Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
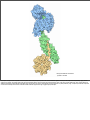
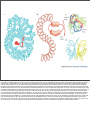
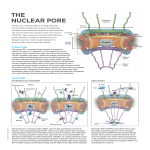
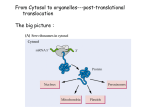
Nuclear Transport • 1000-10000 per cell Nuclear Pore • ~30 proteins (nucleoporins, nups), some in more copies • • ~125 MDa central channel ~25 nm Stewart, M. (2007). Molecular mechanism of the nuclear protein import cycle. Nat Rev Mol Cell Biol, 8, 195-208. André Hoelz and Günter Blobel, Nature 432, 815-816 Cargo to be imported contains a nuclear localization sequence (NLS). The alpha-kap protein is needed for cargo import, and is itself a cargo for export. a, In the cytoplasm, alpha-kap's NLSbinding site is blocked by its auto-NLS. This prevents a cargo's NLS from binding to alpha-kap. b, In the presence of an import beta-kap, the auto-NLS of alpha-kap binds to beta-kap, allowing alpha-kap to bind NLS–cargo (c). d, The ternary complex docks at nucleoporins (proteins of the nuclear pore complex, NPC) and enters the nucleus. e, The complex dissociates when RanGTP interacts with the amino-terminal arch of beta-kap, causing the release of alpha-kap's auto-NLS, which in turn displaces the cargo's NLS. f, The cargo is thus released. g, The import-beta-kap–RanGTP complex is exported. h, GTP is hydrolysed, producing RanGDP, which is released. Export of the alpha-kap, meanwhile, requires an export beta-kap. i, j, The export beta-kap forms a complex with RanGTP. k, This complex binds to alpha-kap and is exported. Matsuura and Stewart1 find that assembly of this ternary complex stores energy by twisting the export beta-kap from an S-shape into a horseshoe-like conformation, in which both arches embrace RanGTP. l, Arrival of the complex on the cytoplasmic face of the NPC triggers GTP hydrolysis. The complex releases the stored energy and pops open. The conformational change in beta-kap liberates alpha-kap and RanGDP. m, The alpha-kap enters a new round of nuclear import, whereas the export beta-kap may relocate to the nucleus for another export cycle. Frey, S. and Görlich, D. (2007). A saturated FG-repeat hydrogel can reproduce the permeability properties of nuclear pore complexes. Cell, 130, 512-23. Figure 1. Only a Saturated FG-Hydrogel Poses a Firm Barrier against Influx of acRedStar (A) A hydrogel containing 0.27 mM unlabelled and 1 μM Alexa 633-labeled Nsp1 fsFG-repeat domain was prepared, mounted to the stage of a confocal laser scanning microscope, and imaged after excitation at 633 nm. (B) Panels show influx of the 117 kDa acRedStar protein into the hydrogel from (A). acRedStar entered the gel rapidly (entry rate ≈5 μm/s). (C) Red points show profile of acRedStar concentration through the buffer-gel boundary at the 25 s time point. A slight delay in gel entry is indicated by the small concentration step at the buffer-hydrogel interphase. The black curve represents fits to error functions (see Experimental Procedures), from which diffusion constants in buffer and within the gel were derived. The data indicate that acRedStar diffused inside this low-concentrated FG-hydrogel not significantly slower than free in solution. See Figure S5 for computer simulations. (D) Right: Cartoon of a saturated FG-hydrogel, where all hydrophobic clusters have engaged into pairwise contacts. The simplest geometry with an orthogonal grid is shown. Since the maximal distance between adjacent clusters (u) is limited by the spacer length, such arrangement requires that the concentration of repeat units exceeds a certain threshold, the “saturation limit.” For details, see main text. Left: Cartoon depicts a gel with a lower concentration of repeat units than the saturation limit. Some hydrophobic clusters fail to find a sufficiently close partner for forming an interrepeat contact. Compared to saturated hydrogels, larger meshes result. The scheme depicts an “undersaturated” gel with defects of minimal size. Alternatively, such small defects could fuse to a larger one. This would make the gel less homogeneous (as observed for the low-concentrated gel