Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Vol.
3, 439-448,
March
Clinical
1997
8-Chioro-cyclic
Factor
Production
Caterina
Bianco,
Gustavo
Baldassarre,
Gabnelia
Fontanini,
A.
AMP
Raffaele
Giampaolo
Bianco,
Inhibits
Autocrine
and
Angiogenic
in Human
Colorectal
and
Breast
Caputo,
Silvana
Chine,
and
Fortunato
Growth
proliferation
Ciardiello2
Cattedra di Oncobogia Medica, Dipartimento
di Endocnnobogia
e
Oncologia
Molecolare
e Clinica, Facolt#{224}
di Medicina
e Chirurgia,
Universit#{224}degli Studi di Napoli Federico II, Via S. Pansini 5, 80131
Napoli [C. B., G. I., G. B., R. C., A. R. B., F. C.], and Cattedra di
Anatomia
e Istologia Patobogica,
Facolt#{224}
di Medicina
e Chirurgia,
Universit#{224}di Pisa, 56100 Pisa [G. F., S. C.], Italy
ating
and
cells
generally
maintaining
growth
factors
factor
these,
growth
factor
a,
colon
and
that
can
paracrine
peptides
TGF-a,
have
or
RNA
cancer
cell
identified
These
stimulate
DNA
growth
endothelial
cell
acts
in mammalian
isoforms
and PKAII
regulatory
of PKA,
have identical
subunits
(defined
respectively;
Ref.
15).
PKAII has been correlated
transformation.
In
in normal
cells
termed
is a novel anticancer
drug that inhibits
the
of various
autocrine
and paracrine
tumor
growth factors that are important
in sustaining
autonomous
local growth
and facilitate
invasion
and metastasis.
level
Received
9/30/96; revised 12/9/96; accepted
12/18/96.
The costs of publication
of this article were defrayed
in part by the
payment of page charges. This article must therefore be hereby marked
advertisement
indicate
1
This
in accordance
with
18 U.S.C.
Section
1734
solely
to
this fact.
study
was
supported
by grants
from
the Associazione
Ia Ricerca
sul Cancro
and from Progetto
Finalizzato
Cliniche della Ricerca Oncologica,
Consiglio
Nazionale
2 To whom
requests
for reprints should be addressed.
7462061;
Fax: 39-81-7462066.
Italiana
per
Applicazioni
delle Ricerche.
Phone: 39-8 1-
regulatory
Rh
the
vessels,
plays
spreading
(10,
VEOF,
TGF-a,
a
1 1).
and
of angiogencells
through
by binding
fact,
preferential
and
paracrine
demonstrated
to either
PKAII
of two
(15).
PKAI
but differ
in the
and Rh in PKAII,
of
expression
tissues
and
PKAI
and
and neoplasof
PKAII
is
in growth-arrested
levels of PKAI
are detected
in tumor
following
exposure
to mitogenic
stim-
in human
subunit
expression
and
catalytic
subunits
as RI in PKAI
nonproliferating
overexpressed
by 8-Cl-cAMP
various
human
have
Angiogenesis,
blood
Differential
expression
with cell differentiation
cells, whereas
enhanced
cells and in normal
cells
generally
by
human
by the tumor
PKAI
RIa
that
colorectal
factors
(10).
cAMP
production
demonstrate
play
inhibits
growth
8-Cl-cAMP
results
and
regulators
are produced
tumors
(16). 8-Cl-cAMP
is a site-selective
specifically
inhibits
PKAI by facilitating
These
prolifAmong
growth
(6-9).
of new
staining
cells.
their
breast
approach
uli (16). Moreover,
we have shown
role in the transduction
of mitogenic
blood
in
of tumor
and CRIPTO
of these
amphiregulin,
CRIPTO,
basic fibroblast
growth factor, and
vascular
endothelial
growth factor production
by the tumor
cells, and of neoangiogenesis,
as detected
by factor VIII
of host
relaxation
regulate
AR,
as potential
proteins
normal
mechanisms
found
exogenous
mechanisms.
and rnetastatic
of synthesis
to the formation
been
esis (12-14).
distinct
The
2). Cancer
,
to the ability
role in tumor survival
and metastatic
growth
factors,
such as bFGF,
P1GF,
tic
cells.
in part
and
primary
breast
leading
central
Several
for
cellular
in initi-
role in the pathogenesis
of human
colon and breast
TGF-a,
AR, and CRIPTO
are expressed
by the
antisense
process
(1
requirement
factors
(4, 5). Inhibition
a specific
can
growth
autocrine
of human
cancers
transformation
to normal
is due
the EGF3-related
majority
transforming
as compared
through
that regulate
normal
and that are important
a decreased
dependency
to synthesize
eration
8-Chboro-cyclic
AMP (8-Cl-cAMP)
is a cAMP analogue
that specifically
down-regulates
type I protein kinase
A, a
signaling
protein directly involved
in cell proliferation
and
neoplastic
transformation,
and that causes growth
inhibition
in a variety of human cancer cell types. In this report, we
have investigated
the effects of 8-Cl-cAMP
on the expression
of several
growth
factors
in human
colon
(GEO
and
LS174T)
and breast (MDA-MB468)
cancer cell lines. 8-ClcAMP
treatment
caused
in the three cancer
cell lines a
significant
doseand time-dependent
inhibition
in the expression
of various
endogenous
autocrine
growth
factors,
such as transforming
growth
factor a, amphiregulin,
and
CRIPTO,
and of two angiogenic
factors,
such as vascular
endothelial
growth factor and basic fibroblast
growth factor,
at both the mRNA and protein levels. Furthermore,
8-ClcAMP treatment
markedly
inhibited
the ability of all three
cell lines to invade a basement
membrane
matrix in a chemoinvasion
assay.
Finally, 8-Cl-cAMP-induced
inhibition
of
GEO tumor growth
in nude mice was accompanied
by a
of
Growth
neoplastic
exhibit
growth
cells
ABSTRACT
suppression
439
Cancer’
factors
are proteins
and differentiation
an important
cancers
(3).
significant
Research
INTRODUCTION
Tortora,
Rosa
Cancer
while
(19).
recently
signals
cancer
up-regulating
Decreased
that PKAI plays
(17, 18). PKAI
cell
lines
in a Phase
and primary
cAMP
analogue
that
the degradation
of the
at the transcriptional
expression
of RIa
treatment
is associated
with growth
cancer cell lines in vitro and in vivo
recently
a
is
I clinical
induced
inhibition
(20-24).
trial
that
in
We
8-Cl-
The abbreviations
used are: EGF, epidermal
growth
factor: TGF,
transforming
growth factor; AR, amphiregulin;
bFGF, basic fibroblast
growth factor; P1GF, placenta growth factor; VEGF. vascular endothehal growth factor; cAMP, cyclic AMP; 8-Cl-cAMP,
8-chboro-cAMP;
PKA,
cAMP-dependent
protein
kinase;
FBS, fetal bovine
serum;
HUVE,
human
umbilical
vein endothelial;
ECGF,
endothelial
cell
growth factor; CM, conditioned
medium;
mAb, monoclonal
antibody;
EGFR, epidermal
growth factor receptor.
3
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
440
Inhibition
of Growth
cAMP
can
patients
be
to
the
with
regulation
and
in the
bFGF
with
membrane
cAMP
tumor
factors
and
8-Cl-cAMP
in nude
AND
(pH
HUVE
Nazionale
4 msi
heat-inactivated
Milan,
Italy),
were
kindly
Difco
agar-complete
Dickinson)
at
mixture
of
10% heatpenicillin,
(ICN).
Early-
Naples,
supplemented
Italy).
ECGF
(Sigma
Chemical
(Sigma),
20 mM HEPES
Co.,
(pH
streptomycin,
and
4 msi
the cells
Kingdom)
human
CRIPTO
Berlex
Biosciences,
scribed
previously
(5
plated
Milan,
in six
Italy)
and
were
trypsinized
(Difco,
were
Detroit,
was
and
in 24 multiwell
with
the cells
colonies
different
seeded
MI)
layered
in
cluster
medium.
and
PBS
stained
were
counted
with
Fragmentation.
tential induction
of programmed
were treated
every
48 h with
of the
cell
Isolation
RNA
was
isolated
the po-
total
RNA
were
(26).
and
Northern
electrophoresed
agarose/2.2
M formaldehyde
gel.
the gels showed
that equivalent
Blotting.
previously
through
Ethidium
amounts
(23).
Total
Twenty
a denaturating
bromide
of RNA
deKDR,
G.
