Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
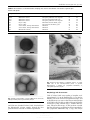
International Journal of Systematic and Evolutionary Microbiology (2000), 50, 2001–2008 Printed in Great Britain Acidilobus aceticus gen. nov., sp. nov., a novel anaerobic thermoacidophilic archaeon from continental hot vents in Kamchatka M. I. Prokofeva,1 M. L. Miroshnichenko,1 N. A. Kostrikina,1 N. A. Chernyh,1 B. B. Kuznetsov,2 T. P. Tourova1 and E. A. Bonch-Osmolovskaya1 Author for correspondence : E. A. Bonch-Osmolovskaya. Tel : j7 95 135 44 58. Fax : j7 95 135 65 30. e-mail : lbo!inmi.host.ru 1 Institute of Microbiology, Russian Academy of Sciences, Prospect 60 Let Oktyabrya 7/2, 117811, Moscow, Russia 2 Bioengineering Centre, Russian Academy of Sciences, Prospect 60 Let Oktyabrya 7/1, 117811, Moscow, Russia New thermoacidophilic organisms that were able to grow anaerobically on starch were isolated from the acidic hot springs of Kamchatka. Strain 1904T, isolated from a hot spring of the Moutnovski volcano, was characterized in detail. Its cells were regular or irregular cocci that were 1–2 µm in diameter, non-motile, and had a cell envelope consisting of one layer of subunits. The new organism was a hyperthermophile, growing in the temperature range 60–92 SC (with an optimum at 85 SC), an acidophile, having the pH range for growth of 20–60 (with an optimum at 38), and an obligate anaerobe. It fermented starch, forming acetate as the main growth product. Other growth substrates were yeast extract, beef extract and soya extract. Growth on yeast extract, beef extract and soya extract was stimulated by elemental sulfur, which was reduced to H2S. Acetate, arabinose, cellulose, formate, fructose, galactose, glucose, glycine, guar gum, lichenan, malate, maltose, methanol, pectin, pyruvate, propionate, xylan, xylose or a mixture of amino acids failed to support growth both in the presence and the absence of sulfur. When starch was used as the growth substrate, yeast extract (100 mg lV1) was required as a growth factor. The GMC content of the DNA was found to be 538 mol %. Comparison of the complete 16S rDNA sequence with databases revealed that the new isolate belonged to the kingdom Crenarchaeota. It was not closely related to any described genera (showing sequence similarity below 908 %) and formed a separate branch of the Crenarchaeota. On the basis of physiological differences and rRNA sequence data, a new genus – Acidilobus – is proposed, the type species being Acidilobus aceticus strain 1904T (l DSM 11585T). Keywords : Acidilobus aceticus, Archaea, hyperthermophiles, anaerobes, acidophiles INTRODUCTION Hyperthermophilic prokaryotes have a temperature optimum for growth of greater than 80 mC and are represented by both the Bacteria and the Archaea (Stetter, 1996). Although only neutrophilic species are known among hyperthermophilic bacteria, hyperthermophilic archaea include neutrophiles, alkaliphiles ................................................................................................................................................. The GenBank accession number for the 16S rDNA sequence of strain 1904T is AF191225. and acidophiles. Alkaliphilic hyperthermophilic archaea are currently represented by a few species of the genus Thermococcus (Keller et al., 1995 ; Dirmeier et al., 1998), which have a pH optimum for growth of 9n0. Thermoacidophilic isolates are much more numerous and comprise mostly organisms with respiratory metabolism, i.e. obligate or facultative aerobes (Scho$ nheit & Scha$ fer, 1995), and just one obligate anaerobe, Stigiolobus azoricus (Segerer et al., 1991), which can grow at pH 1n0–5n5 by means of lithotrophic sulfur respiration by utilizing molecular hydrogen as the sole growth substrate. Among the acidophiles, 01268 # 2000 IUMS 2001 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03 M. I. Prokofeva and others fermentative metabolism has been detected only in Thermoplasma acidophilum (Segerer et al., 1988), a facultative aerobe and moderate thermophile. Here we report the isolation of a new obligately anaerobic organism capable of fermentative growth at 85 mC and pH 3n8. METHODS Sampling and enrichment. Samples of water, mud and soil were taken from solfataric fields (in Kamchatka) and were shown to have pH values of 4n0 or less. The samples were put in 100 ml flasks with screw caps, closed hermetically, transported to the laboratory and used without delay for inoculation of the basal medium, which contained the following (mg lV") : KCl, 330 ; NH Cl, 330 ; KH PO , 330 ; # O,% 500 ; MgCl .6H O, 330 ; CaCl .2H