Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
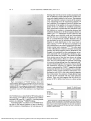
Investigative Ophthalmology & Visual Science, Vol. 32, No. 6, May 1991 Copyright © Association for Research in Vision and Ophthalmology Localization and Characterization of Substance P Binding Sites in Rat and Rabbit Eyes Philippe Denis,*t Veronique Fardin4 Jean-Philippe Nordmann,t Pierre-Paul Elena,§ Laurent Laroche,-(- Henri Saraux,t and William Rostene* Specific and high-affinity binding sites for Substance P (SP) were found in eyes from albino rabbits and rats using an in vitro autoradiographic method with l2SI-Bolton Hunter SP (BHSP). Autoradiograms were generated by apposing 10-20/im-thick cryostat eye sections to 3H-Hyperfilm or liquid emulsion and quantified by means of image-analysis procedures. Kinetic studies showed that equilibrium was reached after a 75-min incubation at room temperature. In rat retina, specific binding corresponding to approximately 90% of total binding, was reversible, of high affinity (dissociation constant [Kd], 0.13 ± 0.02 nM). Half-time for dissociation of 125I-BHSP was about 15 min. I) n la be led SP and the two neurokinins (NK) A and B competed in a concentration-dependent manner for retinal sites labeled by 125 I-BHSP with the following order of potencies: SP > NKA > NKB, in agreement with a pharmacologic profile of a SP receptor site. In both species, specific binding was found in the iris sphincter muscle, choroid, and retina. In rats, detectable amounts of SP-binding sites were also expressed in the corneal epithelium and iridial stroma. Quantitative analysis of the autoradiograms revealed that the highest densities of 125I-BHSP binding sites were localized in the iris sphincter muscle in rabbits and the inner retina in rats. Invest Ophthalmol Vis Sci 32:1894-1902,1991 such as calcitonin gene-related peptide (CGRP) or cholecystokinin, have also been identified in ocular sensory structures8'9 and shown to play a functional role in the neurogenic inflammation (particularly blood-aqueous barrier breakdown for CGRP.l0) However SP involvement in neurogenic inflammation is suggested by several pieces of evidence. First, nerve endings with immunoreactivity to SP are found in the uvea of several species, including humans," mainly in close association with the sphincter muscle of the iris and the smooth blood vasculature in the ciliary body. Second intracameral administration of SP induces a dose-dependent, nonmuscarinic pupil constriction associated with aqueousflareand an increase in intraocular pressure in the rabbit eye.6 Third electrical stimulation of the trigeminal ganglion or intracameral administration of capsaicin (both responsible for SP release in the anterior segment) are able to mimic SP-induced miosis.61213 Fourth, (D-Pro2, D-Trp7-9)SP, a SP antagonist, counteracts this phenomenon in rabbits and could therefore inhibit the ocular inflammatory response to laser iridial burns.14 Biochemical and immunohistochemical studies have localized SP in various vertebrate retinas.15 Cellular expression of SP-encoding mRNA was found recently in the rat retina using RNA blot and in situ hybridization.16 Although physiologic studies indi- Numerous neurogenic mediators released in the anterior segment of the eye by ocular injury, trauma, or noxious stimulation are known to elicit inflammatory effects, such as conjunctival hyperemia, miosis, rise in intraocular pressure, and disruption of the bloodaqueous barrier.' If some of these biologic effects are demonstrated to be mediated directly by metabolites from the arachidonic acid cascade (released from the iris and the ciliary body),2 it is now widely recognized that neural pathways also participate in the initiation of such inflammatory events by releasing neuropeptides from sensory afferent nerves in the uveal tract.3 Substance P (SP), an undecapeptide isolated from intestine in 1931,4 was first proposed as a neurogenic mediator of antidromic vasodilation and plasma extravasation at the peripheral level5 and as a major component in the neurogenic ocular injury responses.6'7 Later, other biologically active substances, From the *1NSERM U55 and tDepartment of Ophthalmology, Hopital Saint Antoine, Paris, JRhone-Poulenc Sante, Department of Biology, Vitry-Sur-Seine, and the §Department of Pharmacology, Faculte de Medecine, Nice, France. Philippe Denis was a recipient of INSERM (Poste d'accueil). Submitted for publication: September 25, 1990; accepted . Reprint requests: Philippe Denis, INSERM U55, 184, rue du Faubourg Saint Antoine, 75012, Paris, France. 