reported to increase water content under drought stress in triploid Bermuda grass (Shaoyun et al 2009). Secondly, under WS, dry weights were maintained better in roots than in shoots in both cultivars. Extensive root growth under drought conditions is considered to be a major drought avoidance mechanism (Shao et al 2008). This process is partly mediated through ABA (Cutler et al 2010). Increased biomass and decreased water content under drought are considered to be the drought tolerant features (Guoxiong et al 2002, Passiosure et al 1993, Lascano et al 2001) and these features were found to be better shown in C306, drought tolerant cultivar than in PBW343, drought sensitive cultivar in this study. 4.2 Effects on antioxidant response In present study antioxidant response was determined in terms of H2O2, ascorbate, ascorbate/dehydroascorbate ratio, malondialdehyde, proline contents and levels of antioxidant enzymes, Ascorbate peroxidase (APX), Guaicol peroxidase (GPOX), Catalase (CAT), Glutathione reductase (GR), Superoxide dismutase (SOD). 4.2.1 Hydrogen peroxides (H2O2) contents H2O2 contents were found to be higher in roots and shoots of control seedlings of PBW343 than of C306 (Table 4.6; Figure 4.6). H2O2 contents increased at 24h stage under all stress treatments in roots of C306 but decreased in the roots of PBW343. These contents were almost unaffected at 24h stage under all stress treatments in shoots of C306 but in 52 shoots of PBW343, it decreased under WS, unaffected under combined stress, increased under ABA treatment at the same stage. H2O2 contents were brought down during later stages in roots of C306 but in shoots of this cultivar, these contents were not affected by any stress treatment and showed increasing trend during germination like as in control seedlings. Comparing both cultivars for this feature, it was clear that there was no increase in H2O2 contents in both root and shoot under WS in drought susceptible cultivar (PBW343). However there was increase in H2O2 level by exogenous ABA in shoots of this cultivar when ABA was applied either individually or under combined stress. Contrary to it, drought tolerant cultivar C306 increased H2O2 level in its roots and maintained higher such contents in its shoots during germination under all stress treatments. As exogenous supply of ABA gave the feature of elevating H2O2 level in PBW343 which indicates that H2O2 production might be the downstream effect of ABA in ABA -dependent pathway which was working in C306 but might be lacking in PBW343 under WS. Rise in H2O2 content in roots of C306 under stresses is understandable as roots of plants are the first tissue to sense water deficit conditions (Selote and Khanna-Chopra 2010). H2O2 content is reported to be increased under ABA and is a downstream effect of ABA. But H2O2 contents have also been found to be decreased under stresses (Loggini et al 1999, Simova-Stoilova et al 2008) similar to as observed in PBW343 in our study. So far H2O2 contents in literature are considered to be related to drought resistance mechanism where lower levels of these contents under stresses were positively correlated to drought tolerance levels as many reports reported lower levels of H2O2 in drought resistant cultivars than drought susceptible cultivars when exposed to similar stresses (Guo et al 2006, KhannaChopra and Selote 2007, Wang et al 2009). These studies considered H2O2 as toxic molecule in plants which causes oxidative damage. Hence its lower level under stresses favors growth and avoids oxidative damage. But recently, another view is given, which considers H2O2 not as toxic molecule rather a signaling molecule to initiate ROS signaling for producing stress response. This is supported by observations that H2O2 content has not been found to be related to lipid peroxidation (Jubany-Mari et al 2010); plants with lower cytosolic APX activity or cAPX knockout were found to be more drought tolerant rather drought susceptible (Ishikawa and Shigeoka 2008, Munns and Tester 2008, Jubany-Mari et al 2010). So higher level of H2O2 contents in drought tolerant cultivar and lower level in drought susceptible cultivar under WS might be related to such kind of observations. 53 Table 4.6: Amount of H2O2 (µmole of H2O2 g-1 DW) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. H2O2 PBW343 CT ABA C306 WS ABA+WS CT ABA WS ABA+WS Shoot 54 0h 1.53 ± 0.18abc 1.53 ± 0.180abc 1.53 ± 0.18abc 1.53 ± 0.18abc 1.12 ± 0.41a 1.12 ± 0.41a 1.12 ± 0.41a 1.12 ± 0.41a 24h 3.08 ± 0.58d 3.88 ± 0.49e 1.80 ± 0.09bc 2.99 ± 0.76d 1.29 ± 0.20ab 1.75 ± 0.02abcd 1.93 ± 0.23abcde 1.58 ± 0.12abc 48h 5.75 ± 0.18f 3.48 ± 0.04de 2.00 ± 0.15c 2.85 ± 0.32d 2.40 ± 1.18cde 2.09 ± 0.38bcde 2.57 ± 0.25def 2.50 ± 0.30def 72h 3.18 ± 0.32d 1.03 ± 0.08a 1.20 ± 0.40ab 1.79 ± 0.38bc 3.67 ± 0.79g 2.74 ± 0.39ef 3.30 ± 0.10f 1.79 ± 0.23abcd cd cd cd 1.11 ± 0.24cd 1.20 ± 0.22ab 1.20 ± 0.22ab 1.20 ± 0.22ab 1.20 ± 0.22ab Root 0h 1.11 ± 0.24 1.11 ± 0.24 1.11 ± 0.24 24h 2.37 ± 0.54e 1.28 ± 0.04cd 0.40 ± 0.05ab 0.81 ± 0.23bc 0.75 ± 0.18a 2.84 ± 0.88def 2.55 ± 0.03de 3.29 ± 0.06ef 48h 6.28 ± 0.70g 3.06 ± 0.35f 2.69 ± 0.07ef 1.41 ± 0.07d 3.57 ± 1.01f 1.65 ± 0.18bc 1.44 ± 0.37ab 1.45 ± 0.34ab 72h 0.19 ± 0.02a 0.23 ± 0.06a 0.16 ± 0.03a 0.11 ± 0.01a 4.89 ± 0.31g *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. 2.36 ± 0.39cd 2.49 ± 0.13d 1.59 ± 0.11b *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. umole of H2O2 per gm DW A 7 0h 24h 48h 72h f 6 5 g e d 4 d de d cde d ef 3 2 abc bc c bc abc ab abc abc bcde abcd a a ab def f g def abcde abcd aabc a a 1 0 CT ABA WS ABA+WS CT ABA 0h 24h ABA+WS C306 PBW343 B 8 WS 48h 72h umole of H2O2 per gm DW g 7 6 g f 5 4 e 3 2 def f ef cd cdcd 1 a cd cd a cdbc ab ef a de bc d ab ab d ab ab ab b ab a a 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.6: H2O2 contents (µmole of H2O2 g-1 DW) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 55 4.2.2 Ascorbic acid contents and ascorbate/dehydroascorbate ratio Ascorbate contents (Table 4.7; Figure 4.7) were found to be more in shoots than in roots of both wheat cultivars. Ascorbate contents increased in shoots and roots of PBW343 at 48h stage following increased contents of H2O2 at 24h stage of shoot under ABA treatment, accompanied by similar increase in ascorbate/dehydroascorbate ratio at the same stage (Table 4.9; Figure 4.8). However under WS as well as under combined stress, such increases in contents and ratios were not observed rather both decreased in roots as well as in shoots of PBW343. But combined stress could be placed intermediate between ABA treatment and WS treatment for contents as well as for ratios. Shoots of C306 control plants had already high ascorbate/dehydroascorbate ratio at the stage corresponding to 24h stage. Ascorbate contents of shoots of C306 increased under ABA treatment from 24-48h accompanied by higher ascorbate/dehydroascorbate ratio during same time period. Ascorbate contents were not altered under WS as well as under combined stress in shoots of C306. But ascorbate/dehydroascorbate ratios first decreased at 24h stage but were maintained higher during later stages compared to control seedlings of same stage under WS and combined stresses. In C306 roots, ascorbate contents were not much affected under all stress treatments but ratios were maintained higher compared to those of control seedlings at 24h to 48h stages under all stresses. Above results indicate that ABA increased H2O2 contents followed by increases in ascorbic acid contents as well as in ascorbate/dehydroascorbate ratios in both cultivars. Such type of ABA-dependent response might be lacked in PBW343 but working in C306 under WS. In literature, there are many reports which correlate drought tolerance of plant with higher ascorbate and ascorbate/dehydroascorbate ratio under stresses (Sairam and Shrivastva 2001, Jiang and Zhang 2002, Guo et al 2006, Khanna-Chopra and Selote 2007) but others reports are also there where ascorbate contents were found to be decreased under stresses (Selote and Khanna-Chopra 2006, Simova-Stilova et al 2008). 4.2.3 Malondialdehyde (MDA) contents MDA contents (Table 4.10; Figure 4.9) were not much altered in roots and shoots of C306 except its shoots showed higher contents at 24h stage under WS. In PBW343 shoots, MDA contents were found to be higher at 24h stage under both WS and combined stress. Roots of PBW343 showed higher level of MDA at 72h stage under all stress treatments and at 48h stage under WS and combined treatments. 56 Table 4.7: Amount of ascorbic acid (µg of ascorbate g-1 DW) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Ascorbic acid PBW343 C306 ABA WS 889.44 ± 153.14ab 490.21 ± 42.87a 2614.76 ± 753.00d 2620.90 ± 144.93d 889.44 ± 153.14ab 540.04 ± 21.98a 3405.77 ± 565.67e 1484.83 ± 89.31c 889.44 ± 153.14ab 639.45 ± 156.61ab 1180.99 ± 154.12bc 759.53 ± 231.41ab ABA+WS Shoot CT ABA WS ABA+WS 889.44 ± 825.33 ± 825.33 ± 825.33 ± 825.33 ± 153.14ab 103.87ab 103.87ab 103.87ab 103.87ab 868.44 ± 1257.20 ± 1470.11 ± 1370.74 1274.48 ± 24h 83.01ab 636.39bc 204.34cd ±476.82bcd 78.94bc 1648.97 ± 2414.57 ± 3979.59 ± 1893.88 ± 2388.96 ± 48h 430.96c 307.24e 608.88f 165.13de 106.74e 471.77 ± 866.84 ± 549.57 ± 946.92 ± 948.17 ± 72h 105.68a 145.18ab 47.26a 274.62abc 67.50abc Root 1302.79 ± 1302.79 ± 1302.79 ± 1302.79 ± 1094.65 ± 1094.65 ± 1094.65 ± 1094.65 ± 0h 134.59d 134.59d 134.59d 134.59d 344.56c 344.56c 344.56c 344.56c 432.85 ± 412.98 ± 187.912 ± 227.64 ± 370.68 ± 410.46 ± 322.63 ± 218.99 ± 24h ab ab a a ab ab ab 6.84 20.38 23.53 90.09 10.00 12.44 27.41 41.93a 1226.21 ± 1924.05 ± 512.17 ± 716.11 ± 865.43 ± 901.11 ± 327.61 ± 508.19 ± 48h d e ab bc c c ab 148.94 665.56 40.95 62.08 114.25 54.88 51.96 26.37b 1230.43 ± 1052.53 ± 1127.81 ± 459.56 ± 858.83 ± 553.56 ± 507.76 ± 332.31 ± 72h 113.72d 166.84cd 132.39d 100.07ab 279.61c 80.80b 134.01b 109.35ab *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 0h 57 CT A 5000 0h 24h 48h ug of ascorbate per gm DW 4500 72h f e 4000 d 3500 d 3000 e 2500 e bc c bcdde cd 2000 1500 c ab 1000 bc ab a ab ab a ab ab ab ab ab bc abc ab ab ab abc a a 500 0 Ct ABA WS ABA+WS Ct ABA 3000 0h 24h 48h ABA+WS C306 PBW343 B WS 72h e ug of ascorbate per gm DW 2500 2000 1500 d d d d d c d c d cd c c c c 1000 bc ab 500 ab ab c b ab ab a b b ab ab abab a a 0 Ct ABA WS ABA+WS PBW343 Ct ABA WS ABA+WS C306 Fig. 4.7: Ascorbic acid (µg of ascorbate g-1 DW) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 58 Table 4.8: Amount of dehydroascorbate (µg of dehydroascorbate g-1 DW) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 C306 Dha CT ABA WS ABA+WS CT ABA WS ABA+WS Shoot 59 0h 1587.64±214.83 1587.64±214.83 1587.64±214.83 1587.64±214.83 1884.08±254.93 24h 1788.36±649.39 1638.07±556.3 1217.64±240.42 1538.33±575.36 240.13±75.85 48h 1261.65±340.29 760.72±188.65 1796.00±318.54 1329.46±256.32 72h 2376.11±228.43 1201.52±354.58 2145.27±655.23 1085.10±350.36 1884.08±254.93 1884.08±254.93 1884.08±254.93 279.82±74.24 447.46±73.31 484.37±166.64 4127.69±1132.39 2203.67±182.76 1699.21±177.24 1562.92±260.69 563.07±64.82 429.32±27.18 735.21±243.73 466.28±26.34 Root 0h 1325±65.57 1325±65.57 1325±65.57 1325±65.57 1486.30±63.29 1486.30±63.29 1486.30±63.29 1486.30±63.29 24h 991.6±238.78 701.55±99.41 606.80±179.02 511.94±141.83 1249.26±106.69 577.59±39.47 1182.84±69.22 982.02±323.53 48h 790.85±134.21 605.63±206.13 1538.73±547.05 319.53±20.78 1287.41±214.44 1321.89±226.44 386.38±83.18 583.04±104.49 72h 1289.39±300.64 718.11±188.76 1799.58±5.00 965.84±5.00 873.18±220.64 551.72±5.00 879.86±170.74 339.95±94.26 *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. A 6 ascorbate/dehydroascorbate 0h 24h 48h 72h 5 4 3 2 1 0 CT ABA WS ABA+WS CT ABA WS ABA+WS C306 PBW343 B 3.5 0h 24h 48h 72h ascorbate/dehydroascorbate 3 2.5 2 1.5 1 0.5 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.8: Ascorbate/dehydroascorbate ratios in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. 60 Table 4.9: Ascorbate/Dehydroascorbate ratios in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Ascorbate/ Dehydroascorbate PBW343 CT ABA C306 WS ABA+WS CT ABA WS ABA+WS Shoot 61 0h 0.560 0.560 0.560 0.560 0.438 0.438 0.438 0.438 24h 0.274 0.330 0.525 0.565 5.236 5.254 3.063 2.631 48h 2.072 4.477 0.658 1.240 0.585 1.806 1.115 1.529 72h 1.103 1.236 0.354 0.435 1.539 1.280 1.288 2.033 Root 0h 0.983 0.983 0.983 0.983 0.736 0.736 0.736 0.736 24h 0.437 0.589 0.310 0.445 0.297 0.711 0.273 0.223 48h 1.550 3.177 0.333 2.241 0.672 0.682 0.848 0.872 72h 0.954 1.466 0.627 0.476 0.984 1.003 0.577 0.978 Table 4.10: Amount of malondialdehyde (nmole of MDA g-1 DW) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. MDA PBW343 CT ABA C306 WS ABA+WS CT ABA WS ABA+WS Shoot 62 0h 25.51±5.44ab 25.51±5.44ab 25.51±5.44ab 25.51±5.44ab 18.97±1.16bcde 18.97±1.16bcde 18.97±1.16bcde 18.97±1.16bcde 24h 25.10±6.92ab 31.24±0.51bc 38.12±10.77cd 46.15±11.3d 3.45±0.2a 12.46±5.41abc 22.30±9.51bcde 10.72±2.21ab 48h 23.98±1.46ab 15.43±1.71a 17.84±2.33a 16.71±2.53a 11.31±2.42ab 16.63±5.03bcd 13.25±4.01abc 11.06±0.87ab 72h 24.88±7.03ab 22.07±6.97ab 27.72±9.56abc 23.18±3.02ab 28.53±12.5e 29.17±10.3e 25.91±1.58de 23.78±8.47cde Root 0h 23.06±1.97de 23.06±1.97de 23.06±1.97de 23.06±1.97de 16.69±6.03bc 16.69±6.03bc 16.69±6.03bc 16.69±6.03bc 24h 41.53±2.81f 27.61±1.65e 14.53±3.83bc 19.56±0.92cd 36.76±11.89def 41.90±0.81efg 46.69±8.26fg 6.67±1.01ab 48h 0.00±0.00a 0.00±0.00a 14.34±2.35bc 8.39±1.33b 25.60±8.72cd 17.60±0.41bc 6.54±0.50ab 3.41±0.50a 72h 0.00±0.00a 8.83±5.95b 17.84±9.90cd 8.11±5.73b 41.90±5.06efg 40.93±3.16efg 32.29±0.32de 50.70±13.96g *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. nmole of MDA per gm DW A 70 0h 24h 48h 72h d 60 cd 50 e e abc 40 ab abab 30 bc ab ab ab ab ab a a 20 cde bcde ab de bcde bcd abc bcde a bcde abc bcde ab 10 abab a 0 CT ABA WS ABA+WS CT ABA WS C306 PBW343 nmole of MDA per gm DW B 70 0h 24h 48h ABA+WS 72h g 60 fg def 50 f efg efg efg cd 40 de 30 e de cd de de 20 b de cd bc bcbc bc bc bc bc b b ab 10 aa a ab a 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.9: Malondialdehyde (nmole of MDA g-1 DW) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 63 PBW343 showed elevated levels of MDA contents as compared to C306. Secondly, ABA treatment produced less MDA contents as compared to WS and combined stress. ABA was reported to alleviate drought- induced oxidative damage (Jiang and Zhang 2001, Shaoyun et al 2009). There was no correlation between MDA content and H2O2 level (Table 4.6, 4.10; Figure 4.6, 4.9). In literature, MDA contents were reported to be increased under stresses and lower levels of MDA was considered to be an important indice for evaluating the redox status and drought resistance (Guo et al 2006). In most of these studies, MDA contents were found to be positively correlated to H2O2 levels. But many recent studies reported opposite to it where MDA contents decreased with increase in H2O2 contents (Jubany-Mari et al 2010). Moreover it has been indicated that molecule responsible for lipid peroxidation is singlet oxygen not H2O2 (Triantaphylides et al 2008). 