O, %330 ; Na S.9H # # # # # starch, 5000 ; yeast extract (Difco), 100 ; resazurin, #1. Traceelement (Kevbrin & Zavarzin, 1992) and vitamin (Wolin et al., 1963) stock solutions were added at 1 ml lV". The medium was prepared anaerobically under a flow of oxygenfree CO . After the reduction of the medium by boiling and # consequent addition of Na S.9H O (700 mg lV") and de# # colourization of resazurin, the pH was adjusted to 3.0 by the addition of 5 M H SO . Portions of the medium (20 ml) were # bottles % dispensed into 50 ml with screw caps, filled with CO and sterilized at 110 mC. Each bottle was inoculated with 1 g# of sample and incubated at 80 mC. Microbial growth was monitored using light microscopy. Isolation of pure cultures. For isolation of colonies, flat 100 ml bottles (Bellco) containing 3 ml of the same medium solidified with 0n8 % (w\v) Gelrite and 0n05 % (w\v) MgSO .7H O were used. Individual colonies were picked up under a% flow# of oxygen-free CO and transferred to the liquid # was tested by using light medium. The purity of isolates microscopy of cultures in different growth conditions. Morphology and ultrastructure studies. The morphology of new isolates was examined using phase-contrast light microscopy. The ultrastructure was studied in a JEM-100 electron microscope, with samples prepared as described elsewhere (Bonch-Osmolovskaya et al., 1990). Metabolic studies. Potential growth substrates (5000 mg lV") were added to the basal medium prepared without starch. Organic acids were added as their sodium salts, and a mixture of 20 amino acids was used (SAG-II). The headspace was filled with 100 % CO ; when molecular hydrogen was # it was used in a mixture with tested as a growth substrate, CO (8 : 2, v\v). Growth was measured by direct cell counting # the light microscope. Growth products were detected under using methods described earlier (Miroshnichenko et al., 1994). The temperature range and optimum for growth were determined on the medium with yeast extract and elemental sulfur at pH 3n8. The pH range and optimum for growth were determined at 80 mC. Possible electron acceptors added to the basal medium included elemental sulfur as sulfur flower powder (10 000 mg lV"), nitrate as the sodium salt, (500, 1000 and 2500 mg lV") and Fe(III) as amorphous ferric oxide [90 mM Fe(III) ; Slobodkin et al., 1999] or Fe(III) citrate (20 mM). Sensitivity to antibiotics. The influence of penicillin, strep- tomycin (both at 500 mg lV") and chloramphenicol (100 mg lV") on the growth of the new isolate was tested. The stability of the antibiotics at high temperature and low pH was checked by observing their effects on the growth of 2002 Thiobacillus ferrooxidans VKM B-458, after preliminary incubation for 3 d at 80 mC and pH 2n5. Determination of DNA GjC content. The GjC content of the genomic DNA was determined as reported earlier (Miroshnichenko et al., 1994). Sequencing and analysis of 16S rDNA. The 16S rRNA gene was selectively amplified from the genomic DNA by a PCR using 5h-AGAGTTTGATCCTGGCTCAG-3h as the forward primer and 5h-TACGGTTACCTTGTTACGACTT-3h as the reverse primer (Lane, 1991). The PCR reaction was carried out in 100 µl reaction mixture containing 1 µg DNA template, 200 µM (each) primer, 200 µM (each) dNTP and 3 U Tet-z polymerase (BioMaster) in reaction buffer (100 mM Tris\HCl pH 8n3, 500 mM KCl, 20 mM MgCl ). The temperature regime involved 30 amplification cycles# of 1 min at 94 mC, 1 min at 42 mC and 1 min at 72 mC. The final extension was carried out at 72 mC for 6 min. The PCR products were purified using the PCR-prep kit (Promega) as recommended by the manufacturer. The 16S rRNA gene was sequenced in both directions using the forward and reverse primers. The DNA sequencing was performed by using USB Sequenase version 2 kit. The 16S rDNA sequence was aligned with a representative set of analogous sequences obtained from the latest versions of the Ribosomal Database Project (RDP) or from GenBank by using the software (Corpet, 1988). Positions that had not been sequenced in one or more reference organisms were omitted and a total of 1247 nucleotides were used in the analysis. Pairwise evolutionary distances were computed by using the correction of Jukes & Cantor (1969). The phylogenetic tree was constructed by the neighbourjoining method (Saitou & Nei, 1987) with bootstrap analysis of 100 trees using the programs of the package (Van de Peer & De Wachter, 1994), with Methanococcus vannielii as the outgroup. Nucleotide sequence accession numbers. The accession numbers of the sequences used as references are as follows : Sulfurisphaera