Viglietto
concentrated
CM
by SDS-PAGE
Milan,
with
or a rabbit
CA)
on
Italy),
trans-
either
a rabbit
(Preprotech,
provided
London,
polycbonal
anti-
by Dr. R. Harkins;
at 1 :500
dilution,
as de-
proteins
were
blotting
kit (Amersharn
Western
Co.)
visualized
cellular
p.g of
1.2%
staining
of
were con-
CM.
grown
For
rpm
diluted
for
10 mm
and
in 70%
(50
ethanol.
temperature
p.g/ml
analysis
cytometer
(Becton
bers
(Becton
35-nim
Biocoat
with
different
cells
were
dishes
concentrations
serum-free
medium
containing
from NIH-3T3
mouse fibroblasts
the chamber
lower compartment
in a humidified
cells on
removed,
atmosphere
dish
and
0.1%
invasion
BSA
was
charn-
(33).
were
96
h, the
in the upper
serum-free
in
Chemical
compartment
CM obtained
was used as chernoattractant
(33). After incubation
for
of 95%
in
48 h
resuspended
(Sigma
air and 5% CO2
the upper
surface
of the filters
were
and the filters were fixed and stained.
GEO,
plated
every
After
PBS,
(26).
assay
and treated
with
cell
a FACScan
previously
cells/dish)
twice
Co.), and placed
(5 X l0 cells/dish)
of the invasion
chamber.
Concentrated
DNA
previously
of 8-Cl-cAMP.
washed
for
staining
chemoinvasion
Dickinson)
with
iodide
with
as reported
p.g/ml
twice
incubated
in PBS).
(l0
stored
in PBS
of 200
were
35-mm
were
and
Cells
(106)
at
CM
resuspended
washed
cells
(Becton
trypsinized,
debris.
concentration
as described
MDA-MB468
culture
cell
trypsinized,
Matrigel
Dickinson),
and
medium
and centrifuged
medium
The
GEO
Chemical
were
Dickinson)
CM,
concentrations
in duplicate
Assay.
using
with
GEO
(Sigma
iodide
performed
washed
or with
in 1 ml of propidiurn
propidium
were
Chemoinvasion
performed
were
then
cell
PBS
collected
at a final
cells
ceased
phases
24 h in serum-free
precipitates
cells
without
24 h, either
different
with
then
with
cells
were
of GEO
twice
FBS
heparin
acetone:serum-free
HUVE
30 mm at room
and
at 4#{176}C
to eliminate
to HUVE
fixed
cells
in
twice
in the G0-G
or without
were
at -20#{176}C.The
proteins.
10%
HUVE
for an additional
in a 2: 1 (v/v)
overnight
containing
preparation
CM
washed
conditions,
ECGF
cells
Dickinson)
for an additional
for 96 h, rinsed
8-Cl-cAMP.
were
48 h. HUVE
in CM with
HUVE
(Becton
accumulation
containing
and incubated
without
these
and incubated
of 8-Cl-cAMP
total
in DMEM
underwent
Cells.
dishes
24 h, the cells
Under
medium
were
of HUVE
in 35-mm
within
PBS
serum-free
flow
cell death by 8-Cl-cAMP,
cells
various
concentrations
of 8-Cl-
as described
dilution
(kindly
After
cycle
with
cells
of
as described
plated
and
cell
dishes
1, 3, or 4 days, both adherent
and detached
cells
and lysed, and DNA was extracted
and electro-
as described
or
antiserum
ECL
incubated
to proliferate
cycle
nitroblue
To evaluate
lysates
separated
Richmond,
and heparin.
ECGF
0.5 ml
concentrations
were
Dr.
and incubated
VEGF
Distribution
complete
LS174T,
of DNA
filters,
(30),
VEGF,
by
(7). Irnmunoreactive
were
solution
in com-
over
for human
Laboratories,
at 1 :500
Cycle
and added
of 8-Cl-cAMP.
103/well)
X
agar
treated
(Bio-Rad
FLT-l
Pascale).
were
antiserum
(105/dish)
PBS,
suspension
and
were
Dickinson,
Noble
12 days,
(Sigma),
(104/well)
(7).
cAMP.
After
were harvested
United
(29),
hy-
probes:
3’-phosphate
provided
Protein
anti-human
complete
by Dr. G. Viglietto
concentrations
medium
kindly
to nitrocellulose
twice
20%
This
and
After
Analysis
RNA
CO2
with
(Becton
different
medium.
(Becton
phoresed
King-
5%
100 lU/mb
Pascale,
i.g/ml
of treatment,
of 0.8%
previously
United
air and
KDR
probes
protein/lane)
gels
Cell
100 .tg/ml
Irvine,
provided
Cells
dishes
0.5 ml of 0.3%
tetrazolium
maintained
penicillin,
(28),
and
eDNA
Corp.).
heat-inactivated
glutamine
in DMEM
Growth.
6 days
8-Cl-cAMP.
growth
were
IL)
human
glyceraldehyde
cDNA
Blotting.
precast
ferred
VEGF
to Hybond-N
Heights,
Tumori-Fondazione
pg of total
3000
48 h with
culture
were
10%
7.4),
4 mi
100
P1GF
polycbonal
complete
grown
in a 1:1 (v/v)
supplemented
with
counted
with a hemocytorneter.
Soft Agar Growth.
Cells
plete
cells
(ICN,
(pH
and
penicillin,
cluster
4 and
endogenous
of 95%
FBS, 100 p.g/rnl
100 p.g/rnl heparmn
every
After
glutamine
cultured
100 IU/ml
treated
an almost
of
Turnori-Fondazione
glutarnine.
Monolayer
multiwell
with
100 lU/mb
cells were
F12 medium
were
8-Cl-
The
transferred
32P-labeled
and
by a chemiluminescence
atmosphere
cells
cells
7.4),
7.4),
streptomycin,
(Istituto
Finally,
Arlington
(7),
Nazionale
10%
of well-established
with
20 mM HEPES
passage
HUVE
mice
production
supplemented
in a humidified
FBS,
assay.
GEO and LS 174T
and
the basement
were
Corp.,
(27),
(32).
Western
(50
METHODS
medium
i.g/ml
to invade
CRIPTO
1 , and
VEGF,
samples
following
3-actin
hydrogenase
(Istituto
treatment
The
the
(27),
(31),
FLT-
down-
peptides.
37#{176}C.
MDA-MB468
DMEM
and Ham’s
100
cells
with
PIGF
and
lane.
(Amersharn
TGF-ca
Treatment
CRIPTO,
the growth
cell
(GEO
time-dependent
in a chemoinvasion
in inhibiting
xenografts
streptornycin,
and
of these
of 8-Clautocrine
cell lines.
AR,
angiogenic
inactivated
colon
human
Moreover,
20 mM HEPES
dom)
in
of TGF-a,
Cell Cultures.
FBS,
factors
the ability
MATERIALS
effects
of several
of cancer
in McCoy’s
the
production
cancer
in each
membranes
bridized
production
matrix
suppression
investigated
lines.
tamed
allow
potential
(25).
a dose-
cell
that
the
and
(MDA-MB468)
is effective
GEO
doses
determines
in all
interferes
at
within
inhibition
have
growth
and breast
patients
concentrations
we
8-Cl-cAMP
by 8-Cl-cAMP
cancer
expression
angiogenic
LS l74T)
to
for growth
study,
on
Production
plasma
range
In this
and
given
reach
therapeutic
cAMP
Factor
in
18 h
at 37#{176}C,
the
mechanically
Ten fields per
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
Clinical
each
experimental
point
were
counted,
as described
Two
previously
(G.
(34).
GEO
female
Xenografts
BALB/c
athymic
Charles
River
was approved,
guidelines
Committee.
Animals
been
Five-
mice
were
injected
were
After
p.1 of
7 days,
Matrigel
when
different
time.
doses
Tumor
diameter
of 8-Cl-cAMP
size
was
X (smaller
measured
using
diameter)2
(35).
Staining.
of
8-Cl-cAMP
nografts
were
containing
peroxidase
blocked
and
for
from
treated
CRIPTO
protein
was
antiserum
with
acid
used
generated
ing to amino
acid
at 1:200
goat
with
the
polycbonal
CA)
dilution
polyclonal
1:200
PC
dom)
was
polyclonal
at 1: 1000
peptide
and treated
antibody
(1:200
Burlingame,
with
PBS,
the
slides
dehydrogenase-biotinylated
rinsed
twice in PBS,
nobenzidine
and
for
were
and
in 0.01%
not cross-react
An anti-VEGF
immunogenic
to + + +) and
percentage
peptide.
cancer
cell
8-Cl-cAMP
was
antigen
used
factor
at
staining,
United
King-
VIII,
a rabbit
was
times
several
30
mm
peroxide.
of immunopositive
semi-
intensity
of the
8-Cl-cAMP
in a wide
used
with
washes
with
The
avidin
slides
intensity
were
cells
rabbit
with the
of staining
were
(-
scored.
determines
variety
of human
lines in vitro and in vivo (20-24).
Treatment
with
for 4 or 6 days induced
a dose-dependent
inhibition
growth
and
in human
GEO)
cancer
20 p.M. respectively.
agar
that
cloning
breast
(MDA-MB468)
cell lines,
A comparable
efficiency
was
with
and colon
an IC50 of 10, 5, and
dose-dependent
caused
in all three
inhibition
of
cell
by
lines
the treatment
with 8-Cl-cAMP
(data not shown).
