Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
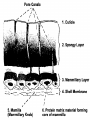
Parental Care IV: Incubation JodyLee Estrada Duek, Ph.D. With assistance from Dr. Gary Ritchison http://people.eku.edu/ritchisong/avianreproduction2.html Incubation • During incubation, birds transfer heat to eggs. Arizona-Sonora Desert Museum • For optimum development, egg temperature must be maintained at about 37 - 38 degrees C (Gill 1995). • Exposure to higher temperatures is lethal, while cooler temperatures will, at minimum, slow down or stop development. • Heat is transferred through brood, or incubation, patches - an area of bare, flaccid skin on the abdomen and/or breast. • Prior to the initiation of incubation, the skin in the area of the brood patch loses its feathers. • In addition, the dermis becomes spongy and richly supplied with blood vessels (Welty and Baptista 1988). Brood patch • Birds of nearly all species temporarily shed their feathers on single or paired areas of the breast or abdomen early in the breeding season. • The bare skin increases in vascularity, which aids it in transferring body heat for incubating the eggs and brooding the chicks. • Development of these incubation (brood) patches is prompted by rising levels of estrogen. • They form in whichever sex cares for the eggs and young, usually females but often males as well. • The lost feathers are replaced in the complete molt following the breeding season (Stettenheim 2000). Incubation reduces microbial growth on eggshells • -- Avian eggshells harbor microbes shortly after laying, and under appropriate ambient conditions they can multiply rapidly, penetrate through shell pores, infect egg contents and cause embryo mortality. • Cook et al. (2005) experimentally examined how incubation affects bacterial processes on the eggshells of Pearl-eyed Thrashers (Margarops fuscatus) nesting in tropical montane and lowland forests in Puerto Rico. • Bacteria and fungi grew rapidly on shells of newly laid, unincubated eggs exposed to ambient conditions, but declined to low levels on shells of eggs incubated by thrashers. • Divergence in bacterial growth between incubated and exposed eggs was more marked at the montane forest than at the lowland site. • Pathogenic microorganisms became increasingly dominant on shells of exposed eggs, but these groups were relatively rare on incubated eggs, where more benign, less invasive groups prevailed. • Some incubation during laying may be necessary to decrease the probability of trans-shell infection by reducing the growth of harmful bacteria and fungi on eggshells, although it may increase hatching asynchrony and the likelihood of brood reduction. Energetic cost of incubation • In birds, the annual peak of energy demand has long been thought to occur when parents provision their offspring with food during the nestling phase. • This led to idea selection on clutch size takes place during nestling phase. • energetic demands during other reproductive phases –egg laying and incubation – have been ignored. • During incubation, avian eggs need external heat provisioning, regular turning and favourable humidity for proper embryonic development; care provided by one or both of parents. • energetic costs of providing heat to the eggs thought to be negligible. • Increasing evidence that below thermo-neutrality the metabolic rate (energy spent per time unit) of an incubating female is higher than that of a nonincubating female at rest. • Because temperatures are normally below thermoneutrality at temperate latitudes, energetic costs of incubation may substantially add to overall daily energy expenditure of attending parents. Energetic costs of incubation • A nest-box modified into a metabolic chamber. • To ensure the top of the nest-box was airtight, a sheet of rubber was inserted between nest-box and lid (a) and a cork was placed in the entrance hole (b). • Reference air was measured close to the inflow of the nest-box (arrows underneath nest-box; c), while sample air was drawn from the nest-box via a tube near the entrance hole (d). • The thickness of the nest was determined by the thickness of the nest cup (e) and the height of the nest rim (f). Clutch size and incubation energetics • de Heij et al. (2007) manipulated the clutch sizes of female Great Tits (Parus major) and monitored their metabolic rates during nocturnal incubation using mobile oxygen analyzers. • found clutch enlargement caused incubating females to expend more energy, but clutch reduction did not lower energy expenditure. • absence of an effect of clutch reduction can be explained by a limit to number of eggs in direct contact with brood patch: threshold clutch size • Above a threshold clutch size, eggs that are not in contact with the female's brood patch cool. • incubating birds will repeatedly rearrange eggs to rewarm • Rewarming energetically more costly than maintaining eggs at incubation temperatures. • energetic costs increase when clutch size is above the threshold clutch size, but do not change when clutch size is at or below threshold clutch size. Nest insulation • de Heij et al. (2007) also found that incubating birds with thicker nests had lower energy expenditure, probably because thicker nests were better insulated. • The fact that not all birds build well-insulated nests suggests there