Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
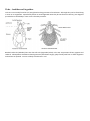
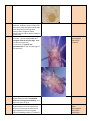
A Key to Soil Mites in the UK th Test version 1: 7 March 2014 Felicity Crotty, and Matthew Shepherd With thanks to David Harding for figures and some text. Introduction Mites (Sub-class Acari) are at first glance similar to spiders (Arachnida). However mites can be distinguished from spiders by their lack of a waist between the thorax (where the legs are attached) and the abdomen. This gives the characteristic appearance of a single oval body, with head and legs attached to it. This key should help you to distinguish the major groups of mites found in UK soil and litter. It aims to enable you to distinguish major groups of soil mites based on untreated, undissected specimens using a compound microscope with top and bottom lighting. Mite Ecology Mites and ticks are a heterogeneous collection of arachnids, including herbivorous, parasitic and saprophagous (detritus-feeding) species as well as predators. In fact there are almost no lifestyles to which mites have not become adapted, with some generating and living within plant galls, and some Demodex sp. even living on human foreheads! This diversity in ecology and behaviour is likely to be the reason for the vast diversity in morphological characteristics, with different groups adapted in different ways to their surrounding environments. Many species of mite change their ecological niches at different stages in their life, with parasitic nymphs, phoretic stages that hitch a lift on other animals, and active hunting adults. The diversity of species of mites within just one ecosystem has perplexed scientists for many years, with many wondering how so many different species can be living in the same space Habitats Mites can be found in all ecosystems globally, from soil in desert and arctic environments to grasslands and tropical rainforests. Mites can be found living in arboreal environments, within plant galls, parasitic on other invertebrates and vertebrates and some mites are even aquatic. This key concentrates on those mites that can be found in soil and litter habitats, however it should also work for mites inhabiting suspended soils in canopies. Feeding Soil mites are generally recognised to be mostly decomposers/microbivores or predators. Feeding mode, will affect the morphology of mouth parts, typically particulate feeders have large chelate chelicerae (e.g. Oribatid and Mesostigmatid mites), whilst for suctorial feeders mouthparts are more elongated (e.g. Prostigmatid mites). Most Mesostigmata are active, well-armoured predators, but some (Uropodidae) are slow-moving fungal feeders. Oribatids are considered to be decomposers, consuming fungi and bacteria on organic matter (mycophagy/microbivory), or the organic debris itself (detritivory/saprophagy). Prostigmatid mites have a more cosmopolitan feeding strategy, being a mixture of predators, fungivores or omnivores, however all known Prostigmata consume their food by fluid feeding. Breeding Mites breed both sexually and asexually (parthenogenetically) depending on species. Sexual reproduction can involve the transfer of sperm either directly or indirectly. Sperm transfer can be via the chelicera, or via the aedeagus directly to the female genital opening or even just by leaving a spermatophore (stalked sperm packet) in the environment to be picked up by a female later. Growth and Development Mites go through five instars between egg laying and maturity. These include a prelarva (sometimes prolarva) stage, larva, protonymph, deutonymph (or deuteronymph also sometimes hypopus depending on species), tritonymph, before becoming an adult. Before full development as an adult it is common for the mites to have a reduced number of legs, have a non-feeding stage of development, or use this stage for dispersal by phoresy. It is very difficult to identify a mite using this key if it is not an adult. Dispersal Mites are known to disperse actively (through movement across / within soils) and passively (via rafting, wind dispersal) as well as by phoresy (hitching a lift on other organisms). Through passive dispersal and phoresy they can be transported many km’s across habitats. The distance covered by active dispersal is species dependent – oribatid’s for example have been found to travel 0.3-2.1 cm per day although predatory Mesostigmata have been found to travel faster than this across habitats. Predators During the growth and developmental stages all mites go through to get to adulthood, all mites are initially at risk from predation, from their own