ohwakuensis TA1T, D85507 ; Sulfolobus acidocaldarius DSM 639T, D14053 ; Thermosphaera aggregans M11TLT, X99556 ; Staphylothermus marinus F1 DSM 3639T, X99560 ; Hyperthermus butylicus DSM 5456T, X99553 ; Sulfophobococcus zilligii K1T, X98064 ; Stygiolobus azoricus DSM 6296T, D85520 ; Acidianus infernus DSM 3191T, D85505 ; Aeropyrum pernix K1T, D83259 ; Metallosphaera sedula DSM 5348T, X90481 ; Stetteria hydrogenophila 4ABCT, Y07963 ; Pyrodictium occultum PL19, M21087 ; Desulfurococcus mobilis, M36474 ; Thermofilum pendens Hvv3T, X14835 ; Thermoproteus tenax, M35966 ; Pyrobaculum islandicum geo3T, L07511 ; Caldivirga maquilingensis IC-167T, AB013926 ; Thermocladium modestius IC-125T, AB005296 ; and ‘ Caldococcus noboribetus ’ NC12, D85038. RESULTS Enrichment and isolation Samples obtained from acidic hot springs from different areas in Kamchatka were used to inoculate anaerobic starch medium at pH 3n0. After 2–5 d incubation at 80 mC, eight samples showed microbial growth (Table 1). Cells of irregular coccoid shape were observed in all cultures. When transferred to Gelritesolidified medium, smooth white colonies approximately 1 mm in diameter appeared after 7 d incubation. By isolating single colonies, strains 1904T, 1919 International Journal of Systematic and Evolutionary Microbiology 50 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03 Acidilobus aceticus gen. nov., sp. nov. Table 1. Characteristics of the Kamchatka sampling sites used for enrichment of anaerobic organotrophic thermoacidophiles Sample no. 1904T 1919 1920 301 310 321 322 345 Location Description Temp. (mC) pH Mutnovski volcano Mutnovski volcano Mutnovski volcano Geyser valley Geyser valley Uzon caldera, Orange thermal field Uzon caldera, Orange thermal field Moutnovski volcano Hot spring with ferric iron deposits Hot well surrounded with grass Hot spring with ferric iron deposits Black mud near the Giant geyser Hot well near the Giant geyser Mud hole Mud hole Mud hole 87 90 90 91 80 70 87 82 4n0 3n0 3n0 4n5 4n5 4n0 4n5 4n0 (a) (b) S ol (c) cm ................................................................................................................................................. Fig. 2. Electron micrographs of ultrathin sections of isolate 1904T cells in the late-exponential phase of growth. Abbreviations : s, subunit ; cm, cytoplasmic membrane ; ol, osmiophilic layer. Bars, 0n5 µm. Morphology and ultrastructure ................................................................................................................................................. Fig. 1. Electron micrographs of isolate 1904T whole cells in the late-exponential phase of growth. Bars, 0n5 µm. and 1920 were obtained. Isolate 1904T, obtained from the Moutnovski volcano sample, showed the best growth and was selected for further experiments. Cells of isolate 1904T were regular to irregular cocci approximately 1–2 µm in diameter (Fig. 1, top). Active motility was never observed in light microscope, and no flagella were ever seen on the electron micrographs of the whole cells. Double or triple cells were frequently observed in young cultures (Fig. 1, middle and bottom). Electron microscopy of thin sections revealed that the cell envelope comprised an S-layer attached to the cytoplasmic membrane (Fig. 2, top). The S-layer International Journal of Systematic and Evolutionary Microbiology 50 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03 2003 7 Cells (i10 –6 ml –1) Doubling time (h) 6 5 4 3 2 70 75 80 85 90 Temperature (°C) 95 T Fig. 3. Influence of temperature on the growth of isolate 1904 on the medium with yeast extract and elemental sulfur at pH 3n8. 25 2·5 20 2·0 15 1·5 10 1·0 5 0·5 2 3 4 5 Time (d) 6 7 8 0 ................................................................................................................................................. Fig. 5. Production of H2S ( ) and acetate (>), and growth ($), of isolate 1904T on the medium with yeast extract and S0. was 2n5 h. The maximum cell concentration achieved was 2–2n5i10( cells mlV". Isolate 1904T was found to be a strict anaerobe unable to grow without prereduction of the medium. 13 Isolate 1904T was found to grow on starch, yeast extract, beef extract and soya extract. None of the following supported growth : acetate, arabinose, cellulose, formate, fructose, galactose, glucose, glycine, guar gum, lichenan, malate, maltose, methanol, pectin, pyruvate, propionate, xylan, xylose or a mixture of 20 amino acids. Yeast extract (100 mg lV") was required as a growth factor when starch was used as the growth substrate. 