Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Observations on the rod and cone layer of
the human retina
A light and electron microscopic study
Ben S. Fine and Lorenz E. Zimmerman
Electron microscopy combined with certain histochemical studies on thin sections of the
human retina has provided additional information concerning the mucinous material that
occupies the intercellular space between the outer limiting membrane and the retinal pigment
epithelium. The myoid region of the inner segments of hotli rods and cones contains an
abundance of organelles associated with the intmcellular elaboration of mucinous and proteinaceous substances. It is suggested that this part of the photoreceptor cells is the most
likely source of the mucoid ground substance in which the inner and outer segments of the
visual cells are suspended. The organelles that are so concentrated in the apical region of the
photoreceptor cells are probably also involved in the production of intracellular materials
required for photoreception and/or generation and transmission of nerve impulses.
-L hotoreceptor cells ("rods" and "cones")
of the retina have been, studied in a number of animal species by electron microscopy.1"-'1 Attention has been directed
mainly toward the study of these cells,
with emphasis upon their outermost segments. Less attention has been directed toward their inner segments and to the
matrix (i.e., extracellular substance) in
which the rods and cones are embedded.
Few studies have been made on human
photoreceptor cells.21"27
Some of the methods of light microscopy'^"1" have been used to study the material that fills the intercellular space of the
rod and cone layer. This indistinct material
has not only been clearly demonstrated in
human and animal retinas, but has been
tentatively characterized as containing a
hyaluronidase-resistant acid mucopolysaccharide.31':"
In certain species, such as the rat and
rabbit, this interstitial matrix appears
smaller in quantity,'" and because this
space is occupied for the most part by the
processes of the pigment epithelial cells in
such species as the frog (Rana pipiens)
there has been some question as to
whether this mucoid material in the human
retina lies actually within or between the
cell processes.
This study, combining techniques of
light microscopy, histochemistry, and elec-
From the Ophthalmic Pathology Branch, Armed
Forces Institute of Pathology, Washington,
D. C.
This investigation was supported by Research
Grants 6X59-01-001 and DAMEDDH 61-51
from the Medical Research and Development
Command, United States Army, Washington 25,
D. C.
446
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Volume 2
Number 5
tron microscopy, was designed to seek
answers to the following questions:
1. Can the mucoid
(Hale-positive)
material be identified by electron microscopy in the human retina?
2. Is this material intracellular, extracellular, or both?
3. What are some of the probable
sources for production of this material?
Materials and methods
The materials and methods used have been202734 described in detail previously.' > 34 Illustrations
are all oriented with the rod and cone outer segments directed downward. The vitreous cavity is
above. All markers are 1 /», unless otherwise indicated.
Rod and cone layer of human retina
447
Observations
The descriptive observations are recorded as
legends to each figure. The figures are arranged
with a light micrograph, as in Fig. 1, for purposes
of orientation, followed by electron micrographs
showing regions of the perikaryon (Figs. 2 and
3). These are followed by a series of micrographs illustrating the external limiting membrane
and myoid portions of the photoceptor cells to 6)a
end of their inner segments (Figs. 4 to 6) and
finally including an adjacent portion of their
outer segments (Fig. 7).
Discussion
The photoreceptor cells of the retina include more than what is conventionally
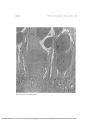
Fig. 1. A light micrograph of human retina taken from approximately the parafoveal region.
The section is celloidin embedded and stained with hematoxylin and eosin. The orientation is
such that the internal limiting membrane (ILM) is above and the external limiting membrane
(XLM) is below. The electron micrographs that follow are arranged to illustrate the region
from A down to B. The free arrow points to the zone of ONL, of outer nu clear segments
ofnuceli the photoreceptor cells. MLM, middle limiting membrane; ONL, ofthe nuclear layer (the
nuclei of the photoreceptor cells); CC, choriocapilla of the
ri
s )
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
448 Fine and Zimmerman
called the layer of rods and cones, since
these cells extend from the free inner
surface of the retinal pigment epithelium
to the inner part of the outer plexiform layer
(region of the middle limiting membrane27) (Fig. 1). In the various cellular
layers occupied by the photoreceptor cells,
only two other histologic elements are
present. Between the outer limiting membrane and the pigment epithelium there
is an intercellular substance that fills the
spaces between adjacent photoreceptor
cells. Inside the outer limiting membrane
the cytoplasm of Miiller's cells fills up all
potential space between the photoreceptors (Figs. 2 to 4). Miiller's cells and the
photoreceptors are joined most firmly at the
level of the outer limiting membrane, where
there are numerous terminal bars. Apparently the outer limiting membrane serves
as an effective barrier preventing the passage of the normal interstitial mucoid
matrix about the outer ends of the visual
cells directly into the retina proper, just as
in pathologic states it serves to prevent extension of exudates and hemorrhages out
into the subretinal area.