The
inhibitory
effect induced
by 8-Cl-cAMP
was cytostatic
growthand not
cytotoxic,
Further-
as assessed
by
trypan
blue
dye
up to 25 p.M for 6 days, as evaluated
analysis
of chromatin
fragmentation
exclusion.
(data
not shown;
Ref. 26).
We next investigated
by gel electrophoresis
into nucleosorne
ladders
the effects
of several
of 8-Cl-cAMP
growth
factors
24, and 96 h, and then
Northern
blotting.
expressed
of 4.8
and
cAMP
caused
total
specific
1.6 kb,
and
isolated
nontreated
TGF-a
in TGF-a
time-
60%
able
inhibition
in TGF-a
mRNA
expression
within
6 h of treatment
with 10 p.M
effect
treated
lines
(Fig.
In fact,
1). An almost
on TGF-a
mRNA
with 8-Cl-cAMP
CRIPTO
LS174T
levels
(data
cells
for 6,
analyzed
by
MDA-MB468
of 4.8 kb and
with
mRNA
8-Cl-
levels
that
an approximately
40-
was already
8-Cl-cAMP
complete
detectin both
down-regulation
in GEO and MDAfor 96 h. A similar
was observed
in LSI74T
not shown).
The expression
cells
of
mRNA
was also significantly
reduced
in GEO
and
cells treated
with the cAMP
analogue
(Fig. 2). In fact,
a reduction
detected
of 60-90%
after
8-Cl-cAMP
A 3.9-kb
LS174T,
in CRIPTO
treatment
of GEO
for 4 days
VEGF-specific
and
8-Cl-cAMP
cells
cancer
cell
cAMP
(Fig.
lines
(Fig.
within
3). VEGF
24
and
mRNA
expressed
was
with
marked
levels
25 p.M
cells.
GEO,
cell
lines,
time-
and
as compared
an almost
complete
suppression
in
was observed
in all three
human
h of treatment
interacts
with
two
with
high
cell membrane
receptors,
FLT- 1 and KDR,
intracellular
tyrosine
kinase
activity
(29-30).
specifically
cells
3). In all three
a rapid
in VEGF
to nontreated
cells. In fact,
VEGF
mRNA
expression
expression
LS174T
to control
nontreated
was
detectable
in
determined
inhibition
mRNA
and
as compared
mRNA
MDA-MB468
treatment
dose-dependent
are
in tumor
1). Treatment
was
cell
dose-dependent.
and
transcripts
(Fig.
reduction
and
GEO
rnRNA
respectively
a marked
was
RNA
Control
treatment
involved
and MDA-MB648
of 8-Cl-cAMP
in TGF-a
mRNA
expression
was observed
MB468
cells treated
with 25 p.M 8-Cl-cAMP
Following
Both
(LSI74T
previously
and differentiation
cancer
using either a similar
concentration
of preimmune
or IgG or by adsorbing
the primary
antibody
appropriate
shown
Santa
then rinsed
in distilled
water, counterstained
with hematoxylin,
and mounted.
Nonspecific
staining
was evaluated
for each specimen
serum
have
inhibition
rabbit
anti-bFGF
horseradish
peroxidase
H complex,
incubated
for 2 mm in 0.05%
diamihydrogen
We
growth
cells
secondary
biotinylated
goat
ABC kit; Vector
Laborafor
cells
was
(8).
An
nuclear
reacted
on color
Biotechnology,
To detect
30 mm.
was
the an-
Oncogene
Science,
Each antibody
was
(Dakopatts,
Gbostrup,
Denmark)
Sections
were then washed
three
CA)
based
cancer
staining
precipitate
(+ , light brown
staining;
color; + + + , an intense
brown color),
growth
and progression.
GEO, LS I 74T,
were treated
with different
concentrations
by Dr. W. J. Gullick
United
Kingdom).
Newcastle,
with an appropriate
dilution;
Vectastain
PBS
tory,
dilution.
anti-AR
correspond-
Biotechnology)
Laboratory,
at 1:200
antibody
dilution.
An
and
cell
a score
previously
on the expression
human
protein
Cruz
Cruz
the
anti-CRIPTO
dilution.
proliferation
(Novocastra
used
I :50
(Santa
For
10 mAb
(Santa
at
antibody
dilution.
of
AR, or CRIPTO
and did
EGF-related
peptides
(5).
used
with
of the rat AR
the
1000
Specific
more,
no evidence
of apoptotic
cell death
was detected
in the
three cancer cell lines treated
with 8-Cl-cAMP
at concentrations
primary
(5).
a 17-mer
kindly provided
Fund,
London,
antibody
was
methanol
endogenous
the sections
washed
13
An anti-human
TGF-a
mouse
mAb (Ab-2;
Manhasset,
NY) was used at 1 : 100 dilution.
Cruz,
any
PBS,
xedoses
were used. An antia l7-mer
peptide
cor-
at 1 :400
scored.
by an investigator
At least
RESULTS
soft
tumor
various
serum,
97-1
Both
ti-AR rabbit antisera
were
(Imperial
Cancer
Research
rabbit
GEO
appropriate
residues
159-175
(5).
specific
for TGF-a,
with the other
two
of
X larger
at 20#{176}C
with
10%
against
residues
dilution
ir/6
i.p. with
for 30 mm
overnight
amino
period
concentrations
peroxide
to block
several
washes
with
mm
to
indicated
different
at 4#{176}C.
The following
antibodies
rabbit antiserum
raised
against
responding
used
45
as described
of monolayer
sections
mice
incubated
incubated
antibody
CRIPTO
rabbit
tumor
0.3% hydrogen
activity.
After
xe-
0.30-0.35
cm3
with i.p. injections
the formula
with
in vitro
s.c. in nude
of 8-Cl-cAMP
PBS,
treated
or GEO
grown
were
Slides
cells
MDA-MB468
and
by assigning
evaluated
code.
441
(Collaborative
well-established
the
counted
diaminobenzidine
a moderately
brown
+ + ,
were
Research
that
and Evaluation
of Immunopercontaining
GEO, LS174T,
and
Immunocytochemistry
oxidase
for
brown
sample
to the treatment
were
quantitated
Care
cells
for each
blinded
per slide
from
Animal
i07 GEO
nografts
were detectable
with an approximate
tumor
size, mice were treated
twice weekly
of
purchased
of Naples
s.c. with
in 200
Products).
to 6-week-old
Milan,
Italy. The research
protocol
were
maintained
in accordance
to
of the University
resuspended
Biomedical
Mice.
(nu+/nu+)
Laboratories,
and mice
institutional
had
in Nude
slides
F.)
Cancer
on
endothelial
10
p.M
affinity
8-Cl-
distinct
which
possess
an
These
receptors
cells
and
on
some
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
442
Inhibition
of Growth
A
Production
by 8-Cl-cAMP
A
ir’
kb
4.8
Factor
;
“
4
-
2.2 kb -
‘-I.--
B
*Iji#{149}
kb -
4.8
4
1.6 kb-
B
.-.
kb -
2.2
l*frJL.
*,
.
I
I
234567
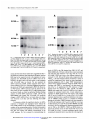
Fig. I Northern
blot analysis
of TGF-a
mRNA
expression
in GEO (A)
and MDA-MB468
(B) cells. A: Lane 1, control
nontreated
GEO cells;
Lanes
2-4,
GEO cells treated with 10 p.M 8-Cl-cAMP
for 6, 24, and
96 h, respectively;
Lanes 5-7, GEO cells treated with 25 p.M 8-Cl-cAMP
for 6, 24, and 96 h, respectively.
B: Lane
1, control nontreated
MDAMB468
cells; Lanes
2-4,
MDA-MB468
cells treated with 10 p.M
8-Cl-cAMP
for 6, 24, and 96 h, respectively;
Lanes 5-7, MDA-MB468
cells
treated
with
25 p.M 8-Cl-cAMP
for 6, 24, and 96 h, respectively.
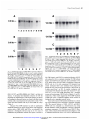
Fig. 2 Northern
blot analysis
of CRIPTO
mRNA
expression
in GEO
(A) and LS174T
(B) cells. A: Lane
1, control
nontreated
GEO cells;
Lanes
2-4,
GEO cells treated with 10 p.M 8-Cl-cAMP
for 6, 24, and
96 h, respectively;
Lanes 5-7, GEO cells treated with 25 p.M 8-Cl-cAMP
for 6, 24, and 96 h, respectively.
B: Lane 1, control nontreated
LSI74T
cells; Lanes 2-4, LS174T cells treated with 10 p.M 8-Cl-cAMP
for 6, 24,
and 96 h, respectively;
Lanes
5-7,
LS174T cells treated with 25 p.M
8-Cl-cAMP
for 6, 24, and 96 h, respectively.
levels
of VEGF
MDA-MB468
cancer
cell lines
(36).