11 Doubling time (h) 3 1 100 ................................................................................................................................................. 9 7 5 3 1 1 30 Acetate, H2S (µmol ml –1) M. I. Prokofeva and others 2 3 4 5 6 pH ................................................................................................................................................. Fig. 4. Influence of pH on the growth of isolate 1904T on the medium with yeast extract and elemental sulfur (incubation temperature, 80 mC). Growth of isolate 1904T on a medium with yeast extract, beef extract and soya extract as the growth substrates was stimulated by elemental sulfur, which was reduced to hydrogen sulfide. On a medium with starch, the stimulatory effect of sulfur was less prominent. Nitrate, at all concentrations tested, did not influence growth on either medium. Fe(III), added as ferric oxide or ferric citrate, did inhibit growth. was composed of a single layer of subunits covered with a thin osmophilic layer that was probably proteinaceous in nature (Fig. 2, bottom). The main metabolic product of isolate 1904T, both on yeast extract (Fig. 5) and on starch, was acetate. Neither H nor ethanol was formed in the course of growth on# any of the substrates used. Isolate 1904T was shown to be resistant to penicillin, streptomycin and chloramphenicol. Physiology The GjC content of the DNA Isolate 1904T grew at temperatures between 60 and 92 mC, with an optimum at 85 mC (Fig. 3), and within a pH range of 2n0–6n0, with an optimum at 3n8 (Fig. 4). The minimum doubling time under optimal conditions The GjC content of the total DNA of isolate 1904T was 53n8 mol %. Similar values were obtained for other isolates : 54n5 mol % (isolate 1919), 55n1 mol % (isolate 1920). 2004 International Journal of Systematic and Evolutionary Microbiology 50 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03 Acidilobus aceticus gen. nov., sp. nov. ................................................................................................................................................. Fig. 6. Phylogenetic tree showing the relationship between novel strain 1904T and other archaea, based on a comparison of 16S rDNA sequences. compared with the corresponding sequence data from the RDP. This analysis revealed that new isolate 1904T was a member of the kingdom Crenarchaeota of the domain Archaea. Additional sequence alignments and phylogenetic analyses performed with type species of the validly published genera of this kingdom showed that strain 1904T was not closely related to any reference organism (82n5–90n8 % sequence similarity). In the phylogenetic tree (Fig. 6), strain 1904T formed a single cluster with Aeropyrum pernix (90n8 % sequence similarity), but the bootstrap probability of this branching point was not high (80 %). More detailed analysis of the 16S rRNA gene of strain 1904T revealed unique deviation from the known crenarchaeotal sequences at positions where all other Crenarchaeota exhibited conserved bases (Table 2). Of these changes, the deviations at positions 592 : 647, 1308 : 1329 and 1310 : 1327 occurred (using the inferred secondary structure) in helixes and involved paired bases (compensatory changes). When the 16S rRNA sequence of isolate 1904T was compared with all available sequences, it exhibited a high degree of 16S rRNA sequence similarity (98n3 %) to crenarchaeotal strain NC12 (‘ Caldococcus noboribetus ’) and had the same signature nucleotides. Table 2. Sequence signatures of the 16S rRNA from isolate 1904T DISCUSSION Sequence position* 321 : 332 592 : 647 678 913 1302 1308 : 1329 1310 : 1327 1335 1393 1414 Corresponding bases† Isolate 1904T Common to crenarchaeotal species‡ C:G C:G C U C U:A G:C C U U A:G G (u,a) : C (a,u) U A U (a) C:G A:U G C C * Numbering according to Escherichia coli nomenclature. † Base pairing was deduced from secondary-structure assignment. ‡ All of the crenarchaeotal 16S rRNA sequences were from the Ribosomal Database Project ; lower-case letters indicate bases found in less than 15 % of assayable cases. Analysis of 16S rDNA The almost complete (1411-nucleotide) sequence of the 16S rRNA gene (corresponding to positions 27–1479 using Escherichia coli numbering) of isolate 1904T was determined. It was found to have a high GjC content (67n3 mol %), like the 16S rRNA genes of other thermophilic prokaryotic organisms. In an initial analysis, the 16S rDNA sequence of strain 1904T was Hot springs with low-pH water are quite common in different volcanic areas of the world (Brock, 1978 ; Stetter et al., 1990). The first