1894 Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 OCULAR SUBSTANCE P BINDING SITES / Denis er al No. 6 cate that SP has a neuromodulator action on ganglion cells in fish17 and in dopamine release from the retina in the rat,18 the role of SP immunoreactive neurons in the processing of visual information is not yet fully understood. The presence of ocular SP binding sites was suggested previously by conventional binding techniques using membranes obtained from rat and bovine retina1920 or bovine and rabbit iris,2122 but the precise distribution of these receptor sites has not yet been investigated extensively. One autoradiographic report briefly mentioned the presence of binding sites in the rat retina,23 but no quantitative data or information on the pharmacologic profile of SP binding were presented. It is important to localize these binding sites since it is believed that most SP biologic actions are receptor -mediated.24 We therefore characterized and determined the anatomic localization of SP binding sites in rat and rabbit eyes using quantitative in vitro autoradiographic methods. We have used BoltonHunter SP (125I-BHSP; Amersham, les Ulis, France), a radiolabeled analogue of the tachykinin which has been extensively used in other organs to label SP receptors.25"27 Materials and Methods Tissue Preparation New Zealand albino rabbits (weighing 3-3.5 kg) were killed by injection of a lethal dose of sodium pentobarbital and Wistar rats (weighing 200-250 g), by decapitation. The eyes were removed, immersed in Tissue Tek medium (Miles Scientific, Naperville, IL), frozen in isopentane cooled (-40°C) in liquid nitrogen, and stored at -80°C. Just before sectioning, the tissues were warmed to -20°C, and sections (20-fxm thick) were cut with a cryostat (Bright), thawmounted onto gelatin-coated glass slides, and stored at -80°C until use. Before incubations, sections were allowed to thaw at room temperature. All investigations described in this paper were done in accordance with the ARVO Resolution on the Use of Animals in Research. Binding Conditions Preliminary experiments showed that preincubation was found to increase specific binding. Thus, slides were washed in a preincubation medium (Tris -HC1 50 mM, pH 7.4, containing 0.2 g/1 of bovine serum albumin) at room temperature for 15 min before incubation with radioligand. After the washing step, the slides were then incubated at room temperature in a solution of 65 pM 125I-BHSP (2000 Ci/ mmol) [Bolton-Hunter is 3-(p-hydroxy-m-(125I)iodophenyl)-propionyl) in 50 mM Tris -HC1, pH 7.4, con- 1895 taining 10 mM MgCl2,2 g/1 of bovine serum albumin, 40 mg/1 of bacitracin, 5 mg/1 of leupeptin, and 4 mg/1 of bestatin. Nonspecific binding was determined on alternate sections in the presence of 10 ju.M unlabeled peptide (SP) added in the incubation medium. The specificity of the binding was studied by incubating sections with increasing concentrations of peptides related to SP, such as senktide, septide, spantide, SP methylester, neurokinin A (NKA), neurokinin B (NKB), and synthetic fragments of SP (SP,_4 and SP4_n). Bacitracin, leupeptin, and bestatin were obtained from Sigma (St. Louis, MO) and SP-related peptides from Bachem (Bubendorf, Switzerland). After incubation with the radioligand, the sections were rinsed four times for 1 min each in the preincubation buffer at 4°C, dipped for 20 sec into distilled water, and quickly dried using a stream of cold air for autoradiography. Autoradiographic Experiments The sections were preincubated as described and incubated with 65 pM 125I-BHSP for 90 min at room temperature. After washing, the dry slides were stored in a Kodak X-ray cassette (Rochester, NY) in tight apposition to a tritium-sensitive film (3H-Hyperfilm; Amersham) and allowed to expose for 1 week in darkness. After exposure, the films were developed in Kodak D19 for 3 min and fixed. To identify the localization of the binding sites, the eye sections were counterstained with hematoxylin and eosin and cover slipped with Fluka (Chemika, Buchs, Switzerland). To investigate precisely the cellular localization of SP binding sites, an autoradiographic technique was used at light microscopic resolution. Briefly, after incubation and washings, selected slides were treated with a 30-min bath of 4% glutaraldehyde at 4°C to fix covalently the radioligand to its binding site, defatted in several baths of increasing concentrations of alcohol (75-100%) and xylene, and then dipped into liquid nuclear emulsion (LM1,; Amersham). Preliminary experiments showed that this treatment did not significantly alter 125I-BHSP binding. After an exposure period of 10 days in darkness, the emulsion dipped microautoradiograms were developed and fixed as described. Corresponding sections were counterstained and examined under a light microscope. Light- and dark-field photomicrographs were taken from the stained sections and silver grains, respectively. Data Analysis Film macroautoradiograms