Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
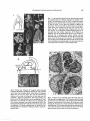
Plant Cell Physiol. 40(7): 657-667 (1999) JSPP © 1999 Transgenic Tobacco Over-Expressing a Homeobox Gene Shows a Developmental Interaction between Leaf Morphogenesis and Phyllotaxy Masanori Tamaoki1' 3, Asuka Nishimura1, Mitsuhiro Aida2, Masao Tasaka2 and Makoto Matsuoka1 1 2 Nagoya University, BioScience Center, Nagoya, Aichi, 464-8601 Japan Graduate School of Biological Science, Nara Institute of Science and Technology, 8916-5 Takayama, Ikoma, Nara, 630-0101 Japan The tobacco gene, NTH1, encodes a polypeptide of 326 amino acids and is a member of the classl KN1type family of homeobox genes. Expression of NTH1 has mainly been observed in vegetative and reproductive shoot apices, not observed in roots or expanded leaves. Over-expression of NTH1 in transgenic plants caused abnormal leaf morphology, consisting of wrinkling and curvature. Interestingly, the direction of leaf curvature tended to be conserved among almost all of the leaves in any given transformant. In transgenic plants exhibiting clockwise or anticlockwise phyllotaxy, leaves curved to the right or left, respectively, when looking from the shoot apex toward the base. Micro-surgical experiments demonstrated that the presence of the shoot apex is necessary for the development of leaf curvature, indicating that the order of formation of leaves on the stem (the generative spiral) affects leaf development. We found a correlation between the severity of leaf curvature and the value of the plastochron ratio, a parameter of phyllotaxy. Transformants with more severe phenotypes had larger plastochron ratios. From these findings, we discuss the possibility that an increase in the plastochron ratio, caused by over-expression of NTH1 in the shoot apex, may be involved in leaf curvature. velopment (Poethig and Sussex 1985). Given that the early development of leaf primordia occurs in such close proximity to the meristem, it is reasonable to expect that this process must involve important developmental interactions between the SAM and the leaf primordia. For example, it has been predicted that the shoot apex plays an important role in the acquisition of dorsoventral character by leaf primordia (Wardlaw 1949, Sussex 1955, Snow and Snow 1959). The arrangement of leaves along the stem, termed phyllotaxy, is strictly determined by the regular initiation of leaf primordia at the SAM, providing another example of their close developmental relationship (Richards 1948, Wardlaw 1949). These examples suggest that such developmental interactions between the SAM and leaf primordia are common features of many plants and may contribute to their variety. While many such examples can be found, very little information is available concerning the actual mechanism(s) underlying the developmental correlation. In the work presented here, we have discovered a clue to the developmental relationship between leaves and the shoot apex through an analysis of transgenic tobacco plants ectopically expressing the tobacco homeobox gene, NTH1 (Nicotiana tabacum homeobox 1). Homeobox genes were first identified as encoding conserved protein motifs, known as homeodomains, that control morphogenesis in the fruit fly, Drosophila. Because the secondary structure of homeodomains resembles the helix-turn-helix motif found in some bacterial DNA binding proteins, the products of homeobox genes are believed to regulate the expression of target genes by acting as transcriptional factors (Affolter et al. 1990). Genes encoding homeodomain proteins have been isolated from many plant species and have been categorized into four groups according to the amino acid sequences of their homeodomains (Kerstetter et al. 1994). Among these, the most well characterized group is the KNl-type homeodomain. Genes in this group show specific expression in the region of the SAM, and their ectopic expression in spontaneous mutants and in transgenic plants alters leaf and flower morphology (Freeling et al. 1992, Matsuoka et al. 1993, Jackson et al. 1994, Lincoln et al. 1994, Tamaoki et al. 1997). Based on these observations, it has been proposed that the KNl-type homeobox genes are involved in lateral organ formation from the SAM; however, their roles in such processes have not been well defined. Key words: Homeobox gene — Micro-surgical experiment — Nicotiana tabacum — Phyllotaxy — Plastochron ratio — Transgenic plant. Although an extraordinary variety of plant shapes exist, they are all formed from only a few organs, including roots, stems and leaves. Among these, stems and leaves constitute the above-ground portion of the plant, termed the shoot system, and are produced from the shoot apical meristem (SAM) throughout the vegetative phases of plant development. The leaf primordium is initiated as a buttress from the flanking region of the SAM during plant deAbbreviations: SAM, shoot apical meristem; DA, divergence angle; PR, plastochron ratio. The nucleotide sequences reported in this paper have been submitted to the DDBJ, EMBL, Genebank under accession number AB025573. 3 Present address: National Institute for Environmental Studies, Onogawa 16-2, Tsukuba, 305-0053 Japan. 