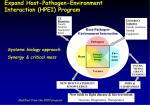
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
REVIEW ARTICLE Internal affairs: investigating the Brucella intracellular lifestyle Kristine von Bargen1,2,3, Jean-Pierre Gorvel1,2,3 & Suzana P. Salcedo1,2,3 1 Faculté de Sciences de Luminy, Centre d’Immunologie de Marseille-Luminy, Aix Marseille Université, UM 2, Marseille Cedex, France; 2INSERM, U 1104, Marseille, France; and 3CNRS, UMR 7280, Marseille, France Correspondence: Suzana P. Salcedo, Centre d’Immunologie de Marseille-Luminy, Aix Marseille Université, Faculté de Sciences de Luminy, Case 906, Marseille, 13288 Cedex 9, France. Tel.: +33 491 269 116 and +33 491 269 115 ; fax: +33 491 269 430; e-mail: [email protected] Received 4 July 2011; revised 10 January 2012; accepted 16 February 2012. Final version published online 22 March 2012. DOI: 10.1111/j.1574-6976.2012.00334.x MICROBIOLOGY REVIEWS Editor: Christoph Dehio Keywords Brucella-containing vacuole; trafficking; type IV secretion system; pathogenicity; bacteria; brucellosis. Abstract Bacteria of the genus Brucella are Gram-negative pathogens of several animal species that cause a zoonotic disease in humans known as brucellosis or Malta fever. Within their hosts, brucellae reside within different cell types where they establish a replicative niche and remain protected from the immune response. The aim of this article is to discuss recent advances in the field in the specific context of the Brucella intracellular ‘lifestyle’. We initially discuss the different host cell targets and their relevance during infection. As it represents the key to intracellular replication, the focus is then set on the maturation of the Brucella phagosome, with particular emphasis on the Brucella factors that are directly implicated in intracellular trafficking and modulation of host cell signalling pathways. Recent data on the role of the type IV secretion system are discussed, novel effector molecules identified and how some of them impact on trafficking events. Current knowledge on Brucella gene regulation and control of host cell death are summarized, as they directly affect intracellular persistence. Understanding how Brucella molecules interplay with their host cell targets to modulate cellular functions and establish the intracellular niche will help unravel how this pathogen causes disease. Introduction Brucellosis is the most common bacterial zoonotic disease worldwide, with over half a million infected people annually (Pappas, 2010). It remains endemic in many parts of the world, including the Middle East, Africa, Latin America, central Asia and several regions of the Mediterranean basin. Human brucellosis is often misdiagnosed and underreported. It is an important travel-associated disease (Memish & Balkhy, 2004), for which no vaccine is available. The treatment requires a combination of different antibiotics for a prolonged period of time (Ariza et al., 2007). Many countries have succeeded in control of the disease in the recent years by nearly eradicating it from livestock, the predominant reservoir for human infections. However, there is growing concern for re-emerging foci of brucellosis in numerous countries, particularly in central Asia, where it is causing significant morbidity (World Health Organization, 2005). Brucella spp. are Gram-negative bacteria that were first isolated in 1887 in Malta by Sir David Bruce from the FEMS Microbiol Rev 36 (2012) 533–562 spleens of soldiers with fatal cases of brucellosis, also known as undulant fever or Malta fever. Brucella spp. belong to the a-2 subdivision of Proteobacteria which includes bacteria that co-evolved with animal or plant hosts, either in a beneficial symbiotic manner such as Sinorhizobium meliloti (a plant symbiont) or as pathogens such as Agrobacterium tumefaciens, Rickettsia spp. and Bartonella spp. At present, ten species of the genus Brucella have been recognized with the nomenclature based on their respective preferential host (Moreno et al., 2002; Audic et al., 2009). The species most pathogenic for humans and that are most relevant for domestic animals are Brucella melitensis that infects goats, sheep and camels; Brucella abortus that causes bovine brucellosis and Brucella suis that is associated with brucellosis in swine. Three other species, Brucella canis (dogs), Brucella ovis (sheep and rams) and Brucella neotomae (desert wood rats) are of low pathogenicity for humans. Since the 1990s, Brucella strains infecting marine mammals have been described in the literature, illustrating the broad mammal host range of ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 534 this bacterial pathogen. Two distinct strains were identified and referred to as Brucella pinnipediae and Brucella cetaceae (Cloeckaert et al., 2001). However, they had not been validly published until 2007 when the species’ names were slightly altered into Brucella ceti and Brucella pinnipedialis (Foster et al., 2007). Brucella ceti is found in dolphins, porpoises and whales, and B. pinnipedialis affects mainly seals. An increasing number of reports associate marine mammal strains with human brucellosis, suggesting that these Brucella species may be pathogenic for humans (Sohn et al., 2003; Whatmore et al., 2008). A more recently described species, Brucella microti, has been isolated from the common vole and from foxes and has been found as a soil contaminant but so far it has not been reported as a cause for human brucellosis (Scholz et al., 2008a, b). Additional strains have been isolated from Australian rodents but have not yet been confirmed as new Brucella species (Tiller et al., 2010a). Finally, so far unknown strains have recently been