is a cost to thick nests. • Knowing females expend more energy during nocturnal incubation when incubating experimentally enlarged clutches is a first step towards determining a potential mechanism underlying negative selection on clutch size during the incubation. • measurements on energy expenditure over a full 24 h are needed to judge how important energy expenditure can be in explaining fitness consequences of incubating experimentally enlarged clutches. Incubation • Most birds sit on their eggs to incubate, but there are exceptions. • For example, male Emperor (D. Attenborough clip) and King penguins place their egg on their feet, wrap their wing around the egg, and incubate it while standing up. • Male emperor penguins http://www.youtube.com/watch?v=6AiCIZ9wM1o • The egg is kept warm by the heat from the male's feet & wing. Source: www.mbr-pwrc.usgs.gov/id/framlst/Photo/p1142.html Foot-Mediated Incubation: Nazca Booby (Sula granti) Feet as Surrogate Brood Patches •Incubation in most birds involves transferring heat from parent to egg through a highly vascularized brood patch. •Some birds, however, do not develop a brood patch. • hold their eggs under the webs of their feet • webs are often positioned between the feathered abdomen and the egg during incubation, suggesting that either the abdomen, the feet, or both could transfer heat to the egg. • Morgan et al. (2003) studied heat transfer from foot webs to eggs in Nazca boobies by spatially separating the feet from the abdomen using an oversized egg. • found feet transfer heat to eggs independently of any heat from the abdomen. • found that incubating boobies had significantly greater vascularization in foot webs, measured as percentage of web area covered by vessels • males, whether incubating or nonincubating, had significantly less vascularization • vascularized Nazca booby feet function during incubation as vascularized brood patches Micronesian megapode Megapodes • They are the only known birds which use heat sources, other than the body, to incubate their eggs. • When the young birds hatch they are fully able to defend for themselves and receive no parental care. • Megapodes are quite heavy-bodied birds and forage on the forest floor, where they search for insects, seeds and fruit. • All members of the Megapode family can fly, but most move around primarily by walking. Megapode mound •found in Southeast Asia and Australia, •use heat sources •geothermal, •solar, or •decomposition of organic material. Maleefowl 1 • Maleefowl mound, Australia Malleefowl 2 • They spend up to eleven months of the year preparing, then maintaining, the mound adapting to daily, as well as seasonal, variations in temperature and rainfall. • mounds, made largely of sand raked up by their powerful feet, are each an immaculate circle approximately 4.5 metres in diameter. • This labour is to establish an exact and constant 33oC - measured by its heat-sensitive beak - in the central chamber of the mound. • This chamber is literally carved out of consolidated layers of vegetation, like compressed paper mixed with sand. • All the eggs will be laid in this chamber - up to thirty at intervals of two to five days apart. • The key elements in incubation are solar heat and fermentation. • malleefowl make the most of rainfall and rely on daily raking of the mound to harness appropriate levels of solar energy. Malleefowl 3 • In winter, when solar energy is minimal, they scratch together dead leaves, twigs, sparse humus and rake them into a path that leads directly into the now volcano-shaped centre of the mound. • It is not an uncommon sight to see a path of sticks and leaves, perhaps half a metre across and 20 metres long, leading from the bush up and over the side of the mound into its centre. • When wet, all the collected vegetation is sealed within the mound and buried to rot and ferment. • This process is repeated again and again and may take four months. • As the season warms and eggs are laid and buried, the malleefowl begin to uncover the mound, altering its architecture to reinforce the slow fermentation with the heat of the sun. • This process increases during the summer months until the mound itself is shaped like a pointed cone and the incubation process becomes dependent on solar energy alone. Malleefowl 4 (Leipoa ocellata) • dedicate 9-11 months per year building and maintaining a large incubation mound of soil, leaves and twigs. • The eggs are laid in the mound, buried and left to incubate by heat generated from the composting litter. • Malleefowl mounds may be used over many generations and can attain an impressive size of 22 meters in circumference and 1 meter high. • The birds maintain the mound temperature of 32-34 degrees C by using their beak as a "thermometer" and adjusting soil cover to either retain or expel heat from the egg chamber. 