657 658 Developmental interaction between the SAM and leaf In this paper, we describe the morphological characteristics of transgenic tobacco plants overexpressing NTH1 and show that a correlation exists between leaf development and phyllotaxy. We have found that the degree of leaf curvature depends on the value of the plastochron ratio, one of the parameters of phyllotaxy. Materials and Methods Plant growth conditions—Tobacco (Nicotiana tabacum cv. Samsun NN) seeds were sterilized in 5% sodium hypochlorite for 5 minutes and germinated on germination medium (Murashige and Skoog salts plus 3 % sucrose and 0.3% Gellangum) under constant white light at 25°C. Seedlings were transplanted to soil and grown at 25°C in a 16 h light-8 h dark cycle. Isolation and sequencing of NTH1 cDNA clones—Total RNA from tobacco shoot apices was amplified by RT-PCR using two oligonucleotide primers corresponding to the conserved region of the homeodomain (5'-AA(A/G)CT(A/C/G/T)CC(A/C/ G/T)AA(A/G)GA(A/G)GC-3', and 5'-TG(A/G)TT(A/T/G)AT(A/G)AACCA(A/G)TT(A/G)TT-3'). Amplified fragments were cloned into the vector pCRII (Invitrogen) and sequenced. One clone corresponding to NTH1 was used as a probe for further screening of cDNA clones. A cDNA library was constructed using total RNA extracted from shoot apices of mature tobacco plants, and poly(A)-enriched RNA was purified by two passes through an oligo d(T) cellulose (type III, Becton Dickinson Labware) column. Thepoly(A) RNA was used to synthesize double stranded cDNA, which was cloned into the EcoRl site of Agtll (Stratagene). Screening was performed in 50% Formamide, 6 x SSC, 5x Denhardt's solution, 0.5% SDS, and 20/jg ml" 1 salmon sperm DNA at 42°C for 14 h. Filters were washed in 2 x SSC, 0.1% SDS at room temperature and then further washed in 0.2 x SSC, 0.1% SDS at 65°C. Nucleotide sequences were determined by the dideoxynucleotide chain-termination method using an automated sequencing system (ABI 373A). The cDNA clone was completely sequenced in both strands. Analysis of cDNA and amino acid sequences was carried out using GENETYX computer software (Software Kaihatsu Co., Japan). Quantitation of NTH1 expression by RT-PCR—RT-PCR amplification using total RNA from various organs as a template was performed to analyze the expression level of NTH1. First strand cDNA was produced from 1 ng of total RNA and was used as a template in quantitative PCR, as described previously (Tamaoki et al. 1995). The locations of oligo-nucleotide primers used for PCR are presented in Fig. 1. Construction of chimeric genes and tobacco transformation—-The full-length NTH1 cDNA clone was cut with EcoRW/ Sad, and the cDNA insert was introduced into Smal/Sad-cut pBI121 to construct the 35S::NTH1 fusion gene in a binary vector. This construct was then introduced into Agrobacterium tumefaciens LBA4404 by electroporation. Agrobacterium-mediated transformation of tobacco leaf discs was performed as previously reported (Tamaoki et al. 1997). Transgenic plants were selected on media containing 100 mg liter" 1 of kanamycin. RNA gel blot analysis—Total RNA was prepared from leaves of transformants for RNA gel blot analysis. Ten fxg of each RNA sample were transferred to Hybond N membrane (Amersham) and probed using a 320 bp EcoRl/Bglll fragment of the NTH1 cDNA, which did not include the homeobox sequence to avoid cross-hybridization with other homeobox genes. Hybridization was performed under the same conditions as cDNA screening. Micro-surgical experiments—For isolation of leaf primordia (P4 or P5 stage) from the SAM, shoot apices were cut tangentially with a razor blade (see Fig. 7A). A cover-glass (thickness 0.13 mm, IWAKI) was inserted into the cut to prevent adhesion of the isolated leaf primordium to the SAM. The shoot apex was covered with a plastic bag to prevent it drying out and treated plants were grown under the same conditions as untreated plants. For culturing excised leaves, shoot apices were separated into two parts using a scalpel; one contained the SAM and several leaf primordia and the other contained only a leaf primordium about 3 mm in length (see Fig. 8A). These excised fragments were transferred to MS basal medium (pH 5.7) consisting of MurashigeSkoog salts supplemented with 100 mg liter"1 myo-inositol, 10 mg liter"1 thiamine-HCl, 1 mg liter"1 nicotinic acid, 1 mg liter"1 pyridoxine-HCl, \% (w/v) sucrose, and 0.3% Gellangum (Wako) and were grown under sterile conditions for 1 month. Histological analysis—Tobacco shoot apices were fixed in solution of 4% paraformaldehyde plus sodium phosphate buffer, pH 7.4, overnight at 4°C then dehydrated through a graded ethanol series followed by a f-butanol series, and finally embedded in Paraplast Plus (Sherwood Medical). Microtome sections (8 tun thick) were mounted on glass slides treated with Vectabond (Vector Labs). For measurements of divergence angle (DA) and plastochron ratio (PR), the sections were briefly stained with 0.1% toluidine blue O (Merck) for 15 minutes and then de-paraffinized in xylene. To obtain clear sections in the region of the SAM, the sections were stained following the method of Sharman (1943). Results Structure and expression ofNTHl—To isolate a novel member of the KNl-type homeobox gene family from tobacco, we designed degenerate oligonucleotide primers corresponding to conserved amino acid sequences from the homeodomain region of previously reported KNl-type homeobox genes (Kerstetter et al. 1994). These primers were used for PCR amplification with total RNA isolated from shoot apices as a template. Several independent clones were isolated from the PCR products and sequenced. Among these, one clone, designated NTH1 (Nicotiana tabacum homeobox 1), was used as a probe to screen for full-length cDNA clones. We analyzed the entire nucleotide sequence of the NTH1 cDNA clone. The predicted 326 amino acid NTH1 protein (Fig. 1) contained a 64 amino acid conserved homeodomain sequence near its C-terminus. This homeodomain sequence was compared to other known KNl-type homeodomains. The KNl-type homeodomains can be subdivided into two classes, class 1 and class 2 (Kerstetter et al. 1994). Class 1 genes are more similar to the maize KN1 gene; their products share high amino acid identity in the homeodomain and are expressed primarily in shoot and floral meristems, not in leaves or roots. The products of class 2 genes are comparatively less similar to KN1 in their homeodomains (only about 60% identity) and are expressed in all tissues. The homeodomain of NTH 1 is highly homologous to that of LG3 from maize (88% identity, Developmental interaction between the SAM and leaf 659 GACCTGTTrcTCTCCCTCTTTAACTTTKMAATAAACCTTAAAACGGTGCAGGTTAATAAACTTCGCAGATTTTATTATTACACACTGTGT GTGTATGTGAAAGGGTTTGTTTGGTTTCTAACC AAGTTACTATTGTTTCTTGTTTAGTACTATTTGATGATGGATGAATTGAGC AAACTT M D E L S K L H S A I A V D C S N N N L E N K N A H S V L V E G M V L E H I S T K G Q A Q Q I Q A E N G H I P N 90 180 7 270 37 360 67 L AACTTATTATCTGCCTACCTTCAATGCCGAAAGGTTGGAGCACCCC