in [A]) but would allow more hydrophobic clusters to engage into mutual contacts. (E) A hydrogel containing 2.2 mM of Nsp1 fsFG-repeat domain was formed and visualized as in (A). (F) Frames show influx of acRedStar into the hydrogel from (E). Only very small amounts entered the gel, even after a >7-fold longer incubation time as compared to the experiment shown in (A)–(C). (G) Red curve shows profile of acRedStar concentration at the 1800 s time point. Quantitation and fitting the data of all time points to diffusion models indicate that the influx of acRedStar into this saturated FG-hydrogel (k1≈1 nm/s) proceeded ≈5000-fold more slowly than into the undersaturated FG-hydrogel from (A)–(C). January 2007 Molecule of the Month by David S. Goodsell The actual mechanisms that importins use to pull molecules through the nuclear pore are still a subject of active debate, but PDB entry 2bpt gives some hints of how it might be done. The thousands of proteins that make up the nuclear pore are covered with special amino acid sequences that are flexible and that contain many phenylalanines. One side of importin-beta binds to these special sequences. The structure shown here includes the full importin-beta (in rainbow-colored cylinders) and a few short pieces from the nuclear pore proteins (shown in spheres at the bottom, with the phenylalanine amino acids in red). Notice that the phenylalanines bind in pockets on the outer surface of the importin. Importin-beta may jump from site to site through the nuclear pore, guided by these special sequences. As you take a look at these structures yourself, take a moment to explore the unusual fold of the chain. Importin-beta is folded like a spring, which is then wraps into a big spiral that traps its binding partners inside. André Hoelz and Günter Blobel, Nature 432, 815-816 Cargo to be imported contains a nuclear localization sequence (NLS). The alpha-kap protein is needed for cargo import, and is itself a cargo for export. a, In the cytoplasm, alpha-kap's NLSbinding site is blocked by its auto-NLS. This prevents a cargo's NLS from binding to alpha-kap. b, In the presence of an import beta-kap, the auto-NLS of alpha-kap binds to beta-kap, allowing alpha-kap to bind NLS–cargo (c). d, The ternary complex docks at nucleoporins (proteins of the nuclear pore complex, NPC) and enters the nucleus. e, The complex dissociates when RanGTP interacts with the amino-terminal arch of beta-kap, causing the release of alpha-kap's auto-NLS, which in turn displaces the cargo's NLS. f, The cargo is thus released. g, The import-beta-kap–RanGTP complex is exported. h, GTP is hydrolysed, producing RanGDP, which is released. Export of the alpha-kap, meanwhile, requires an export beta-kap. i, j, The export beta-kap forms a complex with RanGTP. k, This complex binds to alpha-kap and is exported. Matsuura and Stewart1 find that assembly of this ternary complex stores energy by twisting the export beta-kap from an S-shape into a horseshoe-like conformation, in which both arches embrace RanGTP. l, Arrival of the complex on the cytoplasmic face of the NPC triggers GTP hydrolysis. The complex releases the stored energy and pops open. The conformational change in beta-kap liberates alpha-kap and RanGDP. m, The alpha-kap enters a new round of nuclear import, whereas the export beta-kap may relocate to the nucleus for another export cycle. January 2007 Molecule of the Month by David S. Goodsell The importin complex shown here (built from three separate PDB entries) transports proteins with nuclear localization signals. At the top in blue is importin-beta (PDB entry 1qgk). It is the engine of the complex, recognizing nuclear pores and moving through them. It wraps around the end of importin-alpha, shown at the center in green (PDB entry 1ee5). Importin-alpha is an adapter molecule that connects importin-beta with the cargo. In this picture, the cargo is nucleoplasmin, shown at the bottom in yellow (PDB entry 1k5j), a chaperone protein that is important in nucleosome assembly. Notice how the extended nuclear localization signal of this