Recent
studies
have
suggested
that
and
KDR have different
signal transduction
properties
KDR
may be the receptor
involved
in the regulation
giogenesis
(37).
Both
and
KDR
FLT-l
rnRNAs
cells
FLT-l
and that
of neoan-
were
not
de-
tectable
in the three cancer
cell lines
with
8-Cl-cAMP
(data
not shown).
before
or after treatment
We next
analyzed
the
mRNA
expression
angiogenic
closely
related
the activity
no
of PIGF,
to VEGF
of VEGF
P1GF-specific
another
(14).
It has been
can be potentiated
mRNA
transcript
three cancer
not shown).
cell lines, regardless
Finally,
as control
analysis
G3PD
of
MDA-MB468
mRNA
cells
suggested
by P1GF
could
be
of 8-Cl-cAMP
we performed
expression
treated
with
factor
recently
(38).
is
that
in the
treatment
(data
Northern
blotting
LS174T,
No
changes
in G3PD
mRNA
(Fig. 4) or 3-actin
(data
were
observed
in all three
cell lines
following
and
not shown)
8-Cl-cAMP
expression
8-Cl-cAMP
whether
the
cAMP
reduction
of mitogenic
and angiogenic
treatment
is also accompanied
synthesis
of the corresponding
immunocytochemistiy
were
lines
significant
that
were
analogue.
treated
We
with
first
proteins,
performed
factors
induced
by
by a decrease
in the
Western
blotting
and/or
on all three cancer
cell
different
evaluated
in rnRNA
concentrations
by
immunoblotting
of
the
the
from
cells
121,
may
165,
obtained
were
inducing
and
angiogenesis
species
with
consistent
an
as four
20]
with
(Fig.
levels
MB468
cells
compared
regulation
25
An
approximate
was
observed
treated
with
8-Cl-cAMP
on cell
lysates,
CRIPTO
50-80%
after
treatment
with
in GEO
cells
(Fig.
shown).
ysis
for
pression
8-Cl-cAMP
nostaining
in all three
We
TGF-a,
in all
a significant
In fact,
6) in LS174T
p.M
and
and
marked
in cells
(Fig.
levels
in the expresby Western
were
8-Cl-cAMP
in MDA-MB468
AR,
CRIPTO,
VEGF,
for 4 days.
for all growth
cell
lines
treated
reduced
by
for 2 or 4 days
an immunocytochemical
and
as
downtreated
8-Cl-cAMP
5).
performed
cancer
MDA-
for 4 or 6 days
next
three
is
cell
in VEGF
LS174T,
inhibition
as evaluated
protein
25
which
tumor
reduction
GEO,
for 4 or 6 days
blotting
not
detected
of
in
immunoreactive
10 p.M 8-Cl-cAMP
determined
also
CRIPTO
protein.
(28).
of 45,000,
to nontreated
control
cells. A more
in VEGF
protein
levels
was found
p.M
p.M
con-
respectively
VEGF
60-70%
treatment
sion of
10 or 25
isoforms
secreted
by a variety
has been
implicated
weight
in
and
nontreated
with
acids,
A major
was
LS 174T,
control
different
amino
molecular
VEGF165,
5).
protein
(28).
apparent
GEO,
from
for 4 or 6 days
exist
189,
from
collected
treated
VEGF
of
CM
CM
VEGF165
is the more frequent
isoform
normal
and transformed
cells,
which
with
significant
treatment.
To determine
and
sisting
in the
cells.
8-Cl-cAMP.
lines
However,
detected
in GEO,
8-Cl-cAMP.
that
234567
bFGF
with
cells
(data
analprotein
10 or
ex25
p.M
A mostly
cytoplasmic-specific
immufactors
was observed
in the majority
of
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
Clinical
A
Cancer
Research
443
A
#{149}#{248}#{216}ii #{149}
3.9kb-
1.4kb-
I
2
3
4
5
6
7
8
9 10
B
B
1.4 kb-
ik
kb-
3.9
C
1.4 kbC
..
c
#{149}
‘
1234567
:
3.9kb-
blot analysis ofglyceraldehyde
3-phosphate
dehydrogenase mRNA expression
in GEO (A), LS174T (B). and MDA-MB468
(C) cells. A: Lane
1, control
nontreated
GEO cells: Lanes
2-4.
GEO
cells treated with 10 p.M 8-Cl-cAMP
for 6, 24, and 96 h. respectively:
Lanes
5-7. GEO cells treated with
25 p.M 8-Cl-cAMP
for 6. 24. and
96 h, respectively.
B: Lane I. control
nontreated
LSI74T cells: Lanes
2-4,
LSI74T cells treated with 10 p.M 8-Cl-cAMP
ftr 6, 24, and 96 h.
respectively:
Lanes
5-7. LS 174T cells treated with 25 p.si 8-Cl-cAMP
for 6, 24. and 96 h. respectively.
C: Lane
1. control
nontreated
MDAMB468
cells: Lanes
2-4.
MDA-MB468
cells treated
with 10 p.M
8-Cl-cAMP
for 6. 24, and 96 h. respectively:
Lanes
5-7. MDA-MB468
cells treated with 25 p.M 8-Cl-cAMP
for 6, 24. and 96 h. respectively.
Fig.
I
234567
3 Northern
blot analysis
of VEGF
mRNA
expression
in GEO
(A).
LSI74T (B), and MDA-MB468
(C) cells. A: Lane
1, control
nontreated
GEO cells: Lanes
2-4.
GEO cells treated with 5 p.M 8-Cl-cAMP
for 6.
24, and 96 h, respectively:
Lanes
5-7. GEO cells treated with
10 p.si
8-Cl-cAMP
for 6, 24, and 96 h. respectively:
Lanes
8-10.
GEO cells
Fig.
treated
with 25 p.M 8-Cl-cAMP
1. control
nontreated
LSI74T
for 6. 24, and 96 h. respectively.
B: Lane
cells: Lanes
2-4,
LS174I
cells treated
for 6, 24, and 96 h. respectively:
Lanes
5-7.
with 10 p.M 8-Cl-cAMP
L51741
cells treated with 25 p.51 8-Cl-cAMP
ftr 6. 24. and 96 h.
respectively.
C: Lane I. control nontreated
MDA-MB468
cells: Lanes
2-4.
MDA-MB468
cells treated with 10 p.M 8-Cl-cAMP
for 6, 24, and
96 h, respectively:
Lanes
5-7, MDA-MB468
cells treated with 25 p.M
8-Cl-cAMP
for 6, 24, and 96 h, respectively.
4
Northern
20%
FBS,
heparin,
LS 174T,
percentage
MDA-MB468
lines
with
cells
(Table
I).
Because
activity,
reduction
endothelial
a weak
after
(Table
factors
and
was
expression
treatment
cells
control
nontreated
through
S phase
the cell
raised
for
found
25
increase
the
respectively).
p.M
protein
of GEO
in only
8-Cl-cAMP
step
the production
such
as VEGF,
treatment
of various
bFGF,
is biologically
cell
proliferation.
was
growth
able
factors
and TGF-a.
relevant
HUVE
to significantly
using
cells
with
an
in
require
if this
their
obtained
from
GEO
cells
from
progression
of HUVE
incubation
cells in
with
for 4 days
These
results
causes
a significant
active
The
angiogenic
cell invasion
invasive
suggest
that
8-Cl-cAMP
inhibition
growth
of the basement
membrane
of
blasts,
which
membrane
that
leads
to metastatic
behavior
of
GEO,
LS174T,
a mixture
of
as undefined
(33).
contains
defined
chemotactic
CM
from
mouse
(fibronectin
factors,
was
of
factors.
process
components
treatment
in the secretion
is a critical
spreading
and
cells was evaluated
in a chemoinvasion
chambers
coated
with Matrigel,
a mixture
assay
vitro
stimulated
obtained
a moderate
( I I and 8%,
cells
MB468
Boyden
angiogenic
we evaluated
cells
of quiescent
CM
because
the percentage
5 to 20%.
In contrast,
in the multistage
(39).
8-Cl-cAMP
2). Incubation
concentrated
treated
Tumor
5-25%
for 4 days
cycle
from
(Table
without
in the
25 p.M 8-Cl-cAMP
determined
only
in the percentage
of HUVE cells in S phase
biologically
cancer
h with
GEO
CM
not
in the three
cell cycle
24
HUVE
10% FBS
accumulated
10 or
data
of specific
proliferation.
ofthe
phases
HUVE
in both
the intensity
of each
with
I and
reduction
cells
for all growth
tumor
inhibit
cells
dose-dependent
of immunoreactive
cell staining
cell
and
A substantial
for optimal
G-G1
with
shown).
ECGF
for 24 h in a medium
containing
ECGF
became
quiescent
and
concentrated
GEO,
and
cells cultured
heparin
and
NIH-3T3
and collagen)
used
MDA-
assay
using
of basement
fibroas well
as chemoattractant
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
444
Inhibition
of Growth
Factor
Production
by 8-Cl-cAMP
A
.