AAGAAATGGCATCAATTCTTGAAGAAATCAGCAAAGAGAACC AC N L L S A Y L Q C R K V G A P Q . E M A S I L E E I S K E N H 450 97 TTAATATCCAGCGGCCACAATACTGAAATCGGAACTGATCCAGAACTTGATGATTTTATGGAATCATATTGTGCGGTATTACTAAAATAC H N T E I G T D P E L F M E S Y C A V L L K Y 540 127 K E E L S K P D E A T T F L N N I E S D C G E M E A A Q E L A S G S G L T C S K D E N L D L C E 630 157 D TATAGTGGCTACCTTAGCAGCTTGAGAAAGGAATTTTTGAAGAAGAGGAAGAAAGGCAAGTTACCCAAGGATGCAAGAACTGCATTGCTG Y S G Y L S S L R K E F L K K R K K G K L P K D A R T A L L E W W N N N W T F H Y R W N Q R P K Y R P H T W E R E P E S K N E R D L M S K E Y I T A G L L M D E G P K V S 990 277 Q S 900 247 G 1080 307 AGCATTGCCGGATCCATGTATTTTGATGGAGCTGGAGGCACTGGAAGTATTGGCACTTGAAAAAAGAGAAAAAAACTTGCTGCAACTTTT S I A G S M Y F D G A G G T G S I G T * 1170 326 CAAAGTATAACTAGCTAACTAAC AAGTAGTGC ATAGTATGACAGAAACATTTGTATTTCGAAGTTTATATATTGACTTATACTATTC AGA 1260 AAAAAAAAAAAAAAA Fig. 1 Nucleotide and deduced amino acid sequences of NTH1. Numbers at right indicate nucleotide (upper) and amino acid (lower) positions. Dashed underlining indicates the ELK domain; solid underlining indicates the homeodomain. Arrows indicate positions of the primers used for RT-PCR (see Fig. 2). Kerstetter et al. 1994) which is a class 1 gene. The ELK domain, which is a conserved 21 amino acid stretch adjacent to the KNl-type homeodomain, was also conserved in NTH1. The amino acid sequence of the NTH1 ELK region was more similar to the ELK domain of class 1 gene products than to those of class 2 genes. These structural features indicate that NTH1 is a KNl-type class 1 homeobox gene. We analyzed the expression pattern of NTH1 in various organs by northern blot analysis, but we were unable to detect any NTH1 transcript in any organ. Because of its higher sensitivity, we used quantitative RT-PCR to analyze NTH1 expression in various organs in tobacco. Two synthetic primers were designed outside the homeodomain for this reaction (see Fig. 1). The predicted 325 bp product was amplified from RNA isolated from several organs (Fig. 2). This band was produced from the NTH1 transcript because its sequence was identical to the corresponding sequence of the NTH1 cDNA. Expression of NTH1 was mainly detected in shoot apices, floral buds, flowers, and stems, while a weak band of the same size was also produced from RNA from unexpanded leaves. No NTH1 expression was detected in roots or expanded leaves. This organ-specific expression pattern is similar to that of other class 1 genes, further suggesting that NTH1 is a KNl-type class 1 homeobox gene. Ectopic expression of NTH1 in transgenic tobacco causes altered leaf morphogenesis—To gain further insight into the biological functions of NTH1, the NTH1 coding region was transcriptionally fused to the 35S promoter from cauliflower mosaic virus, and the resulting construct was used to transform tobacco. Eighteen independent transformants were obtained, all of which exhibited abnormal morphology (Fig. 3A). Such morphological alterations were dominantly transmitted to the second and third generations and were linked to kanamycin-resistance, indicating that they were caused by the transgene (data not shown). The most notable changes in morphology in the NTH1 transgenic plants were in leaf development. Leaves of transformants were slightly wrinkled and were curved to the right or left side, whereas leaves of wild-type plants were nearly bilaterally symmetrical (Fig. 3B). In an effort to 660 SA Developmental interaction between the SAM and leaf FB F R ST UL EL Anticlockwise f 1 2 3 Wild M 4 5 Clockwise M 6 7 8 9 11 325 bp Phenotype Fig. 2 RT-PCR analysis of NTH1 expression in wild-type tobacco plants. Total RNA was isolated from vegetative shoot apices (SA), floral buds (FB), mature flowers (F), roots (R), stems (ST), unexpanded leaves (UL), and expanded leaves (EL). Single-stranded cDNA was synthesized from 1 ^g of each RNA sample. The positions of the primers used for PCR are indicated in Fig. 1. The reaction volumes were 20 /A, of which 5//I were loaded per lane. Arrow indicates the 325 bp PCR product which corresponds to the size predicted from the nucleotide sequence of NTH1. determine the cause of the leaf curvature, we examined numerous cross sections of transgenic leaves but could not find any clear differences in the anatomy of laminae between their left and right sides. We also compared the weight of left and right sides of the laminae and found that the outer side of curved leaves was slightly heavier than the inner side (data not shown). This observation indicates that the leaf curvature in transformants is caused by different rates of growth of the two sides of the lamina. To examine transgene expression in leaves of 35S:: NTH1 transformants, RNA gel blot analysis was performed. Total RNA was isolated from leaves of transgenic plants with different degrees of leaf curvature and exhibiting opposite directions of leaf formation order (see below) and was used for RNA gel blot analysis. NTH1 mRNA was barely detectable in mature leaves of wild-type plants, whereas a single 1.3 kb band from the transgene was observed in all transgenic plants (Fig. 4). The severity of leaf curvature correlated with level of expression of the transgene (Fig. 4). Fig. 3 Phenotype of transgenic tobacco carrying the 35S::NTH1 construct. (A) Typical abnormal phenotype caused by NTH1 over-expression. All the leaves are slightly wrinkled and curved. (B) Comparison of mature leaves from a wild-type (left) and N77/7-transformed (right) plants. Both leaves were taken from the middle region of the stem of plants three months after regeneration. Severe Mild Mild Severe Fig. 4 Transgene expression in leaves of 35S::NTH1 transformants. RNA was isolated from leaves of two independent wild-type (lane 5, 6) and eight independent transgenic tobacco plants (lane 1-4, 7-10). Transgenic plants are divided into two groups (Anticlockwise and Clockwise, indicated at top) based on the direction of their generative spiral. Each transgenic group is further divided according to the degree of leaf curvature, Mild to Severe, indicated by arrows at bottom. Severity of phenotype increased from lane 4 to lane 1 in anticlockwise plants, and from lane 7 to lane 10 in clockwise plants. 