isolated from brucellosis patients. Brucella inopinata has been isolated from an infected breast implant (Scholz et al., 2010). The second strain is phenotypically and molecularly similar to B. inopinata and was isolated from a patient with chronic lung infection (Tiller et al., 2010b). These newly described strains highlight the complexity of the growing genus Brucella. Humans are not natural hosts for brucellae, and the source of the infection is usually found in domestic or wild animal reservoirs. Occupational exposure and ingestion of contaminated food products are the main routes of infection for humans. However, its high infectivity via aerosols places Brucella on the category B list for agents of biological warfare and accounts for it being the most common laboratory-acquired bacterial infection. Containment of human brucellosis is dependent on successful vaccination of livestock and imposing strict farming hygiene, surveillance and infection control measures. Although Brucella epidemiological control has seen considerable progress in the last years, several recent reports raise concerns. In some European countries, wild boars and hares are widely infected with B. suis biovar 2 (Bergagna et al., 2009; Cvetnic et al., 2009; Galindo et al., 2010). Approximately 50% of the bison population in the Greater Yellowstone Area in the USA have been found infected with B. abortus (Olsen, 2010; Scurlock & Edwards, 2010). In addition, there are concerns of new habitats being colonized by virulent strains, as illustrated by a report of B. melitensis isolated from fish in the Nile river (El-Tras et al., 2010). Marine Brucella species have been isolated from human brucellosis patients (Sohn et al., 2003; McDonald et al., 2006), demonstrating possible transmission from a yet unconsidered reservoir. The growing incidence of brucellosis in wildlife as a potential ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. source for human infection poses a new challenge to eradication of the disease worldwide. The disease in different hosts In animal primary hosts, Brucella have a particular tropism for the reproductive system, often leading to abortion in pregnant female animals following extensive bacterial colonization of the placenta and sterility in male animals. Transmission between animals is facilitated by the presence of high numbers of bacteria in the aborted foetus, reproductive tract discharges and milk. Animals can present severe complications but may also become carriers and continuously shed bacteria for many years (Moreno & Moriyon, 2006). The chronic nature of brucellosis is a key aspect of the disease that is observed in both the natural host, experimental animal models and during infection in humans. In marine mammals, Brucella can affect the reproductive tract and cause abortion but the disease may also present life-threatening complications such as meningoencephalitis, splenic and liver necrosis (Gonzalez et al., 2002; Nymo et al., 2011). In humans, the onset of brucellosis is most commonly manifested as a flu-like illness, with recurrent fevers associated with muscle and joint pain. As bacteria disseminate systemically, patients present enlarged liver and spleen and swollen lymph nodes. If patients are not treated, Brucella have the propensity to form granulomas and can colonize multiple sites in the body. The most common complication is arthritis, which is often seriously debilitating and difficult to treat. More severe complications include liver abscess formation, neurobrucellosis and endocarditis. Antibiotic treatment is normally effective when administered promptly and mortality is rare. Nonetheless, because of the chronic nature of brucellosis, its tendency to relapse and its propensity to affect joints, patients can suffer long-term disability (Franco et al., 2007). In addition, allergic hypersensitivity to Brucella antigens is a frequent consequence of brucellosis following re-infection or repeated contact with Brucella antigens (Moreno & Moriyon, 2006). The mouse model of brucellosis is the most widely used laboratory experimental model to study Brucella virulence in vivo (extensively reviewed by Silva et al., 2011). In this model, Brucella can be found in multiple tissues including the spleen and liver, where microgranulomas are formed during the chronic stages of the infection. Interestingly, a recent study using bioluminescent Brucella strains (Rajashekara et al., 2005) also found them to target the salivary glands, which could be of significance in relation to human infection, where inoculation occurs through ingestion of contaminated food (Rajashekara et al., 2005). In addition, mice presented chronic infection of tail joints FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle with Brucella resembling osteoarticular brucellosis in humans (Rajashekara et al., 2005). The mouse model has been particularly useful to characterize Brucella factors that mediate intra-host survival and to understand the immune response elicited during brucellosis that ultimately leads to development of chronic disease (reviewed by Martirosyan et al., 2011). An intracellular lifestyle The most common portals of entry for Brucella in animals and humans are mucous membranes of the respiratory and digestive tracts, and in the natural host, also the conjunctiva and membranes covering the sexual organs. The cellular sites of entry remain poorly characterized. Bacteria are eventually taken up by phagocytic cells and reach the regional lymph nodes, leading to subsequent systemic dissemination. Brucella can efficiently colonize cells of the monocyte/macrophage lineage and replicate to high numbers in the liver and spleen. In