45 kDa-
I’1W
*
-
kDa
34
-R
---i:L
B
1
45 kDa-
3
control
grew
carcinomas
_,
#
‘-fl
,,w
-.
shown).
of
suppression
proliferation
cAMP
cAMP
not
inhibition
complete
-
untreated
and 8-Cl-cAMP-treated
as moderately
differentiated
human
(data
dependent
cell
8-Cl-cAMP
GEO
tumor
of tumor
nuclear
cell
antigen
(data
8-Cl-cAMP
I
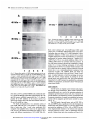
Western blot analysis of VEGF protein expression
in the conCM from GEO (A), LSI74T (B), and MDA-MB468
(C) cells.
A: Lane
1, control
nontreated
GEO cells; Lanes 2 and 3, GEO cells
treated for 4 or 6 days with 10 p.M 8-Cl-cAMP;
Lanes
4 and 5, GEO
cells treated for 4 or 6 days with 25 p.M 8-Cl-cAMP.
B: Lane 1, control
nontreated
LS174T cells; Lanes 2 and 3, LSI74T cells treated for 4 or
6 days with 10 p.M 8-Cl-cAMP;
Lanes
4 and 5, LS174T cells treated for
4 or 6 days with 25 p.M 8-Cl-cAMP.
C: Lane 1, control nontreated
MDA-MB468
cells; Lanes 2 and 3, MDA-MB468
cells treated for 4 or
6 days with 10 p.M 8-Cl-cAMP;
Lanes
4 and 5, MDA-MB468
cells
treated for 4 or 6 days with 25 p.M 8-Cl-cAMP.
5
LS l74T,
ity to invade
and
200
the Matrigel-coated
invasive
pendent
40
and MDA-MB468
to
filter
cells/field,
60%
inhibition
of
flank
of
xenografts
staining
cm3,
with
the animals
different
the abil1 80, 150,
7).
GEO cells and the
growth
factors
were
induced
by 8-Cl-cAMP
Growth
through
uncontrolled
endogenous
growth
factors
several
inhibitors;
to paracrine
(1, 2).
The EGF
the in vivo antitumor
activity
of
were injected
s.c. into the dorsal
AR,
and CRIPTO,
were
treated
twice
weekly
of 8-Cl-cAMP
i.p. for 4
(Fig.
8A).
to controls
8-Cl-cAMP
treatment
with
of growth
evaluation
bFGF,
and
of
VEGF
the
was
intensity
of staining
for all five
(Fig. 8B). The growth
inhibition
treatment
was paralleled
as detected
by a substantial
by factor
VHI
chymal
human
cells
breast
and
stimulation
family
plays
of tumor
of normal
by angiogenic
and
cancer
initiation
These
include:
as a result of the autocrine
factors
by tumor cells; loss
stimulation
cells
concentrations
effect
of 8-Clcytotoxic.
In
at the end of the 4 weeks
of treatin both the percentage
of positive
can regulate
mechanisms.
growth
growth
was
established
GEO
of approximately
by
8-Cl-
staining
of
DISCUSSION
10 or
treatment
than
the production
CRIPTO,
average
observed
inhibition
of angiogenesis,
mouse
blood vessels.
with
after
invasive
A dose-de-
AR,
almost
at 2 mg/dose
immunohistochemical
vivo,
an
as assessed
whether
with
tumor
adenoa dose-
with
rate comparable
termination
of
To ascertain
to interfere
of TGF-a,
activity
lines
the
Fig.
nude
mice.
After
1 week,
when
were palpable
with a tumor
size
0.30-0.35
weeks
exhibited
(approximately
respectively;
observed
in all three cancer cell
25 p.M 8-Cl-cAMP
for 4 days.
We finally
investigated
8-Cl-cAMP.
GEO cells (l0)
cells
in
able
performed
on GEO tumors
rnent.
A marked
reduction
sion
GEO,
also
expression
centrated
not shown).
was
factors
2345
determined
growth
(Fig. 8). The in vivo growth-inhibitory
on GEO tumors
was cytostatic
rather
treatment
(40).
5
GEO
colon
proliferation,
fact, GEO tumors
resumed
a growth
within
10 to 14 days
following
Fig.
4
Western
6
Both
groups
P?!T_
C
45 kDa
2
blot analysis of CRIPTO
protein expression
in GEO
cells. Lane 1, control nontreated
GEO cells; Lanes 2 and 3, GEO cells
treated for 2 or 4 days with 10 p.M 8-Cl-cAMP;
Lanes
4 and 5, GEO
cells treated for 2 or 4 days with 25 p.M 8-Cl-cAMP.
Fig.
host
growth
of growth
a significant
coborectal
cancer
role
(3).
production
of sensitivity
vascularization
endothelial
factors
factors,
and progresautonomous
such
of human
breast
and
colon
mesen-
by cancer
as EGF,
TGF-a,
in the development
We
have
carcinoma
cell
of
demonstrated
previously
that inhibition
of TGF-a,
AR, or CRIPTO
sense oligonucleotide
treatment
or by infection
with
retroviral
expression
vectors
is able to significantly
growth
due
and
secreted
of
to
by antiantisense
block the
lines
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
(6-9).
Clinical
Table
Growth
I
expre ssion
factor
in human
AR
TGF-a
GEO cells
Control
25%
25
15%
(+)
p.M
LS174T
50%
5%
(+)
65%
(+++)
25%
(++)
5%
(+)
30%
(+)
445
bFGF
VEGF
(+++)
Research
treatment”
65%
(+++)
(++)
8-C 1-cAMP
CRIPTO
70%
75%
(+++)
8-Cl-cAMP
10 p.M
cell lines following
cancer
Cancer
(+++)
35%
(++)
10%
(+)
40%
(+)
20%
(+)
cells
60%
Control
65%
(+++)
8-Cl-cAMP
lOp.M
25%
(++)
25 p.M
15%
(+)
MDA-MB468
Control
75%
(+++)
(+++)
(+++)
15%
40%
50%
(+)
(++)
(+)
25%
(+)
5%
(+)
80%
75%
(+++)
55%
(++)
10%
(+)
20%
(+)
cells
80%
65%
70%
(+++)
(+++)
60%
(+++)
65%
(+++)
(+++)
8-Cl-cAMP
10 p.M
25
p.M
The percentage
a
of positive
tumor
20%
35%
(+)
(+)
10%
15%
20%
20%
(+)
(+)
(+)
(+)
cells and the average
60%
intensity
of specific
40%
40%
(++)
(+)
immunostaining
(++)
was determined
20%
(+)
as described
in “Materials
and
Methods.”
2
Cell cycle distribution
with concentrated
CM obtained
Table
of HUVE cells following
from 8-Cl-cAMP-treated
250
incubation
GEO cells
V
4)
Incubation
of HUVE cells with concentrated
CM obtained
from
GEO colon carcinoma
cells treated with the indicated concentrations
of
8-Cl-cAMP
was performed
as described
in “Materials
and Methods.”
Cell cycle distribution
was assessed using propidium
iodide staining and
analyzed with a FACScan flow cytometer.
Data represent the average of
two separate experiments,
each performed
in duplicate.
SD was less than
200
-
150
-
100
-
8
4)
>
C,)
C
5%.
.8
Cell eye le distribut
E
ion (%)
50
-
z
S
G0-G1
88%
74%
Untreated
+GEO CM
+GEOCM+
8-Cl-cAMP
+GEOCM+
8-Cl-cAMP
bFGF
secreted
has
85%
ovary
86%
VEGF
suggested
cells
it does
4%
6%
8%
are potent
types
angiogenic
of human
that
tumor
peptides
malignancies
cells
secreting
that
have
vascularized
tumors
growth-stimulatory
in nude
41).
these
growth
activity
mice,
in
vitro
anti-VEGF
mAb results
size of liver metastases
ing bFGF
A
become
(45).
However,
body
induces
more direct
evidence
of the VEGF
role in tumor-induced
angiogenesis
has been obtained
with the use of specific
anti-VEGF
tumor
neutralizing
by angiogenic
VEGF
in
vitro,
growth
nude
mAbs.
neutralizing
whereas
as xenografts
mice
bearing
In fact,
treatment
antibodies
it exerts
cells
with
anti-
does not affect their proliferation
a potent
inhibitory
effect
on their
in nude
human
of cancer
mice
colon
(43).
carcinoma
Moreover,
xenografts
treatment
with
MDA-MB468
It
whereas
(42).
LSI74T
7
are
(12-13,
have a growth
advantage
in vivo.
In this regard,
of VEGF confers
the ability of Chinese
hamster
to form
not
11%
CEO
Chemoinvasion
assay of GEO, LS 1741, and MDA-MB468
cells. Shown are control nontreated
cells () and cells treated with 10
p.M ()
or with 25 p.M () 8-Cl-cAMP
for 4 days. The cell migration
assay through a Matrigel-coated
filter was performed
as described
in
“Materials
and Methods.”
Values represent the average of two different
experiments,
each performed
in triplicate.