'Incubation' typically takes 60 - 90 days. (Source:http://www.malleefowl.com.au/Pages/TheMalleefowl.htm) Incubation periods • range from about 10 days for some passerines & woodpeckers to as many as 80 days for albatross' and kiwis. • time spent incubating is related to the size of the egg, state of development (precocial vs. altricial), & ambient temperature. Temperatures in an Eastern Bluebird nest cup in Texas (Cooper and Phillips 2002). Nest predation appears to affect parental behavior 1 • Based on an analysis of incubation and provisioning behavior of 97 species of passerines, Conway and Martin (2000) suggested environments with high risk of nest predation favor long on-bouts (long periods on nest) and few foraging trips. • This strategy may prevent frequent feeding by adults and thus compromise future reproductive attempts. • nest predation may influence evolution of avian life-history traits in several ways. • High nest predation favors a strategy of • holding back reproductive effort for renesting attempts and survival, • a short nesting cycle to minimize the time nests are susceptible to predation, • small brood size to minimize noise of begging young Nest predation appears to affect parental behavior 2 • Conway and Martin (2000) suggest nest predation may influence passerines by placing constraints on parental activity and the way an incubating female allocates her time between incubation and foraging. • with high nest predation, natural selection simultaneously favors infrequent nest trips (to reduce the probability of predator detection) and short off-bout duration (to maximize development rates and reduce time of exposure to predators). • These somewhat opposing constraints limit the range of effective incubation strategies available to females in environments with high nest predation. Males Feeding Females during Incubation 1 • See graph on following slide of nest attentiveness (percentage of time that the female is incubating on the nest) relative to the rate that males bring food to the nest (incubation feeding). • The relationship across 19 species of both open- and cavity-nesters is curvilinear and significant. • Nest attentiveness (percentage of time spent on the nest) during incubation represents a parent-offspring conflict; incubating birds must trade-off between caring for embryos by staying on the nest versus caring for themselves by getting off the nest to forage. • For species in which females are the sole incubator, males can potentially affect this trade-off and increase nest attentiveness by feeding incubating females on the nest (incubation feeding). • (From: Martin and Ghalambor 1999). Males Feeding Females during Incubation 2: Theory (Martin and Ghalambor 1999) • Increased nest attentiveness may be required when local microclimate is harsh and requires more incubation feeding (microclimate hypothesis). • Incubation feeding may be constrained by risk of attracting nest predators (nest predation hypothesis), which in turn may constrain female nest attentiveness because of energy limitation. • incubation feeding rates are greater among cavity-nesting than open-nesting birds. • Under microclimate hypothesis, the greater incubation feeding rates of cavitynesting birds generate the prediction that microclimate should be harsher than for open-nesting birds. • results reject this hypothesis because we found the opposite pattern; cavitynesting birds experienced more moderate (less variable) microclimates that were less often below temperatures (i.e., 16°C) that can negatively impact eggs compared with open-nesting species. • In contrast, incubation feeding rates were highly negatively correlated with nest predation both within and between the two nest types, supporting the nest predation hypothesis. • Incubation feeding in turn was positively correlated with nest attentiveness. • Thus, nest predation may indirectly affect female incubation behavior by directly affecting incubation feeding by the male (Check this short video of a male Blue Tit feeding his mate). Food availability and nest attentiveness across species and latitudes Chalfoun and Martin (2007) • Both northern and southern species are expected to show proximate increases in attentiveness in response to increased food availability, • preliminary data further suggest that northern species show slightly stronger responses. • Proximate responses to food availability alone, however, cannot explain why southern species generally show lower nest attentiveness than similar northern species (grey arrows). Latitudinal variation in avian incubation attentiveness 1 • Avian incubation attentiveness has important fitness consequences: • • • number of young quality of hatched young energetic costs imposed on parents. • Nest attentiveness is highly variable across species and geographical regions. • Chalfoun and Martin (2007) reviewed the literature and found a worldwide pattern that nest attentiveness of passerines is generally lower in south temperate and tropical regions than in north temperate regions. • conducted a food manipulation experiment to assess nest attentiveness: does it reflect proximate responses or an evolved behaviour. • Karoo Prinia (Prinia maculosa) in South Africa has very low nest attentiveness (about 49%) compared with many passerines. • provided supplemental food during early incubation to experimental females and compared nest attentiveness and on- and off-bout lengths • Nest attentiveness of females at food-provisioned nests was significantly higher than control females (57% vs. 49%). • Food-supplemented females spent significantly less time off nest than did control females; mean on-bout lengths did not differ. Latitudinal variation in avian incubation attentiveness 2 • mean nest attentiveness of food-provisioned females still substantially