were analyzed by computer-based densitometry. The optical density of the autoradiogram was quantified by means of an image Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 1896 Vol. 32 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / May 1991 analyzer (BIOCOM RAG 200, les Ulis, France). Briefly, autoradiograms were digitized, and each grain density was assigned a relative optical density value. For maximal binding capacity determination, these relative optical density values were then converted to corresponding commercial 125I Amersham standards, and the results were expressed in fmol/mg tissue equivalent. Amersham's microscales are supplied as slices of several layers of polymer containing a range of increasing 1251 concentrations. Since tissue sections are coexposed with iodinated plastic standards, we can do quantitative densitometry using a computer-assisted system. Tissue-equivalent values were provided based on calibration using intact brain gray matter. In all cases, specific binding was defined by subtracting nonspecific binding (obtained in the presence of 10 iiM of unlabeled SP) from total binding. Classic computer analysis was used for the biochemical determination of the binding parameters (dissociation constant [Kd] and concentrations which inhibit 50% of the specific binding [IC50]) on rat retina sections. The kinetics of I25I-BHSP binding was assumed to follow a bimolecular association model (secondorder kinetics). Results Characterization of 125 I-BHSP Binding Kinetic analysis done by densitometry on rat retinal sections indicated that the association of 125I8O - 5O TOO 15O Time(min) Fig. 1. Association and dissociation of I25I-BHSP binding to rat retina sections. Retina sections were incubated with 65 pM 125IBHSP at room temperature for various times. The curve represents specific binding that was calculated as the difference between binding in the presence and absence of 10-^M unlabeled substance P (SP). Dissociation was initiated by the addition of 10-juM unlabeled SP at 120 min, a time at which the association of the ligand had reached the equilibrium. Values are given as the mean of triplicate determinations in a typical experiment. o 00 00 11 10 9 8 SP concentration 7 6 Log.M Fig. 2. Competition between 125I-BHSP and unlabeled SP for binding to rat retina sections. 65 pM 125I-BHSP were incubated alone or in the presence of increasing concentrations of unlabeled peptide SP at room temperature for 90 min. 125I-BHSP binding is expressed as a percent of total binding. Values are given as the mean of triplicate determinations. BHSP resulted in a time-dependent increase in binding (Fig. 1). At room temperature, specific binding reached a plateau in approximately 75 min; nonspecific binding was not significantly increased. About 90% of total binding was specific at 75 min. A routine incubation time of 90 min was then adopted for all subsequent experiments. At equilibrium, bound 125IBHSP could be dissociated specifically from its binding sites by incubating sections with an excess (10 ixM) of unlabeled peptide (Fig. 1). The half-time for dissociation of 125I-BHSP was about 15 min. We investigated competition studies between iodinated SP and increasing concentrations of unlabeled peptide (Fig. 2). These results indicated that SP inhibited 125I-BHSP competitively and with very high affinity. In the range of concentrations studied (10" u to 10~6 M), IC50 values were 0.17 nM. Curve analysis obtained (obtained by regression lines computerized by means of the least-squares method) indicated that 125 I-BHSP bound to high-affinity binding sites in rat retina with an apparent Kd estimated at 0.13 ± 0.02 nM. Competition studies with the tachykinins NKA, NKB, and analogues or fragments of SP were done in attempt to establish the type of site identified by I25IBHSP in the rat retina. The order of potency for inhibition of binding was SP > NKA > NKB (Fig. 3, Table 1). A similar ranking was obtained when rabbit eye sections were incubated with the tachykinins at concentrations in the range of 10"8 to 10~5 M (data not shown). Displacement curves showed that specific Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 1897 OCULAR 5UDSTANCE P BINDING SITES / Denis er al No. 6 .SP ASP 4-11 • NKA 0SP l-4 ANKB 11 10 9 8 7 6 Peptide concentration -Log M Fig. 3. Pharmacology of 125I-BHSP binding. 