4.2.4 Proline contents Proline contents in roots and shoots of C306 were higher than of PBW343 for control seedlings. In control seedlings of C306, proline contents were higher in roots than in shoots (Table 4.11; Figure 4.10). Proline contents were found to be unaltered under ABA in both cultivars, except it was increased in C306 roots at 72h stage and in PBW343 shoots at 48h stage. In literature, proline accumulation under WS was reported to be independent to ABA, hence was a part of ABA independent stress response (de-Carvallo 2008, Sharma and Verslues 2010, Verslues and Zhu 2004). Proline contents were found to be increased under WS and combined stress in both cultivars and this increase was more prominent in shoots than in roots of both cultivars. In conclusions, one can say that Drought tolerant cultivar, C306 contained higher amount proline mainly in roots but under WS, proline contents were more increased in shoots than in roots and this increase in proline might be mediated through ABA independent pathway. MDA contents and proline contents were negatively correlated in shoots of PBW343 under WS and under combined stress at 24h and 48h stages (Table 4.10, 4.11; Figure 4.9, 4.10). This could be correlated to other studies where proline was considered as free radical scavenger (Kavi Kishore 2005) and proline accumulation under severe drought had been found to be linked to the MDA contents and membrane protection among different wheat varieties (Simova-Stilova et al 2008). 4.3 Antioxidant enzymes Plants keep ROS under control by efficient scavenging systems which include enzymatic antioxidants and non-enzymatic antioxidants. Enzymatic antioxidants are superoxide dismutase (SOD), catalase (CAT) and enzymes involved in ascorbate glutathione cycle. These enzymes are located in different compartments of cell except catalase which is located exclusively in peroxisomes. SOD is the front line enzyme in the 64 Table 4.11: Amount of proline (µmole of proline g-1 DW) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 C306 Proline CT ABA WS ABA+WS CT ABA WS ABA+WS Shoot 65 0h 13.52 ± 0.79f 13.52 ± 0.79f 13.52 ± 0.79f 13.52 ± 0.79f 9.93 ± 3.05ab 9.93 ± 3.05ab 9.93 ± 3.05ab 9.93 ± 3.05ab 24h 7.46 ± 0.74abc 6.93 ± 0.81ab 7.73 ± 0.93abc 7.90 ± 1.56abc 7.65 ± 3.09ab 10.86 ± 0.73bc 10.56 ± 0.49abc 15.58 ± 0.67def 48h 8.48 ± 1.40bc 11.68 ± 1.24ef 18.52 ± 1.11g 18.48 ± 1.09g 15.29 ± 1.93de 13.7 ± 1.29cd 17.72 ± 1.16ef 19.38 ± 3.83f 72h 7.47 ± 2.11abc 5.75 ± 0.89a 11.24 ± 1.10de 9.47 ± 1.60cd 8.03 ± 1.04ab 9.93 ± 1.63ab 7.08 ± 0.71a 9.80 ± 1.95ab Root 0h 13.0 ± 1.31de 13.0 ± 1.31de 13.0 ± 1.31de 13.0 ± 1.31de 9.09 ± 0.30a 9.09 ± 0.30a 9.09 ± 0.30a 9.09 ± 0.30a 24h 6.96 ± 0.98ab 5.34 ± 1.99a 8.59 ± 1.97b 5.55 ± 0.62a 24.18 ± 0.95e 16.81 ± 1.56bc 16.20 ± 2.26bc 15.00 ± 0.63b 48h 10.85 ± 0.53cd 5.97 ± 0.56a 13.48 ± 0.24e 10.91 ± 0.99cd 21.23 ± 3.13de 20.50 ± 1.64de 16.70 ± 2.41bc 15.43 ± 0.10b 72h 6.85 ± 1.80ab 8.31 ± 1.62b 12.82 ± 0.07de 8.95 ± 1.37bc 8.75 ± 3.37a 15.09 ± 2.86b 19.34 ± 3.06cd 13.26 ± 0.53b *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. *Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. umole of proline per gm DW A 25 0h 24h 48h f 72h g 20 g ef de 15 10 f f f ef f ab de cd bc abc abc ab bc ab abc abc ab def cd ab ab ab ab abc ab a a 5 0 CT ABA WS ABA+WS CT ABA umole of proline per gm DW 0h 24h 48h 72h e de 25 cd de 20 15 bc de de ab b ab a bcbc b bb de cd 10 ABA+WS C306 PBW343 B 30 WS e de de b cd bc b a a a a a a a 5 0 CT ABA WS ABA+WS PBW343 Fig. 4.9: CT ABA WS ABA+WS C306 Proline (µmole of proline g -1 DW) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 66 ROS attack. Since it rapidly scavenges superoxide which is the first ROS to be produced but it produced another ROS (H2O2). H2O2 scavengers are CAT and APX but these enzymes seem to have different roles because they are located in different compartments of the cell i.e. APX is present in almost every compartment of the cell and CAT exclusively present in peroxisomes and secondly, their affinity for H2O2 which is very high (Km in µM range) in APX but lower (Km in mM range) in catalase. It indicates APX works fine regulator of ROS under steady state level but catalase might be a bulk removal of H2O2 under stress conditions. In the present study, activities of ascorbate peroxidase (APX), catalase (CAT), guaiacol peroxidase (GPOX), superoxide dismutase (SOD), glutathione reductase (GR) were determined in roots and shoots of similarly treated seedlings (as discussed previously) at same four stages (0h, 24h, 48h and 72h). Besides determining activities for these enzymes, database was also searched for availability of gene sequences corresponding to these enzymes at NCBI (www.ncbi.nlm.nih.gov/). Nucleotide, mRNA, ESTs, protein databases were searched for their genes sequences. Primers were designed from these sequences so as to check the status of expression level of these genes by RT-PCR. Expression analysis by RT-PCR was done in shoots of both cultivars at 24h and 48h stages. Gene IDs (in the form of accession no.), gene names and primer sequences are presented in (Table 3.1) 4.3.1 Ascorbate peroxidase (APX) The APX family consists of at least five different isoforms including thylakoid (tAPX) and glyoxisome membrane forms (gmAPX), as well as chloroplast stromal soluble form (sAPX), cytosolic form (cAPX). APX activities were decreased or unaltered in roots of both cultivars under all stress treatments (Table 4.12; Figure 4.11, 4.12). No changes in APX activities were found in shoots of PBW343 under all stress treatments but increases in specific activities were found at 24h stage under WS and at 48h stage under combined stress. In C306 shoots, increases in APX activities as well as specific activities were mainly observed under ABA treatment, however, unaltered under WS as well as under combined stress except 72h stage under combined stress where it decreased. APX activities in both cultivars were concluded to be increased mainly under ABA treatment and in C306 shoots only. For expression studies, only one gene corresponding to peroxisomal ascorbate peroxidase (EF556121, PER-APX) was used. This gene expression was higher in control shoots of PBW343 but expressed very low in control shoots of C306 (Figure 4.13). Gene was induced under all stress treatments in PBW343 mainly at 48h. However in C306, gene was induced at 24h under ABA treatment as well as under WS treatment but continued to be 67 Table 4.12: Ascorbate peroxidase activity (µmole of ascorbate disappeared min-1 g-1 DW) and specific activity (µmole of ascorbate disappeared min-1 mg-1 protein) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 C306 ABA+WS CT ABA WS ABA+WS Shoot 0h 223.22±80.54def 223.22±80.54def 223.22±80.54def 223.22±80.54def 100.53±12.69ab 100.53±12.69ab 100.53±12.69ab 100.53±12.69ab (0.81±0.25)abc (0.81±0.25)abc (0.81±0.25)abc (0.81±0.25)abc (0.46±0.04)a (0.46±0.04)a (0.46±0.04)a (0.46±0.04)a ef cde ef f cde f def 24h 230.58±20.28 179.27±15.99 233.73±16.26 248.43±12.13 164.46±27.50 231.37±39.32 184.34±28.52 129.12±16.48abc abc abc d cd ab ab ab (0.73±0.110) (0.77±0.04) (1.44±0.46) (1.12±0.17) (0.72±0.06) (0.82±0.17) (0.81±0.16) (0.70±0.13)ab bcd cde ab abc abc abc a 48h 171.94±20.29 180.41±30.52 121.15±14.05 151.51±29.51 119.46±12.04 128.83±45.61 88.03±3.72 125.97±12.47abc ab ab bc csd ab ab ab (0.63±0.15) (0.65±0.09) (0.99±0.23) (1.12±0.19) (0.89±0.12) (0.72±0.38) (0.82±0.15) (0.83±0.15)ab abc a a a f ef bcd 72h 132.56±28.88 96.64±14.66 115.37±20.40 98.75±18.02 209.33±28.32 197.63±16.75 142.14±22.80 103.61±4.93ab abc a abc abc c d bc (0.74±0.21) (0.50±0.05) (0.77±0.27) (0.76±0.06) (1.36±0.48) (2.28±0.57) (1.16±0.24) (0.67±0.06)ab Root 0h 263.85±14.56g 263.85±14.56g 263.85±14.56g 263.85±14.56g 217.34±40.31gh 217.34±40.31gh 217.34±40.31gh 217.34±40.31gh bc bc bc bc abcd abcd abcd (1.29±0.22) (1.29±0.22) (1.29±0.22) (1.29±0.22) (0.94±0.14) (0.94±0.14) (0.94±0.14) (0.94±0.14)abcd g f def cd fg ef def 24h 287.60±48.25 186.50±87.47 144.37±6.28 116.85±7.10 175.74±22.47 161.14±25.57 152.83±54.94 126.30±6.29cdef d cd cd cd efg def fg (2.09±0.50) (1.70±0.68) (1.58±0.07) (1.67±0.23) (1.82±0.03) (1.61±0.36) (2.02±0.90) (1.77±0.06)efg ef cde bcd ab cde abc a 48h 176.97±38.64 121.92±23.92 108.61±4.64 54.97±11.03 118.99±13.93 80.39±13.71 51.71±14.85 58.62±3.84ab bc b bcd bc abcd a ab (1.19±0.30) (0.93±0.18) (1.50±0.04) (1.09±0.33) (1.07±0.06) (0.54±0.08) (0.71±0.41) (0.79±0.14)abc bcd a bcd abc h cdef bcd 72h 92.77±23.15 26.737±8.96 92.66±1.69 63.39±6.38 249.22±53.62 127.10±19.01 105.71±14.66 100.77±24.70abcd bc a bc bc g def cdef (1.16±0.39) (0.33±0.08) (1.19±0.13) (1.15±0.40) (2.45±0.59) (1.55±0.17) (1.42±0.19) (1.30±0.39)bcde (1.16±0.39)bc (0.33±0.08)a (1.19±0.13)bc (1.15±0.40)bc (2.45±0.59)g (1.55±0.17)def (1.42±0.19)cdef (1.30±0.39)bcde *Values within parenthesis indicate specific activity and without parenthesis indicate activity. *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the various samples. CT ABA WS 68 A 350 umole of ascorbate disappeared /min/g DW 300 0h 24h def 48h def 72h def ef 250 f f ef f ef cde cde bcd 200 def cde abc abc abc ab a 150 a a bcd abc ab ab ab a 100 abcabc ab ab 50 0 Ct ABA WS ABA+WS Ct ABA B400 0h 24h 48h WS ABA+WS C306 PBW343 72h g 350 umole of ascorbate disappeared /min/g DW def h 300 250 g g g f g gh ef gh fg 200 150 cde bcd def bcd bcd 100 cd cde gh gh def ef cdef bcd cdef abcd abc ababc a ab a 50 0 Ct ABA WS ABA+WS PBW343 Ct ABA WS ABA+WS C306 Fig. 4.11: Ascorbate peroxidase activity (µmole of ascorbate disappeared min-1 g-1 DW) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the various samples. 69 A 0h umole of ascorbate changed per min per mg protein 3 24h 48h 72h d 2.5 d 2 1.5 abc abc abcab 1 c cdcd bc abc abc abc abc abc abc ab a bc ab ab ab ab ab a 0.5 ab abab a ab a a 0 Ct ABA WS ABA+WS Ct ABA umole of ascorbate disappeared per min per mg protein 3.5 0h 24h 48h ABA+WS C306 PBW343 B WS 72h g fg 3 d cd 2.5 cd 2 1.5 bc bc bc cd bc bcd bc bcbc bc bc b def efg abcd abcd efg def abcd ab abcd bcde cdef 1 abcd abc a a 0.5 0 Ct ABA WS ABA+WS PBW343 Ct ABA WS ABA+WS C306 Fig. 4.12: Ascorbate peroxidase specific activity (µmole of ascorbate disappeared min-1 mg-1 protein) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the various samples. 70 PBW343 24hr ABA+WS ABA WS C306 48hr CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT EF556121 (PERAPX) GAPDH 2 24h 48h PBW343 C306 Ratio 1.5 1 71 0.5 0 CT ABA WS ABA+WS CT ABA WS ABA+WS 24h 1.05 1.22 0.105 0.023 0.08 0.35 1 0.11 48h 0.56 1.47 1.66 1.65 0.2 1.12 0 0.33 Figure 4.13: (A) Gel pictures of RT- PCR products of EF556121 (PERAPX) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of EF556121 (PERAPX) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. induced under ABA treatment but not under WS treatment. While comparing the expression data of this gene with activities and specific activities (Table 4.12) of APX obtained in same samples, only higher induction of gene under ABA treatment in C306 might be correlated to higher activities and specific activities of enzymes obtained in same samples. Otherwise this gene expression does not follow the pattern obtained with activity data. Reason might be the contribution of other APX isoforms in activities besides this gene or there might be the regulation at other steps rather than at transcriptional level. 