below similar species worldwide. • Food can be an important proximate influence on parental care behaviour, but proximate influences of food do not explain latitudinal patterns of attentiveness. • Climatic variation across latitudes may influence the amount of time that parents spend on the nest, although temperatures at many south temperate sites often approximate those at north temperate sites during the breeding season. • One possible alternative explanation for geographical patterns in nest attentiveness is variation in adult mortality across latitudes. • According to classic life history theory, if southern birds experience lower adult mortality, they should be less willing to invest as much in nest attentiveness and other components of current reproduction. • Testing for the existence of an adult mortality–nest attentiveness trade-off across latitudes is therefore a critical next step in addressing geographical variation in parental care strategies. Winter habitat influences reproductive success 1 • destruction of tropical forests is creating breeding problems for migratory birds. • Norris et al. (2004) found quality of winter habitat affected ability of American Redstarts to successfully reproduce when they returned north in the spring. • Norris,noted that a relatively small geographic band – across the Caribbean, Greater Antilles, and central America – is the annual destination of an estimated five billion migratory birds flying south each year from Canada. • Norris et al. (2004) measured stable carbon isotopes in blood samples collected from American Redstarts after the birds arrived at their breeding grounds in Ontario. • Since the turnover time of the blood cells is from six to eight weeks, they provide a good indicator of the quality of the birds' previous habitat on the wintering ground. • The carbon signature of each redstart has been deposited by insects the bird ate, and the insects in turn fed on vegetation growing in the winter habitat. • It’s a 'food chain signature' Winter habitat influences reproductive success 2 • Norris et al. (2004) first determined high and low quality habitats for redstarts, which winter in the Caribbean and central America, and breed in deciduous forest throughout Canada and the U.S. • what makes a better winter habitat is primarily the degree of wetness, particularly at the end of the season (late winter/early spring) when low quality habitats tend to dry out. • The second step was to develop isotope "markers" that identify habitat quality for each type of habitat. • Third, blood samples were taken from warblers in their breeding grounds • Fourth, the birds' reproductive success was measured by counting the number of "fledged" offspring to leave their nests. • Analysis revealed that redstarts wintering in high quality habitats, such as mangroves and lowland tropical forests, arrived earlier on the breeding grounds, nested earlier, and were more successful in producing young. • This study shows that destroying high quality winter habitat has a disproportionate effect on the redstart populations: they lose the areas most capable of supporting them. Eggshell • protects the embryo • generally white in cavity-nesters & colored and patterned in open nesters (for camouflage) • color is added to the eggshell from pigments secreted by cells in the wall of the uterus • contains thousands of pores (see diagram) that permit gas exchange Thousands of tiny pores like the ones pictured, cover the shell, providing a passage for gas exchange. (Source: http://www.rit.edu/~tld089 8/SEM.html) Eggshell pores • An egg must exchange gases with its environment • This allows for the removal of excess CO2 and the influx of “fresh air” with additional O2 as gases diffuse through pores in the eggshell • Water loss also occurs through these pores • Water loss leads to a larger and larger air chamber at the blunt end of the egg • Just before hatching the chick needs to inflate its lungs and breathe • This is accomplished as the chick breaks the membranes and has access to the air chamber at the blunt end of the egg • There is a fine line between • not having enough air in the chamber to allow the chick to breathe long enough to break a hole in the shell • losing too much water and the embryo dehydrating before maturity • The rate, and total amount, of water loss depends on • Ambient temperature • Humidity • Elevation and subsequent air pressure Egg water vapor conductance • AR AND RAHN and others at SUNY Buffalo measured the water vapor conductances of fresh eggs of 29 species • showed how this total value increases with egg weight. • change in egg shell thickness with egg weight also determined from the literature • measured periodically the weight loss under known conditions of temperature, humidity, and barometric pressure • rate of water loss per gram egg weight decreases as the eggs get larger; large eggs can save more water than small eggs. Incubation period and water loss • Both Needham (1931) and Romanoff and Romanoff (1949) noted that species with large eggs tend to have longer incubation periods and thus would be exposed longer to water losses. • This problem is discussed in detail by Rahn and Ar (1974) who showed that the extended incubation period in larger eggs is compensated for perfectIy by the reduction in water vapor conductance