species as well as other mites and the other large predators that reside in the soil (e.g. pseudoscorpions, centipedes and staphylinid beetles). However, for some species once they reach maturity they actually suffer from very little predation pressure, usually because they have invested in predator defence mechanisms like sclerotisation/armour, this is why sometimes it is suggested that Oribatids live in an enemy-free space. Trends and threats There is such a huge diversity of mite species, that to the beginner acarologist could seem quite a daunting task, however, everything is not everywhere! Some mites are specialists to certain habitats and there is still a large potential for biodiversity loss through habitat destruction – much like other faunal groups. We unfortunately are also suffering from a taxonomic impediment, where it is difficult to cultivate expertise in mite taxonomy and when it has been taught, it is very difficult to retain these experts within scientific and naturalist jobs, threatening our knowledge of this fascinating group. A Taxonomic Apology The taxonomic groups which this key can identify are not at the same taxonomic levels. This is both for practical reasons – resolving specimens to greater taxonomic detail may require specimens cleared in lactic acid, phase contrast, oil immersion, require both male and female specimens, or complicated approaches based on the locations of hairs (setae) or sensory organs. However, it also reflects the seemingly constant changes in mite taxonomy, with groups that were previously considered at the same level, now appearing at very different places in taxonomic diagrams. Increasing knowledge of the genetics of mites is gradually resolving taxonomy so that it’s no longer based on observation of features, which may be misleading, and becoming based on the pattern of evolutionary change, as evidenced by the clues left in each species genetic material. This should, we hope, enable the taxonomy to become more settled, longer lasting and valid in evolutionary terms. The “trunk” of this evolutionary tree is not so very complicated. Mites fit into the tree of life as part of the animal kingdom, in the ecdysozoa (animals that shed their exoskeletons), among the arthropod phylum (jointed limbs) and in the chelicerata (animals with pincer-like mouthparts) and the class Arachnida (spiders, scorpions, pseudoscorpions, ticks and mites). However, within the sub-class Acari things get more complex. There is a major division between Super-order Acariformes and Parastiformes, and these are further divided into six orders. Some orders only contain a single super-family, and so once you’ve identified the order, you’ve already got down to super-family level. However, most of the soil-dwelling orders have a variety of levels, some with sub-orders within them, these are split (in some cases) into super-cohorts, then cohorts, some with sub-cohorts, under which lie superfamilies, familes and (finally!) genera and species. Figures 1 and 2 provide schematics showing how the two superorders are subdivided, indicating which groups are unlikely to be found in the UK, and showing the taxonomic level to which this key can take you. These levels are used for general identification of mites for ecological monitoring purposes, and identification to these groups should help to separate groups playing different roles within the soil and litter ecosystem. If you’re really bitten by the acarology bug, following the keys in Dindal or “The Manual of Acarology”, may help, but these may require a more complicated process, access to both male and female specimens, and may require some dissection and clearing of specimens. Figure 1: Taxonomy of Acari in the super-order Parasitformes. Grey boxes are groups not normally found in the UK, and the blue box shows those not normally found in soil. Superfamilies are indicated by bold italic text, and are included under those groups for which there are no further subdivisions. These are named where there is only one superfamily, or the number of superfamilies given. The most detailed taxonomic levels which this key can identify are identified in red text. Figure 2: Taxonomy of Acari in the super-order Acariformes, following the system used in figure 1 above. Mite Anatomy Where possible, terms in common usage have been used, but it’s worth getting to know some specific terms. The basic division of the acarine body is into two sections: the gnathosoma (jaw-body or “head”) and the idiosoma (distinct-body or main body) although depending on group it can be referred to differently (Figure 3). The gnathosoma bears mouthparts comprising