SD was less than 10% for
each point.
Fig.
(25 p.M)
and
factors
may
overexpression
0
7%
6%
(10 p.M)
in several
been
G2-M
5%
20%
of
an
(45).
factors
that
The
specific
aggressive
with
a significant
growth
anticancer
more
treatment
of angiogenic
therapy
in a marked
decrease
(44). Similarly,
tumor
Therefore,
factors
has
cytostatic
cAMP
(46,
metastatic
the
been
proposed
with
more
and
with
signaling
in vivo
neutralizing
of primary
interference
can be combined
drugs
and highly
an anti-bFGF
inhibition
or with
in the number
and
cells overexpressanti-
metastatic
the production
pathways
regulated
as a form
of cancer
conventional
cytotoxic
47).
antiproliferative
analogue
that
effects
inhibits
PKAI
of 8-Cl-cAMP,
expression
a
and
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
446
Inhibition
of Growth
Factor
Production
by 8-Cl-cAMP
8-Cl-cAMP
treatment
levels.
These
results
A
7
previous
6
I
E
C)
observation
mRNA
(23).
a)
E 4
in mouse
the TGF-a
The
the reduction
and further
of a specific
and protein
pressing
5
and precedes
are in agreement
gene
reduction
in protein
extend
our
down-regulation
mammary
following
of TGF-a
epithelial
treatment
in the production
cells
with
of various
overex-
8-Cl-cAMP
peptide
growth
factors
induced
by 8-Cl-cAMP
treatment
does not appear
as a
nonspecific
phenomenon
due to cell growth
inhibition.
In fact, a
3
2- to 4-fold
increase
2
expression
of the
I
and MDA-MB468
with 8-Cl-cAMP
The
0
PKAI
0
5
10
15
20
25
30
35
40
the
results
B
and
This
of TGF-a,
overexpressed
PCNA
Control
TOFu
65%
8O#{176}.’
(+++)
AR
CRIPTO
VEGF
70%
65%
(+++)
(+++)
(+4+)
8-Cl-cAMP:
o5 mg/dose
63%
45%
(++)
35%
(++)
60%
(+++)
I mg/dose
35%
25%
(+)
20%
(++)
40%
15%
15%
(+)
10%
2mg/dose
15%
(+)
bFGF
Factor
VIII
50%
+++
40’.’o
(4)
(4)
30%
20%
(+4)
20%
(+)
10%
10%
(+)
5%
suggest
sustained
clinical
AR,
and
by
which
and
activation
of
TGF-a
++
+
(4)
+1.
(4)
8 A, antitumor
activity of 8-Cl-cAMP
treatment
on established
GEO human colon carcinoma
xenografts.
The mice were injected s.c.
into the dorsal flank with l0 GEO cells as described
in “Materials
and
Methods.”
After 7 days (average tumor size, 0.30-0.35
cm3) the mice
were treated i.p. twice weekly with 8-Cl-cAMP
at the following
doses:
L, 0.25 mg/dose;
0, 0.5 mg/dose; U, 1 mg/dose; and E, 2 mg/dose for
4 weeks. #{149},
control. Each group consisted
of 15 mice. Bars, SD. B,
immunohistochemical
evaluation
of growth factor expression
and of
angiogenesis
in GEO xenografts.
Two mice per group were killed, and
tumors were excised on day 35 after GEO cell injection. The percentage
of positive cancer cells and the average intensity of specific immunostaining were determined
as described
in “Materials
and Methods.”
Fig.
EGFR,
and AR,
vitro
and in vivo
49).
We
growth
have
is the
blocking
may
determine
a synergistic
breast cancer
The inhibition
factors
such
as VEGF
treated
cancer
cells
is functionally
endothelial
GEO
cells
with
and
of HUVE
cell
The
cell
and
proliferation.
lines
(48,
and
combination
an
and
on human
colon
secretion
of angiogenic
in 8-Cl-cAMP
biologically
relevant.
In
of growth
factorto GEO cell CM reare able to stimulate
In contrast,
significantly
growth-stimulating
decreased
cell
EGF,
the in
obtained
and bFGF
8-Cl-cAMP
cancer
effect
fact, analysis
of the cell cycle
distribution
depleted
quiescent
HUVE
cells exposed
veals that GEO cells secrete
factors
that
normal
inhibits
in
growth-inhibitory
for
mAbs
8-Cl-cAMP
used
cell lines (26).
in the synthesis
growth
therapies.
In this
that blocking
the
that
be
as cornrepresent
receptor
human
can
frequently
cancer
recently
mAb
because
are
breast
specific
of several
demonstrated
anti-EGFR
and
that
EGF-related
epitheliurn,
by the use of anti-EGFR
of
blocks
relevance
CRIPTO,
and breast
inhibition
treatment
potential
colorectal
treatment
that
by 8-Cl-cAMP
an important
goal of novel biological
cancer
respect,
several
studies
have demonstrated
(+-t-+)
(4-f)
study
in the
LS174T,
following
signaling
colorectal
dependent
of GEO,
has
in human
to normal
dose
surface
be observed
activity
mitogenic
factors.
pared
could
and
cell
of the present
autocrine
inhibition
is time
on the
cells
(26).
expression
growth
Days
that
EGFR
treatment
inhibits
the
of
secretion
factors.
production
of
autocrine
and
paracrine
growth
factors
induced
by 8-Cl-cAMP
treatment
is paralleled
by
the reduced
ability
of the tumor
cells to invade
the basement
activity,
have
cancer
cell
cently
entered
Phase
has
cycles
widely
in cancer
that
plasma
for tumor
The
results
iv.
and
cancer
cells
The
at doses
that
a range
membrane
rea
therapies
in
allow
the
potentially
for the first
drug
effec-
time
is accompanied
growth
and dose
levels
growth
In fact,
factors
in
inhibition
MB468
cells
cells,
that
tumor
by a
type
nents
(39).
and
inhibition
human
dependent
a significant
is detected
and
colon
probably
reduction
within
of growth
and
cell
of
treatment
for
colon
4 weeks
results
may
enzymes,
basement
membrane
is highly
that
studies
proteolytic
carcinoma
that 8-Clof cancer
process
Further
in conferring
and MDAis observed
suggest
behavior
8-Cl-cAMP
degrade
8-Cl-cAMP
filters
in the multistep
of various
that
are involved
GEO
a dose-dependent
LS 174T,
results
invasive
metastases.
whether
production
and that
growth
event
of distant
to determine
Finally,
of tumor-induced
treatment.
These
interfere
with the
IV collagenase,
In fact,
of GEO,
the Matrigel-coated
is a critical
formation
assay.
in the capacity
to penetrate
which
to the
necessary
demonstrate
cell
in a chemoinvasion
40-60%
after 8-Cl-cAMP
cAMP
may also
several
(25).
peptides
level.
these
In fact,
to standard
8-Cl-cAMP-induced
a transcriptional
for
patients.
administered
of 8-Cl-cAMP
cancer
angiogenic
is time
has
in the synthesis
and secretion
of several
as TGF-a,
AR, CRIPTO,
VEGF,
and bFGF,
modulators
through
autocrine
and paracrine
of human
factors
study
effect
be
within
inhibition
of this
marked
inhibition
growth
factors
such
which are important
neoangiogenesis.
infusion
concentrations
the antiproliferative
pathways
refractory
can
of human
8-Cl-cAMP
in cancer
patients
8-Cl-cAMP
growth
in a variety
Furthermore,
evaluation
of continuous
to reach
demonstrated
(20-24).
clinical
I trial
shown
tive
been
types
inhibit
effective
such
in inhibiting
in
vivo.
8-Cl-cAMP
in a dose-dependent
at
in mRNA
6 to 24 h of
4
F. Ciardiello
and G. Tortora,
unpublished
as
phenotype
breast
occurs
be
the
compo-
the invasive
cells
leads
will
observations.
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
suppres-
the
Clinical
sion
of GEO
nude
mice.
panied
AR,
xenografts
The
growth
by a significant
CRIPTO,
VEGF,
inhibition
of host
staining
of endothelial
in the absence
suppression
in vivo
reduction
in the
and bFGF
in tumor
neoangiogenesis,
In summary,
of toxicity
of tumor
growth
expression
cells
local
and
growth
the
of the
present
study
relevance.
growth
and facilitate
obtained
by treatment
lular targets
and that
can
otherapy
In this
that
13.
of endogenous
sustain
spreading
respect,
be proposed
in cancer
factors
rnetastatic
demonstrate
autonomous
of tumor
long-term
in adjunct
cells
treatment
is
Goldfarb,
M. The
Ferrara,
and
N.,
family
chem-
patients.
G. Borriello
for
helpful
for the excellent
discussions,
technical
and
Dr.
G.
assistance,
Viglietto
providing
early-passage
HUVE cells and the eDNA
VEGF, KDR, FLI- 1 , and PIGF.
for
probes
1 . Aaronson,
2.
Sporn,
Ann.