per unit egg weight which they express as a common water loss coefficient for all eggs. • Whether the increase in pore area with weight is achieved by increasing the number of pores or their diameter or some combination of the two is still being explored. Eggshell pores http://www.cooper.org/pdf/p1977Rahn1974.pdf • Gas exchange in the avian embryo has been shown to be dependent on, and limited by, the diffusive properties of gases across the resistance offered by the shell and shell membranes • The diffusive rate of water loss from eggs depends on: • permeability constant of the shell – Pore size – Number of pores per unit area • surface area of the shell ( cm2) • water vapor pressure difference across the eggshell Maculation in eggshells James P. Higham and Andrew G. Gosler 2006 Oecologia • • • • • • Many small passerine birds worldwide lay white eggs speckled with red, brown and black protoporphyrin pigment spots (maculation). Unlike some patterns of avian eggshell pigmentation which clearly serve a crypsis or signalling function, the ubiquity of maculation among passerines suggests that its origins lie in another function, not specific to any particular ecological or behavioural group. There is evidence that protoporphyrin pigments serve a structural function related to eggshell thickness and calcium availability: eggshell maculation in the great tit Parus major increases with decreasing soil calcium levels, pigments demarcate thinner areas of shell, and both the pigment intensity and distribution are related to shell thickness. maculation also affects the rate of water loss from the egg during incubation (≈ Mass Loss per Day or MLD, which is critical to egg viability), but not that of unincubated eggs. They demonstrate, both by observation and experiment, that the effect of female incubation behaviour on MLD compensates in some way for variation in egg characteristics, and that differences between females in the degree of such compensation are related to differences in clutch maculation. results suggest that, while a principal function of maculation in this species may be to strengthen the eggshell, it may also reduce eggshell permeability when large amounts of pigment are used, and that this necessitates a behavioural adjustment from the female during incubation. A special problem with mount nesters and gas exchange Condor, 1987 Elevation and water loss • Among species found at wide ranges altitudinally, there is a local variation in number of eggshell pores per unit area, OR in size of individual pores • Higher elevation results in more rapid diffusion; therefore fewer pores, or smaller pores, are needed • Birds that are moved altitudinally fairly quickly adjust pore numbers and/or pore sizes in the eggs that they lay Adaptation of the Avian Egg to High Altitude 1 CYNTHIA CAREY American Zoologist 1980 • Theoretical predictions and experiments on eggs of domesticated birds indicate that the diffusion coefficient of gases is inversely proportional to barometric pressure. • Therefore potentially lethal losses of CO2 and water vapor from eggs laid at high altitude might result if the increased tendency of gases to diffuse at reduced barometric pressure were not counteracted • data from two wild populations indicate water loss is independent of altitude over a 3000 m gradient. • Four different possibilities by which compensation for increased diffusion of water vapor might be achieved at high elevations – 1) a reduction in eggshell conductance (GH2O) – 2) an increase in the initial water content of the eggs – 3) an increase in shell thickness – 4) alteration of water vapor pressure in the nest microenvironment or incubation temperature by variation in parental behavior Adaptation of the Avian Egg to High Altitude 2 • Mean GH2O of eggs of two precocial and four altricial species breeding above 2800 m is significantly reduced below values of related birds breeding at lower elevations, but no change in initial water content or shell thickness has been observed in such eggs • That data contradicts hypotheses 2 and 3 • Observations of parental behavior in species breeding over wide elevational gradients have not yet been made (hypothesis 4) • More research needed for identification of the mechanisms – ways eggshell structure is modified to achieve a reduced GH2O – environmental cues used by females to determine elevation of nest location – the rapidity with which shell structure can be modified Elevation and water loss H. Rahn, T. Ledoux, C. V. Paganelli and A. H. Smith J Appl Physiol 53: 1429-1431, 1982; 8750-7587 • Hens acclimated to an altitude of 3,800 m (PB 480 Torr) were transferred to 1,200 m (PB 657 Torr). • Eggs were collected before departure and daily after the transfer so that changes in eggshell conductance could be studied. • Over the next 2 mo eggshell conductance increased 30%, presumably to compensate for the 37% reduction (from 657 to 480 Torr) in gas diffusivity at the lower altitude. • Measurements of shell thickness and number of pores in the shell allow one to calculate that most of the change in total pore area occurred by an increase in cross-sectional area of individual pores. Altitude and eggshells Monge-C. and F. León-Velarde (1993) • At the end of incubation, partial pressures of oxygen and carbon dioxide in the air cell of sea-level