That part of the photoreceptor cells between the nucleus and the retinal pigment
epithelium is the free or apical half of the
cell (Fig. 2). As is true of less highly
specialized epithelia, the apical part of
these neuroepithelial cells near the nucleus
contains the Golgi complex, associated
secretory vesicles, ribonucleoprotein particles, and smooth-surfaced endoplasmic
reticulum. The apical cytoplasm of the
photoreceptors includes that part of the cell
Inoestigative Ophthalmology
October 1963
passing through the external limiting membrane, together with the inner segments of
the rod and cone layer. The outer segments
of these cells are considered as remarkably
specialized ciliated villous projections of
the apical cytoplasm.
The Golgi complex, generally considered
to play an important role in the secretory
activities of cells, is very prominent in the
apical part of the photoreceptors, where
there are also large numbers of associated
vesicles. Many of the larger vesicles contain
a finely granular or filamentous material
that is electron lucent (Figs. 3 to 5). This
material resembles closely that which occupies the intercellular spaces between the
outer limiting membrane and the pigment
epithelium (Figs. 5 and 7). These observations suggest the possibility that the apical
part of the photoreceptor cells (i.e., the
rod and cone inner segments) is the source
of most of the mucoid matrix in which the
free ends of these cells are enveloped.
The occurrence of a film of mucoid material within the lumen of well-differentiated
rosettes of retinoblastomas and in those
of dysplastic retinas31 provides additional
support for the belief that the visual
cells produce this mucoid ground substance.
The human rod and cone inner segments
have long been subdivided into a refractile
outer part, or ellipsoid, and a nonrefractile,
basophilic inner part, or myoid. This subdivision is more prominent in cones than in
rods.
The "ellipsoid" is filled with mitochondria,20 while the "myoid" (Fig. 3) contains
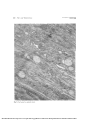
Fig. 2. This electron micrograph for orientation shows an area corresponding to region A of
the light micrograph. The nuclei (N) are those of the rod and cone photoreceptor cells,
which make up the outer nuclear layer (see Fig. 1, ONL). The external limiting membrane
(a series of terminal bars) is indicated by the free arrows toward the bottom of the micrograph. The very lucent cytoplasm of the Miiller cell (MC) can be seen occupying the
"spaces" between the photoreceptor cells internal to the external limiting membrane. The
basal cytoplasm of these photoreceptor cells is attenuated into rod axons (RA) and cone axons
(CA). The apical cytoplasm, which contains the large Golgi complex (G), extends outward
"through" the external limiting membrane as the photoreceptor inner segments. Here, between these inner segments, lie the narrow villous extensions of the Miiller cells, which cannot be seen at this low magnification but can be in Fig. 3 (MV). (Nasal macula, untreated.
x6,800.)
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Volume 2
Number 5
Rod and cone layer of human retina. 449
Fig. 2. For legend see opposite page.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
450
Investigative OiMbalnmlogy
Octoficr JS63
Fine and Zimmerman
ss
Fig. 3. For legend see page 453.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Rod and cone layer of human retina 451
Volume 2
Number .5
ro
ss
DE
SS
MV
Fig. 4. For legend see page 453.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
rxlifiatiirt' Ophthalmology
October 1963
452 Fine and Zimmerman
SS
SS
" " w v ? ^ -MP
• FR
MP ,
M
Fig. 5. For lopMid sec opposite page.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Rod and cone layer of human retina
the large Golgi complex, many of its associated vesicles, and considerable quantities
of both free and membrane-bound ribosomes. The granular endoplasmic reticulum
("ergastoplasm") shows little orientation.
Many segments of agranular endoplasmic
reticulum are also present, as are a few
electron-dense bodies.15
The term "myoid" is derived from a
similar, but contractile, portion of some
amphibian cones and is not a meaningful
term in relation to the human, for there is
as yet no evidence, either physiologic or
morphologic (including electron microscopic), for the properties of a muscle here.