a pair of chelicerae (often pincer like) and paired palps (or pedipalps) which are leg-like appendages either side of the mouthparts. The idiosoma bears four pairs of legs in the vast majority of species, although young mites often start with only three pairs, whilst gall mites have only two pairs. Legs and palps are divided into segments which are named from their tip (distal end - furthest from the body), which may also bear another structure (claws, hooks, pads or projections) called an apotele, or setae (hairs) which often act as sensory organs. Counting back from the distal end, the leg segments are: Tarsus – Tibia – Genu – Femur – Trochanter – Coxa Where segments have been lost through evolutionary change, the naming runs from the distal end (tip), thus a 4segmented palp has only a tarsus, tibia, genu and femur. The simple body division into gnathosoma and idiosoma is most easily seen in ticks and Mesostigmata, but can be subject to fusion and subdivision in other groups. For this reason mites in different groups often have different names for different parts of the body. In several groups the idiosoma is divided by a furrow called the sejugal furrow into two sections, which may be evident on the dorsal (back) side or on both dorsal and ventral side. These two sections are called the opisthosoma (rear body / abdomen) and the propodosoma (front-foot-body) which bears the first two pairs of legs. Figure 3: Arbitrary/jargon body divisions in mites. Many mites breathe through holes in their exoskeleton called stigmata (singular is stigma) which is often associated with a channel called a peritreme. These peritremes can be located on the sides of the animal (Mesostigmata), or around the mouthparts (Prostigmata), or may not be visible at all (Oribatida). The body parts of each of the major groups of acari are illustrated in the following sections. Ticks – Ixodidae and Argasidae Ticks are not normally found in soil, being blood-sucking parasites of vertebrates. Although they can be found living in nests or on vegetation. Superficially similar to mestostigmatid mites they do not feature in this key, but diagrams provided here should help in case a tick is found by accident. Besides hard ticks (ixodidae) there are also soft ticks (argasidae) which, in the UK, are parasites of bats, pigeons and seabirds. Subcapitulum with denticulate hypostome (attachment organ), palps usually with four or fewer segments and without an apotele. You are unlikely to find these in soil. Mesostigmata About one quarter of all mite species identified belong to the Mesostigmata. Mesostigmata are able to move all 6 leg segments independently, whereas in other orders the coxae are fused or absent. Mesostigmata undersides are a series of shields which differ between male and females, with flexible tissue between the shields called integument. Mesostigmata have a pair of lateral stigmatal openings usually around the level of coxae II-IV, often associated with elongated peritremes. A transverse genital aperture (cat flap) in the intercoxal region of the female covered by 1-4 epigynal shields. Astigmata Astigmata (also referred to as Astigmatina, Acaridaiae and Acaridida in some text books) are part of the Sarcoptiformes, and allied to the Oribatids, and recent phylogenies place them within the Oribatida. They tend to be white, hairy, range from relatively small to quite large and have only five free leg segments with the coxae fused to their body and reduced to internal sclerotized channels called apodemes. They can be very common within disturbed soils, and dust mites are part of this group. Astigmata do not have stigmatal opening or peritremes, and the gnathosoma is exposed. Perpendicular genital and anal openings (“bomb bay doors”), males aedeagus visible, females usually with two genital papillae. apodemes Oribatida Oribatids (sometimes referred to as beetle, box or moss mites): most oribatid mites inhabit the soil-litter layer and are considered to be the main decomposers of litter and organic matter. They have their idiosoma split into at least two sections when viewed from above (dorsal), comprising a front shield (the “prodorsum” ) covering the front of the body (proterosoma) often extending far forward covering the “head”. The rear “notogaster” (back-stomach), covering the hysterosome (rear of body). The prodorsum often supports two trichobothria, which are feather-like or other-shaped projections set in pits in the shield called bothridia (sing. bothridium). The true gnathosoma of these mites is often hidden under the projecting front part of the proterosome called a camerostome (“chamber-hole”). In