65 pM 125I-BHSP were incubated alone or in the presence of increasing concentrations of unlabeled peptides at room temperature for 90 min. I25IBHSP binding is expressed as a percent of initial binding. Values are given as the mean of four determinations. binding of I25I-BHSP was strongly inhibited by SP,_n (IC50=, 0.17 ± 0.02) (Fig. 2) and to a lesser extent by SP4_n and NKA (IC50=, 3.39 and 8.04 nM, respectively). On the other hand, senktide and septide was ineffective, and S P ^ , NKB, and spantide had some potency to inhibit retinal I25I-BHSP binding (Table 1). Autoradiographic Experiments Representative autoradiograms of 125I-BHSP binding sites are shown in Figure 4-6. In the rat anterior segment, SP binding sites were found in the iris, mainly in the sphincter region (Fig. 4, Panel a). Analysis of histologic sections confirmed that labeling on the pupil margin coincided with the iris muscular tissue. The density of silver grains was reduced dramatically in the presence of 10 IJM unlabeled SP (Fig. 4, Panel b), indicating specific labeling in that structure. In contrast, moderate 125I-BHSP binding densities were noted over the other iridial structures and in the corneal epithelium. No labeling occurred in structures such as the sclera and the ciliary processes. Binding seen on the lens fibers was not considered to be specific since SP did not displace 125I-BHSP labeling. In the posterior segment, the retina showed intensive labeling; low concentrations of binding were noted in the choroid. Binding sites were not uniformly distributed throughout the retina since a slight decrease of binding at the ora serrata was observed. Autoradiograms of the emulsion-dipped slides provided detailed histologic resolution of the retinal SP binding sites (Fig. 5). Silver grains were localized primarily in the inner plexiform (IPL) and ganglion cell layers (GCL). A few scattered grains were also detected in the outer plexiform layer and proximal inner nuclear layer (INL). Much lower SP binding was present in other regions, particularly in the outer retina (outer nuclear layer and photoreceptor cell layer). Similarly, very low, but significant, concentrations of SP binding sites were seen in the choroidal tissue. Some scleral blood vessels also exhibited some binding activity (Fig. 5, see arrows). In the rabbit eye (Fig. 6), the topographic distribution of SP binding sites was similar to that found in the rat although there were some quantitative differences (Table 2). Very high labeling was seen in association with the iridial sphincter muscle; no labeling was found either in the cornea, iris stroma, or ciliary body. Low binding was also identified in the choroid. As in the rat, lens fibers gave important nonspecific binding, ie, not displaceable in the presence of an excess of unlabeled peptide. The retina also had high densities of specific SP binding sites, mainly concentrated in the IPL and GCL, as described in the rat. Discussion These results demonstrate that several structures from albino rat and rabbit eyes (mainly the retina and the iris sphincter muscle) have the capacity to bind specifically a derivative analogue of SP, I25I-BHSP. The characteristics of I25I-BHSP binding in the rat retina strongly suggest that the ligand binds to SP receptors. The apparent Kd of the high-affinity (0.13 nM) binding site identified in this study was similar to those reported in rat retina homogenates (0.2 nM),19 in rabbit optic sections (0.54 nM),28 and in human retina (0.27 nM).29 Recent studies have described the existence of multiple tachykinin receptors in the rat retina.23 The agent, SP, belongs to a group of closely Table 1. Peptides ICso (nM) Substance P Substance P^,, Neurokinin A Substance P,_4 Neurokinin B Spantide Substance P methyl ester Septide Senktide 0.17 ±0.02 3.39 ± 0.40 8.04 ± 0.57 35 ±4.34 52.8 ± 4.06 53 ± 1.53 54 ± 6.34 >1000 >1000 Displacement of 125I-BHSP binding on rat retina sections by substance P (SP), peptides related to SP, and synthetic fragments of SP. Sections were incubated for 90 min at room temperature with 65 pM I251-BHSP and increasing concentrations (10"" to 10~6 M) of unlabeled peptides. Nonspecific binding of the radioligand was determined in the presence of 10-MM unlabeled SP. IC50 is defined as the molar concentration of the tested drug that displaced 50% of the specific binding of I25I-BHSP on retinae sections. Autoradiograms were analyzed by computer-based densitometry with an image analyser. Values are given as the mean of triplicate determinations. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / May 1991 1898 ISM /|SM the mammalian tachykinins have been described, namely SP (or NK-1) receptors, found in both central and peripheral tissues, NKA (or NK-2) receptors in peripheral tissues, and NKB (or NK-3) receptors mainly detectable in the central nervous system.23'32 Our data suggest that 125I-BHSP binds preferentially to NK-1 receptor sites; SP is the most potent tachykinin in inhibiting 25I-BHSP binding and the order of potency of the competitors for 125I-BHSP binding was SP > NKA > NKB. The IC50 values calculated for NKA and NKB were somewhat lower than those found for the displacement of I25I-BHSP binding in a preliminary report.19 Variations in the methods, including the incubation buffer and the lower ligand concentration used in our study (65 pM), may be the cause of these differences. It is unlikely that I25I-BHSP binds to NKB receptors since senktide, a highly selective NK-3 agonist, showed no activity at the micromolar level in displacing 125I-BHSP from its binding Fig. 4. Autoradiograms of I25I-BHSP binding in albino rat eye. Sections were incubated for 90 min at room temperature with 65 pM I251-BHSP alone (A) or in presence of 10-^M unlabeled SP to determine nonspecific binding (B). Autoradiograms were generated by apposition to 3H-Hyperfilm for 7 days. (C) Histological section, (c, cornea; ism, iris sphincter muscle; cp, ciliary processes; 1, lens; ch, choroid; r, retina). Magnification X3. related peptides known as tachykinins, all of which contain a common C-terminal amino acid sequence: Phe-X-Gly-Leu-Met-NH2.30 In 1983, two additional tachykinins, NKA and NKB, were isolated in mammalian tissues.31 To date, three distinct receptors for Vol. 02 B Fig. 5. Microautoradiograms of I25I-BHSP binding in albino rat posterior segment. Dark field (A) and bright field (B) were obtained after incubation of 65 pM '"l-BHSP alone. Silver grains visualized in the retina and choroid indicated specific SP binding sites since labeling disappeared after incubation with an excess of unlabeled peptide (not shown). GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; SV, scleral vessel. Magnification X500. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 No. 6 OCULAR SUBSTANCE P BINDING SITES / Denis er ol ISM B Fig. 6. Autoradiograms of I25I-BHSP binding in albino rabbit anterior segment. Sections were incubated for 90 min at room temperature with 65 pM I25I-BHSP alone (A) or in presence of 10-^M unlabeled SP (B). Auloradiograms were generated by apposition to 3 H-Hyperfilm for 7 days. (C) Histological section, C, cornea; ISM, iris sphincter muscle; CP, ciliary processes; L, lens. Magnification X3, binding sites were found to be mainly associated with the iris sphincter muscle although other iridial structures were slightly labeled in the rat eye. The presence of SP was reported in sensory neurons of the anterior uvea originating from the trigeminal ganglion, 3334 and a number of investigators extensively studied the involvement of the peptide in the acute irritation response of the eye. Besides its vasodilator properties, early experiments reported that the tachykinin peptide induces an increase in intraocular pressure and moderate alterations in the blood-aqueous barrier in rabbit eyes.6'714'35 Subsequent studies observed that these effects were relatively weak and inconsistent and that the rise in intraocular pressure was mainly due to a miosis-induced pupillary blockade since it was abolished in part by peripheral iridectomies. 12 Despite its wide distribution in the uveal tract in many species,11'33'34'36'37 SP has been proposed to be only responsible for the miotic component of the antidromic ocular injury response.12'3538 The other parts of the inflammatory response (elevated intraocular pressure and blood-aqueous barrier breakdown) are thought to be mediated by other mechanisms; in animals pretreated with prostaglandin inhibitors, injection of SP caused only miosis.12 The release of CGRP from sensory nerves has also been shown to induce marked inflammatory effects in cat39 and rabbit 40 eyes. The two neuropeptides are partially colocalized in terminal endings arising from the trigeminal cells41 and are released together into aqueous humor during the antidromic response. In addition, SP-induced miosis is potentiated by CGRP, 42 suggesting biologic interactions between the two peptides. Our autoradiographic results provide essential information concerning the role of SP in the ocular anterior segment; they demonstrate the presence of high-affinity binding sites for SP associated with the iridial sphincter muscle. They may also suggest that the miotic response to SP in rabbits may be due to a direct action Table 2. Specific l2iI-BHSP bound (fmol/mg tissue equivalent) Area Cornea Iris stroma Iris sphincter muscle site. Furthermore, as described for SP binding sites in other tissues, l9'25>26 affinity is encoded in the carboxy terminal of SP since SP,_, j and SP4_n had the greatest potency in inhibiting I25I-BHSP binding. Autoradiographic visualization of 125I-BHSP binding sites in the anterior segment of the eye shows a selective pattern of distribution. In both species, SP 1899 rat rat rabbit rat Retina rabbit rat 0.12 0.11 2.93 0.17 0.95 0.63 ±0.01 ±0.01 ± 0.42 ±0.01 ±0.17 ± 0.07 Regional distribution of specific BHSP binding sites in the albino rat and rabbit eye. Results were obtained by transformation of optical densities as described in the text to fmol/mg tissue equivalent ± SEM and were obtained from at least 10 determinations throughout the area under examination. Nonspecific binding was subtracted from each total value, so that data are expressed as specific binding. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 1900 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / May 1991 on the iris muscle through specific receptors. No binding sites was found in dilator or ciliary muscles, corroborating the fact that SP has no contractile effect on these tissues.3843 One of the most interesting findings of our study was the absence of high-affinity binding sites in the rabbit iris and ciliary epithelia, known to be the major site of the blood-aqueous barrier. It is unlikely that the lack of binding sites in rabbit ciliary body may be due to occupied receptor sites since sections were washed in a preincubation medium for 15 min before incubation with 125I-BHSP. Nevertheless, we cannot exclude the possibility of the presence of lower-affinity receptors or other unknown subtypes in ciliary process. Such sites may be absent or not detectable by our autoradiographic procedures. However, these results suggest that, in the rabbit, the peptide is not directly involved in the blood-aqueous barrier disruption observed in ocular neurogenic inflammation, as suggested by its weak and inconsistent effect. In addition, we cannot exclude the possibility that SP exerts some inflammatory actions in the eye through an indirect mechanism; it has been demonstrated that SP has a wide spectrum of inflammatory properties, in particular mast cell degranulation with release of histamine44 and arachidonic acid45 (and consequently prostaglandin biosynthesis). Finally, the involvement of SP in ocular neurogenic inflammation cannot be generalized since it has been reported that the ocular response to SP was species -dependent (cat, baboon, or human iris are relatively insensitive to SP46). This species variation is well illustrated in the our study by the fact that the overall density of iridial SP binding sites was higher in the rabbit than in the rat. The distribution of uveal SP binding sites and SP nerve terminals was not correlated well. In our study, regions that have relatively high densities of SP binding sites, such as the iris sphincter muscle, have been reported previously to have the highest amounts of SP immunoreactivity.47 In contrast, regions such as the ciliary processes or the iridial stroma, also known to contain SP immunoreactivity, did not have detectable SP binding sites. The existence of discrepancies between the localization of receptors and the distribution of fibers or neurotransmitters has been clearly demonstrated for SP in the brain.48 The SP binding activity in the scleral vessels (Fig. 5) may be associated with the endothelium or vascular constituents. This may be attributed to the chemotactic peptide, f-Met-Leu-Phe (fMLP)49 or the phagocytosis promoting peptide, tuftsin,50 which share structural similarities with SP. Furthermore, SP has been shown to interact with the tuftsin receptors present on the macrophage and polymorphonuclear leukocyte plasma membrane.50 However, under our experimen- Vol. 32 tal conditions, competition studies with increasing concentrations of tuftsin (10~4 to 10~7 M) showed that 125 I-BHSP binding was not inhibited by tuftsin (data not shown). Although SP-containing neurons have been identified in the retina of mammals,51 including humans,52 very little is known about their functional role in this tissue. These neurons are present in most species in amacrine cells whose somata are located at the border of the INL and IPL (which send processes primarily to the IPL). Evidence for SP localization to ganglion cells has also been demonstrated in some species, particularly rabbits.53 The localization of SP binding sites we found was similar to the pattern of SP-immunoreactive cells previously described for this region. Neonatal monosodium glutamate treatment of rats (which induces degeneration of the inner layers of the retina) has been reported to cause a marked reduction in 125I-BHSP retinal binding.19 Our autoradiographic findings also agreed with this experiment since most of the SP binding sites identified our study were concentrated in the IPL and GCL. Several observations suggest a neuromodulator role for SP in retinal functions; it has been found to elicit 3H-dopamine release from rabbit retinas18 and to exert excitatory effects on carp cholinergic-sensitive ganglion cells activity.17 It has been also shown to stimulate the accumulation of inositol triphosphates in rabbit retinal cultures.54 The presence of SP binding sites in the inner retina is an additional indication that the peptide probably has some regulatory action in visual processing. Key words: substance P, substance P receptor, autoradiography, eye Acknowledgments The authors thank Odile Flamand and Yves Issoulie for technical assistance. References 1. Duke-Elder PM and Duke-Elder WS: The vascular response of the eye. Proc R Soc Lond [Biol] 109:19, 1931. 