The myoid region of the cone has, however, long been known to be basophilic,
a characteristic that is accounted for by
the densely congregated ribosomes. The
453
ribosomes (ribonucleoprotein particles)
produce most of the cytoplasmic basophilia30 and are involved in the synthesis
of certain proteins.37' HS For other reasons,"4
these particles and/or associated membrane
systems have been suspected of playing a
significant role in the synthesis of a variety
of mutinous materials, which are then collected and probably concentrated in the
Golgi complex,3""11 from which they are
eventually extruded from the cell (Fig. 4).
The similarity in appearance of the material within the Golgi complex here with
that occupying the extracellular spaces
does not imply chemical identity, especially
if the intracellular material is a precursor
material, the composition of which may be
undergoing change prior to discharge from
the cell.
Fig. 3. Higher magnification of the apical region of the cone photoreceptor, above, and two
rod photoreceptors, lower left portion of the micrograph. The terminal bars (TB) that form
the external limiting membrane are evident, and the lucent cytoplasm of the Miiller cell (MC)
is indicated between the photoreceptor cells. The apical part of the visual cell, which is
continuous with the myoid portion, contains a very large Golgi complex (G) with many
associated vesicles varying in size. Some of these larger vesicles contain a lucent, almost filamentous material (free arrows) that closely resembles the extracellular material observed
more distally (Figs. 4, 5, and 7). The cytoplasm here also contains large quantities of ribonucleoprotein particles (RNP), most of which lie "free" or in small clusters. Moderate numbers of these ribosomes are observed to be associated with short fragments of a doublemembrane system, which here shows little orientation. Considerable quantities of agranular
double membranes (smooth-surfaced endoplasmic reticulum) (SS) are also present. MV,
Miiller cell vilh; N, nucleus. (Nasal macula, uranyl acetate treated. x27,000.)
Fig. 4. Photoreceptor (rod) inner segments at the region of the external limiting membrane
(TB) and extending outward from the retina, separated here for the most part by the
delicate villous processes (MV) of the Miiller cells (MC). Large vesicles, rjresumably a part of
the Colgi complex, are present, filled with a lucent, finely granular to filamentous material
(free arrows). A small segment of agranular reticulum (V,) lies close to the surface membrane
of the cell. In another region the cell surface membrane protrudes inward (V), suggesting the
possibility of fusion here with an intracellular vesicle. Throughout the cytoplasm are large
aggregates of smooth-surfaced endoplasmic reticulum (SS), as well as small clusters of
granular endoplasmic reticulum (ER) and widespread groupings of free ribosomes (R). OB, •
dense body. (Nasal macula, uranyl acetate treated. x27,000.)
Fig. 5. Photoreceptor (rod) inner segments just external to the portions illustrated in Fig.
4. The villi of Miiller's cells protrude between these inner segments, and one can be seen
cut obliquely at MV. Beyond this villous process, lying in the space between adjacent cells,
is a lucent, finely granular to filamentous material (MP). This material occupies more space
as the cells are more widely separated distally (toward the lower right of the micrograph)Within these cells is an extensive Golgi complex system (G) together with several large
vesicles frequently observed to contain a finely granular to filamentous, lucent material (free
arrows) that resembles the material lying between the cells (MP). Many segments of granular
endoplasmic reticulum (ER), as well as many free clusters of ribosomes and agranular membranes (SSJ, are also present. CR, portion of a cross-banded ciliary rootlet; DB, dense body;
M, mitochondria. (Nasal macula, uranyl acetate treated. x25,200.)
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
•.stigalioe Ophthalmology
October .1963
454 Fine and Zimmerman
The presence of such large quantities of
ribosomes, membrane systems, and an
enormous Golgi complex in this "myoid
region" of the photoreceptor inner segment strongly points to the synthesis of
various mutinous and proteinaceous materials at this site within the cell. Such a
relationship has also been observed in
other actively secreting cells that produce
both mutinous and proteinaceous materials.'-"11 What portion of this synthetic
activity is related to production of the
interstitial mutinous materials, and what
portion to possible use within the cell for
purposes of photoreception and/or nerve
impulse generation and conduction, is not
at all clear. The relative paucity of highly
organized granular endoplasmic reticulum
and. the presence of large quantities of
"free" ribosomes also suggest considerable
synthetic activity at this site for local or
intracellular use.4Si 4"
The inner segments of rods and cones
contain similar components, but, in general, the materials in the rod inner segment
are more diffusely arranged,-'1 and the
division of inner segment into ellipsoid
and myoid regions is not as distinct, nor is
the histochemical staining as distinct as in
the cone inner segment (Fig. 6).