most oribatids, the mouthparts can be seen to support two club-like organs called rutella (sing. rutellum) when viewed from the underside. The presence or absence of these, and the line where these join the underside of the head can be an important features depending on whether they form a straight line (diarthric) typical of most brachypyline mites, an angled line (stenarthric like a Ʌ), or are not jointed at all (anarthric). The chelicerae are often hard to see, tucked deep within the camerostome and are usually twosegmented and commonly chelate-dentate (although occasionally pelopsiform (elongated / suctorial)). The underside of Oribatids bear genital and anal plates, which in this group characteristically are arranged to enable them to open up along a vertical suture, rather like the bomb-bay doors of an aircraft. Prostigmata These mites are very variable in their form, and have the largest size range across all the mite groups (from very small <0.1mm to very large >1cm). They are mainly soft-bodied, with the idiosoma often wider towards the front than at the back. They can have eyes, which are rare among other groups, and may have strongly modified palps and chelicerae. They can either be brightly coloured (especially in larger species such as velvet mites) or very tiny and more or less transparent. Their genital and anal openings are not typically sclerotized and often only a suture line is visible (although still opening vertically like bomb-bay doors). They often have highly modified legs, which can finish in brooms of long seta, or large claws. The stigmatal opening is often difficult to see under a dissecting scope, however it is located near the chelicerae or at the top margin of the idiosoma. Collecting, handling and examining soil mites Soil mites can be collected for casual observation or recording simply by searching through litter or soil and collecting moving mites with a pooter. When you appreciate the scale of soil mites, however, the size of the meshes often provided on standard entomologists’ pooters can seem alarmingly large, and it probably worth adapting a small pooter with as fine a mesh as you can find. Placing litter or soil into a white plastic tray makes it easier to locate mites, by moving the litter to one side and watching for mites moving against the tray’s base. Placing litter or soil into a coarse mesh (>2mm) and gently agitating it over the plastic tray to let the mites fall into the tray along with smaller debris, can also be effective. It is worth noting that the faster moving predatory mites are relatively easy to spot using this technique, but slower moving microbivores may be harder to spot (particularly the box mites, which close up when disturbed). The hardest part is adapting your search technique to see the mites which are much smaller than other invertebrates obtained with a pooter – but once you have adapted to this scale, you will see them everywhere! Sweep nets can also be used to collect mites from vegetation. A more thorough and rigorous method for collecting mites is to extract the mites and other mesofauna from a litter sample or soil core using a Tullgren funnel. Litter or very fine flakey soil may benefit from being placed on top of a layer of wide-mesh cloth (e.g. muslin or cheesecloth) to prevent too many mite-sized particles from falling through the Tullgren funnel. Soil cores can be collected in any manner, but if you’re interested in using your data to describe the numbers or density of animals it’s best to collect a soil core of known volume. Because most soil mesofauna inhabits the topsoil, monitoring work such as that carried out for the Countryside Survey, or by Natural England, collects soil cores from 0 – 10cm deep, by driving lengths of plumber’s waste pipe into the soil and then digging it out, intact, with a trowel. Taping a gauze over the top of these pipes before driving them into the soil will stop the larger more active surface fauna from escaping once the core has been driven in. Once extracted from the soil, you should wrap up the core in clingfilm, or place within an individual plastic sample bag and keep it cool (a fridge at 4oC is ideal) until you’re ready to extract the soil animals. Tullgren funnels can be easily set up by amateurs using kitchen supplies and household goods, but if you’re doing a lot of work on mites, you’ll probably need to build something more elaborate. A Tullgren funnel is a funnel lined with a 2mm mesh suspended over a container of liquid, and held under a 40 watt incandescent light bulb (energy saving bulbs don’t emit enough heat!). This set up can also be created by using fairy