2. Cole DF and Unger WG: Prostaglandins as mediators for the response of the eye to trauma. Exp Eye Res 17:357, 1973. 3. Unger WG and Butler JM: Neuropeptides in the uveal tract. Eye 2:202, 1988. 4. Von Euler US and Gaddum JH: An unidentified depressor substance in certain tissue extracts. J Physiol (Lond) 72:74, 1931. 5. Lembeck F and Holzer P: Substance P as neurogenic mediator of antidromic vasodilation and neurogenic plasma extravasation. Naunyn Schmiedebergs Arch Pharmacol 310:175, 1979. 6. Bill A, Stjernschantz J, Mandahl, Brodin E, and Nilsson G: Substance P: Release on trigeminal nerve stimulation, effects in the eye. Acta Physiol Scand 106:371, 1979. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 No. 6 OCULAR SUBSTANCE P BINDING SITES / Denis er ol 7. Nishiyama A, Masuda K, and Mochizuki M: Ocular effects of substance P. Jpn J Ophthalmol 25:362, 1981. 8. Stone RA, Kuwayama Y, Laties AM, McGlinn AM, and Schmidt ML. Guinea-pig ocular nerves contain a peptide of the cholecystokinin/gastrin family. Exp Eye Res 39:287, 1984. 9. Stone RA and McGlinn AM: Calcitonin gene-related peptide immunoreactive nerves in human and rhesus monkey eyes. Invest Ophthalmol Vis Sci 29:305, 1988. 10. Unger WG, Terenghi G, Ghatei MA, Ennis KW, Butler JM, Zhang SQ, Too HP, Polak JM, and Bloom SR: Calcitonin gene-related peptide as a mediator of the neurogenic ocular injury response. J Ocul Pharmacol 1:189, 1985. 11. Stone RA and Kuwayama Y: Substance P-like immunoreactive nerves in the human eye. Arch Ophthalmol 103:1207, 1985. 12. Stjernschantz J, Sears M, and Stjernschantz L: Intraocular effects of substance P in the rabbit. Invest Ophthalmol Vis Sci 20:53, 1981. 13. Ueda N, Muramatsu I, and Fujiwara M.: Capsaicin and bradykinin induced substance P-ergic responses in the iris sphincter muscle of the rabbit. J Pharmacol Exp Ther 230:469, 1984. 14. Holmdahl G, Hakanson R, Leander S, Rosell S, Folkers K, and Sundler F: A substance P antagonist, (D-Pro2, D-Trp7-9) SP, inhibits inflammatory responses in the rabbit eye. Science 214:1029, 1981. 15. Brecha N, Elred W, Kuljis RO, and Karten HJ: Identification and localization of biologically active peptides in the vertebrate retina. In Progress in Retinal Research, Osborne N and Chader J, editors. Pergamon Press, 1984, pp. 185-226. 16. Brecha NC, Stemini C, Anderson K, and Krause JE: Expression and cellular localization of substance P/neurokinin A and neurokinin B mRNAs in the rat retina. Vision Neuroscience 3:527, 1989. 17. Glickman RD and Adolph AR: Acetylcholine and substance P: Action via distinct receptors on carp retinal ganglion cells. Invest Ophthalmol Vis Sci 22:804, 1982. 18. Tsang D: Effect of substance P on dopamine release in rat retina. In Contemporary Themes in Biochemistry, Kon OL, Chung MCM, Hwang PLH, Leong SF, Loke KH, Thiayagarajah P, and Wong PTH, editors. 1986, pp. 588-589. 19. Lee CM and Cheung WT: Effects of neonatal monosodium glutamate treatment on substance P binding sites in the rat retina. Neurosci Lett 92:310, 1988. 20. Osborne NN: Substance P in the bovine retina: Localization, identification, release, uptake and receptor analysis. J Physiol (Lond) 349:83, 1984. 21. Hirata H, Baba S, Mishima H, and Choschi K: Specific binding of 125I-substance P in the bovine iris-ciliary body. Nippon Ganka Gakkai Zasshi 10:935, 1983. 22. Too HP, Unger WG, and Hanley MR: Evidence for multiple tachykinin receptor subtypes on the rabbit iris sphincter muscle. Mol Pharmacol 33:64, 1988. 23. Mantyh PW, Gates T, Mantyh CR, and Maggio JE: Autoradiographic localization of tachykinin receptor binding sites in the rat brain and peripheral tissues. J Neurosci 1:258, 1989. 24. Pernow B: Substance P. Pharmacol Rev 35:85, 1983. 25. Regoli D, Drapeau G, Dion S, and D'Orleans-Juste P: Receptors for substance P and related neurokinins. Pharmacology 38:1, 1989. 26. Stephenson JA, Summers RJ, and Burcher E: Autoradiographic localization and characterization of substance P binding in dog kidney. Eur J Pharmacol 142:391, 1987. 27. Beaujouan JC, Torrens Y, Daguet MC, Glowinski J, and Prochiantz A: Specific binding of an immunoreactive and biologi- 1901 cally active 125I-labeled substance P derivative to mouse mesencephalic cells in primary culture. Mol Pharmacol 22:48, 1982. 28. Mantyh PW, Johnson DJ, Boehmer CG, Catton MD, Vinters HV, Maggio JE, Too HP, and Vigna SR: Substance P receptor binding sites are expressed by glia in vivo after neuronal injury. Proc Natl Acad Sci U S A 86:5193, 1989. 