10 fig of total RNA were loaded per lane. The blot was probed with a fragment of the NTH1 cDNA which did not include the homeobox sequence (see Materials and Methods). The approximate size of transcripts was 1.3 kb. Correlation between the direction of generative spiral and the direction of curvature of the malformed leaves in transgenic plants—Transgenic tobacco plants produced malformed leaves with a curved midvein and wavy leaf blade (Fig. 3B). We noticed an interesting relationship between the direction of leaf curvature and the direction of leaf formation in all of the transgenic plants. The order of Fig. 5 Correlation between the order of leaf formation and the direction of leaf curvature in transgenic plants. Transgenic plants exhibited a clockwise generative spiral (A) and an anticlockwise generative spiral (D). Transverse section through the shoot apex of a clockwise spiral plant (B) and an anticlockwise spiral plant (E). Leaf primordia are numbered in the order of their appearance. The direction of the generative spiral is defined by tracing through these numbers counting down from the fourth leaf. Leaves from a clockwise plant (C) and an anticlockwise plant (F). In each case, all leaves curve in the same direction. Developmental interaction between the SAM and leaf leaf formation can be visualized as a spiral drawn to pass through the series of leaves differing in number by one. This spiral, which is drawn as a helical pattern ascending the stem in order of decreasing leaf age, is commonly referred to as the generative spiral and defines the direction of leaf formation around the shoot apex (Callos and Medford 1994). In wild-type tobacco, the generative spiral is maintained throughout vegetative development, and clockwise and anticlockwise spirals are observed with equal frequency. Generative spirals were also seen in the NTH1 transformants (Fig. 5B, E); transformants with a clockwise generative spiral exhibited malformed leaves curved to the right side (Fig. 5A, B, C), whereas the leaves of plants with an anticlockwise spiral curved to the left side (Fig. 5D, E, F). For quantitative analysis, we determined whether the correlation between the directions of the generative spiral and leaf curvature held for all of the transformants. The Tl transformants were grouped according to the direction of their generative spiral, and leaves were counted. Five plants exhibited a clockwise spiral, and the remaining thirteen had an anticlockwise spiral. An average of twenty leaves were formed on each transgenic plant. The leaves were categorized into three groups, left, straight or right, Clockwise 661 based on the direction of their curvature. On average, transgenic plants with a clockwise spiral formed about three leaves curved to the left, three straight leaves, and eleven leaves curved to the right (Fig. 6). In the plants with an anticlockwise spiral, the average number of leaves curved to left, straight, and right was fourteen, five, and three, respectively (Fig. 6). These results strongly suggest that the direction of leaf curvature is related to the order of leaf formation in the transgenic plants. The shoot affects the development of malformed leaves in transformants—The previous results strongly suggest that the direction of the generative spiral of leaves affects the direction of leaf curvature. Leaves arise at regular intervals and are regularly spaced around the SAM. Such an arrangement might be expected to be controlled by influences of the SAM (see Discussion), so it seemed logical to test whether the observed asymmetry in leaf development also required the SAM. Therefore, it seemed possible that the curvature of transgenic leaves might be diminished or changed if a leaf primordium were isolated from the shoot apex, we surgically isolated leaf primordia from the meristem and examined leaf shape during the developmental process. Two different means of isolating leaf primordia from the shoot apex were employed. First, we Anticlockwise Right Straight Left 20 CD JO #1-3 #1-6 #1-14 #1-18 #1-19 Ave. #1-1 #1-4 #1-5 #1-7 #1-8 #1-9 #1-10 #1-11 #1-12 #1-13 #1-15 #1-16 #1-20 Ave. Transgenic line Fig. 6 Quantitative analysis of the relationship between the direction of the generative spiral and the direction of leaf curvature in eighteen iV77/7-transformed plants. Transgenic plants were classified as clockwise or anticlockwise according to the direction of their generative spiral. Leaves of each plant were counted according to the direction of their curvature (left, straight or right). All Tl transformants were tested at three months after regeneration. Ave.; Average of the number of leaves per plant in each category. Bars indicate standard error. 662 Developmental interaction between the SAM and leaf incised the shoot apex tangentially to isolate a leaf primordium from the meristem (Fig. 7A). Leaf primordia were separated when they were about 2-3 mm in length. At that time, the lamina were already present, and the primary lateral veins had started to expand from the midvein (Poethig and Sussex 1985). The morphology of these isolated leaves was observed at eight days after incision. A total of over forty T2 progeny of the primary transformants were incised, but only ten leaves had developed at eight days after incision. All the living leaves developed with similar morphology. At two days after incision, the leaves showed no significant growth (Fig. 7B). They then started to expand and developed into mature leaves by eight days after incision (Fig. 7C, D). Isolated leaves developed to about 6 cm in length by eight days after incision. Leaves of this length in untreated transgenic plants already exhibited a curved phenotype (Fig. 7G). The isolated leaves of transformants were slightly wrinkled but were not curved, and the leaf blades developed almost symmetrically, as did leaves of non-transgenic plants (Fig. 7E, F). Leaves below the isolated leaf on transgenic plants were curved