cargo is gripped by importin-alpha. Stewart, M. (2007). Molecular mechanism of the nuclear protein import cycle. Nat Rev Mol Cell Biol, 8, 195-208. a | Cargoes for nuclear import have nuclear localization signals (NLSs). The karyopherin importin-alpha recognizes classic NLSs based on one (monopartite) or two (bipartite) clusters of basic residues that are shown in red. b | Structure of the nucleoplasmin NLS (red) bound to importin-alpha23. The 10 different Armadillo (ARM) repeats of importin-alpha are shown in different colours. The NLS-binding sites are located in a groove on the inner surface of importin-alpha: the major binding site spans ARM repeats 1–4, whereas the minor site spans ARM repeats 6–8. c | Schematic illustration of the interactions between the SV40 large-T antigen NLS (shown in red with each residue identified in black one-letter code) and importin-alpha that are crucial for molecular recognition23. The key residues in the binding site are referred to as P1–P5. The hydrophobic portions of the basic side chains of residues in sites P3 and P5 are sandwiched between the indole side chains of residues W273, W184 and W142 of importin-alpha (yellow rectangles), whereas their charged (+) nitrogens form salt bridges with acidic (blue) residues D270, E266 and E180 (-) of importin-alpha. Lys P2 is neutralized by D192 (refs 21–24). In addition, hydrogen bonds (dotted lines) are formed between Asp residues (N235, N188 and N146) and the NLS main-chain amides. d | Similar schematic showing the interactions between the nucleoplasmin NLS (shown in red) and importin-alpha, for which, in addition to the interactions at the major site (P2–P5), there are also similar interactions at the minor site (P'1–P'2). e | Schematic illustration of the autoinhibitory role of the importin-beta binding (IBB) domain. In the import complex, the IBB is bound to importin-beta (blue), but when released by RanGTP (cyan), it can compete with NLSs for binding to the importin-alpha ARM domain (green), leading to cargo release32. André Hoelz and Günter Blobel, Nature 432, 815-816 Cargo to be imported contains a nuclear localization sequence (NLS). The alpha-kap protein is needed for cargo import, and is itself a cargo for export. a, In the cytoplasm, alpha-kap's NLSbinding site is blocked by its auto-NLS. This prevents a cargo's NLS from binding to alpha-kap. b, In the presence of an import beta-kap, the auto-NLS of alpha-kap binds to beta-kap, allowing alpha-kap to bind NLS–cargo (c). d, The ternary complex docks at nucleoporins (proteins of the nuclear pore complex, NPC) and enters the nucleus. e, The complex dissociates when RanGTP interacts with the amino-terminal arch of beta-kap, causing the release of alpha-kap's auto-NLS, which in turn displaces the cargo's NLS. f, The cargo is thus released. g, The import-beta-kap–RanGTP complex is exported. h, GTP is hydrolysed, producing RanGDP, which is released. Export of the alpha-kap, meanwhile, requires an export beta-kap. i, j, The export beta-kap forms a complex with RanGTP. k, This complex binds to alpha-kap and is exported. Matsuura and Stewart1 find that assembly of this ternary complex stores energy by twisting the export beta-kap from an S-shape into a horseshoe-like conformation, in which both arches embrace RanGTP. l, Arrival of the complex on the cytoplasmic face of the NPC triggers GTP hydrolysis. The complex releases the stored energy and pops open. The conformational change in beta-kap liberates alpha-kap and RanGDP. m, The alpha-kap enters a new round of nuclear import, whereas the export beta-kap may relocate to the nucleus for another export cycle. importin-beta and RanGTP importin-alpha, CAS and RanGTP January 2007 Molecule of the Month by David S. Goodsell Once the importin-beta/importin-alpha/cargo complex gets inside the nucleus, the cargo must be released and the importins must be recycled back to the cytoplasm. The Ran protein, shown here in red, is responsible for releasing the cargo. It binds to importin-beta and causes a significant change in shape, leading to the release of importin-alpha