At the base of these photoreceptor inner
segments (just external to the outer limiting membrane), villi of the Muller cells
protrude into the extracellular spaces for a
distance approximately the length of the
myoid portions (Figs. 4 and 5). These villi
might contribute to the extracellular material, but evidence supporting such a possibility is, at present, lacking. Because of
their considerable morphologic similarity to
the apical microvilli of the brush border
of intestinal epithelium, or to those of the
actively absorptive cells of the proximal
convoluted tubule of the kidney nephron,17
one might consider the possibility of a
greater absorptive function here than secretory, although determination of their actual
function must await other methods of investigation.
Another possible source of some of the
interstitial mucoid material is the retinal
pigment epithelium, but these cells, like the
Muller cells, have not been found to have
the high degree of cytoplasmic organization that is generally associated with this
type of secretory activity.
The outer segments of both rods and
cones in man consist (as in other animals)
of finely laminated structures, in reality
layers of membranes and tubules. Both of
these laminated structures are intensely
periodic acid-Schiff positive'113 and are enveloped within an expansion of the cell
surface membrane that includes the projecting cilium. The limiting plasma membrane of the cylindrical rod outer segment
is very closely applied to its laminated
inner structure, whereas that of the cone
is usually more widely separated (Fig. 7).
This slight separation of the cell membrane
in the cone outer segment introduces the
possibility that a small amount of Halepositive material may occupy this region,
which cannot be adequately resolved by
light microscopic techniques.
The human rod and cone outer segments
are easily distinguished from one another
by a number of morphologic criteria, which
will be discussed in more detail elsewhere.
Conclusions
From these observations and interpretations one may conclude, therefore, that the
Hale-positive region of the rod and cons
layer in the human retina represents mainly
the extracellular material in which the
photoreceptor outer segments are embedded. This material contains an acid
mucopolysaccharide that is not sensitive to
either bovine testicular or streptococcal
hyaluronidase.31';|- The major portion of
this material is probably synthesized within the apical cytoplasm (inner segments)
of the photoreceptor cells and extruded
from there into the intercellular space.
From these observations and conclusions,
a further possibility appears that some of
the sites of the synthetic processes involved
in elaboration of these various materials
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Investigative Ophthalmology
Volume 2 Number 5
Fine and Zimmerman:
Rod and cone layer of human retina 455
Fig. 6. A, A 1 to 1.5 A thick section of osmium-fixed, methacrylate-embedded human retina
treated by a combination of the periodic acid-Schiff reaction and the Hale colloidal-iron
technique for acid mucopolysaccharides. The photoreceptor outer segments are clearly P.A.S.
positive, while the material between the cells is P.A.S. negative, but intensely Hale positive.
Note that within the inner segments, P.A.S. positivity is greater in the ellipsoid portion,
where most of the mitochondria are congregated in the cone cell, while the myoid portion,
where most of the Golgi complex is to be found, is somewhat Hale positive. The dark
granules at the bottom of the figure are the pigment granules of the retinal pigment epithelium. B, Several of the rod photoreceptor segments are seen in cross-section. They are intensely P.A.S. positive. The material that fills the space between the photoreceptor segments
is Hale positive and considered to contain an acid mucopolysaccharide. (Nasal retina,
periodic acid-Schiff, and Hale treated, x 1,440.)
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Volunw 2
Number 5
Rod and cone layer of human retina
Fig. 7. For legend see pa£e 458.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
457
458 Fine and Zimmerman
! Ophthalmology
October 1963
Fig. 7. Junction of outer and inner segments of a rod and a cone from the nasal macular
area of a 40-year-old man. Only the apexes of the receptor inner segments are seen here. The
cone inner segment contains a large number of mitochondria (M) as well as large numbers
of "free" ribosomes. The cone-connecting cilium, identical to that of the rod, is seen at
ClL. The cross striations of a cilium rootlet are seen at R. The laminations or discs of the
rod outer segment are clearly seen, as are the staggered groupings of the cut ends of tubules.
The cone outer segment is not cylindrical but tapered, somewhat like the cones of lower
animals. No cut ends of tubules resembling those of the rod are seen in the cone, and the
more closely arranged double lamellas of the cone in this particular section end along one
side as dilated tubules and vesicles. The lamellas themselves frequently present as loops (L)
at right angles to the longitudinal axis of the segment. The limiting plasma membrane (PL)
of the cone cell here is more widely separated from the lamellated internal structure than is
that of the rod cell. The large intercellular space is occupied by a lucent, finely granular
to filamentous material (MP) that corresponds to most of the Hale-positive region seen in
Fig. 6 (Nasal macula, uranyl acetate treated. x23,800.)
may also be concerned here with the production of some compounds necessary for
the processes of photoreception and/or
generation and propagation of the nerve
impulse in these special neuroepithelial
cells.