in the same direction (Fig. 7C, D). The incision did not affect the leaf morphology of wild-type plants (Fig. 7H). The second method by which leaves were isolated was by completely excising the meristem from the stem and dividing it into two parts. The separated "block B" contained the SAM with several leaf primordia and the remaining part (block G) contained only a leaf primordium approximately 3 mm long (Fig. 8A). After removal, the isolated fragments were grown in sterile culture. A total of at least thirty pairs of primordia were cultured. One month after excision, only five pairs were alive and growing. Leaf -primordia still associated with the SAM developed curved leaves, while isolated leaf primordia developed leaves that were nearly symmetrical and wrinkled (Fig. 8B, C). These results strongly suggest that the presence of the SAM is essential for the curvature of the leaves in NTHl-trans- formed tobacco. Correlation between severity of leaf curvature and plastochron ratio—The relationship that exists between the direction of the generative spiral of a stem and the direction of leaf curvature, indicates that phyllotaxy affects leaf development in the transgenic plants (Fig. 5,6). Phyllotaxy is defined as the arrangement of leaves along the axis of a stem and is described using three parameters: order of leaf formation, divergence angle (DA), and plastochron ratio (PR) (Richards 1951). To more precisely examine the relationship between leaf curvature and phyllotaxy, we compared these parameters between wild-type and transgenic tobacco plants. The order of leaf formation in transformants showing clockwise or anticlockwise generative spirals was the same as in non-transgenic tobacco (Fig. 6). The parameters DA and PR were obtained by examining transverse sections through the shoot apex. In order to estimate these parameters, it is necessary to determine the centers of the leaf primordia and the SAM. Once these centers are determined, lines can be drawn between the center of each leaf primordium and the center of the SAM. DA and PR are then estimated from these lines. DA is determined by the smaller angle between the lines of two successively formed leaf primordia. PR is calculated by dividing the distance from the apical center to the center of a leaf primordium by the distance from the apical center to the center of the next younger leaf primordium. For determination of values of DA and PR, we examined sections through the shoot apices from several wild-type and transgenic plants (Fig.9A, B). We did not find any significant differences in DA between non-transformants (140° ±1) and the transformants (141 °±1) when we compared the values of DA between leaf primordia P3-P7 (Table 1). In contrast to DA, PR differed between the transformants (1.6±0.08) and non-transformants (1.3±0). We also estimated the value of PR in transformants exhibiting weaker leaf curvature. The value of PR for these plants was estimated to be 1.4±0, which is intermediate between that of Table 1 The divergence angle (DA) and plastochron ratio (PR) for leaves in the shoot apex of wild tobacco and mildand severe-transgenic plants PR DA Wild-type Average 139±7 141 ±4 141 ±5 139±5 140±6 140±l Transgenic (Mild) Transgenic (Severe) Wild-type 140±5 138±6 139±5 142 ±8 141 ±7 141±3 143 ±3 141±5 141±6 141 ±1 1.3±0.15 1.3±0.15 1.3 ±0.04 1.3±0.05 1.3 ±0.05 1.3±0 — 140 ±1 Transgenic (Mild) Transgenic (Severe) 1.4±0.15 1.4±0.07 1.4±0.15 1.4±0.15 1.7±0.33 1.5 ±0.04 1.5 ±0.07 1.6±0.12 1.5 ±0.08 1.6±0.08 — 1.4±0 Each data is mean of DA or PR value obtained from P3 to P7 leaves of a plant. The average shown at the bottom of each column is presented as a mean ±standard error obtained from five (mild transformants are four) independent plants. Developmental interaction between the SAM and leaf 663 Fig. 7 Leaf primordia isolated from the SAM develop normally in transgenic plants. (A) Diagram indicating the method of isolating a leaf primordium from the SAM. A tangential incision (dashed line) was made between a leaf primordium at stage P4 or P5 and the SAM. A cover glass was inserted to separate the primordium from the stem and was held in place with parafilm. Upper leaves were removed, and the apex was covered with a plastic bag. (B) A surgically treated transgenic plant two days after incision. (C, D) Surgically treated transgenic plants at six and eight days after incision, respectively. (E, F) Higher magnification views of the isolated leaves shown in (C) and (D), respectively. (G) A transgenic plant without incision. One leaf primordium (P4 or P5) remained at the shoot apex, and all other leaves around the shoot apex were removed. The remaining leaf grew to about 6 cm in length after eight days and was curved. (H) A leaf exhibiting normal morphology at six days after incision; it was isolated from the SAM of a non-transgenic plant. Fig. 8 Excised leaf primordia of transgenic plant developed symmetric leaf blades. (A) Diagram showing how leaf primordia were excised from transgenic plants. Shoot apices of transgenic plants were cut into two parts. One part (B) contained the meristem with several leaf primordia, and the other part (C) contained a single leaf primordium. The excised blocks were grown on the MS basal medium, and the morphology of developed leaves was observed one month after the operation. (B) Curved leaves developed from blocks containing the SAM. This block generated several leaves and many roots during one month of incubation. (C) Blocks containing only an excised leaf primordium (block C) developed a wrinkled leaf with a nearly symmetrical blade. Fig. 9 Position of leaf primordia around the shoot apex of a wild-type and a transgenic plant. (A) Transverse section through the shoot apex of a wild tobacco. The direction of leaf primordia formation is anticlockwise in this plants. (B) Transverse section through the shoot apex of a severe transformant with the clockwise phyllotaxy. Five successive leaf primordia are indicated as P3 to P7. Each white line is drawn between the center.of the shoot apical meristem and the center of each leaf primordium. Developmental interaction between the SAM and leaf 664 (A) WildType(PR=1.3) P(n) P(n+2) Transgenic (PR=1.6) P(n+3) P(n+3) P(n) P(n+2) p(n.3)j --._.