representative of extremely thermophilic archaea, Sulfolobus solfataricus, was isolated in 1972 and was a thermoacidophile growing optimally at 70–75 mC and pH 2n0–3n0 (Brock et al., 1972). More recently, significant progress has been made in investigations of both the phenotypic and the phylogenetic diversity of hyperthermophiles (Blo$ chl et al., 1995 ; Stetter, 1996) ; 10 genera of thermoacidophilic archaea have been described (Table 3). Most of the genera belong to the Crenarchaeota, with the exception of Thermoplasma (Segerer et al., 1988) and Picrophilus (Schleper et al., 1995), which belong to the order Thermoplasmales of the kingdom Euryarchaeota. The majority of the thermoacidophilic archaea are aerobes capable of lithotrophic growth with elemental sulfur, sulfides or molecular hydrogen as electron donors (Stetter, 1996 ; Scho$ nheit & Scha$ fer, 1995). Some of them, like Acidianus infernus (Segerer et al., 1986), are facultative anaerobes and are capable of anaerobic growth with molecular hydrogen by using elemental sulfur as an alternative electron acceptor. The metabolism of Thermoproteus tenax (Zillig et al., 1981) and Stigiolobus azoricus (Segerer et al., 1991), which are obligate anaerobes, is also based on this reaction. Whilst most of the organisms mentioned above have coccoid or flat irregular cells, representatives of the genus Thermoproteus have rod-shaped cells (Zillig et al., 1981). In this genus, it is only the type species, Thermoproteus tenax, that is moderately acidophilic, growing at pH values in the range 2n5–6n0 (with International Journal of Systematic and Evolutionary Microbiology 50 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03 2005 M. I. Prokofeva and others Table 3. Characteristics of thermoacidophilic archaea ................................................................................................................................................................................................................................................................................................................. Abbreviations : R, respiration ; F, fermentation ; Org, organic substrates. Genus/type species Topt. (mC) pHopt. Type of metabolism Sulfolobus\S. acidocaldarius 70–75 2–3 R Thermoproteus\T. tenax Sulfurococcus\S. mirabilis Acidianus\A. infernus Thermoplasma\T. acidophilum Metallosphaera\M. sedula Stigiolobus\S. azoricus Picrophilus\P. oshimae Sulfurisphaera\S. ohwakuensis Thermocladium\T. modestius 90 70–75 90 59 75 80 60 84 75 5 2–3 2 1–2 1–4n5 2n5–3 0n7 2n0 4n0 R R R F\R R R R R R 85 3n8 F(R?) Isolate 1904T an optimum at pH 5n0). Other species, as well as representatives of the genus Pyrobaculum (Huber et al., 1987), grow optimally at pH 6n0–7n0. Members of these two genera also have a respiratory metabolism and are capable of lithotrophic or organotrophic growth via sulfur respiration (Scho$ nheit & Scha$ fer, 1995) or, in the case of Pyrobaculum aerophilum (Vo$ lkl et al., 1993), via aerobic respiration or nitrate reduction. The fermentative growth capacity was observed only with Thermoproteus uzoniensis (BonchOsmolovskaya et al., 1991), which is an obligate anaerobe and neutrophile. This type of metabolism is quite common among representatives of the neutrophilic, hyperthermophilic archaea (Scho$ nheit & Scha$ fer, 1995). In the Euryarchaeota kingdom, they are represented by the genera Thermococcus (Zillig et al., 1983) and Pyrococcus (Fiala & Stetter, 1986). In the Crenarchaeota, obligately anaerobic fermentative micro-organisms belong to the genera Desulfurococcus (Zillig et al., 1982), Staphylothermus (Fiala et al., 1986), Hyperthermus (Zillig et al., 1990), Thermosphaera (Huber et al., 1998), Sulphophobococcus (Hensel et al., 1997) and Pyrodictium (Pley et al., 1991). All these organisms are capable of fermenting peptides and\or polysaccharides and produce volatile fatty acids, hydrogen and CO . Stetteria hydrogenophila # (Jochimsen et al., 1997) requires molecular hydrogen and elemental sulfur for its organotrophic growth and thus was considered to possess a respiratory type of metabolism. All these organisms are extreme thermophiles or hyperthermophiles, and neutrophiles. Our findings show that obligately anaerobic organisms with fermentative metabolism are widespread in Kamchatka hot springs with low-pH water. Organic substrates for their growth might be synthesized by lithotrophic components of the same microbial community (Bonch-Osmolovskaya et al., 1999) or might 2006 Electron donors Electron acceptors H , org, S!, O # # Fe#V, S O#V % ' S! H ,org # Org, S!, Fe#V, S O#V O % ' # S!, Fe#V, H