and the cargo. Then, the complex of importin-beta and Ran (shown on the left, PDB entry 2bku) travels back through the pore. Outside, a GTP molecule in Ran (shown in bright red) is cleaved and the Ran dissociates, leaving importin-beta ready to carry the next cargo protein inside. Importin-alpha can't get back to the cytoplasm by itself, so it gets some help from CAS (a nuclear export factor), shown here on the right in purple (PDB entry 1wa5). CAS is similar to importin-beta, but moves through the nuclear pore in the opposite direction. It binds to importin-alpha and Ran and carries them out of the nucleus. Then, a similar cleavage of the GTP in Ran releases importin-alpha for another round of transport. Importin-beta56, 74, 75 (which is shown complexed with the importin-beta binding (IBB) domain of importin-alpha (red) in part a of the figure and in Supplementary information S2 (movie)) is constructed from 19 HEAT repeats, each of which is made up of two alpha-helices (A and B, part b). These HEAT repeats stack together to generate an elongated helicoidal molecule that is formed from two C-shaped arches56, 74, 75. The export carrier of importin-alpha, known as CAS (see Supplementary information S7, S8 (movies)) has a similar helicoidal configuration based on 19 HEAT repeats33, 79. The two alpha-helices (cylinders A and B) from which HEAT repeats are constructed function like the coils of a spring (part b) and impart considerable flexibility to importin-beta and CAS that is crucial for orchestrating their binding and release of partners. Ran (part c and Supplementary information S3 (movie)) has the Ras-family GTPase fold. Most of the Ran structure shows only marginal changes in confirmation between the GTP- (cyan) and GDP-bound (yellow) states, but two loops (the switch I and switch II loops) change conformation markedly depending on whether GDP (red) or GTP (blue) is bound33, 73, 74, 94, 95. The C terminus also changes with nucleotide state, although the conformation taken up in the GTP-bound state appears to vary depending on the partner to which Ran is bound. NTF2 (ref. 96) (part d and Supplementary information S11 (movie)) is constructed from two chains (cyan and yellow) and is the nuclear transport factor for RanGDP17, 18. NTF2 (ref. 96) is a dimer in which there is a hydrophobic cavity that binds RanGDP95 (Supplementary information S12 (movie)) and also a hydrophobic patch formed by residues from each chain at the opposite end of the molecule, which binds nucleoporin FXFG repeats41, 44, 95, 96. Ran guanine nucleotide-exchange factor (RanGEF) has a 7-bladed propeller structure97 (see Supplementary information S13 (movie)), and, by stabilizing the nucleotide-free state of Ran, increases the rate of nucleotide exchange by approx105. Ran GTPase activating protein (RanGAP)98 is based on a leucine-rich repeat (LLR) fold (see Supplementary information S14 (movie)) and increases the Ran GTPase activity by approx105 (ref. 99). Movies showing the structure of each molecule rotating can be accessed online (Supplementary information S1, S2, S3, S4, S5, S6, S7, S8, S9, S10, S11, S12, S13, S14 (movies)). Part b is reproduced with permission from Ref. 81 © (2003) American Association for the Advancement of Science. Stewart, M. (2007). Ratcheting mRNA out of the nucleus. Mol Cell, 25, 327-30. (i) A large mature mRNP (shown schematically as a blue line although it will have extensive secondary structure) probably has several Mex67:Mtr2 carrier complexes (red) attached along its length. Interactions between Mex67:Mtr2 and FG-nucleoporins (F) that line the NPC transport channel (gray) facilitate movement of the mRNP backward and forward by thermal motion (Brownian movement). (ii) When one of the Mex67:Mtr2 complexes reaches the cytoplasmic face of the NPC, it is removed from the mRNP by the DEAD-box helicase Dbp5, the ATPase activity of which is stimulated by Gle1 and IP6. (iii) Removal of Mex67:Mtr2 prevents this segment of the mRNP from moving back into the transport channel and so functions as a molecular ratchet. (iv) Brownian motion soon