lights placed directly within top of drainpipe, over mesh (Figure). Your litter or soil core is placed in the funnel and the soil organisms will be driven out of the sample by the light, heat and dryness, go through the mesh and fall into the liquid trap at the bottom. When using soil cores carefully remove any mesh from the core, and place it upside down in the funnel, with the original soil surface facing downwards. This is because larger organisms tend to inhabit the upper layers of soil, and may not be able to travel through the smaller pores found lower down in the soil. It normally takes 5 days to complete the extraction (however a lot depends on the size and wetness of the soil core), with a slow drying helping the animals to move through the soil, rather than drying them out in situ. The liquid can be water, if you’re interested in keeping your specimens alive (however you should aim to change collecting vial daily to prevent losses through predation). Springtails often float on the surface, while mites sink to the bottom. More commonly, you can use alcohol to kill and preserve your specimens until you’re ready to examine them. You can buy 70% industrial methylated spirits (without the purple dye) from suppliers of cleaning products, which is fine for preserving specimens, but if you’re interested in submitting your samples for genetic analysis then stronger alcohol is recommended. Of course, if you’re using alcohol you should keep it in a safe place, away from children and set up your extraction in a well ventilated area, child-free and away from sources of ignition. Preserving and recording You should aim to record details of your collection, even if you can’t identify the animals to species level, as is the case with this key. A good way to do this is to keep your specimens in at least 70% alcohol in a small glass or plastic vial, sealed with a bung, and include a slip of paper with key details of the specimen. You should use pencil (biro will dissolve in alcohol) and write your name, the date the sample was collected, the location (postcode is good, grid reference is better), habitat and what you think the specimen is. Because mites are usually very small, the immediate location can be described according to micro-habitat. Unfortunately most biological recording systems (such as www.irecord.org) will only allow you to record data at the level of species, or sometimes genus. This is a tall order for an amateur acarologist! This does not mean that your records are not useful, however. Much scientific research has been conducted usefully using taxonomic levels that are similar to, or at lower resolution, than the levels provided in this key. The Key This key is dichotomous, which means it relies on following one of two choices, based on a description of features. Choose the description that fits your specimen best, and if this seems to lead to the wrong answer, retrace your steps using the numbers in brackets, and explore whether you’ve gone down the correct route. The 4 major groups of mites in soil have their backgrounds colour coded with orange for Mesostigmata, pale blue for Astigmata, pale brown for Oribatida and pink for Prostigmata. Parts of the key leading to one or more of these groups are in white. Photographs of features as they appear under a light microscope, with bottom or top lighting, illustrate the features described. Please note that the features on your specimen may not always look exactly like the pictures here. Mites are very variable creatures. 1. 2.(1) 3.(2) Three pairs of legs (do not confuse palps with legs!) Four pairs of legs Six free leg segments (ie. coxae not fused into ventral shield or absent); palps with apotele on last segment. Stigmata and lateral peritremes always visible. Either disc-shaped animals or predatory mites “spider-like”, no trichobothria. Mite larva1 Five free leg segments (not counting claw, coxae fused to body) OR leg segments are inside a shell-like arrangement of shields and cannot easily be counted. Rear leg (leg IV) with small plate inserted near the base of the underside of the tarsus (segment nearest foot) bearing 2 bristles (setae). Epigynal shield either with 6 setae (rarely 2-4) Sejida; or a complex of 2-4 genital shields Trigynaspida. 11 Rear leg has no such small plate at the base of the tarsus. One epigynal shield with only 0-1 pairs of setae. 1 2 Order: Mesostigmata 3 Small plate (intercalary sclerite) Order: Mesostigmata Sub-orders: Trigynaspida or Sejida 10 Order: Mesostigmata Sub-order: Monogynaspida 4 You may still be able to key out a major group for this specimen – go to question 2 but be careful that some features may not match exactly. The exoskeletons of six-legged larvae may not be as well-armoured, for example. 