S!, O # # Org O # V H , org, S!, S O# O # % ' # H S! # Org O # H , org, S! S!, O # V # Org S!, SO# , S O#V % # $ -cystine Org S! Reference Brock et al. (1972) Zillig et al. (1981) Golovacheva et al. (1985) Segerer et al. (1986) Segerer et al. (1988) Huber et al. (1989) Segerer et al. (1991) Schleper et al. (1995) Kurosawa et al. (1998) Itoh et al. (1998) This work come from surrounding areas with abundant vegetation. Isolate 1904T, described in this work, is a true hyperthermophile, having a temperature optimum for growth of 85 mC, and is a true acidophile, having a pH optimum for growth of 3n8. Isolate 1904T produced significant growth on organic substrates in the absence of elemental sulfur ; acetate was the main metabolic product. The metabolism of this isolate, therefore, could be characterized as fermentative. The growthstimulating action of sulfur could be explained by the need for an additional electron sink for the fermentation of some of the yeast-extract components. This phenomenon was shown for several organotrophic, hyperthermophilic archaea (Fiala & Stetter, 1986 ; Bonch-Osmolovskaya & Miroshnichenko, 1994). However, the possibility of respiratory metabolism in this organism cannot be excluded until additional experiments have been performed. Three other strains, isolated from different terrestrial thermal habitats of low pH, phenotypically resemble isolate 1904T and have similar GjC DNA contents. All of the isolates probably represent the same species. Comparison of an almost complete 16S rDNA sequence from isolate 1904T with nucleotide sequences of reference micro-organisms showed that it does not belong to any validly described Crenarchaeota genera. Isolate 1904T exhibited a high degree of 16S rDNA sequence similarity (98n3 %) and shared signature nucleotides only with crenarchaeotal strain NC12 (‘ C. noboribetus ’ ; Aoshima et al., 1996). The description of this organism, however, has never been published and strain NC12 was not available for the comparison. Of the representatives of the validated Crenarchaeota taxa, the closest to our isolate was A. pernix (90n8 % similarity). This organism is a neutrophile and an aerobe (Sako et al., 1996). On the basis of the differentiating phenotypic and genomic features of International Journal of Systematic and Evolutionary Microbiology 50 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03 Acidilobus aceticus gen. nov., sp. nov. isolate 1904T, we propose that this isolate should be assigned to a new genus – Acidilobus – the type species being Acidilobus aceticus. Description of Acidilobus gen. nov. Acidilobus (A.ci.di.lohbus. L. masc. adj. acidus acid ; Gr. masc. n. lobos lobe ; M.L. masc. n. Acidilobus acid lobe). Cells are regular to irregular cocci. The cell envelope consists of an S-layer attached to the cytoplasmic membrane. Archaeon. Hyperthermophile. Acidophile. Obligate anaerobe. Organotroph. Peptides and polysaccharides serve as energy and carbon sources. Acetate is the major growth product. Elemental sulfur stimulates growth and is reduced to hydrogen sulfide. Resistant to antibiotics. The type species is Acidilobus aceticus. The habitat is terrestrial acidic hot springs. nov. – a new thermophilic sulfur-reducing eubacterium. Arch Microbiol 153, 151–155. Bonch-Osmolovskaya, E. A., Miroshnichenko, M. L., Kostrikina, N. A., Chernyh, N. A. & Zavarzin, G. A. (1991). Thermoproteus uzoniensis sp. nov., a new extremely thermophilic archaebacterium from Kamchatka continental hot springs. Arch Microbiol 154, 556–559. Bonch-Osmolovskaya, E. A. & Miroshnichenko, M. L. (1994). The influence of molecular hydrogen and elemental sulfur on the metabolism of extremely thermophilic archaea of genus Thermococcus. Microbiology (English translation of Mikrobiologiya) 63, 777–782. Bonch-Osmolovskaya, E. A., Miroshnichenko, M. L., Slobodkin, A. I. & 7 other authors (1999). Biodiversity of anaerobic lithotrophic prokaryotes in terrestrial hot springs of Kamchatka. Microbiology (English translation of Mikrobiologiya) 68, 398–406. Brock, T. D. (1978). Thermophilic Microorganisms and Life at High Temperatures. New York : Springer. Brock, T. D., Brock, K. M., Belly, R. T. & Wiess, R. L. (1972). Description of Acidilobus aceticus sp. nov. Acidilobus aceticus (a.cehti.cus. L. masc. adj. aceticus producing acetate). Cells are non-motile, regular to irregular cocci approximately 1–2 µm in diameter. The cell envelope consists of an S-layer attached to the cytoplasmic membrane. Growth occurs in the temperature range 60–92 mC (optimum at 85 mC) and in the pH range 2n0–6n0 (optimum at 3n8). Strictly