4.3.2 Catalase (CAT) Catalases are tetrameric heme containing enzymes which directly dismutate H2O2 into H2O and oxygen. Catalases play important role in removal of H2O2 generated in peroxisomes by oxidases involved in ß-oxidation of fatty acids, photorespiration and purine catalases. Higher CAT specific activities were found in C306 than PBW343 in both roots and shoots of control seedlings. In roots of both cultivars, catalase activities as well as specific activities were mostly decreased under all stress treatments (Table 4.13; Figure 4.14, 4.15). Catalase was found to be maintained at higher levels (at activities and specific activities) mainly under ABA treatment in shoots of C306 only but not in shoots of PBW343. Under other stress treatments (WS and combined stress), catalase was not found to be altered in shoots of both cultivars. Catalase activities and H2O2 contents were in positive correlation in C306 shoots of control as well as stress treated seedlings which are in agreement with other reports which suggests that catalase gene expression and activity are controlled by H2O2 through H2O2 signal transduction pathway initiated under dehydration signal (Bailly et al 2008). Two catalase genes CAT-1 and CAT-2 (D86327, CAT-1; X94352, CAT-2) were used for gene expression analysis. Expression of CAT-1 was slightly induced at 24h stage under ABA treatment in shoots of PBW343 only but not expressed in any other samples (Figure 4.16(a), 4.16(b)). CAT-2was also induced under ABA treatment in both cultivars but expression level was higher in PBW343 than in C306. Comparing (CAT-1 and CAT-2) gene expression with activities and specific activities of catalases obtained in the same samples (Table 4.13); a good correlation was not found between two, though catalase activities were higher under ABA treatment. There can be many reasons for it like catalase isoforms contributing to activities in these samples might be different than the forms encoded by these two genes (CAT-1 and CAT-2). Secondly, catalase enzyme is suggested to be controlled at other levels (translational and protein turnover number) rather than at transcriptional level (Luna et al 2004). There are reports in literature where catalase (CAT-1 and CAT-2) gene expression was reduced but activities increased in the same samples as studied in leaves of wheat plant under drought ( Luna et al 2004) and these genes were suggested to be regulated through circadian control and by light. Our observations for catalase gene expression and activities in this study might be due these factors. 72 Table 4.13: Catalase activity (µmole of H2O2 disappeared min-1 g-1 DW) and specific activity (µmole of H2O2 disappeared min-1 mg-1 protein) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 0h 24h 73 48h 72h 0h 24h 48h 72h C306 CT ABA WS 752.70±113.05a (2.82±0.78)a 6147.62±779.19fg (20.10±4.79)bcde 6978.87±1748.79g (15.22±5.42)bc 4335.98±1255.26de (23.19±1.88)cde 752.70±113.05a (2.82±0.78)a 531.64±52.86a (2.73±0.06)a 1t241.78±244.58c (8.98±2.38)bc 1759.76±534.39d (12.16±3.81)bcd 1836.65±541.24d (23.02±7.26)e CT ABA WS ABA+WS 752.70±113.05a (2.82±0.78)a ABA+WS Shoot 752.70±113.05a (2.82±0.78)a 695.56±102.26a (2.93±0.27)a 695.56±102.26a (2.93±0.27)a 4350.13±32.71de (18.89±2.32)bcde 5363.81±1137.34ef (19.41±4.88)bcde 5165.66±131.40ef (27.19±2.55)e 4449.53±599.99de (27.29±8.58)e 2696.73±354.51bc (21.70±0.33)bcde 3966.98±522.94cde (26.20±7.38)de 3177.38±118.22bcd (14.17±2.68)b 2457.94±266.41b (18.41±1.56)bcd 2493.75±503.81b (20.09±2.55)bcde 3592.25±1311.03b (16.07±6.87)b 4720.88±406.16bcd (35.14±4.48)ef 5376.84±190.55cd (30.48±7.11)de 6208.23±739.72d (21.58±1.66)bcd 5353.06±951.68cd (35.12±7.20)ef 4226.50±318.38bc (45.72±8.34)f 695.56±102.26a (2.93±0.27)a 4530.66±1549.42bc (19.64±5.12)bc 3768.07±655.35b (34.65±0.75)e 3346.98±562.31b (27.15±4.53)cde 695.56±102.26a (2.93±0.27)a 3443.31±527.43b (18.56±4.09)bc 3629.56±462.47b (23.50±1.50)bcd 3771.95±1231.40b (19.93±8.48)bc 531.64±52.86a (2.73±0.06)a 1097.49±305.06abc (9.93±2.79)bcd 531.64±52.86a (2.73±0.06)a Root 531.64±52.86a (2.73±0.06)a 474.75±80.12a (2.08±0.42)a 474.75±80.12a (2.08±0.42)a 474.75±80.12a (2.08±0.42)a 474.75±80.12a (2.08±0.42)a 2972.55±510.02c (27.03±3.56)g 2491.50±154.22c (21.74±3.46)fg 1725.97±402.22b (16.04±3.75)de 817.72±363.58a (8.25±1.99)bc 535.14±136.47a (7.33±1.96)bc 554.12±86.28a (7.93±1.45)bc 2471.64±539.37c (16.31±2.87)de 995.56±181.29a (12.11±1.62)bcd 1604.85±76.86b (20.40±4.86)ef 1520.28±127.23b (19.73±2.39)ef 978.15±294.19a (12.58±4.34)cd 2045.43±468.46d (12.72±5.21)bcd 1038.46±395.77abc (11.29±4.07)bcd 511.34±177.03a (6.71±1.78)ab 806.12±11.48abc (11.38±0.68)bcd 629.38±122.07ab (11.81±0.28)bcd 747.53±76.29abc (10.59±1.62)bcd 1169.41±225.95bc (14.93±4.07)cd 751.50±146.22abc (16.65±3.72)de 534.58±66.55a (7.09±0.68)b *Values within parenthesis indicate specific activity and without parenthesis indicate activity. *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the various samples. umole of H2O2 disappeared /min/g DW A 10000 0h 24h 48h 72h g 9000 8000 7000 fg d ef 6000 de cd ef 5000 cd bcd b de de cde bc b bc b b b b 4000 bc bcd b 3000 b 2000 1000 a a a a a a a a 0 CT ABA WS ABA+WS CT ABA 4000 0h 24h 48h ABA+WS C306 PBW343 B WS 72h c 3500 umole of H2O2 disappeared /min/g DW c 3000 c d dd 2500 b 2000 b c 1500 a abc 1000 a a b abc bc abc a a abcab a a a abc a a a a a a a 500 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.14: Catalase activity (µmole of H2O2 disappeared min-1 g-1 DW) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the various samples. 74 umole of H2O2 disappeared per min per mg protein A 60 0h 24h 48h 72h f 50 ef ef 40 e de de e cde e 30 bcde cde bcde bcde bcde bc b bcd bcde 20 bcd 10 a a a a a a a a 0 ABA WS ABA+WS CT ABA 35 0h 24h 48h WS ABA+WS C306 PBW343 umole of H2O2 disappeared per min per mg protein bc b CT B bcd bc 72h g e 30 fg ef 25 ef 20 cd bcd bcd 15 bc de de cd bcd bcd bcd bcd bcd bcd bc ab 10 5 de a a a a a bc a a bc b a 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.15: Catalase specific activity (µmole of H2O2 disappeared min-1 mg-1 protein) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the various samples. 75 (A) PBW343 C306 24hr ABA+WS ABA 48hr WS CT ABA+WS ABA 24hr WS CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT X94352 (CAT-2) GAPDH (B) 24h PBW343 48h C306 0.8 0.6 0.4 76 0.2 0 Figure 4.16(a): CT ABA WS ABA+WS CT ABA WS ABA+WS 24h 0.045 0.69 0 0 0 0.08 0 0 48h 0 0 0 0 0 0 0 0 (A) Gel pictures of RT- PCR products of X94352 (CAT-2) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of X94352 (CAT-2) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. PBW343 (A) C306 24hr ABA+WS ABA WS 48hr CT ABA+WS ABA 24hr WS CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT D86327(CAT-1) GAPDH 24h PBW343 48h C306 0.08 (B) 77 Ratio 0.06 0.04 0.02 0 CT ABA WS ABA+WS CT ABA WS ABA+WS 24h 0 0.076 0 0 0 0 0 0 48h 0 0 0 0 0 0 0 0 Figure 4.16(b): (A) Gel pictures of RT- PCR products of D86327 (CAT-1) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of D86327 (CAT-1) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. 4.3.3 Peroxidases Peroxidases have a role in biosynthesis of lignin and in defense against biotic and abiotic stresses by consuming H2O2. Increased peroxidase activities are reported under abiotic stresses (Gill and Tuteja 2010). Peroxidases and ascorbate peroxidases (APX) can be distinguished from each other in terms of their gene sequences and their functions. Peroxidases prefer aromatic electron donor such as guaiacol and pyrogallol but oxidise ascorbate at the rate of around 1% to that with guaiacol. While ascorbate peroxidases have high degree of specificity for ascorbate as electron donor. Secondly, peroxidases participate in a number of processes like biosynthesis of lignin and are involved in processes of various plant responses like senescence, wounding of pathogens. APX has elemental role in scavenging H2O2 in chloroplast mainly. In this study, POX activities were determined using guaiacol as substrate hence enzyme was named as guaiacol peroxidases (GPOX). GPOX specific activities were not altered (Table 4.14, Figure 4.17, 4.18) but activities were decreased in roots of both cultivars under all stress treatments. Under ABA treatment, GPOX specific activities were increased in shoots of both cultivars but these increases were more prominent in shoots of C306. Under WS and combined stress, GPOX was almost unaltered in shoots of both cultivars. Higher APX, POD, CAT activities were reported to be related to drought tolerance in different plants (Guo et al 2006, Khanna-Chopra and Stelote 2007, Hameed et al 2011). These enzymes were also reported in downstream effect of ABA under WS in maize leaves where sequential effects downstream to ABA lead to these increases following increased levels of superoxide radicals and H2O2 (Jiang and Zhang 2002). In this study, higher CAT, APX, and POD level were found under ABA treatment in shoots of both cultivars but the effect was more pronounced in shoots of C306. Similar to results obtained with contents (H2O2, ascorbic acid, ascorbate/dehydroascorbate ratio) (Table 4.9) measurements, these three enzymes followed almost same pattern. It could be concluded that ABA led to increased contents of H2O2 at 24h mainly followed by increased contents of ascorbate as well as increased ascorbate/dehydroascorbate ratio and higher levels of these three antioxidant enzymes (CAT, APX, POD), can be ABA-dependent pathway under WS. 4.3.4 Glutathione reductase (GR) GR is a flavoprotein oxidoreductase which is potential enzyme in ascorbate-glutathione cycle. Secondly, it produces reduced glutathione (an antioxidant), hence maintains ROS. It is localized predominantly in chloroplast and small amount in cytosol and mitochondria. In roots of PBW343, GR activities were mostly decreased under all stress treatments (Table 4.15, Figure 4.19, 4.20). However in roots of C306, GR activities were not altered under almost all stress treatments, which might be the reason for maintaining higher ascorbate/dehydroascorbate ratio in these tissues (Table 4.9). In shoots of both cultivars, GR 78 79 Table 4.14: Guaiacol peroxidase activity (µmole of tetraguaicol appeared min -1 g-1 DW) and specific activity (µmole of tetraguaicol appeared min -1 mg-1 protein) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. GPX Shoot PBW343 C306 CT ABA WS ABA+WS CT ABA WS ABA+WS 0h 140.22±47.42a 140.22±47.42a 140.22±47.42a 140.22±47.42a 84.44±15.66a 84.44±15.66a 84.44±15.66a 84.44±15.66a (0.51±0.13)a (0.51±0.13)a (0.51±0.13)a (0.51±0.13)a (0.36±0.09)a (0.36±0.09)a (0.36±0.09)a (0.36±0.09)a 24h 576.92±27.92e 397.86±103.55bcd 374.79±6.07bcd 371.85±83.02bcd 290.32±60.01b 442.71±54.75d 342.30±46.22bc 317.52±20.52b bcde bcd cde bc ab bc bc (1.90±0.49) (1.69±0.29) (2.30±0.63) (1.63±0.33) (1.28±0.28) (1.55±0.22) (1.52±0.29) (1.70±0.20)bc 48h 313.27±138.99bc 469.28±96.62de 419.52±91.58cd 267.06±62.62b 459.00±33.58d 575.52±66.33e 413.51±59.33cd 431.32±23.63cd (1.13±0.47)ab (1.66±0.09)bc (3.38±0.67)f (1.97±0.49)cde (2.00±0.05)bcd (1.94±0.36)bcd (1.87±0.38)bcd (1.76±0.21)bcd cd de bcd bc cd bcd b 72h 412.57±32.30 474.83±24.11 392.21±26.06 295.21±5.24 412.40±77.72 385.73±62.69 309.66±50.51 292.40±43.46b (2.29±0.42)cde (2.50±0.20)de (2.57±0.56)ef (2.26±0.31)cde (2.80±1.28)de (3.25±1.28)e (2.49±0.24)cde (1.89±0.23)bcd Root 0h 800.00±26.20defg 800.00±26.20defg 800.00±26.20defg 800.00±26.20defg 634.33±132.57def 634.33±132.57def 634.33±132.57def 634.33±132.57def (3.94±0.48)a (3.94±0.48)a (3.94±0.48)a (3.94±0.48)a (2.78±0.61)a (2.78±0.61)a (2.78±0.61)a (2.78±0.61)a 24h 1461.94±336.20h 976.35±250.82g 954.26±13.92fg 825.09±155.38defg 710.02±10.39f 626.91±82.30def 458.84±126.77ab 423.39±41.20a ef def ef f bc bc b (10.35±1.38) (8.90±2.59) (10.42±0.39) (11.59±1.66) (7.16±1.11) (5.72±0.37)b (6.70±0.49) (5.73±1.07) 48h 868.64±74.48efg 665.23±138.62bcde 425.42±150.92ab 333.67±46.58a 1310.49±129.07h 1114.84±40.48g 495.85±146.85abcd 520.36±135.79abcde (5.99±0.48)abc (5.19±1.12)ab (5.72±2.06)abc (6.49±1.87)bcd (10.73±1.33)d (9.58±1.51)cd (8.69±2.60)bcd (9.65±4.12)cd cdef bcd bcde abc f ef abc 72h 704.26±8.50 588.60±34.79 682.08±4.02 464.91±16.51 749.26±37.05 676.59±36.96 459.88±15.19 447.25±78.42a (8.76±0.23)de (7.93±0.27)cde (8.64±0.87)de (8.24±1.55)cde (7.15±0.58)bc (5.70±0.96)b (8.29±0.87)bcd (6.12±0.32)b *Values within parenthesis indicate specific activity and without parenthesis indicate activity.*Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. A 700 0h 24h 48h 72h e umole of tetraguaicol produced /min/g DW e de 600 500 cd bcd de bccd d cd d bcd bcd bcd 400 b b b bc 300 a a a 200 a a a 100 a a 0 ABA WS ABA+WS CT ABA 2000 1800 0h 24h 48h ABA+WS 72h h 1600 h 1400 g g 1200 1000 defg 800 WS C306 PBW343 umole of tetraguaicol produced /min/g DW cd bc b b CT B cd bcd efg fg defg bcde defg cdef bcd defg defg bcde ab deff f defdef ef def def ababcd abcde abc 600 abc a a a 400 200 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.17: Guaiacol peroxidase activity (µmole of tetraguaicol appeared min -1 g-1 DW) in shoot (A) and of root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 80 umole of tetraguaicol produced per min per mg protein A 5 0h 24h 48h 72h e de 4.5 f 4 ef 3.5 de cde 3 cde 2.5 ab cde cdecde bcde bcdbc bc a a a a a 1 bcbcd bc bc ab 2 1.5 bcd bcd bcd a a bcd a 0.5 0 CT ABA WS ABA+WS CT ABA umole of tetraguaicol produced per min per mg protein 0h 24h 48h ABA+WS C306 PBW343 B 16 WS 72h cd f 14 ef 12 d def cd ef 10 de de cde 8 abc bcd cde bcd bcd abc bc ab bc bc b b b b 6 a a a a a 4 a a a 2 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.18: Guaiacol peroxidase specific activity (µmole of tetraguaicol appeared min -1 mg-1 protein) in shoot (A) and of root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 81 activities were almost unaffected under all stress treatments. GR was reported to be induced under water stress through ABA-dependent pathway (Jiang and Zhang 2002). In this study, unlike other antioxidants-enzymes (APX, POD, CAT), GR activities were not found to be increased under ABA. Secondly higher GR and glutathione contents are assumed to be related to drought resistance of plant under various stresses (Gill and Tuteja 2010). However in this study, GR in stressed samples was not found to be significantly different from unstressed samples. But one conclusion can be drawn that GR activities were decreased in roots of drought susceptible cultivar, PBW343 under stresses but these decreases were not observed in roots of C306, drought tolerant cultivar where higher ascorbate/ dehydroascorbate ratios were also maintained under stresses as compared to the ratios observed in roots of control seedlings (Table 4.9). 