results in another Mex67:Mtr2 complex reaching the cytoplasmic face, where again it is removed by Gle1:IP6-stimulated Dbp5, and now (v) a longer segment of the mRNP is prevented from returning. By iteration of this cycle, even an extremely large mRNP can be ratcheted into the cytoplasm using the ATPase activity of Dbp5 to rectify thermal motion. The Mex67:Mtr2 liberated is recycled to the nucleus to participate in another mRNP export cycle. Although this mechanism envisages Mex67 release though the Gle1:IP6 stimulation of the Dbp5 ATPase, other mRNP-remodeling processes at the NPC cytoplasmic face could generate directionality in an analogous manner. André Hoelz and Günter Blobel, Nature 432, 815-816 Cargo to be imported contains a nuclear localization sequence (NLS). The alpha-kap protein is needed for cargo import, and is itself a cargo for export. a, In the cytoplasm, alpha-kap's NLSbinding site is blocked by its auto-NLS. This prevents a cargo's NLS from binding to alpha-kap. b, In the presence of an import beta-kap, the auto-NLS of alpha-kap binds to beta-kap, allowing alpha-kap to bind NLS–cargo (c). d, The ternary complex docks at nucleoporins (proteins of the nuclear pore complex, NPC) and enters the nucleus. e, The complex dissociates when RanGTP interacts with the amino-terminal arch of beta-kap, causing the release of alpha-kap's auto-NLS, which in turn displaces the cargo's NLS. f, The cargo is thus released. g, The import-beta-kap–RanGTP complex is exported. h, GTP is hydrolysed, producing RanGDP, which is released. Export of the alpha-kap, meanwhile, requires an export beta-kap. i, j, The export beta-kap forms a complex with RanGTP. k, This complex binds to alpha-kap and is exported. Matsuura and Stewart1 find that assembly of this ternary complex stores energy by twisting the export beta-kap from an S-shape into a horseshoe-like conformation, in which both arches embrace RanGTP. l, Arrival of the complex on the cytoplasmic face of the NPC triggers GTP hydrolysis. The complex releases the stored energy and pops open. The conformational change in beta-kap liberates alpha-kap and RanGDP. m, The alpha-kap enters a new round of nuclear import, whereas the export beta-kap may relocate to the nucleus for another export cycle. Stewart, M. (2007). Molecular mechanism of the nuclear protein import cycle. Nat Rev Mol Cell Biol, 8, 195-208. a | An import complex is formed in the cytoplasm between cargoes bearing nuclear localization signals (NLSs), importin-alpha and importin-beta. After passing through the nuclear pore complex (NPCs), the binding of RanGTP to importin-beta dissociates importin-beta from importin-alpha. The NLS-containing cargo is then displaced from importin-alpha and the importinalpha is recycled to the cytoplasm by its nuclear export factor, CAS, complexed with RanGTP. In the cytoplasm, RanGAP stimulates GTP hydrolysis, releasing the importins for another import cycle. Nucleoporins such as NUP50 catalyse cargo dissociation and function as molecular ratchets to prevent futile cycles. b | Ran also cycles between the nucleus and cytoplasm and, because the Ran guanine nucleotide-exchange factor (RanGEF) is nuclear and Ran GTPase activating protein (RanGAP) is cytoplasmic, this cycle is associated with changes in the nucleotide state. Therefore, cytoplasmic RanGDP is imported into the nucleus by nuclear transport factor-2 (NTF2), where RanGEF catalyses nucleotide exchange and generates RanGTP. RanGTP then binds to transport factors, such as importin-beta and CAS, and is exported to the cytoplasm, where RanGAP stimulates GTP hydrolysis. The structure of two loops in Ran (the switch I and II loops) changes markedly with nucleotide state (see Box 2 and Supplementary information S3 (movie)) and this change enables Ran to orchestrate the binding and release of its partners during the import cycle. • monopartite: PKKKRKV in SV40 large T antigen • bipartite: nucleoplasmin, KR[PAATKKAGQA] KKKK • others: of U snRNPs acidic M9 domain of hnRNP A1, the sequence KIPIK in yeast transcription repressor Matα2, and the complex signals