S. A. Growth
1146-1153,
M. B., and
Intern.
Med.,
3. Salomon,
dermal
I.,
Science
R.,
Rev.
Ciardiello,
peptides
F., and
and
Oncol.-Hematol.,
Stromberg,
K.,
Qi,
their
19:
C-F.,
Normanno,
183-232,
Gullick,,
W.
permeability
16. Cho-Chung,
Res.,
later.
Tahara,
E.,
20. Tagliaferri,
Neckers.
L.,
Selvam,
M.
P.,
Pepe, S.,
a transreduces
cell line.
Basolo,
F.,
G., Pacifico, F., Normanno,
N., Brandt, R., Persico, M. G.,
D. S., and Bianco, A. R. Inhibition ofCRIPTO
expression
and
tumorigenicity
in
human
oligodeoxynucleotides.
8. Normanno.
N.,
colon
Oncogene,
Bianco,
C.,
cancer
cells
9: 291-298,
Damiano,
V.,
by antisense
1994.
de
Angelis,
RNA
E.,
and
Selvam,
I.,
Ally,
R. A., Avery,
G. W., Robins,
I.
I.,
modulation
of gene
mice by site-selective
22.
lortora,
Katsaros,
protein
expression
8-Cl-cAMP.
G., Ciardiello,
I.,
Ally,
Z., Chang,
induces
S., Tortora,
G.,
Y. A., Revanker,
F., Ally,
Y. S. Synergistic
cancer
cell
1642-1650,
G.,
Yokozaki,
Y. S. Inhibition
in human
Cancer
lung
Res.,
S., Clair,
lines
1988.
by
H., Finch,
of growth
and
carcinoma
in athymic
49: 5650-5655,
1989.
I.,
Salomon,
D. S., and
Cho-Chung,
Y. S. Site-selective
8-chloro-adenosine
3,5’ monophosphate inhibits transformation
and transforming
growth factor a production in Ki-ras-transformed
rat fibroblasts.
FEBS
Lett., 242: 363-367,
1989.
23.
Ciardiello,
F., lortora,
G..
Kim,
N.,
Clair,
I.,
Ally,
S., Salomon,
D. S., and Cho-Chung,
Y. S. 8-Cl-cAMP
inhibits transforming
growth
factor a transformation
of mammary epithelial cells by restoration
of the
normal mRNA patterns for cAMP-dependent
protein kinase regulatory
subunit
Chem.,
isoforms
which
265: 1016-1020,
show disruption
1990.
upon
transformation.
J. Biol.
24. Ciardiello,
F., Pepe, S., Bianco,
C., Balda.ssarre,
G., Ruggiero,
A.,
Bianco,
C., Selvam,
M. P., Bianco,
A. R., and Tortora,
G. Downregulation
of RIa subunit
of the cAMP-dependent
protein
kinase
induces
26.
and the esbreast
can-
Y. S. 8-Cl-cAMP
D., Tortora,
L., and Cho-Chung,
antisense
mRNA
inhibits
trogen-induced
proliferation
Physiol.,
to ther-
AMP-dependent
1993.
and colon
human
Cancer
Res., 48:
Bianco,
cer cells. J. Cell.
in growth,
approaches
R. K., and Cho-Chung.
of breast
analogues.
9. Kenney, N. J., Saeki, I., Gottardis,
M., Kim, N., Garcia-Morales,
P.,
Martin, M. B., Normanno,
N., Ciardiello,
F., Day, A., Cutler, M. L., and
Sabomon,
D. S. Expression
of transforming
growth factor a (TGFa)
production
human
of cyclic
D., Clair,
Parandoosh,
growth inhibition
of human mammary
c-Ha-ras
and c-erbB-2
protooncogenes.
estrogen-induced
of estrogen-responsive
156: 497-514,
1993.
proteins
new
and Cho-Chung,
M. P., Grassi, M., Magliulo, G., Tortora, G., Bianco, A. R., Mendelsohn,
J., Salomon, D. S., and Ciardiello,
F. Growth inhibition of human colon
carcinoma
cells by combinations
of anti-EGF-related
growth factor
antisense
oligonucleotides.
Clin. Cancer
Res., 2: 601-609,
1996.
the
Sci.
W. cAMP-dependent
of regulatory
enzymes.
receptors
4: 359-365,
P., Katsaros,
Rubalcava,
B.,
S., Clair,
C.,
& Differ.,
C., Clair,
21.
1992.
Panico, L., D’Antonio,
A., Salvatore, G., Mazza, E., lortora.,
G., Dc
Laurentiis,
M., Dc Placido, S., Giordano,
I., Memo,
M., Salomon,
D. S., Gullick, W. J., Pettinato,
G., Bianco, A. R., and Ciardiello,
F.
Differential
immunohistochemical
detection
of transforming
growth
factor a, amphiregulin
and CRIPTO
in human normal and malignant
breast tissues. Int. J. Cancer, 65: 51-56,
1996.
Bianco,
Acad.
P., and
related
USA, 88:
truncation
and down-regulation
of the RIa subunit
and up-regulation
of
the RIIb subunit
of cAMP-dependent
protein
kinase
leading
to type
holoenzyme-dependent
growth
inhibition
and differentiation
of HL-60
leukemia
cells. J. Biol. Chem., 268: 5774-5782,
1993.
3467-3473,
G.,
Natl.
Delli-Bovi,
for a protein
18. Tortora,
G., Pepe,
S., Bianco,
C., Baldassarre,
G., Budillon,
A.,
Clair, I., Cho-Chung,
Y. S., Bianco,
A. R., and Ciardiello,
F. The RIa
subunit
of protein
kinase
A controls
serum
dependency
and entry into
cell cycle ofhuman
mammary
epithelial
cells. Oncogene,
9: 3233-3240,
1994.
5.
Tortora,
1992.
G.,
cDNA
Proc.
of cAMP
type I isozyme
Growth
of growth
cAMP
F.,
18-32,
Viglietto,
placenta
suppression
of malignancy:
50: 7093-7100,
1990.
inhibition
site-selective
Ciardiello,
D. W. Molecular
growth
factor
endothelial
J. A., and Yonemoto,
for a diverse
family
971-1005,
1990.
Y. S. Role
and
G. R., Crabtree,
7.
13:
factor.
15. Taylor,
S. S., Buechler,
protein
kinase:
framework
Annu.
Rev. Biochem.,
59:
Normanno,
N., Ciardiello,
F., Kenney, N., Johnson, G. R., and Salomon,
D. S. Differential
immunohistochemical
detection
of amphiregulin
and
cripto in normal colon and human colorectal
tumors. Cancer Res., 52:
Fontanini,
Salomon,
Rev.,
vascular
cells involve
1995.
6. Ciardiello,
F., Bianco, C., Normanno,
N., Baldassarre,
G.,
Tortora, G., Bianco, A. R., and Salomon,
D. S. Infection with
forming growth factor a antisense retroviral
expression
vector
the in vitro growth and transformation
of a human colon cancer
mt. J. Cancer, 54: 952-958,
1993.
L., and Leung,
of the
Cell
in human
J.,
&
kinase.
N. Epi-
receptors
Growth
thyroid
for human
ten years
Cell
Dr. P.
(Washington
secretion:
family.
1991.
19. Rohlff,
Roberts,
A. B. Autocrine
117: 408-414,
1992.
factor-related
Crit.
Saeki,
and cancer.
factor
of angiogenesis.
kindly
1991.
D. S., Brandt,
growth
malignancies.
4.
factors
Regulation
17. Tortora,
G., Pepe, S., Cirafici,
A. M., Ciardiello,
F., Porcellini,
A.,
Clair,
I., Colletta,
G., Cho-Chung,
Y. S., and Bianco,
A. R. Thyroidstimulating
hormone-regulated
growth
and cell cycle distribution
of
REFERENCES
DC),254:
growth
K., Jakeman,
Endocr.
to the vascular
differentiation
ACKNOWLEDGMENTS
We thank
fibroblast
properties
of proteins.
apy. Cancer
Tagliaferri
P. A.
1991.
1990.
Houck,
biological
9267-9271,
with
to conventional
and D’Amore,
53: 217-239.
14. Maglione,
D., Guerriero,
V.,
Persico,
M. G. Isolation
of a human
with an agent that has different
intraceldoes not antagonize
the effect of cytotoxic
drugs.
8-Cl-cAMP
inhibition
M.,
Rev. Physiol.,
Differ., I: 439-445,
with effects
on the
factors
that have
In fact,
Klagsbrun,
Annu.
VIII
12.
results
paracrine
antineoplastic
by factor
447
Research
10. Folkman,
J. Tumor angiogenesis.
In: J. Mendelsohn,
P. Howley,
L. A. Liotta, and M. Israel (eds.), The Molecular
Basis of Cancer, pp.
206-232.
Philadelphia:
W. B. Saunders,
1995.