anaerobic. Heterotrophic. Yeast extract, beef extract, soya extract and starch may serve as growth substrates. Elemental sulfur stimulates growth on yeast extract but is not obligately required. Growth products are acetate and, in the presence of S!, H S. Resistant to penicillin, strep# tomycin and chloramphenicol. The source of isolation was a solfataric field near the Moutnovski volcano, Kamchatka. The GjC content of the DNA is 53n8 mol %. The type strain is Acidilobus aceticus 1904T (l DSM 11585T). ACKNOWLEDGEMENTS This work was supported by the Russian Foundation for Basic Research, grants no. 96-04-49463 and 99-04-48360 and by the ‘ Biodiversity Programme ’ of the Russian Ministry of Science and Technology. We also thank T. A. Pivovarova (Institute of Microbiology, Russian Academy of Sciences) for the strain of Thiobacillus ferrooxidans used in this work. REFERENCES Aoshima, M., Nishibe, Y., Hasegawa, M., Yamagishi, A. & Oshima, T. (1996). Cloning and sequencing of a gene encoding 16S ribosomal RNA from a novel hyperthermophilic archaebacterium NC12. Gene 180, 183–187. Blo$ chl, E., Burggraf, S., Fiala, G. & 7 other authors (1995). Isolation, taxonomy and phylogeny of hyperthermophilic microorganisms. World J Microbiol Biotechnol 11, 9–16. Bonch-Osmolovskaya, E. A., Sokolova, T. G., Kostrikina, N. A. & Zavarzin, G. A. (1990). Desulfurella acetivorans gen. nov., sp. Sulfolobus : a novel genus of sulfur-oxidizing bacteria living at low pH and high temperature. Arch Microbiol 84, 54–68. Corpet, F. (1988). Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res 16, 10881–10890. Dirmeier, R., Keller, M., Hafenbradl, D., Braun, F.-J., Rachel, R., Burggraf, S. & Stetter, K. O. (1998). Thermococcus acidamino- vorans sp. nov., a new hyperthermophilic alkalophilic archaeon growing on amino acids. Extremophiles 2, 109–114. Fiala, G. & Stetter, K. O. (1986). Pyrococcus furiosus sp. nov. represents a novel genus of marine heterotrophic archaebacteria growing optimally at 100 mC. Arch Microbiol 45, 56–61. Fiala, G., Stetter, K. O., Jannasch, H. W., Langworthy, T. H. & Madon, J. (1986). Staphylothermus marinus sp. nov. represents a novel genus of thermophilic submarine organotrophic archaebacteria growing up to 98 mC. Syst Appl Microbiol 8, 106–113. Golovacheva, R. S., Val’ekho-Roman, K. M. & Troitskii, A. V. (1985). Sulfurococcus mirabilis gen. nov., sp. nov., a new thermophilic archaebacterium with the ability to oxidize sulfur. Microbiology (English translation of Mikrobiologiya) 56, 100–107. Hensel, R., Matussek, K., Michalke, K., Tacke, L., Tindall, B. J., Kohlhoff, M., Siebers, B. & Dielenschneider, J. (1997). Sulpho- phobococcus zilligii gen. nov., sp. nov., a novel hyperthermophilic archaeum isolated from hot alkaline spring of Iceland. Syst Appl Microbiol 20, 102–110. Huber, R., Kristiansson, J. K. & Stetter, K. O. (1987). Pyrobaculum gen. nov., a new genus of neutrophilic, rod-shaped archaebacteria from continental solfataras growing optimally at 100 mC. Arch Microbiol 149, 95–101. Huber, G., Spinnler, C., Gambacorta, A. & Stetter, K. O. (1989). Metallosphaera sedula gen. nov. and sp. nov. represents a new genus of aerobic, metal-mobilizing, thermoacidophilic archaebacteria. Syst Appl Microbiol 12, 38–47. Huber, R., Dyba, D., Huber, H., Burggraf, S. & Rachel, R. (1998). Sulfur-inhibited Thermosphaera aggregans sp. nov., a new genus of hyperthermophilic archaea isolated after its prediction from environmentally derived 16S rRNA sequences. Int J Syst Bacteriol 48, 31–38. Itoh, T., Suzuki, K. & Nakase, T. (1998). Thermocladium modestius gen. nov., sp. nov., a new genus of rod-shaped, extremely thermophilic crenarchaeote. Int J Syst Bacteriol 48, 879–887. Jochimsen, B., Piensman-Simon, S., Volker, H., Stuben, D., Botz, International Journal of Systematic and Evolutionary Microbiology 50 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03 2007 M. I. Prokofeva and others R., Stoffers, P., Dando, P. R. & Thomm, M. (1997). Stetteria hydrogenophila, gen. nov. and sp. nov., a novel mixotrophic sulfur-dependent crenarchaeote isolated from Milos, Greece. Extremophile 1, 67–73. Jukes, T. H. & Cantor, C. R. (1969). Evolution of protein molecules. In Mammalian Protein Metabolism, pp. 21–132. Edited by H. N. Munro. New York : Academic Press. Keller, M., Braun, F.