4.(3) Roundish disc shaped animal (flying saucer like), each leg can be withdrawn into a hollow beside it (pedofossae). Beware that some oribatids can do this – count leg segments carefully and look for peritremes and stigmata! Order: Mesostigmata Cohort Uropodina, Sub-cohort Uropodiae pedofossa Stigmata 5.(4) 6(5) 7.(6) Not matching this description Animals with a pair of disc-shaped suckers either side of the anus, roughly circular and flattened. Parasites of myriopods. (rare) Not matching this description 5 Order: Mesostigmata Cohort Heterozerconina 6 Dorsum covered by more than two shields, including a pygidial shield/plate (back rear end) with setae Order: Mesostigmata Cohort Microgynoidea Order: Mesostigmata Cohort Gamasina 7 Order: Mesostigmata Sub-cohort Epicriiae, Dorsum (usually) covered by single shield no free mesonotal or pygidial plates with setae Dorsal side of body covered in patterns of numerous 3-4 lobed warts, sides with paired tubercles and long hairs – peritremes not visible. Tarsi with clubbed setae and paired claws. 8.(7) Body without patterns of warty lumps, tubercles. Predatory mites usually with peritremes visible along the lateral sides of the body behind the legs when viewing venter. Stigmata visible between legs III and IV. With or without paired claws. Dorsal shield resembles a police badge. In female – genital shield comes to a triangular point at its front edge. Male has spurs on leg IV only. In male there is a groove-like spermatotreme on the movable digit of the chelicerae. 8 Female genital shield squarer or axeshaped. Male has either unmodified chelicerae or long spermatodactyl, and may have spurs on leg II. 9 Adult females with a truncated dorsal shield not covering all of opisthonotal region (Males all covered). Pygidial sclerite resembling infinity (∞) (without Order: Mesostigmata Sub-cohort: Arctacariae, Order: Mesostigmata Sub-cohort: Parasitiae 9.(8) setae). Chelicerae enlarged. Peritremes extending to level of legs I or II only. (Very rare in UK) Animal not matching this description. Peritremes may be long (over many coxae) or short. Male: moveable digit of chelicerae with spermatodactyl, male genital opening at the front edge of ventral shield, males may have spurs or tubercles on legs II and or IV. Sternal shield entire with 2-4 pairs of setae. May lack claws on tarsus I. (Most common group). 10.(3) 11.(2) 12.(11) Females have a truncated diamond shaped genital plates made up of a small, rear central shield and two larger lateral shields (Tri = three and gynaspid = genital shield), although can be 2-4 shields. Males have genital opening at the front edge of a plate. Chelicerae may have mop- or brush-like excrescences at their bases. Female genital plate is a simple flap, males genital plate is a sub-circular valve set into the plate between the coxae (leg bases). Chelicerae without excresences, body often with tubercles or horn-like protrusions. Epigynal shield with 6 (or more) setae (occasionally only 2-4). Animals with only vestigial head and mouthparts present between front legs (Deuteronymph stage), and sucker-discs at the rear ventral side Animals with fully formed mouth parts and no sucker plates Often tiny animals, adult females with stigmatal opening either side of gnathosoma (near palps). End of leg IV not developed in females, hind legs either modified into long brush/broomlike structures, reduced to thin prongs, long whip-like hairs or single massive Superfamily Arctacaroidea Order: Mesostigmata Sub-cohort: Dermanyssiae Superfamilies within sub-cohort: 1) Veigaioidea 2) Rhodacaroidea 3) Eviphidoidea 4) Ascoidea 5) Phytoseioidea 6) Dermanyssoidea Order: Mesostigmata Sub-order: Trigynaspida Order: Mesostigmata Sub-order: Sejida Order: Sarcoptiformes; Sub-order: Oribatida; Cohort: Astigmata, Hypopi (phoretic life stage) 12 Order: Trombidiformes; Sub-order: Prostigmata; Super-Cohort: Eleutherengonides; Cohort: 13.(12) claws. Palps usually reduced to 2-3 segments. Heterostigmata Animal may be strongly or weakly sclerotised, but hind legs with claws or pads, all legs fully developed - never with brush/broom- or whip-like structures. Stigmatal openings at base or on gnathosoma, or not visible at all. Palps variable but usually 3-5 segments. Teardrop-shaped animals, leg coxae fused into ventral plate with apodemes, body normally white, often with long hairs, trichobothria always absent, with single claws or no claws. Females with v-shaped genital plates, male with aedegus and anal suckers. 