REFERENCES
1. Cohen, A. I.: The infrastructure of the rods
of the mouse retina, Am. ]. Anat. 107: 23,
1960.
2. Cohen, A. I.: The fine structure of the extrafoveal receptors of the rhesus monkey, Exper.
Eye Res. 1: 128, 1961.
3. Cohen, A. I.: The fine structure of the visual
receptors of the pigeon, Exper. Eye Res. 2:
88, 1963.
4. De Robertis, E.: Electron microscopic observations on the submicroscopic organization of the retinal rods, J. Biophys. &
Biochem. Cytol. 2: 319S 1956.
5. De Robertis, E., and Lasansky, A.: Submicroscopic organization of retinal cones of
the rabbit, j . Biophys. & Biochem. Cytol. 4:
743, 1958.
6. De Robertis, E.: Some observations on the
ultrastructure and morphogenesis of photoreceptors, J. Gen. Physiol. 43 (Suppl.): 1,
1960.
7. De Robertis, E., and Lasansky, A.: Ultrastructure and chemical organization of photoreceptors, in Smelser, G. K., editor: The
structure of the eye, New York, 1961,
Academic Press, Inc., pp. 29-49.
8. Eakin, R. M., and Westfall, J. A.: Fine structure of the retina in the reptilian third eye, ].
Biophys. & Biochem. Cytol. 6: 133, 1959.
9. Fernandez-Moran, H.: Fine structure of the
light receptors in the compound eyes of insects, Exper. Cell. Res. 5 (Suppl.): 586,
1958.
10. Fernandez-Moran, H.: The fine structure of
vertebrate and invertebrate photoreceptors
as revealed by low-temperature electron
microscopy, in Smelser, C. K., editor: The
structure of the eye, New York, 1961, Academic Press, Inc., pp. 521-556.
11. Lasansky, A., and De Robertis, E.: Electron
microscopy of retinal photoreceptors. The use
of chromation following formaldehyde fixation as a complementary technique to osmium
tetroxide fixation, J. Biophys. & Biochem.
Cytol. 7: 493, 1960.
12. Miller, \V. H.: Fine structure of some invertebrate photoreceptors, Ann. New York
Acad. Sc. 74: 204, 1958.
13. Moody, M. F., and Robertson, J. D.: The
fine structure of some retinal photoreceptors,
J. Biophys. & Biochem. Cytol. 7: 87, 1960.
14. Porter, K. R.: The submicroscopic morphology of protoplasm, Harvey Lect. 51: 175,
1957.
15. Sjostrand, F. S.: The ultrastructure of the
outer segments of rods and cones of the eye
as revealed by the electron microscope, J.
Cell. & Comp. Physiol. 42: 15, 1953.
16. Sjostrand, F. S.: The ultrastructure of the
retinal receptors of the vertebrate eye,
Ergebn. Biol. 21: 128, 1959.
17. Sjostrand, F. S.: Electron microscopy of the
retina, in Smelser, G. K., editor: The structure of the eye, New York, 1961, Academic
Press, Inc., pp. 1-28.
18. Villegas, G. M.: Electron microscopic study
of the vertebrate retina, J. Gen. Physiol. 43
(Suppl.): 15, 1960.
19. Wolken, J. ]., Capenos, J., and Turano, A.:
Photoreceptor structures. III. Drosophila
melanogaster, J. Biophys. & Biochem. Cytol.
3: 23, 1957.
20. Wolken, J. J.: Studies of photoreceptor structures, Ann. New York Acad. Sc. 74: 161,
1958.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017
Volume 2
Number 5
21. Wolken, J. J.: A structural model for a
retinal rod, in Smelser, G. K., editor: The
structure of the eye, New York, 1961,
Academic Press, Inc., pp. 173-192.
22. Yamada, E.: Observations on the fine structure of photoreceptive elements in the vertebrate eye, J. Electron Microscopy (Chiba)
9: 1, 1960.
23. Carasso, N.: Etude au microscope electronique des synapses des cellules visuelles
chez le t£tard d'Alytes obstetricans, Compt.
rend. Acad. sc. 245: 216, 1957.