__P(n-3); • P(n-2) *s'«'iPl ,- O : Center of the SAM P(n) (C) Distance from P(n) Wild type Transgenic D(n+3) D(n+2) D(n±/)2 = a? +1>2 - 2 a- b • cose When / = 2 , 8 = 80° and when ( = 3 . 9 = 60°. D(n-2) D (n-3) T/W 1.94 1.42 0.95 1.05 Fig. 10 Comparison of leaf positions around the SAM between wild and transgenic tobacco. (A) Diagram of the shoot apices of wild-type (left) and transgenic (right) plants showing the arrangement of leaf primordia. Anticlockwise organization of seven leaf primordia; P(n — 3) to P(n + 3), is shown for both apices and their positions are described by the values of DA and PR shown in Table 1. Both wild-type and transgenic plants showed the same value of DA (about 140°). The PR value of transformants exhibiting a severe phenotype was 1.6, while that of wild-type plants was 1.3. (B) When the distance of P(n) from the center of the SAM (O) is described as a, the distance between P(n) and other leaf primordia, denned as D(n±/), can be described as a trigonometric function of a. (C) Distances between P(n) and the adjacent leaves in wild-type and transgenic plants. The distances are indicated as D(n±2) or D(n±3). The calculated results were obtained using the estimated values of DA and PR. T/W, the ratio of the distance from P(n) to P ( n ± 0 . is calculated by dividing D ( n ± 0 for transformants by D ( n ± 0 for wild-type plants. non-transformants and severe transgenic plants, while DA (140° ±1) did not change (Table 1). These observations demonstrate a correlation between the value of PR and the severity of leaf curvature. Arrangement of leaf primordia in wild-type and transgenic tobacco—Applying the values of DA and PR of wildtype and transgenic tobacco plants, a leaf primordium n, defined as P(n), is flanked by previously-initiated leaf primordia P(n + 2) and P(n + 3) and by later-initiated primordia P ( n - 2 ) and P ( n - 3 ) (Fig. 10A). The distance between P(n) and these flanking leaf primordia, defined as D(n±2) or D(n±3), is a function of a, where a is the distance of P(n) from the center of the SAM (Fig. 10B). Using these functions, D(n + 3)2, D(n + 2)2, D(n-2) 2 and D ( n 3)2 are calculated as a2 + (PR) 6 a 2 -2(PR) 3 a 2 cos 60°, a2 + (PR)V-2(PR) 2 a 2 cos 80°, a 2 + ( l / P R ) V - 2 ( l / P R ) 2 a 2 cos 80°, and a2 + (l/PR) 6 a 2 -2(l/PR) 3 a 2 cos 60°, respectively. When the value of PR is applied to these functions, the values of D(n + 3), D(n + 2), D ( n - 2 ) and D ( n - 3 ) in nontransformants (PR = 1.3) are V3.63a, V3.27a, Vl.l4a, and V0.75a, respectively (Fig. 10C). In transgenic plants (PR = 1.6), the values for the same parameters are Vl3.7a, V6.66a, Vl.02a, and V0.82a, respectively. The values of D(n + 3), D(n + 2), D(n —2) and D ( n - 3 ) in transformants with weaker curvature were calculated to be V5.79a, V4.16a, Vl.l3a, and V0.76<z, respectively (data not shown). To compare the change in distance from P(n) and P(n±2 or 3), the ratio of the distance from P(n) to P(n±2 or 3) between wild-type and transgenic plants (T/W as defined in the figure legend) is calculated by dividing D(n +3), D(n + 2), D(n-2) and D(n —3) for transformants by these for wild-type plants, respectively. The values for T/W of P(n + 3), P(n + 2), P ( n - 2 ) and P(n-3) are 1.94, 1.42, 0.95, and 1.05, respectively (Fig. 10C). Discussion In this study, we showed that transgenic tobacco plants which over-expressed a tobacco homeobox gene, NTH1, formed abnormal leaves. Although there are several studies of over-expression of the KNl-type homeobox gene in tobacco (Sinha et al. 1993, Kano-Murakami et al. 1993, Tamaoki et al. 1997), the following new findings are made for the first time in this study. (1) Only the mild class of KNl-type homeobox gene misexpression phenotype is observed in NT/ZV-transformants. (2) The direction of leaf curvature correlates with the direction of leaf production order. (3) The expression of leaf curvature is dependent on the presence of the SAM. (4) The severity of leaf curvature correlates with the value of plastochron ratio. These points are discussed below. As reported in our previous study, the malformed Developmental interaction between the SAM and leaf leaves of tobacco plants transformed with 35S::NTH15 can be categorized into three groups, ranging from mild to severe on the basis of their phenotype (Tamaoki et al. 1997). According to this classification scheme, all the NTH1transformed tobacco plants produced leaves with a mild phenotype (Fig. 3B). Indeed, we never observed plants with an intermediate or severe phenotype in this experiment. The differences in leaf phenotypes caused by NTH1 and other KNl-type genes may not be due to the expression level of the transgenes, because their expression was driven by the same promoter, 35S. In fact, transgene expression showed a range of levels in different NTT/i-transformants (Fig. 4). Therefore, the narrow range of phenotypic severity among the N77/7-transformants depends upon a unique structural character of the NTH1 gene. In the Nterminal region of some KNl-type homeodomain proteins, homopolymeric amino acid stretches are observed (Vollbrecht et al. 1991, Matsuoka et al. 1993, Schneeberger et al. 1995, Tamaoki et al. 1997); no similar characteristic structures are found in NTH1 (Fig. 1). The biological role of these homopolymeric amino acid stretches has not yet been determined. However, it has been suggested that they may be involved in transcriptional activation, because polyproline and polyglutamine stretches often function as activation domains in trans-acting factors (Gerber et al. 1994). Therefore, these N-terminal homopolymeric amino acid stretches may be important for the severity of leaf malformation in transgenic tobacco plants, and the lack of such structures in NTH1 may reduce the severity of leaf malformation. Our preliminary experiments involving domain swapping between NTH1 and NTH 15 show that the Nterminal region outside the homeodomain is important in determining the