11.
by a strong
cells.
therapeutic
autocrine
of TGF-a,
and
as measured
that 8-Cl-cAMP
is a novel anticancer
drug
production
of several
growth
and angiogenic
potential
in the
is accom-
Cancer
epithelial
cells transformed
by
Int. J. Cancer, 53.- 438-443,
1993.
25.
Tortora,
G.,
Ciardiello,
C., Guarrasi,
with 8-chloro-cAMP
patients.
Clin. Cancer
Ciardiello,
F., Pepe.
R., Mild,
S., Tagliaferri,
K., Bianco,
and evaluation
of immunologic
Res., I: 377-384,
1995.
F., Damiano,
V., Bianco,
P.,
A. R. Phase
C., di Isernia,
Caraglia, M., Tagliafem,
P., Baselga, J., Mendelsohn,
and Tortora, G. Cooperative
antiproliferative
effects
Ruggiero,
I clinical
effects
A.,
study
in cancer
G., Ruggiero,
A..
J., Bianco, A. R.,
of 8-Cl-cAMP
and
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
448
Inhibition
528
Factor
anti-epidermal
human
27.
of Growth
cancer
growth
cells.
Ciardiello,
Production
F.,
factor
Clin. Cancer
McGeady,
by 8-Cl-cAMP
receptor
monoclonal
1: 161-167,
1995.
antibody
on
Res.,
M.
L.,
Kim,
N.,
Basolo,
F.,
factor a complementary
DNA
Differ., 1: 407-420,
1990.
Hynes,
N.,
protooncogene
but not by the
of the transforming
growth
leads to transformation.
factor
702-709,
Langton,
B. C., Yokozaki,
H., Saeki, I., Elliott, J. W., Masui, H.,
Mendelsohn,
J., Soule, H., Russo, J., and Salomon,
D. S. Transforming
growth factor a expression
is enhanced
in human mammary
epithelial
cells transformed
by an activated
c-Ha-ras
c-neu
protooncogene
and overexpression
growth
Cell Growth
&
37.
receptor.
Biochem.
Biophys.
Res.
Commun.,
190:
1993.
Mustonen,
involved
I.,
and
Alitalo,
in angiogenesis.
K. Endothelial
129:
receptor
J. Cell Biol.,
tyrosine
895-898,
kinase
1995.
38. Park, J. E., Chen, H., Winer, J., Houck, K., and Ferrara, N. Placenta
growth factor: potentiation
of vascular endothelial
growth factor bioactivity, in vitro and in vivo, and high affinity binding to FIt-I but not to
FIkIIKDR.
J. Biol. Chem., 269: 25646-25654,
1994.
28. Houck,
K., Ferrara,
N., Winer,
J., Cachianes,
G., Li, B., and Leung,
D. W. The vascular
endothelial
growth factor family: identification
of a
39. Stetler-Stevenson,
W. G., Aznavoorian,
S., and Liotta, L. Tumor
cell interactions
with extracellular
matrix during invasion and metastasis. Annu.
Rev. Cell Biol.,
9: 541-573,
1993.
fourth
RNA.
40. Mensing,
H., Pontz, B. F., Muller, P. K., and Gauss-Muller,
study on fibroblast
chemotaxis
using fibronectin
and conditioned
molecular
species
Mol. Endocrinol.,
and characterization
5:
of alternative
splicing
of
1991.
1806-1814,
29. Terman,
B. I., Vermazen,
D., Carrion,
M. E., Dimitrov,
D., Armellino, D. C., Gospodarovicz,
D., and Bohelen,
P. Identification
of the
KDR
tyrosine
kinase
as a receptor
for vascular
endothelial
growth
factor. Biochem.
Biophys. Res. Commun.,
187: 1579-1586,
1992.
30. Shibuya, J., Yamaguchi,
S., Yamane, A., Ikeda, I., Tojo, A., and
Sato, M. Nucleotide
sequence
and expression
of a novel human receptor-type tyrosine kinase (fit) closely related to the fms family. Oncogene, 8: 519-524, 1990.
31.
M.
Maglione,
D., Guemero,
G. Translation
of the
affected
region.
by a small
Growth
32. Budilbon,
G., Clair, I.,
V., Rambaldi,
M.,
Russo,
G., and
Persico
placenta
growth factor mRNA is severely
open reading frame localized in the 5’ untranslated
Factors,
8: 141-152,
A., Cereseto,
A.,
1993.
Kondrashin,
A.,
Nesterova,
M.,
Merlo,
and Cho-Chung,
Y. S. Point mutation of the autophosphorylation site or in the nuclear location signal causes protein kinase A
RII3 regulatory
subunit to lose its ability to revert transformed
fibroblasts. Proc. Nail. Acad. Sci. USA, 92: 10634-10638,
1995.
33. Albini,
A., Iwamoto,
Y., Kleinman,
S. A., Kozlowski,
J. M., and McEwan,
quantitating
the invasive
3239-3245,
1987.
34.
Giunciuglio,
potential
H. K., Martin,
G. R., Aaronson,
R. N. A rapid in vitro assay for
of tumor cells. Cancer Res., 47:
Fassina,
G.,
Masiello,
L.,
Melchiori,
A.,
Baselga,
J., Norton,
L., Masui,
H., Pandiella,
A., Coplan,
W. H., and Mendelsohn,
nation with anti-epidermal
J. Antitumor
effects of doxorubicin
growth factor receptor monoclonal
ies.
85:
J. Natl.
Cancer
Inst.,
1327-1333,
K., Miller,
in combiantibod-
1993.
36. Gitay-Goren,
H., Halobon,
R., and Neufeld,
G. Human
melanoma cells but not normal melanocytes
express vascular
endothelial
as chemoattractants.
41 . Ferrara,
pathological
1995.
Eur.
N. The role
angiogenesis.
J. Cell
I.,
vascular
does
endothelial
confers
a growth
J. Clin.
Invest.,
43.
growth
91:
Biol.,
29:
268-273,
1983.
of vascular
endothelial
growth
Breast Cancer Res. Treat., 36:
42. Ferrara,
N., Winer,
J., Burton,
H. S., Terrel,
I., Keller, G. A.,
advantage
160-170,
Rowland,
A., Siegel,
and Levinson,
factor
factor in
127-137,
M., Phillips,
A. D. Expression
not promote
to Chinese
in vivo
of
transformation
hamster
but
ovary
cells.
1993.
Kim,
K. J., Li, B., Winer,
N. Inhibition
induced angiogenesis
suppresses
362: 244-250,
1993.
H. S., and Ferrara,
J., Armanini,
M., Gillet, N., Phillips,
of vascular endothelial
growth factortumor growth in vivo. Nature (Lond.),
44. Warren, R. S., Yuan, H., Math, M. R., Gillett, N., and Ferrara, N.
Regulation
by vascular endothelial
growth factor of human colon cancer
tumorigenesis
J. Clin.
45.
in
Invest.,
Hori,
growth
basic
D.,
Paglialunga,
G., Culty, M., Arand, G., Ciardielbo, F., Basolo, F., Thompson, E. W., and Albini, A. Invasive
phenotype
of MCF-lOA
cells
overexpressing
c-Ha-ras
and c-erbB-2
oncogenes.
mt. J. Cancer, 63:
815-822,
1995.
35.
dium
V. A
me-
model
1789-1797,
A., Sasada,
R., and
fibroblast
growth
N. Engl. J. Med.,
Gasparini,
333:
G., and
experimental
Matsutani,
Cancer
applications
1757-1763,
Harris,
Mendelsohn,
monocbonal
J. Potential
antibodies.
Cancer
metastasis.
Res.,
Cells,
tumor
of research
on
angiogenesis.
1995.
A. L. Clinical
clinical
of solid
antibody
against human
51: 6180-6184,
1991.
implications
mination of tumor angiogenesis
in breast carcinoma:
new prognostic
tool. J. Clin. Oncol., 13: 765-782,
48.
liver
E. Suppression
monoclonal
factor.
J. Clinical
of
1995.
by immunoneutralizing
46. Folkman,
47.
a mouse
95:
applications
7: 359-362,
of the
much
deter-
more than a
1995.
of anti-EGF
receptor
1989.
49. Baselga, J., and Mendelsohn,
J. The epidermal
growth factor receptor as a target for therapy in breast carcinoma.
Breast Cancer Res. Treat.,
29: 127-138,
1994.
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.
8-Chloro-cyclic AMP inhibits autocrine and angiogenic growth
factor production in human colorectal and breast cancer.
C Bianco, G Tortora, G Baldassarre, et al.
Clin Cancer Res 1997;3:439-448.
Updated version
E-mail alerts
Reprints and
Subscriptions
Permissions
Access the most recent version of this article at:
http://clincancerres.aacrjournals.org/content/3/3/439
Sign up to receive free email-alerts related to this article or journal.
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Department at [email protected].
To request permission to re-use all or part of this article, contact the AACR Publications
Department at [email protected].
Downloaded from clincancerres.aacrjournals.org on June 16, 2017. © 1997 American Association for Cancer
Research.