4.3.5 Superoxide dismutase (SOD) Three types of SOD have been reported in plants; Mn-SOD, Cu/Zn-SOD, Fe-SOD. Mn-SOD are located in mitochondria of eukaryotic cells and peroxisomes. Cu/Zn-SOD isozymes are located in chloroplast as well as in cytosol of higher plants. Fe-SOD is often not detected in plants if present then located in chloroplast. All SOD enzymes are encoded by nuclear genes and targeted to different compartments. SOD enzymes act on superoxide radical and dismutate it to H2O2 . Its activity has been found to be increased under stresses in various tissues of various plants (Gill and Tuteja 2010). In our studies, SOD activities and specific activities were determined (Table 4.16; Figure 4.21, 4.22). SOD specific activities increased in roots as well as in shoots of C306 under all stress treatments. However in PBW343, its specific activity increased at 48h and 72h stages under WS, at 48h stage under combined stress, unaltered under ABA treatment in shoots and increased under combined stress only in roots. It was clear that SOD level was higher in C306 than in PBW343 under all stress treatments. SOD is the only enzyme among all other studied in this study, which act on ROS particles other than H2O2, however all other were peroxidases which removes H2O2. SOD is usually considered to be the first line of defense against oxidative stress. SOD has also been reported to be increased under ABA treatment as well as under WS through ABA dependent pathway (Jiang and Zhang 2002). So higher level of SOD found in drought tolerant cultivar C306 might be related to its drought tolerance mechanism. In control seedlings of both cultivars, there were increases or decreases of antioxidant enzymes during growth of seedlings. Reason for such changes might be related to increasing or decreasing contents of H2O2 at the corresponding stages. As out of five antioxidant enzymes, four enzymes i.e. APX, CAT, GPOX and GR are involved in the regulation of H2O2 among other ROS species and H2O2 contents are related to metabolic activities of the cells, so changing trends of antioxidant activity might be controlling changing levels of H2O2 in these tissues. 82 Table 4.15: The glutathione reductase activity (nmole of NADPH+H+ disappeared min-1 g-1 DW) and specific activity (as nmole of NADPH+ H+ disappeared min-1 mg-1 protein) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 CT ABA C306 WS ABA+WS CT ABA WS ABA+WS 5785.24±2117.55abc (23.52±4.39)ab 5785.24±2117.55abc (23.52±4.39)ab 5785.24±2117.55abc (23.52±4.39)ab 83 0h 5119.10±1446.33a (18.90±5.42)a 5119.10±1446.33a (18.90±5.42)a 5119.10±1446.33a (18.90±5.42)a Shoot 5119.10±1446.33a 5785.24±2117.55abc (18.90±5.42)a (23.52±4.39)ab 24h 9458.01±1716.62b (30.66±6.46)abc 6035.39±1299.27a (25.81±3.38)ab 4829.16±1298.69a (29.25±8.90)abc 6096.53±1632.93a (26.60±4.97)ab 6314.62±1110.95abcd (27.81±3.71)ab 9063.40±459.11d (31.66±3.05)b 5411.47±2104.59abc (24.57±11.88)ab 5188.37±283.44abc (27.76±1.76)ab 48h 17370.55±4999.64c (64.11±26.28)d 7784.88±766.69ab (28.26±6.05)abc 5799.16±2004.15a (47.18±18.59)cd 6485.86±75.31ab (47.76±2.29)cd 5222.56±557.78abc (22.77±1.89)ab 5576.97±1626.84abc (18.44±4.39)ab 4483.11±1515.95abc (19.72±4.80)ab 4025.58±945.71a (16.18±2.73)a 72h 5652.02±572.79a (28.05±8.09)abc 4643.79±730.23a (24.50±5.06)a 5046.04±723.81a (28.69±2.34)abc 5864.60±600.41a (44.55±1.92)bcd 7317.43±2801.31cd (45.35±7.93)c 6867.32±1693.13bcd (56.49±20.57)c 3533.45±1062.17a (28.03±5.59)ab 4550.53±1214.44abc (29.35±6.82)ab 5739.84±983.47c 5739.84±983.47c 5739.84±983.47c (25.03±3.77)abc 2105.57±764.39a (25.03±3.77)abc 3391.18±215.59ab (34.49±5.66)abc (25.03±3.77)abc 2464.09±713.21a 0h bcd 3734.43±794.47 (18.29±4.90)a 24h 4486.06±956.59d 48h (31.79±2.70)abc 6300.71±1405.41e (43.51±10.19)cd 72h 5660.00±508.21 f e 3734.43±794.47 bcd 3734.43±794.47 bcd (18.29±4.90)a 3082.92±898.77bc (18.29±4.90)a 2648.69±247.27ab (28.28±10.38)abc 3317.78±546.98bcd (28.90±2.05)abc 2844.70±233.06abc (26.18±6.18)ab 3096.53±20.99bc (38.21±1.21)bcd 3958.59±373.22cd (51.68 ± 11.84)de cd Root 3734.43±794.47bcd 5739.84±983.47c (18.29±4.90)a (25.03±3.77)abc 1734.66±56.76a 4858.33±165.83bc ab (24.49±1.92) (44.30±0.27)cd 3354.50±470.58bcd (63.62±22.8)ef 3179.71±210.69bc (62.08 ± 6.77)ef 2645.80±1089.82a (21.63±9.07)ab 6641.13±2007.03 (63.82 ± 22.03)d c (24.47±7.57)abc 2048.67±168.45a (17.47±0.69)a 5047.16±1628.35c 1754.30±442.76a (32.88±6.16)abc 1771.12±725.09a (31.27±10.81)abc 3167.60±280.58a (33.27±18.79)abc 3040.74±687.10a (70.79±7.88) (43.73±1.95) (61.93±20.36)d (42.05±2.10)bcd (38.60±7.68)abc *Values within parenthesis indicate specific activity and without parenthesis indicate activity. *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. nmole of NADPH2 disappeared /min/g DW A 25000 0h c 24h 48h 72h 20000 15000 b cd ab 10000 a a aa a a a a a a a abc abcd ab a abc d abc bcd abc abc abc abc a abc abca abc 5000 0 CT ABA WS ABA+WS CT ABA 0h 24h 48h 72h c 9000 nmole of NADPH2 disappeared /min/g DW ABA+WS C306 PBW343 B 10000 WS e 8000 c 7000 c c c c e 6000 d 5000 bcd bc bcd bcbcd 4000 bc 3000 bcd cd ab abc bcd bcd bc a ab a a a a 2000 a a a a 1000 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.19: The glutathione reductase activity (nmole of NADPH+H + disappeared min-1 g-1 DW) shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 84 nmole of NADPH2 disappeared per min per mg protein A 100 0h d 24h 48h 72h c 90 80 cd 70 c 60 cd bcd 50 40 abc abc 30 a abc abc ab a a abc a ab a ab ab b ab ab ab ab ab ab a 10 0 ABA WS ABA+WS CT ABA 0h 100 90 24h 48h ABA+WS 72h ef f 80 d d ef de 70 cd 60 cd 50 abc ab a a abc a abc abc abcbcd abc abc cd bcd abc 40 30 WS C306 PBW343 nmole of NADPH2 disappeared per min per mg protein abab 20 CT B ab ab ab abc ab a abc abc abc abc a 20 10 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.20: The glutathione reductase specific activity (as nmole of NADPH+ H + disappeared min-1 mg-1 protein) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 85 Table 4.16: SOD activity (SOD units g-1 DW) and specific activity (SOD units mg-1 protein) in shoot (upper panel) and root (lower panel) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. PBW343 0h 24h 48h 72h C306 ABA WS 00.00±0.00 (0.00±0.00) 145.47±41.99a (0.50±0.27)ab 80.94±23.27a (0.31±0.13)a 98.26±39.30a (0.76±0.26)b 00.00±0.00 (0.00±0.00) 134.03±64.37a (0.56±0.23)ab 149.83±40.39ab (0.53±0.08)ab 123.93±32.38a (0.65±0.15)ab 00.00±0.00 (0.00±0.00) 135.60±17.07a (0.83±0.20)b 95.64±7.21a (0.78±0.14)b 227.61±71.98b (1.46±0.36)c 00.00±0.00 (00.00±0.00) 201.69±52.86c (1.48±0.57)bc 97.61±12.58ab (0.67±0.10)ab 99.75±24.25ab (1.24±0.33)abc 00.00±0.00 (00.00±0.00) 197.72±110.23c (1.74±0.86)cd 84.18±3.04ab (0.66±0.07)ab 53.31±31.80a (0.69±0.34)ab 00.00±00.00 (00.00±0.00) 144.00±17.55bc (1.57±0.16)c 43.25±2.32a (0.58±0.07)a 96.11±12.54ab (1.21±0.04)abc 86 CT 0h 24h 48h 72h ABA+WS Shoot 00.00±0.00 (0.00±0.00) 114.55±51.54a (0.51±0.25)ab 96.46±9.74a (0.71±0.09)b 96.36±45.14a (0.71±0.25)b Root 00.00±0.00 (00.00±0.00) 168.69±19.45c (2.39±0.38)de 48.75±22.84a (0.97±0.58)abc 75.91±39.23ab (2.97±0.97)e CT ABA WS ABA+WS 00.00±0.00 (0.00±0.00) 144.02±59.24c (0.61±0.17)cd 66.32±28.68ab (0.30±0.15)ab 45.02±6.23a (0.26±0.10)a 00.00±0.00 (0.00±0.00) 116.80±70.79bc (0.42±0.27)abc 109.01±46.31abc (0.36±0.15)abc 278.89±47.50d (1.14±0.09)e 00.00±0.00 (0.00±0.00) 82.87±41.85abc (0.39±0.23)abc 57.19±2.33ab (0.26±0.03)a 106.39±16.46abc (0.88±0.25)de 00.00±0.00 (0.00±0.00) 44.76±3.90a (0.25±0.001)a 83.76±34.10abc (0.35±0.18)abc 89.79±16.77abc (0.60±0.15)bcd 00.00±0.00 (00.00±0.00) 154.316±14.88cd (1.45±0.10)bcd 127.44±12.71bc (1.04±0.13)abc 122.16±20.70bc (1.17±0.23)abc 00.00±0.00 ((00.00±0.00)) 197.91±53.43d (2.04±0.12)e 116.27±7.54bc (1.00±0.19)ab 55.17±9.75a (0.65±0.09)a 00.00±0.00 (00.00±0.00) 95.78±9.59ab (1.04±0.10)abc 100.79±51.32ab (1.70±0.66)de 112.99±42.73bc (1.51±0.60)bcde 00.00±0.00 (00.00±0.00) 77.82±7.37ab (1.11±0.07)abc 88.35±18.69ab (1.57±0.06)cde 89.94±23.03ab (1.16±0.35)abc *Values within parenthesis indicate specific activity and without parenthesis indicate activity. *Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. A 350 0h 24h 48h 72h d b 300 SOD units /g DW 250 200 a a c ab a a 150 bc a a a abc a abc abc a a abc abc ab 100 ab a a 50 0 CT ABA WS ABA+WS CT ABA 0h 24h 48h ABA+WS C306 PBW343 B 350 WS 72h c 300 SOD units / g DW c d 250 c 200 cd bc bcbc 150 ab ab ab ab a 100 a ab ab bc bc ab ab ab ab a a 50 0 CT ABA WS ABA+WS CT ABA WS ABA+WS C306 PBW343 Fig. 4.21: SOD activity (SOD units g-1 DW) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. SOD activity could not be detected at 0h stage in any of above mentioned samples. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 87 A 2 0h 24h 48h 72h c SOD units per mg protein 1.8 1.6 1.4 e 1.2 b b 1 0.8 b b ab ab ab de ab b cd bcd abc ab 0.6 abc ab a abc a 0.4 abc a a 0.2 0 CT ABA WS ABA+WS CT ABA 4.5 0h 24h 48h ABA+WS C306 PBW343 B WS 72h e SOD units per mg protein 4 3.5 de 3 2.5 2 cd bc c abc abc abc 1.5 1 de bcde e ab ab abc ab cde bcd abc ab abc abc abc a a 0.5 0 CT ABA WS ABA+WS PBW343 CT ABA WS ABA+WS C306 Fig. 4.22: SOD specific activity (SOD units mg-1 protein) in shoot (A) and root (B) of germinating wheat seedlings of two cultivars PBW343 and C306 where stress was applied on 4 th day after germination as ABA (20 µM ABA), WS (6% Mannitol) and ABA+WS (20 µM ABA and 6% Mannitol) treatments. Data was collected at 24h, 48h, 72h after stress treatment and compared with similarly aged control seedlings (Ct) grown on distilled water as well as with 0h stage taken just before applying stress treatment. SOD activity could not be detected at 0h stage in any of above mentioned samples. Estimations were done on samples extracted in triplicates. Mean ± SD was calculated. Data of root and shoot in each cultivar was analyzed separately by Duncan’s Multiple Test at P < 0.05 for test of significance, where superscripts of similar alphabets indicate no significant difference among the values. 88 4.4 Late Embryogenesis Abundant Protein (LEA) LEA (Late Embryogenesis Abundant Protein) proteins are expressed at high levels during later stages of embryo development (post abscission) in plant seeds as at this stage in the developmental process, orthodox seeds acquire ability to withstand extreme dehydration. LEA proteins have been associated with dessication tolerance (Tunnacliffe and Wise 2007). LEA proteins have been classified into 9 groups based upon sequence similarities and conserved domains (Hundertmark and Hincha 2008). Among nine groups, role of only group 1, 2 and 3 has been investigated in imparting abiotic stress tolerance. ABA induces the expression of LEA genes during seed development and abiotic stresses (Finkelstein et al 2002, Dalal et al 2009). However all LEA genes were not responsive to ABA , they represent a major group of downstream genes involved in ABA-dependent and –independent signal pathways (Hong-Bo et al 2005, Kobayashi et al 2008a, 2008b). In the present study, LEA genes belonging to three groups; group 2, group 3, group 4 were studied for their expression level by semi-quantitative RT- PCR, where internal control of the constitutive gene GAPDH (glyceraldehyde-3-phosphate dehydrogenase) was used. This work was done in shoots of both cultivars at 24h and 48h after applying stress. All four similar treatments were used i.e. 20µM for ABA stress, 6% mannitol for WS and 20µM for ABA plus 6% mannitol for combined stress and shoots of control seedlings growing on distilled water was used as control to compare the expression level under stresses versus unstressed. This work was done by designing primers from known sequences of LEA genes available in database. Database was searched for nucleotide, mRNA, ESTs and protein sequences for LEA genes/proteins in wheat plant at NCBI (www.ncbi.nlm.nih.gov/). Primers were designed from sequences either manually or using Primer BLAST available at NCBI. Gene ID (NCBI Accession no.), gene name and primer sequences are summarized in Table 3.1. Some of these sequences belong to cDNA sequences isolated from Triticum aestivum and found to be 100% similar to dehydrin sequences isolated from Triticum durum. SemiQuantitative RT-PCR was done on total RNA isolated from shoots of above mentioned samples. RT-PCR products were run on 1.5% agarose gel and were pictured using Gel DocTM XR (Biorad). The intensities of bands were measured using Quantity One software (Biorad Laboratories Inc.). The ratio for each band was calculated by dividing the intensity of that band with intensity of band obtained with GAPDH primers in same sample. Results are presented in Figure 4.23- 4.33. 