phenotypic severity of transformants (Sakamoto et al. unpublished result) may support this idea. The micro-surgical experiments showed that P5 leaf primordia, 2-3 mm in length, isolated from the SAM developed into nearly symmetric leaves (Fig. 7, 8). Moreover, the isolated leaves showed a wrinkled morphology that was also observed in non-treated transgenic tobacco plants. These data indicate that the expression of abnormal leaf morphology observed in the present study is divided into two independent developmental steps. One is expression of leaf curvature that requires an interaction between the leaf primordia and the SAM, and the other is an expression of wrinkled phenotype that is independent from the SAM and only requires over-expression of NTH 1. The abnormal leaf morphology observed in this experiment has also been reported in transgenic tobaccos over-expressing other KN1type homeobox genes (Sinha et al. 1993, Kano-Murakami et al. 1993, Tamaoki et al. 1997). In these reports, the expression of abnormal leaf morphologies is explained as a result of disorganization of cell division in leaves that follows the over-expression of A7V7-type homeobox genes (Sato et al. 1996). This hypothesis can elucidate the ab- 665 normal leaf development in transgenic plants but not explain how the direction of leaf curvature is determined, because disorganized cell division may occur on both sides of leaf blade in equal frequency. Thus, the present study proves for the first time that the expression of leaf curvature in transgenic tobacco is due to the activity of the SAM. It is interesting to speculate how the direction of leaf curvature is determined. The direction of leaf curvature is correlated with the direction of phyllotaxy in transgenic tobacco, as if the expression of leaf curvature requires the phyllotaxy (Fig. 5, 6). Moreover, there is a positive correlation between the severity of leaf curvature and the value of PR (Table 1), a parameter of phyllotaxy that expresses the extent of leaf primordia at the SAM (Callos and Medford 1994). Therefore, the expression of leaf curvature may depend on the arrangement of leaf primordia around the SAM. The diagram shown in Fig. 10, which illustrates the arrangement of leaf primordia around the SAM in wild-type and transgenic tobacco, demonstrates that when the value, of PR increased as in the transformants, only D(n —2), the distance between P(n) and P ( n - 2 ) is decreased while all the other distances are increased. In other words, the increase of PR in the transformants causes P ( n - 2 ) to be closer to P(n), while P(n + 3), P(n+2) and P(n —3) are further away from P(n). In this case, if adjacent leaf primordia affect the development of P(n), then the effect of P(n —2) in the transformants becomes stronger and that of other leaf primordia becomes weaker in comparison to non-transformants with lower PR values. Wardlaw (1949) hypothesized that an inhibitory substance may be secreted from the center of the SAM and leaf primordia and inhibits leaf initiation around the SAM. If such a hypothetical inhibitor also inhibits leaf blade expansion, then the effect of P(n —2) on the development of the P(n) leaf blade would be greater than that of other leaf primordia. In this situation, leaf primordia might develop to avoid the inhibitory substance at P(n —2), resulting in leaves on the transformants curving in the same direction as their generative spiral. Such an inhibitory substance derived from leaf primordia has not yet been identified; however, it would be a good candidate for explaining the relationship between the direction of leaf curvature and the direction of phyllotaxy. The value of PR is defined as a measure of the radial expansion of the shoot apex during a plastochron, that is, the initiation of successive leaf primordia (Steeves and Sussex 1989, Callos and Medford 1994). Thus, an increase in the PR value is interpreted as an increase in the radial growth of the shoot apex during a plastochron. In other word, it is expected that the extent of the SAM of transformants is greater than that of wild-type plants. This resembles an Arabidopsis mutant, clavatal (clvl); the SAM of this mutant is significantly larger than that of wild-type plants (Leyser and Furner 1992, Clark et al. 1993). In con- Developmental interaction between the SAM and leaf 666 trast, plants with mutation at the SHOOT MERISTEMLESS (STM) locus show the opposite phenotype of clvl plant because the weak stm allele has a small SAM consisting of a small number of cells (Clark et al. 1996, Endrizzi et al. 1996). Genetic analyses of CLKand STM interaction reveal that the size of SAM is regulated, in part, by balancing CLV and STM levels (Clark et al. 1996). According to this hypothesis, when the STM level is larger than the CLV one, the meristem size might increase. The STM gene is a member of KN1 -type homeobox gene (Long et al. 1996); thus, it is possible that NTH1 may have a function similar to that of this gene. If so, over-expression of NTH 1 causes unbalancing between NTH1 level and the level of the CLV counterpart in tobacco; this would cause the extension of the SAM that is identified as increasing of the PR value in this experiment. The availability of transformants with similar phenotypes allowed us to notice an interesting relationship between the direction of curvature of malformed leaves and the direction of the order of leaf formation. This relationship was also observed in mild-phenotype tobacco plants transformed with other tobacco homeobox genes, such as NTH9, NTH15, and NTH22 (Tamaoki et al. 1997, Nishimura et al. unpublished result), indicating that it does not depend specifically on some characteristic of NTH1 but it is a common feature of transformants carrying KNl-type homeobox genes. Such a relationship between the order of leaf formation on a stem and leaf development is also observed in other plant species in the natural context. For example, pine trees develop a set of two needles bundled with several scale leaves. The needles twist in the same direction; this direction depends on the direction of the