-J., Dirmeier, R., Hafenbradl, D., Burgraff, S., Rachel, R. & Stetter, K. O. (1995). Thermococcus alkaliphilus sp. nov., a new hyperthermophilic archaeum growing on polysulfide at alkaline pH. Arch Microbiol 164, 390–395. Kevbrin, V. V. & Zavarzin, G. A. (1992). The influence of sulfur compounds on the growth of halophilic homoacetic bacterium Acetohalobium arabaticum. Microbiology (English translation of Mikrobiologiya) 61, 812–817. Kurosawa, N., Itoh, Y. H., Iwai, T., Sugai, A., Uda, I., Kimura, N., Horiuchi, T. & Itoh, T. (1998). Sulfurisphaera ohwakuensis gen. nov., sp. nov., a novel extremely thermophilic acidophile of the order Sulfolobales. Int J Syst Bacteriol 48, 451–456. Lane, D. J. (1991). 16S\23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics, pp. 115–147. Edited by E. Stackebrandt & M. Goodfellow. New York : Wiley. Miroshnichenko, M. L., Gongadze, G. M., Lysenko, L. M. & BonchOsmolovskaya, E. A. (1994). Desulfurella multipotens sp. nov., a new sulfur-respiring thermophilic eubacterium from Raoul Island (Kermadec archipelago, New Zealand). Arch Microbiol 161, 88–93. Pley, U., Schipka, J., Gambacorta, A., Jannasch, H. W., Fricke, H., Rachel, R. & Stetter, K. O. (1991). Pyrodictium abissi sp. nov. represents a novel heterotrophic marine archaeal hyperthermophile growing at 110 mC. Syst Appl Microbiol 14, 245–253. Saitou, N. & Nei, M. (1987). The neighbour-joining method : a new method for reconstructing phylogenetic trees. Mol Biol Evol 4, 406–425. Sako, Y., Nomura, N., Uchida, A., Ishida, Y., Morii, H., Koga, Y., Hoaki, T. & Maruyama, T. (1996). Aeropyrum pernix gen. nov., sp. nov., a novel aerobic hyperthermophilic archaeon growing at temperatures up to 100 mC. Int J Syst Bacteriol 46, 1070–1077. Schleper, C., Puehler, G., Holz, I., Gambacorta, A., Janecovic, D., Santarius, U., Klenk, H.-P. & Zillig, W. (1995). Picrophilus gen. nov., fam. nov. : a novel aerobic, heterotrophic, thermoacidophilic genus and family comprising archaea capable of growth around pH l 0. J Bacteriol 177, 7050–7059. Scho$ nheit, P. & Scha$ fer, T. (1995). Metabolism of hyperthermophiles. World J Microbiol Biotechnol 11, 26–57. Segerer, A., Neuner, A., Kristjansson, J. K. & Stetter, K. O. (1986). Acidianus infernus gen. nov., sp. nov., and Acidianus brierleyi 2008 comb. nov. : facultatively aerobic, extremely acidophilic thermophilic sulfur-metabolizing archaebacteria. Int J Syst Bacteriol 36, 559–564. Segerer, A., Langworthy, T. A. & Stetter, K. O. (1988). Thermoplasma acidophilum and Thermoplasma volcanium sp. nov. from solfataric fields. Syst Appl Microbiol 10, 161–171. Segerer, A., Trincone, A., Gahrtz, M. & Stetter, K. O. (1991). Stigiolobus azoricus gen. and sp. nov. represents a novel genus of anaerobic, extremely thermoacidophilic archaea of the order Sulfolobales. J Bacteriol 41, 495–501. Slobodkin, A. I., Tourova, T. P., Kuznetsov, B. B., Kostrikina, N. A., Chernyh, N. A. & Bonch-Osmolovskaya, E. A. (1999). Thermoanaerobacter siderophilus sp. nov., a novel dissimilatory Fe(III)-reducing, anaerobic, thermophilic bacterium. Int J Syst Bacteriol 49, 1471–1478. Stetter, K. O. (1996). Hyperthermophilic procaryotes. FEMS Microbiol Rev 18, 149–158. Stetter, K. O., Fiala, G., Huber, G., Huber, R. & Segerer, A. (1990). Hyperthermophilic microorganisms. FEMS Microbiol Rev 75, 351–382. Van de Peer, Y. & De Wachter, R. (1994). for Windows : a software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. CABIOS 10, 569–570. Vo$ lkl, P., Huber, R., Drobner, E., Rahel, R., Burggraf, S., Tricone, A. & Stetter, K. O. (1993). Pyrobaculum aerophilum sp. nov., a novel nitrate-reducing hyperthermophilic archaeum. Appl Environ Microbiol 59, 2918–2926. Wolin, E. A., Wolin, M. J. & Wolfe, R. S. (1963). Formation of methane by bacterial extracts. J Biol Chem 238, 2882–2888. Zillig, W., Stetter, K. O., Schafer, W., Jankovic, D., Wunderl, S., Holz, I. & Palm, P. (1981). Thermoproteales : a novel type of extremely thermoacidophilic anaerobic archaebacteria isolated from Icelandic solfataras. Zentbl Mikrobiol Parasitenkd Infektionskr Hyg Abt 1 Orig C 2, 205–227. Zillig, W., Stetter, K. O., Prangishvili, D., Schafer, W., Wunderl, S., Janekovic, D., Holz, I. & Palm, P. (1982). Desulfurococcaceae, the second family of the extremely thermophilic, anaerobic sulfurrespiring Thermoproteales. Zentbl Bakteriol Hyg 1 Abt Orig C 3, 304–317. Zillig, W., Holz, I., Janekovic, D., Schafer, W. & Reiter, W. D. (1983). The archaebacterium Thermococcus celer represents a novel genus within the thermophilic branch of archaebacteria. Syst Appl Microbiol 4, 88–94. Zillig, W., Holz, I., Janekovic, D. & 7 other authors (1990). Hyperthermus butilicus, a hyperthermophilic sulfur-reducing archaebacterium that ferments peptides. J Bacteriol 172, 3959–3965. International Journal of Systematic and Evolutionary Microbiology 50 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Sat, 17 Jun 2017 15:21:03