89 (A) PBW343 C306 24hr ABA+WS ABA 48hr WS CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT Td16 GAPDH (B) 3.5 24h 48h PBW343 C306 3 Ratio 2.5 2 90 1.5 1 0.5 0 24h 48h CT 1.52 0.39 ABA 1.37 1.68 WS 3.26 1.59 ABA+WS 2.4 1.92 CT 2.22 0 ABA 1.81 1.6 WS 1.52 2.71 ABA+WS 2.22 0.88 Figure 4.23: (A) Gel pictures of RT- PCR products of CV762802 (similar to X78429 (Td16) of durum wheat) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of CV762802 (similar to X78429 (Td16) of durum wheat) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. PBW343 (A) C306 24hr ABA+WS ABA WS 48hr CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT Td25a GAPDH (B) 1.2 24h 48h PBW343 C30 1 91 Ratio 0.8 0.6 0.4 0.2 0 24h 48h Figure 4.24: CT 1.07 0 ABA 0.563 0.36 WS 0.369 0 ABA+WS 0.161 0 CT 0.31 0 ABA 0.12 0.18 WS 1.09 0.79 ABA+WS 0.39 0.14 (A) Gel pictures of RT- PCR products of DR739608 (similar to X78431 (Td25a) of durum wheat) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of in the form of ratio of DR739608 (similar to X78431 (Td25a) of durum wheat) intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. (A) PBW343 C306 24hr ABA+WS ABA WS 48hr CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT Td27e GAPDH (B) 3 24h 48h C306 PBW343 2.5 92 Ratio 2 1.5 1 0.5 0 24h 48h CT 1.12 0.69 ABA 0.81 1.41 WS 2.15 1.33 ABA+WS 1.81 1.31 CT 1.45 0.77 ABA 1.27 1.38 WS 1.04 2.57 ABA+WS 1.59 0.52 Figure 4.25: (A) Gel pictures of RT- PCR products of AL815683 (similar to X78431 (Td27e) of durum wheat) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of in the form of ratio of AL815683 (similar to X78431 (Td27e) of durum wheat) intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. (A) PBW343 C306 24hr ABA+WS ABA 48hr WS CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT Td11 GAPDH 3 (B) 24h 48h PBW343 C306 2.5 93 Ratio 2 1.5 1 0.5 0 24h 48h Figure 4.26: CT 1.49 1.66 ABA 1.08 1.6 WS 2.67 1.56 ABA+WS 2.22 1.49 CT 1.85 0.96 ABA 1.54 1.39 WS 1.42 2.27 ABA+WS 1.89 1.1 (A) Gel pictures of RT- PCR products of U73211 (WCOR410 (similar to AJ890140, Td11 of durum wheat) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of U73211 (WCOR410 (similar to AJ890140, Td11 of durum wheat) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. PBW343 (A) C306 24hr ABA+WS ABA 48hr WS CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT Wdhn 13 GAPDH 24h 2.5 48h (B) PBW343 C306 94 Ratio 2 1.5 1 0.5 0 24h 48h Figure 4.27: CT 1.72 1.31 ABA 1.43 1.35 WS 1.19 1.61 ABA+WS 2.33 1.4 CT 0.77 0.44 ABA 1.51 1.04 WS 1.11 0.77 ABA+WS 0.92 0.55 (A) Gel pictures of RT- PCR products of AB076807 (Wdhn 13) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of AB076807 (Wdhn 13) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. Among five LEA genes belonging to group 2 (Figure 4.23- 4.27), Wdhn 13 LEA gene was induced more under ABA than under WS in C306 otherwise other LEA genes were induced more under WS than under ABA in both cultivars. This gene was neither responsive to ABA nor induced under WS in PBW343 but expressed in control shoot. Among other four LEA genes; two LEA genes CV762802 (similar to Td16 of durum plant) and AL815683 (similar to Td27e of durum plant) were responsive to ABA and other two, DR739608 (similar to Td25a of durum plant) and U73211 (WCOR410, similar to Td11 of durum plant) were weakly responsive to ABA in both cultivars. Under WS treatment, all four were induced under WS in both cultivars but DR739608 (similar to Td25a of durum plant) was expressed only in C306 and Al815683 (similar to Td27e of durum plant) was induced by more amounts in C306 than PBW343. Td16, Td27e, Td25a were reported to be more expressed under WS, while Td11 responded to cold, salt stress, ABA application but not to WS in wheat (Ali-Benali et al 2005). In another study, WCOR410 (Td11) was found to expressed only in freezing tolerant cultivar (Danyluk et al 1994, 1998). Among these five group 2 Lea genes; CV762802 (similar to Td16 of durum plant) and AL815683 (similar to Td27e of durum plant) and U73211 (WCOR410 similar to Td11 of durum plant) were induced earlier (24h) in PBW343 but later (48h) in C306. Accumulation of Td27e, Td25a was reported to be delayed (67h) in drought tolerant cultivar but earlier (30h) in drought susceptible cultivar in both root and shoot, moreover these two genes were found to be responsive to ABA and induced by more amount in drought tolerant cultivar than drought susceptible cultivar (Labhili et al 1995). In present study, these two genes (Td25a and Td27e) were similarly found to be more induced or induced only in drought tolerant cultivar but for their sensitivity towards ABA, Td27e was more responsive to ABA than Td25a in drought tolerant cultivar and Td27e was induced earlier (24h) in drought susceptible cultivar, PBW343 and later (48h) in drought tolerant cultivar, C306 where Td25a was induced only in tolerant cultivar not in drought susceptible cultivar. Comparing cultivars for ABA response in terms of expression status of group 2 LEA genes in shoots, Wdhn13 gene was responsive to ABA as well as induced under WS in drought tolerant cultivar C306 but was neither responsive to ABA nor induced under WS in drought susceptible cultivar, PBW343 (though expressed in its control shoots), hence it might be involved in ABA- dependent pathway of drought tolerance. In literature, it is found to be ABA-responsive in Triticum aestivum (Kobayashi et al 2006) as well as in Ae. Tauschii (Kurahashi et al 2009) and its level of expression was also correlated to freezing tolerance, drought tolerance as well as ABA-sensitivity. It is also reported to be induced under salt stress besides drought and low temperature (Ohno et al 2003). 95 (A) PBW343 C306 24hr ABA+WS ABA WS 48hr CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT AF255052 (Wrab 19) GAPDH (B) 3 24h 48h PBW343 C306 96 Ratio 2.5 2 1.5 1 0.5 0 24h 48h CT 1.35 0 ABA 0.63 0.68 WS 1.5 1.19 ABA+WS 1.18 0.74 CT 1.18 0 ABA 1.33 0.69 WS 0.93 2.7 ABA+WS 1.77 0 Figure 4.28: (A) Gel pictures of RT- PCR products of AF25505 (Wrab 19) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of AF255052 (Wrab 19) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. PBW343 (A) C306 24hr ABA+WS ABA 48hr WS CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT AB115913 (Wrab 15) GAPDH 2.5 (B) 24h 48h PBW343 C306 97 Ratio 2 1.5 1 0.5 0 24h 48h Figure 4.29: CT 1.43 0.56 ABA 0.8 0.88 WS 1.85 1.76 ABA+WS 1.92 1.68 CT 0.54 0 ABA 1.56 0.68 WS 1.27 2.01 ABA+WS 1.94 1.08 (A) Gel pictures of RT- PCR products of AB115913 (Wrab 15) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of AB115913 (Wrab 15) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. (A) PBW343 C306 24hr ABA+WS ABA WS 48hr CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT AF255053 (Wrab 17) GAPDH (B) 2 24h 48h PBW343 C306 1.8 1.6 98 Ratio 1.4 1.2 1 0.8 0.6 0.4 0.2 0 24h 48h CT 1.21 0.75 ABA 0.89 0.82 WS 1.87 1.36 ABA+WS 1.28 0.98 CT 1.05 1.46 ABA 1.07 1 WS 1.03 0.31 ABA+WS 0.8 0 Figure 4.30: (A) Gel pictures of RT- PCR products of AF255053 (Wrab 17) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of AF255053 (Wrab 17) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. PBW343 (A) C306 24hr ABA+WS ABA WS 48hr CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT AB115914 (Wrab18) GAPDH (B) 3 24h 48h PBW343 C306 99 Ratio 2.5 2 1.5 1 0.5 0 24h 48h CT 1.39 0 ABA 0.64 0.76 WS 1.66 2 ABA+WS 1.22 1.63 CT 1.18 0 ABA 1.39 0.17 WS 1.01 2.49 ABA+WS 1.54 0 Figure 4.31: (A) Gel pictures of RT- PCR products of AB115914 (Wrab18) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of AB115914 (Wrab18) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. (A) PBW343 C306 24hr ABA+WS ABA 48hr WS CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT AY148490 GAPDH (B) 1.6 24h 48h PBW343 C306 1.4 10 0 Ratio 1.2 1 0.8 0.6 0.4 0.2 0 24h 48h CT 1.5 0 ABA 0.044 0 WS 0.881 0.69 ABA+WS 0.312 0.79 CT 0.61 0 ABA 1.42 0.31 WS 0.78 1.34 ABA+WS 1.43 0 Figure 4.32: (A) Gel pictures of RT- PCR products of AY148490 (TaLEA-3 like) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4 th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of AY148490 (TaLEA-3 like) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. (A) PBW343 C306 24hr ABA+WS ABA 48hr WS CT ABA+WS ABA WS 24hr CT ABA+WS ABA WS 48hr CT ABA+WS ABA WS CT AY148490 GAPDH (B) 3.5 24h 48h PBW343 C306 3 10 1 Ratio 2.5 2 1.5 1 0.5 0 24h 48h Figure 4.33: CT 1.75 0.9 ABA 1.89 1.76 WS 3.28 1.71 ABA+WS 2.72 1.67 CT 1.7 0 ABA 1.62 1.22 WS 1.31 2.94 ABA+WS 1.92 0.95 (A) Gel pictures of RT- PCR products of GH729039 (similar to AJ890139 (Td29) of durum wheat) and GAPDH (internal control) where semiquantitative RT- PCR was done on total RNA isolated from shoots of germinating wheat seedlings of two cultivars PBW343 and C306 treated on 4th day of germination with ABA (20 µM), WS (6% mannitol), ABA+ WS (20 µM and 6% mannitol). Experiment was performed at 24h and 48h after stress applications and compared with shoots of control seedling (CT) growing on distilled water for similar time period. (B) Quantitative expression level of GH729039 (similar to AJ890139 (Td29) of durum wheat) in the form of ratio of intensity of band obtained with gene specific primer to the intensity of band obtained with GAPDH specific primer. LEA Group3 Wrab genes (Wrab15, Wrab17, Wrab18, Wrab19) were observed as weakly- or non-responsive to ABA e.g. Wrab17 and Wrab18 were non-responsive (rather downregulated under ABA treatment), Wrab19 was weakly responsive to ABA in both cultivars but Wrab 15 and another gp3 gene (LEA gp 3-like gene) were responsive to ABA mainly in C306 cultivar. As opposed to ABA treatment, these genes were induced under WS, where Wrab19, Wrab15, Wrab18 and Ta LEA-3 like were more induced in C306 than in PBW343, while Wrab17 was induced only in PBW343 not induced in C306. All five LEA gp3 genes were induced earlier (24h) in PBW343 while later (48h) in C306. In literature, Wrab genes were reported to be responsive to ABA, low temperature, drought (Kobayashi et al 2004, 2006, 2008, Kurahashi 2009, Egawa et al 2006) but almost all of this work was done in two wheat cultivars M808 (winter wheat, freezing tolerant) and CS (spring wheat freezing susceptible) where expression levels of these genes were more in tolerant cultivar, M808 than in susceptible cultivar, CS under ABA treatment but were not different between cultivars under low temperature (Egawa et al 2006, Kobayashi et al 2004, 2006, 2008). But these genes (except Wrab17) were actually not so good responsive to ABA, if these expressed, stayed for 2-5 hr after the treatment and not stayed like as under low temperature treatment (Kobayashi et al 2004, 2006). Reasons for such temporary response under ABA but stable response under stress treatment were not clear though suggested to be probable metabolization of exogenously applied ABA (Kobayashi et al 2004). In this study, adding exogenous ABA with WS (combined stress) gave response more similar to ABA treatment than to WS in drought tolerant C306 cultivar as 5of 11 LEA genes (belonging to all three gps 2,3,4) as these were induced under WS and remain un-induced or less induced under combined stress (Figures 4.23 to 4.33). It indicated that ABA might be inhibiting or down-regulating the expression of these genes in C306 cultivar. Our observations agreed to the results (Kobayashi et al 2006) of these genes expression in another wheat cultivar Kitakei1354 (ABA hypersensitive and dormant winter wheat) where unlike M808 cultivar, these genes were downregulated under ABA treatment while in another cultivar EH47-1 (derived from Kitakei1354 by single mutation, which became ABA-sensitive and nondormant but more freezing tolerant than Kitakei1354), these genes were responsive to ABA like as observed in freezing tolerant M808 cultivar. Under low temperature, these genes were almost equally induced in both EH47-1 and Kitakei-1354. Drought tolerant cultivar, C306 used in this study was also found to be higher sensitive to ABA than drought susceptible cultivar, PBW343 (16% germination in C306 as opposed to 81% in PBW343 in the presence of 20 µM ABA on 6th day of germination, unpublished results). It can be concluded that LEA genes’ expressions are controlled by both ABA-dependent and ABA-independent pathways under water stress but dominating effects of ABA over the effects of WS when applied both 102 together in combined stress observed in C306 not in PBW343, might be due to higher ABAsensitivity of C306 and ABA-insensitivity of PBW343. Only one gene GH729039 (similar to AJ890139 (Td29) of durum wheat) of LEA gp 4 was studied. Response of this gene was similar to gp2 LEA genes (studied in the present study) than to gp3 LEA genes. This gene was poorly responsive to ABA but induced under WS in both cultivars, however induction level was higher in C306 than in PBW343. It was induced earlier in PBW343 (24h) and later in C306 (48h). 