13 Sejugal furrow Simple setae – no trichobothria Order: Sarcoptiformes; Sub-order: Oribatida; Cohort: Astigmata V-shaped genital plates Male anal region 14.(13) Animals not fitting this description, may/may not have trichobothria, may / may not have sejugal furrow, zero, single or multiple claws. Males do not have anal region as above. Animals with genital and anal plates open longitudinally like bomb-bay doors on aircraft. Commonly strongly sclerotised and redish, brownish or blackish, at least on legs. Body shape narrower at front than back (tear-drop). Mouthparts may have rutella, and 14 Order: Sarcoptiformes; Sub-order: Oribatida 15 chelicerae typically chelate (rarely modified) and palps are simple (never with thumbclaw, although not always easy to see). Often with dark-spots (oil glands) on both the rear flanks. Legs I-IV with 1 or 3 claws (rarely bidactyl), empodium claw or sucker-like never pad-like. Stigmata and peritremes NEVER present or visible. 15.(14) Genital and anal openings without plates. Often weakly sclerotised (beanbag-like), although some forms strongly sclerotized (labidostommatides), body often wider at front than at back, colour can be white, yellow, green, red. Mouthparts without rutella. Palps and chelicerae often modified (rarely chelate) always fairly visible. At least legs II and III with 2 lateral claws, empodium may be pad-like, often with tenant hairs. Lacking oil glands. Stigmata opening between cheliceral bases on gnathosoma. Order: Trombidiformes; Sub-order: Prostigmata 22 Animals ptychoid – prosoma and opisthosoma hinged (dorsal shields split into front and back that can fold up to enclose the animal – a bit armadillo/hedgehog-like). 16 16.(15) 17. (15) Animals not capable of folding up like this Body sclerotisation strong. Mouthparts with rutella broad (massive). Rear sides of body (opisthosome) may have oil glands Hyseterosoma not segmented. Genital and anal openings touching each other (resembles a sash window with four panes). Pretarsi often monodactyl or tridactyl Mouthparts lacking rutella/rutella narrow. Body sclerotisation strong. Opisthosomal glands absent. Hysterosoma sometimes segmented. Pretarsi various although usually monodactyl. Animals with obvious “knees” – genu is shorter than other leg segments (particularly tibia) and leg usually bends more here. Legs often within a deep pockets in body – like a ball and socket joint (acetabula). Genital and anal plates are set apart from each other, with no adjacent small adanal or adgenital shields, and are entirely enclosed in a single fused ventral shield, which continues to the mouthparts. Subcapitulum usually diarthric (no ventral sejugal furrow). [most common super-cohort]. 17 Sub-order: Oribatida Super-cohort: Mixonomata, Infra-orders: Euptyctima and Dichosomata. (Common name – box mites) Sub-order: Oribatida Super-cohort: Enarthronota Sub-order: Oribatida Super-cohort: Brachypylina – Higher oribatids (Euoribatida) Infra-orders: Pycnonoticae and Poronoticae. Small “knees” Genital and anal plates set apart in single shield Animals with most leg segments a similar size - genu not noticeably different from other leg segments. Genital and anal plates can be adjacent to each other, or not completely surrounded by ventral shield, or with adgenital or adanal plates. Ventral shield may be separated from coxal plates, and sejugal furrow may be present. Genu ~same as tibia not knee like Genital and anal plates may be adjacent 18.(17) Sub-order: Oribatida Macropyline – Super-cohorts: Parhyposomata, Palaeosomata, Desmonomata. Lower Oribatids 18 Adanal and adgenital plates present Weakly sclerotised bodies, often with long black hairs. Mouthparts always visible from above (astegasime) without rostral tectum. Femur (fourth from end of leg) divided – so appears to have 6 free leg segments (do not confuse with Mesostigmata). Never with oil glands on the sides of the opisthosoma, and legs never ending with single claw (2 or 3 claws). Sub-order: Oribatida Super-cohort: Palaeosomata Femur divided here 2 or 3 (as here) claws Femurs not divided, legs with only 5 free segments. Mouthparts can be hidden under projecting dorsal shield. Bodies usually light/dark brown with noticeable sclerotisation (rarely 19 19.(18) unsclerotised to some extent). Opisthosomal oil glands may be present, single claws may be present. Normal hairs on notogaster. Holoid body type - always lacking flexible cuticle between leg pairs II and III (though a furrow may be present). Adults always strongly sclerotised (though nymphs can be weakly sclerotised, wrinkly, with unsclerotised anal plates and single claws). Usually with distinct adgenital and/or adanal plates, always with opisthosomal oil glands, mouthparts always concealed under projecting front dorsal shield (stegasime). Sub-order: Oribatida Super-cohort: Desmonomata Dichoid body type - with flexible cuticle between legs 2 and 3, strongly or weakly sclerotised animals with or without oil glands, mouthparts concealed or visible from above. 