avian eggs are similar to those in the expiratory air of adult birds. • At high altitude, changes in the permeability of the shell and probably in embryo metabolism partially compensates the increase in the gas diffusion constant resulting from the low barometric pressure. • tested whether-despite of the adaptive responses of the high altitude avian embryo-the air cell values would be similar to those of the alveolar air of high altitude human natives. • Air cell O2 (48.3±1.6 torr) and CO2 (20.9±0.85 torr) pressure values were obtained by studying naturally incubated eggs of the Andean gull (Larus serranus) at 4650m. • Sea-level chicken (Gallus gallus) air cell pressure values of O2 (102.3±2.7 torr) and of CO2 (43.3±1.3 torr) were obtained from the literature for comparison. • values similar to alveolar air of humans at sea level (O2: 104.4±0.4 torr, CO2: 40.1± 0.25 torr) and at high altitude (4540 m) (O2:50.5±0.53 torr, CO2: 29.1±0.37 torr). • Despite large evolutionary changes in morphology and physiology of respiratory organs, head pressure of O2 that oxygenates the blood keeps a constant value in the pre-pipping avian embryo and in alveolar air of adult mammals. • This constancy holds valid at high altitude. Differences in egg size, shell thickness, pore density, pore diameter and water vapour conductance between first and second eggs of Snares Penguins Eudyptes robustus and their influence on hatching asynchrony MELANIE MASSARO, LLOYD S. DAVIS (2005) Ibis 147 (2) , 251–258 • Brood reduction in birds is frequently induced by hatching asynchrony. • Crested penguins (genus Eudyptes) are obligate brood reducers, but in contrast to most other birds, first-laid eggs are considerably smaller in size than second-laid eggs; furthermore, first-laid eggs hatch after their siblings • The mechanisms underlying this reversal in size and hatching order remain unclear. • tested whether the second-laid eggs of Snares Penguins have a higher eggshell porosity allowing them to maintain a higher metabolic rate throughout incubation and to hatch before their first-laid siblings. • investigated differences in egg size, shell thickness, pore density, pore diameter and water vapour conductance between first and second eggs within clutches and examined influence of shell characteristics on hatching asynchrony. • First-laid eggs were approximately 78% the size of larger second eggs. • Second-laid eggs had considerably thicker shells and more pores per cm2 than first eggs; pore diameter did not differ Differences in egg size, shell thickness, pore density, pore diameter and water vapour conductance between first and second eggs of Snares Penguins Eudyptes robustus and their influence on hatching asynchrony MELANIE MASSARO, LLOYD S. DAVIS (2005) Ibis 147 (2) , 251–258 • Water vapour conductance was greater in second- (16.8 mg/day/torr) than in firstlaid eggs (14.9 mg/day/torr). • The difference in water vapour conductance between first- and second-laid eggs within clutches was related to hatching patterns. • In nests where second eggs hatched before first-laid eggs, second eggs had a considerably greater water conductance than their sibling, whereas in nests where both eggs hatched on the same day, the difference in water conductance between eggs was very small, and in a few nests where small first eggs hatched before their larger sibling, they had a greater water conductance than their larger second-laid nestmate. • Surprisingly few studies have investigated differences in shell characteristics between eggs within clutches and associated effects on hatching asynchrony. • This study has demonstrated that such differences exist between eggs within clutches and that they can influence hatching patterns. EGG SIZE, EGGSHELL POROSITY, AND INCUBATION PERIOD IN THE MARINE BIRD FAMILY ALCIDAE Karen Zimmermann and J. Mark Hipfner The Auk • • • • • • • ultimate factors that influence duration of avian incubation are well known, we know much less about the proximate mechanisms by which birds adjust incubation period tested the hypothesis that an adjustment in eggshell porosity is one such proximate mechanism (i.e., that avian species with higher ratios of incubation period to egg size lay eggs with less porous shells). Eggshell porosity affects the rate of gaseous exchange between the developing embryo and the external environment; thus, to the extent that embryonic metabolism is diffusionlimited, eggshell porosity could directly determine incubation period. collected eggs from seven species of Alcidae, a family of marine birds that exhibits an unusual degree of interspecific variation in incubation period, and measured egg mass and eggshell porosity (determined by the number and size of pores and the thickness of the shell). Incubation periods were obtained from the literature. Egg mass and eggshell porosity combined explained 87% of variation in incubation period among the species, which included at least one member of each of six main alcid lineages. As predicted, eggshell porosity and incubation period were negatively related, after controlling for egg mass. results are consistent with the hypothesis that evolutionary changes in avian incubation period may be attributed, at least in part, to adjustments in eggshell porosity. Canada goose hatching • http://www.youtube.com/watch?v=TYJiQ-03tBI