103 Chapter V SUMMARY Two wheat cultivars contrasting in drought tolerance were compared for their response to exogenous ABA, water stress (WS) and combined stress (ABA plus WS) during germination. Their response was studied in the form of growth and antioxidant potential in roots and shoots at different hours (0h, 24h, 48h, 72h) and in the form of LEA gene expression in shoots at 24h and 48h after stress treatment where stress was given on 4th day of germination. Responses were compared with similarly old control seedlings growing on distilled water and with 0h stage taken on 4th day of germination before applying stress treatment. Such responses were also compared among stresses and between cultivars. ABAdependent maintenance of dry weights under WS as well as combined stress was found in shoots and roots of drought tolerant cultivar, C306 but lacked in drought susceptible cultivar, PBW343. Drought tolerant cultivar, C306 showed lower water contents throughout its stress period while water contents were higher and regained after 24h of stress in drought susceptible cultivar, PBW343. Exogenous application of ABA led to increased H2O2 contents in roots of C306 at 24h, followed by increase in ascorbate content, ascorbated/ dehydroascorbate ratio, activities of catalase, ascobate peroxidase and guaicol peroxidase in shoots; decreased membrane toxicity in both shoots and roots. Such types of responses were poorly given by PBW343 under exogenous ABA supply. Under WS, both cultivars behaved differently where above mentioned ABA-dependent responses were given by C306 but were lacked in PBW343. Under combined stress, above mentioned responses were similarly produced as under WS and as under ABA supply in C306, however in PBW343, such responses were more similar to as under ABA than to as under WS. It indicated the existence of different pathways for ABA and for WS where ABA-dependent pathway was working in C306 but lacked in PBW343 under WS. Drought tolerant cultivar showed lesser level of membrane oxidation and higher levels of SOD. MDA contents were not related to H2O2 contents. Proline contents were higher in roots of drought tolerant cultivar, C306 under unstressed conditions and on exposure to WS, this cultivar accumulated more proline contents in its roots. However drought susceptible cultivar, PBW343 contained lesser contents of proline under unstressed conditions and on exposure to WS, it accumulated proline contents mainly in its shoots. Catalases and H2O2 content were positively correlated in shoots of C306 in control as well as in stressed samples. Eleven LEA genes, five of gp2, five of gp3 and one of gp4, were studied for their expression status in shoots of both cultivars at 24h and 48h after stress treatment under same stresses (ABA, WS, ABA plus WS). Results were compared with control shoots of same age, among stresses and between cultivars. All 11 LEA genes were more induced under WS than under ABA in both cultivars except Wdhn13 which was more induced under ABA than under WS in C306. All LEA genes were responsive to ABA in both cultivars except Wdhn13 (responsive to ABA in C306 only but not in PBW343) and Wrab17 (ABA-responsive in PBW343 not in C306). Among all 11 LEA genes, 6 (three of gp2, Td25a, Td27e, Wdhn13; two of gp3 Wrab1, Ta LEA gp3-like; one gp4, Td29) were more induced or induced only in C306, one LEA gene of gp3 (Wrab17) was induced only in PBW343 and others four (two of gp2, Td11, Td16; two of gp3, Wrab18, Wrab15) were equally induced in both cultivars under WS. Gp2 and Gp4 LEA genes were induced earlier (24h) in PBW343 and later (48h) in C306 while gp3 genes were induced mainly at 48 h in both cultivars under WS. 105 REFERENCES Abu-Abied M, Golomb L, Belausov E, Huang S, Geiger B, Kam Z, Staiger C J and Sadot E (2006) Identification of plant cytoskeleton-interacting proteins by screening for actin stress fiber association in mammalian fibroblasts. Plant J 48: 367–379. Addicott F T, Lyon J L, Ohkuma K, Thiessen W E, Carns H R, Smith O E, Cornforth J W, Milborrow B V, Ryback G and Wareing P F (1968) Abscisic acid: a new name for abscisin II (dormin). Science 78: 1479: 1493. Agarwal P K, Agarwal P, Reddy M K and Sopory S K (2006) Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep 25: 1263-1274. Agarwal S and Pandey V (2004) Antioxidant enzyme responses to NaCl stress in Cassia Angustifolia. Biol Plant 48 (4): 555-560. Alexieva V, Sergiev L, Mapelli S and Karanove (2001) The effects of drought and ultraviolet radiations on growth and stress markers in pea and wheat. Plant Cell Environ 24: 1337-1344. Al-Hakimi and Hamada A M (2001) Counteraction of salinity stress on wheat plants by grain soaking in ascorbic acid, thiamine or sodium salicylate. Biol Plant 44: 253–261. Ali-Benali Mohamed, Alary Remi, Joudrier Philippe and Gautier Marie-Francoise (2005) Comparative expression of five Lea Genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochemica et Biophysica Acta 1730: 56-65. Alscher R G, Erturk N and Heath L S (2002) Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J Exp Bot 53: 1331–1341. Apel K and Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55: 373–399. Arvind P and Prasad M N V (2003) Zinc alleviates cadmium-induced oxidative stress in Ceratophyllum demersum L: a free-floating freshwater macrophyte. Plant Physol Biochem 41: 391-397. Athara H, Khanb A and Ashraf M (2008) Exogenously applied ascorbic acid alleviates saltinduced oxidative stress in wheat. Env Exp Bot 63: 224-31. Bailly Christophe, EL-Maarouf-Bouteau Hayat and Corbineau Francoise (2008) From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. C R Biologies 331: 806-814. Bailly Christophe, Leymarie Juliette, Lehner Arnaud, Rousseau Sandra, Come Daniel and Corbineau Francoise (2004) Catalase activity and expression in developing sunflower seeds as related to drying. J Exp Bot 55: 475-483. Barth C, De Tullio M and Conklin P L (2006) The role of ascorbic acid in the control of flowering time and the onset of senescence. J Exp Bot 57: 1657–65. Bartosz G (1997) Oxidative stress in plants. Acta Physiol Plant 19: 47-64. Bates L S, Waldren R P and Teare I D (1973) Rapid determination of free proline for water stress studies. Plant Soil 39:205-207. Battaglia M, Olvera-Carrillo Y, Garciarrubio A, Campos F and Covarrubias A (2008) The enigmatic LEA proteins and other hydrophilins. Plant Physiol 148: 6–24. Battista J R, Park M J and McLemore A E (2001) Inactivation of two homologues of proteins presumed to be involved in the desiccation tolerance of plants sensitizes Deinococcus radiodurans R1 to desiccation. Cryobiology 43: 133–139. Bellaire B A, Carmody J, Braud J, Gossett D R, Banks S W, Lucas M C and Fowler T E (2000) Involvement of abscisic acid-dependent and –independent pathways in the upregulation of antioxidant enzyme activity during NaCl stress in cotton callus tissue. Free Radical Res 33: 531-45. Bies-Ethève N, Gaubier-Comella P, Debures A, Lasserre E, Jobet E, Raynal M, Cooke R and Delseny M (2008) Inventory, evolution and expression profiling diversity of the LEA (late embryogenesis abundant) protein gene family in Arabidopsis thaliana. Plant Mol Biol 67: 107–24. Blokhina O, Virolainen E, Kurt V and Fagerstedt (2002) Antioxidants, Oxidative Damage and Oxygen Deprivation Stress: Annu Rev Plant Biol 91: 179-194. Bohnert H J, Nelson D E and Jensen R G (1995) Adaptations to environmental stresses. The Plant Cell 7: 1099-1111. Bolwell G P, Bindschedler L V, Blee K A, Butt V S, Davies D R, Gardner S L, Gerrish C and Minibayeva F (2002) The apoplastic oxidative burst in response to biotic stress in plants: a three-component system. J Exp Bot 53: 1367–1376. Bravo L A, Gallardo J, Navarrete A, Olave N, Martı´nez J, Alberdi M, Close T J and Corcuera L J (2003) Cryoprotective activity of a cold-induced dehydrin purified from barley. Plant Pysiol 118: 262–69. Bray E A (1993) Molecular responses to water deficit. Plant Physiol 103: 1035-1040. Browne J, Tunnacliffe A and Burnell A (2002) Anhydrobiosis: Plant desiccation gene found in a nematode. Nature 416: 38-42. Bueno P, Piqueras A, Kurepa J, Savoure A, Verbruggen N, Van Montagu M and Inze D (1998) Expression of antioxidant enzymes in response to abscisic acid and high osmoticum in tobacco BY-2 cell cultures. Plant Sci 138: 27-34. 107 Chalapathi Rao A S V and Reddy A R (2008) Glutathione reductase: a putative redox regulatory system in plant cells. in: N A Khan, S Singh and S Umar (Eds.) Sulfur Assimilation and Abiotic Stresses in Plants. Springer, The Netherlands 111-147. Chang C C, Ball L, Fryer M J, Baker N R, Karpinski S and Mullineaux P M (2004) Induction of ascorbate peroxidase 2 expression in wounded Arabidopsis leaves does not involve known wound-signalling pathways but is associated with changes in photosynthesis. Plant J 38: 499–511. Chen T H H and Murata N (2002) Enhancement of tolerance of abiotic stress by metabolic controlling oxidative stress in plants. J Exp Bot 53: 1331-1341. Chen† L, Ren† F, Zhong H, Jiang W and Li* X (2010) Identification and expression analysis of genes in response to high-salinity and drought stresses in Brassica napus. Acta Biochim Biophys Sin 42:154-162. Cheng W H, Endo A, Zhou L, Penney J, Chen H C, Arroyo A, Leon P, Nambara E, Asami T, Seo M, Koshiba T and Sheen J (2002) A unique short-chain dehydrogenase/ reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 14: 2723–2743. Chinnusamy V, Jagendorf A and Zhu J K (2005) Understanding and improving salt tolerance in plants. Crop Sci 45: 437–448. Chinnusamy V, Shumaker K and Zhu J K (2004) Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J Exp Bot 55: 225-236. Cho U and Seo N (2005) Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci 168: 113-120. Choi H, Hong J, Ha J, Kang J and Kim S Y (2000) ABFs, a family of ABA-responsive element binding factors. J Biol Chem 275: 1723–1730. Cornforth J W, Milborrow B V, Ryback P F and Wareing P F (1965) Identity of sycamore ‘dormin’ with abscisin II. Nature 205: 1269. Cramer G R (2002) Response of ABA mutants of Arabidopsis to salinity. Funct Plant Biol 29: 561–67. Cutler Sean R, Rodriguez Pedro L, Finkelstein Ruth R and Abrams Suzanne R (2010) Abscisic acid: Emergence of core signaling network. Annu Rev Plant Biol 61: 651679. Dalal M, Tayal D, Chinnusamy V and Bansal K C (2009) Abiotic stress and ABA-inducible group 4 LEA from Brassica napus plays a key role in salt and drought tolerance. J Biotechnol 139: 137-45. 108 Danyluk J, Houde M, Rassart E and Sarhan F (1994) Differential expression of a gene encoding an acidic dehydrin in chilling sensitive and freezing tolerant gramineae species. FEBS Letters 344(1): 20-24. Danyluk J, Perron A, Houde M, Limin A, Fowler B, Benhamou N and Sarhan F (1998 ) Accumulation of an acidic dehydrin in the vicinity of the plasma membrane during cold acclimation of wheat. Plant Cell 10(4): 623-38. Dat J, Vandenabeele S, Vranová E, Van Montagu M, Inzé D and Breusegem F (2000) Dual action of the active oxygen species during plant stress responses. CMLS Cell Mol Life Sci 57: 779–795. Davey M W, Van Monatgu M, Sanmatin M, Kanellis A, Smirnoff N, Benzie I J J, Strain J J, Favell D and Fletcher J (2000) Plant L-Ascorbic Acid: chemistry, function, metabolism, bioavailability and effects of processing. J Sci Food Agric 80: 825–860. Davletova S, Rizhsky L, Liang H, Shenqiang Z, Oliver D J, Coutu J, Shulaev V, Schlauch K and Mittler R (2005) Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen network of Arabidopsis. Plant Cell 17: 268–281. de Carvallo (2008) Drought stress and reactive oxygen species. Plant signaling and behavior 3(3): 156-165. de-Lacerda C F, Cambraia J, Oliva M A, Ruiz H A and Prisco J T (2003) Solute accumulation and distribution during shoot and leaf development in two sorghum genotypes under salt stress. Environ Exp Bot 49: 107–120. Desikan R, Hancock J T and Neill S J (2003) Oxidative stress signalling. Top Curr Genet 4: 129–149. Dubouzet J G, Sakuma Y, Ito Y, Kasuga M, Dubouzet E G, Miura S, Seki M, Shinozaki K and Yamaguchi-Shinozaki K (2003) OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high salt- and cold-responsive gene expression. Plant J 33: 135-149. Egawa C, Kobayashi F, Ishibashi M, Nakamura T, Nakamura C and Takumi S (2006) Differential regulation of transcript accumulation and alternative splicing of a DREB2 homolog under abiotic stress conditions in common wheat. Gene Genet Syst 81: 77–91. Eyidogan F and Oz M T (2005) Effect of salinity on antioxidant responses of chickpea Seedlings. Acta Plant Physiol 29: 485-493. Farooq M, Wahid A, Lee D J, Ito O and Siddique H M (2009) Advances in drought resistance of rice. Crit Rev Plant Science 28: 199-217. Finkelstein R R, Gampala S S L and Rock C D (2002) Abscisic acid signaling in seeds and seedlings. Plant Cell 14: 15-45. 109 Fowler S and Thomashow F (2002) Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 14: 1675-90. Foyer C H and Noctor G (2003) Redox sensing and signaling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol Plant 119: 355–364. Foyer C H and Noctor G (2005) Oxidant and antioxidant signaling in plants: a re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ 28: 1056-1071. Fujii H, Zhu J K (2009) Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction, and stress. Proc Natl Acad Sci USA 106: 8380–8385. Gapinska M, Skodowska M and Gabara M (2008) Effect of short- and long-term salinity on the activities of antioxidative enzymes and lipid peroxidation in tomato roots. Acta Physiol Plant 30: 11-18. Garay-Arroyo A, Colmenero-Flores J M, Garciarrubio A and Covarrubias (2000) Highly hydrophilic proteins in prokaryotes and eukaryotes are common during conditions of water deficit. J Biol Chem 275: 5668–5674. Garg N and Manchanda G (2009) ROS generation in plants: boon or bane? Plant Biosys 143: 83-96. Gill S S and Tuteja N (2010) Reactive Oxygen Species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48: 909-30. Gilles G J, Hines K M, Manfre A J and Marcotte W R Jr (2007) A predicted N-terminal helical domain of a group 1 LEA protein is required for protection of enzyme activity from drying. Plant Physiol Biochem 45: 389–399. Goeres D C, Van Norman J M, Zhang W, Fauver N A, Spencer M L and Sieburth L E (2007) Components of the Arabidopsis mRNA decapping complex are required for early seedling development. Plant Cell 19: 1549–1564. Goyal K, Tisi L, Basran A, Browne J, Burnell A, Zurdo J and Tunnacliffe A (2003) Transition from natively unfolded to folded state induced by dessication in an anhydrobiotic nematode protein. J Biol Chem 278: 12977-84. Goyal K, Walton L J and Tunnacliffe A (2005) LEA proteins prevent protein aggregation due to water stress. Biochem J 388: 151–157. Grelet J, Benamar A, Teyssier E, Avelange-Macherel M H, Grunwald D, and Macherel D (2005) Identification in pea seed mitochondria of a late embryogenesis abundant (LEA) protein able to protect enzymes from drying. Plant Physiol 137: 157–167. Guan L, Zhao J and Scandalios J G (2000) Cis-elements and trans-factors that regulate 110 expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response. Plant J 22: 87-95. Guo Z, Ou W, Lu S, Zhong Q (2006) Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol Biochem 44: 828-836. Guoxiong Chen, Krugman Tamar, Fahima Tzion, Korol B Abraham and Nevo Eviatar (2002) Comparative study on morphological and physiological traits related to drought resistance between xeric and mesic Hordeum spontaneum lines in Israel. Barley Genetics Newsletter 32: 22-23. Hameed Amjad, Bibi Noreen, Akhter Javed and Iqbal Nayyer (2011) Differential changes in antioxidants, proteases, and lipid peroxidation in flag leaves of wheat genotypes under different levels of water deficit conditions. Plant Physiol Biochem 49: 178-185. Hameed Amjad, Iqbal Nayyer and Malik Salman (2009) Mannose-Induced Modulation in Antioxidants, Protease Activity, Lipid Peroxidation, and Total Phenolics in Etiolated Wheat Leaves. J Plant Growth Regul 28: 58-65. Harinasut P, Poonsopa D, Roengmongkol K and Charoensataporn R (2003) Salinity effects on antioxidant enzymes in mulberry cultivar. Sci Asia 29: 109-113. Hartung W, Sauter A and Hose E (2002) Abscisic acid in the xylem: where does it come from, where does it go to? J Exp Bot. 53: 27–32. Heath and Packer (1968) Photoperoxidation in isolated chloroplasts. I. kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125: 180-198. Hoekstra F A, Golovina E A, Tetteroo F A and Wolkers W F (2001) Induction of desiccation tolerance in plant somatic embryos: how exclusive is the protective role of sugars? Cryobiology 43: 140–150. Hong-Bo S, Zong-Suo L and Ming-An S (2005) LEA proteins in higher plants: Structure, function, gene expression and regulation. Colloid and Surface B 45: 131-35. Honjoh K, Matsumoto H, Shimizu H, Ooyama K, Tanaka K, Oda Y, Takata R, Joh T, Suga K and Miyamoto T (2000) Cryoprotective activities of group 3 late embryogenesis abundant proteins from Chlorella vulgaris C-27. Biosci Biotechnol Biochem 64: 1656–1663. Hoth S, Morgante M, Sanchez J P, Hanafey M K, Tingey S V and Chua N H (2002) Genomewide gene expression profiling in Arabidopsis thaliana reveals new targets of abscisic acid and largely impaired gene regulation in the abi1-1 mutant. J Cell Science 115: 4891–4900. 111 Hsu S Y, Hsu Y T and Kao C H (2003) The effect of polyethylene glycol on proline accumulation in rice leaves. Biol Plant 46: 73–78. Hu X, Jiang M, Zhang A and Lu J (2005) Abscisic acid-induced apoplastic H2O2 accumulation up-regulates the activities of chloroplastic and cytosolic antioxidant enzymes in maize leaves. Planta 223: 57-68. Hua B and Guo W Y (2002) Effect of exogenous proline on SOD and POD activity of soyabean callus under salt stress. Acta Agric Boreali-Sinica 17: 37–40. Hundertmark M and Hincha D K (2008) LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 9: 118-140. Ishikawa Takahiro and Shigeoka Shigeru (2008) Recent advances in ascorbate biosynthesis and the physiological significance of ascorbate peroxidase in photosynthesizing organisms. Biosci Biotechnol Biochem 72 (5): 1143-1154. Jannat R, Uraji M, Morofuji M, Islam M, Bloom R E, Nakamura Y, McClung C R, Schroeder J I, Mori I C and Murata Yoshiyuki (2011) Roles of intracellular hydrogen peroxide accumulation in abscisic acid signaling in Arabidopsis guard cells. J Plant Physiol 18. Jia W, Xing Y, Zhao J, Zhang J and Zhang S Q (2002b) Cellular stress signaling from water deficit-perception to ABA accumulation. Adv Plant Sci 4: 205–212. Jiang M and Zhang J (2001) Effect of abscisic acid on active oxygen species, antioxidant defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol 42: 1265-73. Jiang M and Zhang J (2002a) Role of abscissic acid in water stress-induced antioxidant defense in leaves of maize seedlings. Free Radical Res 36: 1001-15. Jiang M and Zhang J (2002b) Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J Exp Bot 53: 2401-10. Jiang M and Zhang J (2002c) Involvement of plasma-membrane NADPH oxidase in abscisic acid- and water stress-induced antioxidant defense in leaves of maize seedlings. Planta 215: 1022-30. Jiang X and Wang Y (2004b) Elimination coupled with tandem mass spectrometry for the identification of in vivo and in vitro phosphorylation sites in maize dehydrin DHN1 protein. Biochemistry 43: 15567–15576. Jubany Mari T, Munne-Bosch S and Alegre L (2010) Redox regulation of water stress responses in field-grown plants. Role of hydrogen peroxide and ascorbate. Plant Physiol Biochem 48:351-358. 112 Kacperska A (2004) Sensor types in signal transduction pathways in plant cells responding to abiotic stressors: do they depend on stress intensity? Physiol Plant 122: 159–168. Kaminaka H, Morita S, Tokumoto M, Masumura T and Tanaka K (1999) Differential gene expressions of rice superoxide dismutase isoforms to oxidative and environmental stresses. Free Radical Res 219-225. Karlson D T, Fujino T, Kimura S, Baba K, Itoh T and Ashworth E N (2003a) Novel plasmodesmata association of dehydrin-like proteins in cold acclimated red-osier dogwood (Cornus sericea). Tree Physiol 23: 759–767. Karlson D T, Zeng Y, Stirm V E, Joly R J and Ashworth E N (2003b) Photoperiodic regulation of a 24-kD dehydrin-like protein in red-osier dogwood (Cornus sericea L.) in relation to freeze-tolerance. Plant Cell Physiol 44: 25–34. Karpinska B, Karlsson M, Schinkel H, Streller S, Süss K–H, Melzer M and Wingsle G (2001) A novel superoxide dismutase with a high isoelectric point in higher plants. Expression, regulation, and protein localization. Plant Physiol 126: 1668–1677. Karpinska B, Wingsle G and Karpinski S (2000) Antagonistic effects of hydrogen peroxide and glutathione on acclimation to excess excitation energy in Arabidopsis. IUBMB Life 50: 21–26. Kaur N and Gupta A (2005) Signal transduction pathways under abiotic stresses in plants. Curr Sci 88: 1771-80. Kavi Kishore P B, Sangam S, Amrutha R N, Laxmi P S, Naidu K R, Rao K R S S, Rao S, Reddy K J, Theriappan P and Sreenivasulu N (2005) Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Curr Sci 88: 424–438. Keles Yuksel and Oncel Isil (2002) Response of antioxidative defence system to temperature and water stress combinations in wheat seedlings. Plant Sci 163: 783-790. Khanna-Chopra Renu and Selote Devarshi S (2007) Acclimation to drought stress generates oxidative stress tolerance in drought-resistant than –susceptible wheat cultivar under field conditions. Environ Exp Bot 60: 276- 283. Koag M C, Fenton R D, Wilkens S and Close T J (2003). The binding of maize DHN1 to lipid vesicles. Gain of structure and lipid specificity. Plant Physiol 131: 309–316. Kobayashi F, Ishibashi M and Takumi S (2008a) Transcriptional activation of Cor/Lea genes and increase in abiotic stress tolerance through expression of a wheat DREB2 homolog in transgenic tobacco. Transgenic Res 17: 755-67. Kobayashi F, Maeta E, Terashima A, Kawaura K, Ogihara Y and Takumi S (2008b) Development of abiotic stress tolerance via bZip-type transcription factor LIP19 in common wheat. J Exp Bot 59: 891-905. 113 Kobayashi F, Takumi S, Egawa C, Ishibashi M and Nakamura C (2006) Expression patterns of the low temperature responsive genes in a dominant ABA-less-sensitive mutant of common wheat. Physiol Plant 127: 612-23. Kobayashi F, Takumi S, Nakata M, Ohno R, Nakamura T and Nakamura C (2004) Comparative study of the expression profiles of the Cor/Lea gene family in two wheat cultivars with contrasting levels of freezing. Physiol Plant 120: 585-94. Köhler B, Hills A and Blatt M R (2003) Control of guard cell ion channels by hydrogen peroxide and abscisic acid indicates their action through alternate signaling pathways. Plant Physiol 131: 385–388. Kreps J A, Wu Y, Chang H S, Zhu T, Wang X and Harper J F (2002) Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol 130: 2129–2141. Kukreja S, Nandval A S, Kumar N, Sharma S K and Sharma P K (2005) Plant water status, H2O2 scavenging enzymes, ethylene evolution and membrane integrity of Cicer arietinum roots as affected by salinity. Biol Plant 49: 305-308. Kurahashi Y, Terashine A, Takumi S (2009) Variation in dehydration tolerance, ABA sensitivity and related gene expression patterns in D-Genome progenitor and synthetic hexaploid wheat lines. Int J Mol Sci 10: 2735-51. Kwak J M, Mori I C, Pei Z M, Leonhardt N, Torres M A, Dangl J L, Bloom R E, Bodde S, Jones J D G and Schroeder J I (2003) NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J 22: 2623–2633. Labhilili Mustapha, Joudrier Philippe and Gautier Marie- Francoise (1995) Characterization of cDNA encoding Triticum durum dehydrins and their expression patterns in cultivars that differ in drought tolerance. Plant Sci 112: 219-230. Lascano Hernan R, Antonicelli Gerardo E, Luna Celina M, Melchiorre Mariana N, Gomez Leonardo D, Racca Roberto W, Trippi Victorio S and Casano Leonardo M (2001) Antioxidant system response of different wheat cultivars under drought: field and in vitro studies. Aust J Plant Physiol 28:1095-1102. Leonhardt N, Kwak J M, Robert N, Waner D, Leonhardt G, Schroeder J I (2004) Microarray expression analyses of Arabidopsis guard cells and isolation of a recessive abscisic acid hypersensitive protein phosphatase 2C mutant. Plant Cell 16: 596–615. Lin C C and Kao C H (2001) Abscisic acid induced changes in cell wall peroxidase activity and hydrogen peroxide level in roots of rice seedlings. Plant Sci 160: 323-329. Loggini Barbara, Scartazza Andrea, Brugnoli Enrico and Navari-Izzo Flavia (1999) Antioxidative defense system pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physiol 119:1091-1099. 114 Lowry O H, Rosebrough N J, Farr A L and Randall R J (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193: 265-75. Luna Celina L, Pastori M Gabriela , Driscoll Simon, Groten Karin, Bernard Stephanie and Foyer H Christine (2004) Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J Exp Bot 56: 411. Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y and Christmann A (2009) Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 324: 1064–1068. Macrae T H (2005) Diapause: diverse states of developmental and metabolic arrest. J Biol Res 3: 3–14. MacRobbie E A C (2000) ABA activates multiple Ca2+ fluxes in stomatal and guard cells, triggering vacuolar K+ (Rb+) release. Proc Natl Acad Sci USA 97: 12361–12368. Mahalingam R and Federoff N (2003) Stress response, cell death and signaling: the many faces of reactive oxygen species. Physiol Plant 119: 56–68. Manfre A J, Lanni L M and MarcotteW R Jr (2006) The Arabidopsis group 1 late embryogenesis abundant protein ATEM6 is required for normal seed development. Plant Physiol 140: 140–149. Melcher K, Ng L M, Zhou X E, Soon F F, Xu Y and Suino-Powell K M (2009) A gate-latchlock mechanism for hormone signaling by abscisic acid receptors. Nature 462: 602– 608. Millar A H, Mittova V and Kiddle G (2003) Control of ascorbate synthesis by respiration and its implications for stress responses. Plant Physiol 133: 443-447. Milone M T, Sgherri C, Clijters H and Navari-Izzo F (2003) Antioxidative responses of wheat treated with realistic concentrations of cadmium Env Exp Bot. 50: 265-273. Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7: 405410. Møller I M (2001) Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu Rev Plant Physiol 52: 561–591. Mouillon J M, Gustafsson P and Harryson P (2006) Structural investigation of disordered stress proteins. Comparision of full length dehydrins with isolated peptides of their conserved segments. Plant Physiol 141: 638-650. Munns Rana and Tester Mark (2008) Mechanism of salinity tolerance. Annu Rev Plant Biol 59: 651-681. Nayyar H and Walia D P (2003) Water stress induced proline accumulation in contrasting wheat genotypes as affected by calcium and abscisic acid. Biol Plant 46: 275–279. Neill S, Desikan R and Hancock J (2002a) Hydrogen peroxide signalling. Curr Opin Plant Biol 5: 388–395. 