24. Yamada, E., Tokuyasu, K., and Iwaki, S.:
The fine structure of retina studied with
electron microscope. III. Human retina, J.
Kurame M. A. 21: 1979, 1958.
25. Missotten, L.: Etude des batonnets de le
retine humaine au microscope electronique,
Ophthalmologica 140: 200, 1960.
26. Fine, B. S.: Limiting membranes of the
sensory retina and pigment epithelium, an
electron microscopic study, A. M. A. Arch.
Ophth. 66: 847, 1961.
27. Fine, B. S., and Zimmerman, L. E.: Miiller's
cells and the "middle limiting membrane" of
the human retina, INVEST. OPHTH. 1: 304,
1962.
28. Howard, A. D.: The visual cells in vertebrates chiefly in Necturus maculosus, J.
Morphol. 19: 561, 1908.
29. Willmer, E. N.: A physiological basis for
human colour vision in the central fovea,
Docum. Ophth. (Den Haag) 9: 235, 1955.
30. Sidman, R. L.: Histochemical studies on
photoreceptor cells, Ann. New York Acad.
Sc. 74: 182, 1958.
31. Zimmerman, L. E., and Eastham, A. B.:
Acid mucopolysaccharide in the retinal pigment epithelium and visual cell layer of the
developing mouse eye, Am. J. Ophth. 47:
488, 1959.
32. Zimmerman, L. E.: Acid mucopolysaccharides
in ocular histology and pathology, Proc. Inst.
Med. Chicago 23: 267, 1961.
33. Fine, B. S.: Unpublished observations.
34. Fine, B. S., and Zimmerman, L. E.: Light
and electron microscopic observations on the
ciliary epithelium in man and rhesus monkey,
INVEST OPHTH. 2: 105, 1963.
35. Fine, B. S.: Ganglion cells in the human
retina with particular reference to the macula
Rod and cone layer of human retina 459
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
lutea, an electron microscopic study, A. M.
A. Arch. Ophth. 69: 83, 1963.
Palade, G. E.: A small particulars component
of the cytoplasm, in Palay, S. L., editor:
Frontiers in cytology, New Haven, 1958, Yale
University Press.
Siekevitz, P., and Palade, G. E.: A cytochemical study on the pancreas of the guinea
pig. V. In vivo incorporation of leucine-l-C"
into the chymotrypsinogen of various cell
fractions, J. Biophys. & Biochem. Cytol. 7:
619, 1960.
Siekevitz, P., and Palade, C. E.: A cytochemical study on the pancreas of the guinea pig.
III. In vivo incorporation of leucine-1-C14
into the proteins of cell fractions, J. Biophys.
& Biochem. Cytol. 4: 557, 1958.
Palay, S. L.: The morphology of secretion,
in Palay, S. L., editor: Frontiers in cytology,
New Haven, 1958, Yale University Press.
Caro, L. C : Electron microscopic radioautography of thin sections: The Golgi zone
as a site of protein concentration in pancreatic acinar cells, J. Biophys. & Biochem.
Cytol. 10: 37, 1961.
Dalton, A. J.: Golgi apparatus and secretion
granules, in Brachet, J., and Mirsky, A. E.,
editors: The cell: biochemistry, physiology,
morphology, New York, 1961, Academic
Press, Inc., vol. 2, pp. 603-619.
Helander, H. F.: Ultrastnicture of fundus
glands of the mouse gastric mucosa, J. Ultrastruct. Res. 4 (Suppl.): 1962.
Ekholm, R., Zelander, T., and Edlund, Y.:
The ultrastructural organization of the rat
exocrine pancreas. I. Acinar cells, J. Ultrastruct. Res. 7: 61, 1962.
Scott, B. L., and Pease, D. C : Electron
microscopy of the salivary and lacrimal glands
of the rat, Am. J. Anat. 104: 115, 1959.
Munger, B. L.: A phase and electron microscopic study of cellular differentiation in
pancreatic acinar cells of the mouse, Am. J.
Anat. 103: 1, 1958.
Ham, A. W.: Histology, Philadelphia, 1957,
J. B. Lippincott Company.
Rhodin, J.: Anatomy of kidney tubules, Internat. Rev. Cytol. 7: 485, 1958.
Manual of histologic and special staining
technics, ed. 2, New York, 1960, Blakiston
Division, McGraw-Hill Book Company, Inc.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933144/ on 06/17/2017