generative spiral of the scale-leaves (Schmucker 1924). A more specialized example is seen in Cyperaceae. One species in this family forms leaves curved in the same direction as the spiral arrangement of leaves on the stem (Moore and Clark 1995). These observations indicate that developmental interaction between leaves and the SAM is a general phenomenon in leaf development and is not limited to plants transformed with KNl-type homeobox genes. Whether a relationship between the direction of curvature of malformed leaves and the direction of the generative spiral in other species results from over-expression of class 1 gene homologues remain to be determined. This work was supported by a Grant-in Aid for Scientific Research on Priority Areas (The Molecular Basis of Flexible Organ Plans in Plants) from the Ministry of Education, Science and Culture (Japan), and a Research Fellowship of the Japan Society for the Promotion of Science for Young Scientists to M.T. References Affolter, M., Schier, A. and Gehring, W.J. (1990) Homeodomain proteins and regulation of gene expression. Curr. Opin. Cell Biol. 2: 485-495. Callos, J.D. and Medford, J.I. (1994) Organ positions and pattern formation in the shoot apex. Plant J. 6: 1-7. Clark, S.E., Jacobson, S.E., Levin, J. and Meyerowitz, E.M. (1996) The CLAVATA and SHOOT MERISTEMLESS loci competitively regulate meristem activity in Arabidopsis. Development 122: 1567-1575. Clark, S.E., Running, M.P. and Meyerowitz, E.M. (1993) CLA VA TA1, a regulator of meristem and flower development in Arabidopsis. Development 119: 397-418. Endrizzi, K., Moussain, B., Haecker, A., Levin, J.Z. and Laux, T. (1996) The SHOOT MERISTEMLESS gene is required for maintenance of undifferentiated cells in the Arabidopsis shoot and floral meristems and acts at a different regulatory level than the meristem genes WUSCHEL and ZWILLE. Plant J. 10: 967-979. Freeling, M., Bertrand-Garcia, R. and Sinha, N. (1992) Maize mutants and variations altering developmental time and their heterochronic interactions. BioEssays 14: 227-236. Gerber, H.P., Seipel, K., Georgiev, O., Hofferer, M., Hug, M., Rusconi, S. and Schaffner, W. (1994) Transcriptional activation modulated by homopolymeric glutamine and proline stretches. Science 263: 808-811. Jackson, D., Veit, B. and Hake, S. (1994) Expression of maize KNOTTED-1 related homeobox genes in the shoot apical meristem predicts patterns of morphogenesis in the vegetative shoot. Development 120: 405-413. Kano-Murakami, Y., Yanai, T., Tagiri, A. and Matsuoka, M. (1993) A rice homeotic gene, OSH1, causes unusual phenotypes in transgenic tobacco. FEBS Lett. 334: 365-368. Kerstetter, R., Vollbrecht, E., Lowe, B., Veit, B., Yamaguchi, J. and Hake, S. (1994) Sequence analysis and expression patterns divide the maize knottedl-hke homeobox genes into two classes. Plant Cell 6: 1877-1887. Leyser, H.M.O. and Furner, I.J. (1992) Characterization of three shoot apical meristem mutants of Arabidopsis thaliana. Development 116: 397-403. Lincoln, C , Long, J., Yamaguchi, J., Serikawa, K. and Hake, S. (1994) A knotted l-Mkz homeobox gene in Arabidopsis is expressed in the vegetative meristem and dramatically alters leaf morphology when overexpressed in transgenic plants. Plant Cell 6: 1859-1876. Long, J.A., Moan, E.I., Medford, J.I. and Barton, M.K. (1996) A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 379: 66-69. Matsuoka, M., Ichikawa, H., Saito, A., Tada, Y., Fujimura, T. and Kano-Murakami, Y. (1993) Expression of a rice homeobox gene causes altered morphology of transgenic plants. Plant Cell 5: 1039-1048. Moore, R. and Clark, W.D. (1995) Plant form and Function. In Botany. Vol. 1, pp. 276. Wm. C. Brown Publishers, U.S.A. Poethig, R.S. and Sussex, I.M. (1985) The developmental morphology and growth dynamics of the tobacco leaf. Planta 165: 158-169. Richards, F.J. (1948) The geometry of phyllotaxis and its origin. Symp. Soc. Exp. Biol. 2: 217-245. Richards, F.J. (1951) Phyllotaxis: its quantitative expression and relation to growth in the apex. Phil. Trans. R. Soc. London, Ser. B 235: 509-564. Sato, Y., Tamaoki, M., Murakami, T., Yamamoto, N., Kano-Murakami, Y. and Matsuoka, M. (1996) Abnormal cell divisions in leaf primordia caused by the expression of the rice homeobox gene OSH1 lead to altered morphology of leaves in transgenic tobacco. Mol. Gen. Genet. 251: 1322. Schmucker, T. (1924) Rechts-und Linkstendenz bei Pflanzen. Beih. Bot. Zentralbl. 41: 51-81. Schneeberger, R.G., Becraft, P.W., Hake, S. and Freeling, M. (1995) Ectopic expression of the knox homeo box gene rough sheathl alters cell fate in the maize leaf. Genes Dev. 9: 2292-2304. Sharman, B.C. (1943) Tannic acid and iron alum with safranin and orange G in studies of the shoot apex. Stain Techno!. 18: 105-111. Sinha, N.R., Williams, R.E. and Hake, S. (1993) Overexpression of the maize homeo box gene, KNOTTED-1, causes a switch from determinate to indeterminate cell fates. Genes Dev. 7: 787-795. Snow, M. and Snow, R. (1959) The dorsiventrality of leaf primordia. New Phytol. 58: 188-207. Steeves, T.A. and Sussex, I.M. (1989) Patterns in Plant Development. 2nd edition. Cambridge University Press, Cambridge, U.K. Developmental interaction between the SAM and leaf Sussex, I.M. (1955) Morphogenesis in Solarium tuberosum L.: experimental investigation of leaf dorsiventrality and orientation in the juvenile shoot. Phytomorphology 5: 286-300. Tamaoki, M., Kusaba, S., Kano-Murakami, Y. and Matsuoka, M. (1997) Ectopic expression of a tobacco homeobox gene, NTH15, dramatically alters leaf morphology and hormone levels in transgenic tobacco. Plant Cell Physiol. 38: 917-927. Tamaoki, M., Tsugawa, H., Minami, E., Kayano, T., Yamamoto, N., 667 Kano-Murakami, Y. and Matsuoka, M. (1995) Alternative RNA products from a rice homeobox gene. Plant J. 7: 927-938. Vollbrecht, E., Veit, B., Sinha, N. and Hake, S. (1991) The developmental gene Knotted-} is a member of a maize homeobox gene family. Nature 350: 241-243. Wardlaw, C.W. (1949) Experiments on organogenesis in ferns. Growth (Suppl.) 13: 93-131. (Received February 5, 1999; Accepted April 16, 1999)