115 Neill S, Desikan R, Clarke A, Hurst R D and Hancock J (2002b) Hydrogen peroxide and nitric oxide as signalling molecules in plants. J Exp Bot 53: 1237–1247. Nobuhiko Arakawa, Tsutsumi Kimiko, Sanceda G Norlita, Kurata Tadao and Inagaki Choten (1981) A rapid and sensitive method for the determination of Ascorbic acid using 4,7Diphenyl-1,10-phenanthroline. Agric Biol Chem 45 (5): 1289-1290. Nylander M, Svensson J, Palva E T and Welin B V (2001) Stress-induced accumulation and tissue-specific localization of dehydrins in Arabidopsis thaliana. Plant Mol Biol 45: 263–279. Ohkuma K, Lyon J L, Addicott F T and Smith O E (1963) Abscisin II, an abscissionaccelerating substance from young cotton fruit. Science 142: 1592–93. Ohno Y, Ooura C, Rahman A, Aspuria E T, Hayashi K, Tanaka A and Uchimiya H (2003) pclorophenoxyisobutyric acid impairs auxin response in Arabidopsis root. Plant Physiol 133: 1135–1147. Okuma E, Soeda K, Tada M and Murata Y (2000) Exogenous proline mitigates the inhibition of growth of Nicotiana tabacum cultured cells under saline conditions. Soil Sci Plant Nutr 46: 257–263. Ozturk L and Demir Y (2002) In vivo and in vitro protective role of proline. Plant Growth Regul 38: 259–264. Ozturk Z N, Talame V, Deyholos M, Michalowski C B, Galbraith D W, Gozukirmizi N, Tuberosa R and Bohnert H J (2002) Monitoring large scale changes in transcript abundance in drought- and salt-stressed barley. Plant Mol Biol 48: 551–573. Park S Y, Fung P, Nishimura N, Jensen D R, Fujii H and Zhao Y (2009) Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 324: 1068–1071. Passiosure J B, Condon A G and Richard R A (1993) Water deficits, the development of leaf area crop productivity. In‘Water deficits’ (Eds JAC Smith and H Griffiths). Bios Scientific Publishers: Oxford: 253-263. Pei Z M, Murata Y, Benning G, Thomine S, Klusener B, Allen G J, Grill E and Schroeder J I (2000) Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 406: 731–734. Puhakainen T, Li C, Boije-Malm M, Kangasjarvi J, Heino P and Palva E T (2004b) Short-day potentiation of low temperature-induced gene expression of a C-repeat-binding factor-controlled gene during cold acclimation in silver birch. Plant Physiol 136: 4299–4307. Qin F, Kakimoto M, Sakuma Y, Maruyama K, Osakabe Y, Phan Tran L-S, Shinozaki K and Yamaguchi-Shinozaki K (2007) Regulation and functional analysis of ZmDREB2A 116 in response to drought and heat stresses in Zea mays L. Plant J 50: 54–69. Radotic K, Ducic T and Mutavdzic D (2000) Changes in perxidase activity and isoenzymes in spruce needles after exposure to different concentrations of cadmium. Env Exp Bot 44: 105-113. Ramanjulu S and Bartels D (2002) Drought- and desiccation- induced modulation of gene expression in plants. Plant Cell Environ 25: 141-151. Reddy A R, Raghavendra A S, Photooxidative stress in: K V Madhava Rao, A S Raghavendra and K J Reddy (Eds.) (2006). Physiology and Molecular Biology of Stress Tolerance in Plants. Springer, The Netherlands 157-186. Rhoads D M, Umbach A L, Subbaiah C C and Siedow J N (2006) Mitochondrial reactive oxygen species. Contribution to oxidative stress and interorganellar signaling. Plant Physiol 141: 357–366. Rio L A del, Sandalio L M and Corpas F J (2006) Reactive oxygen species and reactive nitrogen species in peroxisomes. Production, scavenging, and role in cell signaling. Plant Physiol 141: 330-335. Rio L A del, Sandalio L M, Altomare D A and Zilinskas B A (2003) Mitochondrial and peroxisomal magnese superoxide dismutase: differential expression during leaf senescence. J Exp Bot 54: 923-933. Rodriguez M, Canales E and Borras-Hidalgo O (2005) Molecular aspects of abiotic stress in plants. Biotechnologia Aplicada. 22: 1-10. Roe J and Oesterling M (1943) The determination of dehydroascorbic acid and ascorbic acid in plant tissues by the 2,4-dinitrophenylhydrazine. J Biol Chem 152: 511-21. Romero-Puertas M C, Corpas F J, Sandalio L M, Leterrier M, Rodriguez- Serrano M, Rio L A del and Palma J M (2006) Glutathione reductase from pea leaves: response to abiotic stress and characterization of the peroxisomal isozyme. New Phytol 170: 43-52. Rorat T, Grygorowicz W J, Irzykowski W and Rey P (2004) Expression of KS type dehydrins is primarily regulated by factors related to organ type and leaf developmental stage during vegetative growth. Planta 218: 878–885. Sairam R K and Srivastva G C (2001) Water stress tolerance of wheat (Triticum aestivum L.) variations in hydrogen peroxide accumulation and antioxidant activity in tolerant and susceptible genotypes. J Agron Crop Sci 186: 63-70. Sairam R K, Srivastava G C, Agarwal S and Meena R C (2005). Differences in antioxidant activity salt tolerance and reduced lipid peroxidation. J Exp Bot 364: 2207–2211. Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M, Shinozaki K and Yamaguchi-Shinozaki K (2006a) Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 18: 1292–1309. 117 Satoh R, Nakashima K, Seki M, Shinozaki K and Yamaguchi-Shinozaki K (2002) ACTCAT, a novel cis-acting element for proline- and hypoosmolarity-responsive expression of the ProDH gene encoding proline dehydrogenase in Arabidopsis. Plant Physiol 130: 709–719. Schwartz S H, Tan B C, McCarty D R, Welch W and Zeevaart J A (2003) Substrate specificity and kinetics for VP14, a carotenoid cleavage dioxygenase in the ABA biosynthetic pathway. Biochim Biophys Acta 1619: 9–14. Seki M, Ishida J, Narusaka M, Fujita M, Nanjo T, Umezawa T, Kamiya A, Nakajima M, Enju A and Sakurai T (2002b) Monitoring the expression pattern of around 7000 Arabidopsis genes under ABA treatments using a full-length cDNA microarray. Funct Integr Genomics 2: 282–291. Seki M, Narusaka M, Ishida J, Nanjo T, Fujita M, Oono Y, Kamiya A, Nakajima M, Enju A and Sakurai T (2002a) Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray. Plant J 31: 279–292. Selote D S and Khanna-Chopra R (2006) Drought acclimation confers oxidative stress tolerance by inducing co-ordinated antioxidant defense at cellular and subcellular level in leaves of wheat seedlings. Physiol Plant 127: 494-506. Selote Devarshi S and Khanna-Chopra Renu (2010) Antioxidant response of wheat roots to drought acclimation. Protoplasma 245:153-163. Seo M, Peeters A J M., Koiwai H, Oritani T, Marion-Poll A, Zeevaart J A D , Koornneef M, Kamiya Y and Koshiba T (2000). The Arabidopsis aldehyde oxidase 3 (AAO3) gene product catalyzes the final step in abscisic acid biosynthesis in leaves. Proc Natl Acad Sci U S A 97: 129-142. Serraj R and Sinclair T R (2002) Osmolyte accumulation: can it really help increase crop yield under drought conditions? Plant Cell Environ 25: 333–34. Shalata A and Neumann P M (2001) Exogenous ascorbic acid (vitamin c) increases resistance to salt tolerance and reduced lipid peroxidation. J Exp Bot 364: 2207–2211. Shao Hong-Bo, Chu Li-Ye, Cheruth Abdul Jaleel and Chang-Xing Zhao (2008a) Waterdeficit stress-induced anatomical changes in higher plants. Science Direct 331: 215225. Shao Hong-Bo, Chu Li-Ye, Shao Ming-An, Cheruth Abdul Jaleel and Mi Hong-mei (2008b) Higher plant antioxidants and redox signaling under environmental stresses. C R Biologies 331: 433-441. Shao Hong-Bo, Chu Li-Ye, Wu Gang, Zhang Jin-Heng, Lu Zhao-Hua and Hu Ya-Chen (2007) Changes of some anti-oxidative physiological indices under soil water deficits 118 among 10 wheat (Triticum aestivum L.) genotypes at tillering stage. Colloid Surface B 54: 143-149. Shao Hong-Bo, Liang Zong-Suo and Shao Ming-An (2005) Changes of anti-oxidative enzymes and MDA content under soil water deficits among 10 wheat (Triticum aestivum L.) genotypes at maturation stage. Colloid Surface B 45: 7-13. Shaoyun Lu, Wei Su, Haihang Li and Zhenfei Go (2009) Abscisic acid improves drought tolerance of triploid Bermuda grass and involves H2O2 – and NO- induced antioxidant enzyme activities. Plant Physiol Biochem 47: 132-138. Sharma P and Dubey R S (2005) Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: role of osmolytes as enzyme protectant. J Plant Physiol 162: 854-864. Sharma S and Verslues P E (2010) Mechanisms independent of abscisic acid (ABA) or proline feedback have a predominant role in transcriptional regulation of proline metabolism during low water potential and stress recovery. Plant Cell Environ 33 (11): 1838-51. Sharp R E and LeNoble M E (2002) ABA, ethylene and the control of shoot and root growth under water stress. J Exp Bot 53: 33–37. Shen Y G, Zhang W K, He S J, Zhang JS, Liu Q and Chen S Y (2003) An EREBP/AP2-type protein in Triticum aestivum was a DRE-binding transcription factor induced by cold, dehydration and ABA stress. Theor Appl Genet 106: 923–930. Shih M, Lin S, Hsieh J, Tsou C, Chow T, Lin T and Hsing Y (2004) Gene cloning and characterization of a soybean (Glycine max L.) LEA protein, GmPM16. Plant Mol Biol 56: 689–703. Shinozaki K and Yamaguchi-Shinozaki K (2000) Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Curr Opin Plant Biol 3: 217–223. Simova – Stilova Lyudmila, Demirevska Klimentina, Petrova Tatyana, Tsenov Nikolay and Feller Urs (2009) Antioxidative protection and proteolytic activity in tolerant and sensitive wheat (Triticum aestivum L.) varieties subjected to long-term field drought. Plant Growth Regul 58: 107-117. Simova-Stilova L, Vaseva I, Grigorova B, Demirevska K and Feller U (2010) Proteolytic activity and cysteine protease expression in wheat leaves under severe soil drought and recovery. Plant Physiol Biochem 48: 200-206. Simova-Stilova, Demirevska K, Petrova T, Tsenov N and Feller U (2008) Antioxidative protection in wheat varieties under severe recoverable drought at seedling stage. Plant Soil Environ 54 (12): 529-536. 119 Smirnoff N (2000) Ascorbic acid: metabolism and functions of a multi-facetted molecule. Curr Opin Plant Biol 3: 229–235. Soulages J L, Kim K, Walters C and Cushman J C (2002) Temperature-induced extended helix/random coil transitions in a group 1 late embryogenesis abundant protein from soybean. Plant Physiol 128: 822–832. Spollen W G, LeNoble M E, Sammuels T D, Bernstein N and Sharp R E (2000) Abscisic acid accumulation maintains maize primary root elongation at low water potentials by restricting ethylene production. Plant Physiol 122: 967–976. Su L, Zhao C, Ping Y, Wan S, Xia Z and Wang X (2011) Isolation and expression analysis of LEA genes in peanut (Arachis hypogea L.). J Biosci 36 (2): 223-228. Tai H H, Tai G C and Beardmore T (2005) Dynamic histone acetylation of late embryonic genes during seed germination. Plant Mol Biol 59: 909–925. Tan J, Zhao H, Hong J, Han Y Li H and Zhao W (2008) Effects of exogenous nitric oxide on photosynthesis, antioxidant capacity and proline accumulation in wheat seedlings subjected to osmotic stress. World J Agric Sci 4(3): 307-13. Tolleter D, Jaquinod M, Mangavel C, Passirani C, Saulnier P and Manon S (2007) Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell 19: 1580–1589. Tompa P, Szasz C and Buday L (2004) The role of structural disorder in the function of RNA and protein chaperon. FASEB J 18: 1169-75. Triantaphylides C, Krischke M, Hoeberichts F A, Ksas B, Gresser G, Havaux M, Breusegem Van and Mueller M J (2008) Singlet oxygen is the major reactive oxygen species involved in photo-oxidative damage to plants. Plant Physiol 148: 960-968. Tunnacliffe A and Wise J Michael (2007) The continuing conundrum of the LEA proteins. Naturwissenschaften 94:791-812. Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K and Yamaguchi- Shinozaki K (2000) Arabidopsis basic leucine zipper transcription factors involved in an abscisic aciddependent signal transduction pathway under drought and high salinity conditions. Proc Natl Acad Sci USA 97: 11632–11637. Veljovic-Jovanovic S, Noctor G and Foyer C H (2002) Are leaf hydrogen peroxide concentrations commonly overestimated? The potential influence of artefactual interference by tissue phenolics and ascorbate. Plant Physiol Biochem 40: 501–507. Verslues P E and Zhu J K (2004) Before and beyond ABA: upstream sensing and internal signals that determine ABA accumulation and response under abiotic stress. Biochem Soc Trans 33: 375-379. 120 Vysotskaya Lidiya B, Korobova Alla V and Kudoyarova Guzel R (2008) Abscisic acid accumulation in the roots of nutrient-limited plants: its impact on the differential growth of roots and shoots. Science Direct 165:1274-1279. Wang C Q and R C Li (2008) Enhancement of superoxide dismutase activity in the leaves of white clover (Trifolium repens L.) in response to polyethylene glycol-induced water stress. Acta Physiol Plant 30: 841-847. Wang Wen-Bin, Kim Yun-Hee, Lee Haeng-Soon, Kim Ki-Yong, Deng Xi-Ping and Kwak Sang-Soo (2009) Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol 47: 570-577. Wingler A, Lea P J, Quick W P and Leegood R C (2000) Photores- piration: metabolic pathways and their role in stress protection. Phil Trans R Soc Lond B 355: 1517– 1529. Wise M J (2003) LEAping to conclusions: a computational reanalysis of late embryogenesis abundant proteins and their possible roles. BMC Bioinform 4: 52-68. Wise M J and Tunnacliffe A (2004) POPP the question: what do LEA proteins do? Trends in Plant Sci 9: 13–17. Wolkers W F, McCready S, Brandt W F, Lindsey G G and Hoekstra F A (2001) Isolation and characterization of a D-7 LEA protein from pollen that stabilizes glasses in vitro. Biochim Biophys Acta 1544: 196–206. Xiong L, Ishitani M, Lee H and Zhu J K (2001) The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cofactor sulfurase and modulates cold stress and osmotic stress responsive gene expression. Plant Cell 13: 2063–2083. Xue G P and Loveridge C W (2004) HvDRF1 is involved in abscisic acid-mediated gene regulation in barley and produces two forms of AP2 transcriptional activators, interacting preferably with a CT-rich element. Plant J 37: 326–339. Yamaguchi-Shinozaki K and Shinozaki K (2005) Organization of cis-acting regulatory elements in osmotic- and cold-stress responsive promoters. Trends Plant Sci 10: 88– 94. Yan H, Gang L Z, Zhao C Y and Guo W Y (2000) Effects of exogenous proline on the physiology of soyabean plantlets regenerated from embryos in vitro and on the ultra structure of their mitochondria under NaCl stress. Soybean Sci 19: 314–319. Yang Y, Han C, Liu Q, Lin B and Wang J (2008) Effect of drought and low light on growth and enzymatic antioxidant system of Picea asperata seedlings. Acta Physiol Plant 30: 433-440. Yu C-W, Murphy T M and Lin C-H (2003) Hydrogen peroxide-induced chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Funct Plant Biol 30: 955–963. 121 Zhang J X and Kirharn M B (1994) Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol 35: 785-791. Zhang W, Ruan J, Ho TD, You Y, Yu T and Quatrano R S (2005) Cis-regulatory element based targeted gene finding: genome-wide identification of abscisic acid- and abiotic stress-responsive genes in Arabidopsis thaliana. Bioinformatics 21: 3074-3081. Zhang X, Zhang L, Dong F, Gao J, Galbraith D W and Song C P (2001) Hydrogen peroxide is involved in abscisic acid-induced stomatal closure in Vicia faba. Plant Physiol 126: 1438–1448. Zhu J K (2001b) Cell signaling under salt, water and cold stresses. Curr Opin Plant Biol 4: 401–406. Zhu J K (2002) Salt and drought stress signal transduction in plants. Plant Mol Biol 53: 247– 273. Zlatev Z S, Lidon F C, Ramalho J C and Yordanov I T (2006) Comparison of resistance to drought of three bean cultivars. Biol Plant 50: 389-394. 122