29. Kieselbach GF, Ragaut R, Knaus HG, Konig P, and Wiedermann CJ: Autoradiographic analysis of binding sites for 125IBolton-Hunter-substance P in the human eye. Peptides 11:655, 1990. 30. Maggio JE: Tachykinins. Annu Rev Neurosci 11:13, 1985. 31. Kanazawa I, Ogawa T, Kimura S, and Munekata E: Regional distribution of substance P, neurokinin alpha, and neurokinin beta in the rat central nervous system. Neurosci Res 2:11,1984. 32. Saffroy M, Beaujouan JC, Torrens Y, Besseyre J, Bergstrom L, and Glowinski J: Localization of tachykinin (NK1, NK2, NK3 ligands) in the rat brain. Peptides 9:227, 1988. 33. Butler JM, Powell D, and Unger WG: Substance P levels in normal and sensorily denervated rabbit eyes. Exp Eye Res 30:311, 1980. 34. Tervo K, Tervo T, Eranko, L, Eranko O, and Cuello AC: Immunoreactivity for substance P in the gasserian ganglion, ophthalmic nerve and anterior segment of the rabbit eye. Histochem J 13:435, 1981. 35. Beding-Barnekow B, Brodin E, and Hakanson R: Substance P, neurokinin A and neurokinin B in the ocular response to injury in the rabbit. Br J Pharmacol 95:259, 1988. 36. Miller A, Costa M, Furness JB, and Chubb IW: Substance P immunoreactive nerves supply the rat iris and cornea. Neurosci Lett 23:243, 1981. 37. Shimizu Y, Kuwayama Y, Fukuda M, Ishimoto I, Shiosaka S, Inagaki S, Takagi H, Sakanaka M, Senba E, Kawai Y, Takatsuki K, and Tohyama M: Localization of substance P-like immunoreactivity in the anterior segment of squirrels: An immunohistochemical analysis. Invest Ophthalmol Vis Sci 22:259, 1982. 38. Soloway MR, Stjernschantz J, and Sears M: The miotic effect of substance P on the isolated iris. Invest Ophthalmol Vis Sci 21:47, 1981. 39. Oksala O: Effects of calcitonin gene-related peptide and substance P on regional blood flow in the cat eye. Exp Eye Res 47:283, 1988. 40. Krootila K: CGRP in relation to neurogenic inflammation and cAMP in the rabbit eye. Exp Eye Res 47:307, 1988. 41. Kuwayama Y and Stone RA: Distinct substance P and calcitonin gene-related peptide immunoreactive nerves in the guinea pig eye. Invest Ophthalmol Vis Sci 28, 1947, 1987. 42. Wahlestedt C, Beding B, Ekman R, Oksala O, Stjernschantz J, and Hakanson R: Calcitonin gene-related peptide in the eye: Release by sensory nerve stimulation and effects associated with neurogenic inflammation. Regul Pept 16:107, 1986. 43. Suzuki R and Kobayashi S: Different effects of substance P and vasoactive intestinal peptide on the motor function of bovine intraocular muscles. Invest Ophthalmol Vis Sci 24:1566, 1983. 44. Hagermark O, Hokfelt T, and Pernow B: Flare and itch induced by substance P in human skin. J Invest Dermatol 71:233, 1978. 45. Yousufzai SYK, Akhtar RA, and Abdel-Latif AA: Effects of substance P on inositol triphosphate accumulation, on contractile responses and on arachidonic acid release and prostaglandin biosynthesis in rabbit iris sphincter muscle. Exp Eye Res 43:215, 1986. 46. Unger WG and Tighe J: The response of the isolated iris Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017 1902 47. 48. 49. 50. INVESTIGATIVE OPHTHALMOLOGY b VISUAL SCIENCE / Moy 1991 sphincter muscle of various mammalian species to substance P. Exp Eye Res 39:677, 1984. Cole DF, Bloom SR, Burnstock, Butler JM, McGregor GP, Saffrey MJ, Unger WG, and Zhang SQ: Increase in SP-immunoreactivity in nervefibresof rabbit iris and ciliary body one to four months following sympathetic denervation. Exp Eye Res 37:191, 1983. Herkenham M: Mismatches between neurotransmitter and receptor localizations in brain: Observations and implications. Neuroscience 23:1, 1987. Marasco WA, Showell HJ, and Becker EL: Substance P binds to the formylpeptide chemotaxis receptor on the rabbit neutrophil. Biochem Biophys Res Commun 99:1065, 1981. Bar-Shavit Z, Goldman R, Stabinsky Y, Gottlieb P, Fridkin M, Teichberg VI, and Blumberg S: Enhancement of phagocytosis: A newly found activity of substance P residing in its N-terminal 51. 52. 53. 54. Vol. 32 tetrapeptide sequence. Biochem Biophys Res Commun 94:1445, 1980. Brecha N: Retinal neurotransmitters. In Chemical Neuroanatomy, Emson PC, editor. New York, Raven Press, 1983, pp. 85-129. Tornqvist K and Ehinger B: Peptide immunoreactive neurons in the human retina. Invest Ophthalmol Vis Sci 29:680, 1988. Brecha NC, Johnson D, Bolz J, Sharma S, Parnavelas JG, and Lieberman AR: Substance P immunoreactive retinal ganglion cells and their central axon terminals in the rabbit. Nature 327:155, 1987. Osborne NN and Ghazi H: The effect of substance P and other tachykinins on inositol phospholipid hydrolysis in rabbit retina, superior colliculus and retinal cultures. Vision Res 29:754, 1989. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933387/ on 06/17/2017