20 Flexible cuticle 20.(19) 21.(20) Rutella either diarthic (connected to underside of head with joints in a relatively straight line) or fixed (not jointed). Oil glands absent. Usually only one claw on pretarsi. Notogastral scissures present (1-3). Rutella stenarthric (joined to underside of head with angled joints). Oil glands may be present. Notogastral scissures either not present or only 1 present. Notogaster entire with no scissures (cracks), never divided into 3 sections. Mouthparts normally hidden under projecting front dorsal shield. Oil glands may be absent (secondarily lost) or present. Brown, reddish, beige or yellow. Sub-order: Oribatida Super-cohort: Enarthronota (non – ptychoid) May have 1 scissure in notogaster, however notogaster often split into 3 Sub-order: Oribatida 21 Sub-order: Oribatida Super-cohort: Mixonomata (non ptychoid) 22.(14) sections – Trichoid body type (opistonotum divided after leg IV), with lateral, usually protruding oil glands. Mouthparts sometimes visible from above. White or tan coloured, weakly sclerotised. Legs ending in 1, 2 or 3 claws. Palps with fewer than 5 segments, no thumb claws on palp tibia. Palps with 5 segments, palp tibia with thumb claws. 23.(22) 24 (23) Super-cohort: Parhyposomata 23 25 24 Palps with 4 segments, large/obvious. Stigmatal opening at base of gnathosoma (no peritreme), on gnathosoma (with peritreme) or absent. Cheliceral bases not fused together. Relatively large mite >1mm. Palps with 3 or fewer segments – may be hard to see, although legs usually have a well-developed terminal claw and pad-like empodium. In females stigmata on either side of gnathosoma (near palps / shoulders of mite) no peritremes. Males often have genital suckers for copulation. White yellow or brown colouring. Never more than 1 pair of trichobothria present (on females only), and always capitate, like a blob on a short stalk). Usually smaller animals (<1mm), often have a parasitic or phoretic (travelling on insects) stage. Palps with 4 segments, with unusual fusion of genu and tibia, lacks thumbclaw. Coxae on venter expanded/plate like (Arnold Schwarzenegger mite – looks like a 6pack!). Dorsum and venter with alveolar ornamentation (to varying extents dependent on species). Stigmata opens at base of chelicerae. Femora of legs subdivided (up to 3 times). Heavily sclerotized. Sub-order: Prostigmata; Super-Cohort: Eleutherengonides; Cohort: Heterostigmata Sub-order: Prostigmata; Super-cohort: Labidostommatides 25 26.(25) Palps with 4 segments (rarely 1-5), sometimes borne on the sides of an extended snout. May have up to 2 pairs of trichobothria (various shapes), eyes and naso usually present. Can be large and brightly coloured animals (white, pink, yellow, green, red), although still weakly sclerotised. Leg I can be long and antenna-like, and femur IV is often swollen. Sub-order: Prostigmata; Super-cohort Eupodina (incl. superfamily Bdelloidea - snout mites) Trichobothria absent, leg tarsi with tenent (flattened tip – resembles head of a nail) hairs. Normally with modified chelicerae (as whips, blades, stylets), naso always absent, eyes usually present. Genital papillae absent in all stages of development. Adults are small to medium in size. Males with an aedagous. Trichobothria present (1 or 2 pairs). Genital papillae present normally 2-3 (but can be 0-3 pairs or reduced in size). Tenent hairs absent. Peritremes usually absent, however if present, located between cheliceral bases. Eye lenses well developed. Often hypertrichous (hairy) in terrestrial species. Have 1-2 pairs of trichobothria. Often medium to large animals, red or orange in colour, many with velvet-like coating. Sub-order: Prostigmata; Super-Cohort: Eleutherengonides; Cohort Raphignathae Peritremes well developed often as grooves across prodorsum at the base of the mouthparts, linear, or may emerge to form prongs with channels. Chelicerae often sickle-like, bases separate. Animals may be smaller, often purple, red, pink or brown. Not normally velvety. Usually with naso and well-defined thumb-claw. Not hypotrichous. 26 Sub-order: Prostigmata; Super-Cohort: Anystides; Cohort Parasitengona (Velvet Mites) (Large and diverse clade, with four sub-cohorts. Larvae typically parasitic) Sub-order: Prostigmata; Super-Cohort: Anystides; Cohort: Anystina