animals, Brucella also multiply in mammary glands and reproductive organs. In humans, any organ can become infected. Histopathology of the tissues from infected animals and patients clearly shows that Brucella extensively replicate within host cells. Much work remains to fully characterize the cellular targets of Brucella during different stages of the infection but it is clear that intracellular replication is 535 directly linked to Brucella pathogenicity. The progression of brucellosis into a chronic disease in either animals or humans is related to the ability of Brucella to persist for prolonged periods within host cells and to resist the host immune response (Martirosyan et al., 2011). Study of Brucella interaction with host cells has mostly relied on in vitro analysis using cultured murine, bovine or human cells (Fig. 1) and more recently, the mouse experimental model of brucellosis. Macrophages Macrophages have been shown to constitute an important site for Brucella intracellular replication within tissues. Despite the fact that over 90% of Brucella internalized by macrophages are killed soon after phagocytosis, a few bacteria escape and establish an intracellular niche permissive for replication without affecting the survival of these phagocytic cells. Activated macrophages are more efficient at killing intracellular Brucella (Jiang & Baldwin, 1993; Eze et al., 2000; Sathiyaseelan et al., 2000). However, wild-type virulent strains are still able to replicate, albeit at later time points after infection (Baldwin & Goenka, 2006). Comparison of different Brucella species shows efficient survival and replication within human and murine macrophages for B. abortus, B. suis and B. melitensis. In the case of the marine species B. ceti and B. pinnipedialis, some Fig. 1. The various cell types of different hosts where Brucella establishes intracellular niches. Depiction of the main differences between the intracellular replication niches set up by Brucella (Brucella abortus, Brucella melitensis or Brucella suis) within specific cell types originating from different hosts: murine (top), human (middle) and natural hosts, mostly caprine and bovine (bottom). White ellipses containing question marks refer to cells or compartment types that have not yet been described in the context of Brucella infection. ER-derived replicative niches are established within murine macrophages infected with either opsonized or nonopsonized Brucella, murine bone marrow-derived DCs infected with nonopsonized Brucella, trophoblast giant cells from infected pregnant mice, human epithelial cells and trophoblasts from infected goats and cows. Although Brucella has been shown to replicate within bovine macrophage cell lines, human monocyte/macrophage cell lines and human DCs, the respective intracellular niches remain uncharacterized in the case of infection with nonopsonized bacteria. In the human monocyte/ macrophage cell line THP1, opsonized Brucella have been shown to replicate in a non-ER vacuole that retains LAMP1. The VirB T4SS has been shown to be required for intracellular replication within murine macrophages, murine DCs, human monocyte/macrophage cell lines, human DCs, human epithelial cells and bovine macrophages. Lys, lysosomes; AP, autophagosome. FEMS Microbiol Rev 36 (2012) 533–562 ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. 536 strains were also able to replicate within human macrophages to the same level as B. melitensis, including a clinical isolate obtained from a patient with osteomyelitis, whereas other strains were cleared within 48 h or were not phagocytosed at all (Maquart et al., 2009). Interestingly, B. microti replicate more efficiently than the virulent B. suis in human and murine macrophages and showed enhanced lethality in mice (Jimenez de Bagues et al., 2010). This emphasizes the high pathogenic potential of these newly isolated Brucella species for humans. Infection of macrophages is not only associated with intracellular bacterial multiplication but also results in interference with macrophage functions. In human monocytes/macrophages, B. abortus inhibits the IFN-cinduced expression of the FccRI receptor and FccRI-mediated phagocytosis (Barrionuevo et al., 2011); in murine macrophages, B. abortus lipopolysaccharide (LPS) reduces their capacity for antigen presentation and subsequent T cell activation (Forestier et al., 2000). Moreover, yet unidentified Brucella factor(s) specifically act on human, but not on murine, macrophage-like cells to inhibit TNFa expression (Caron et al., 1996). Dendritic cells Another important cellular target recently identified is the dendritic cell (DC). Brucella was found to efficiently proliferate within human and murine DCs (Billard et al., 2005; Salcedo et al., 2008). In vitro, Brucella intracellular replication within DCs results in reduction of DC susceptibility to activating signals which may compromise their ability to induce an appropriate immune response (Billard et al., 2007; Salcedo et al., 2008) and might contribute to the development of a chronic infection. However, Brucella infection in these cells is not silent as there is induction of low levels of pro-inflammatory cytokines and increased expression of co-stimulatory molecules as well as MHC class II on the surface of murine DCs (Salcedo et al., 2008). This was also observed in human DCs (Billard et al., 2007; Zwerdling et al., 2008). However, Zwerdling et al. (2008) noted high expression of surface co-stimulatory molecules following Brucella infection comparable to that of Escherichiacoliinfected DCs (but not pro-inflammatory cytokine secretion) at 24 h postinfection and therefore suggested that Brucella induces significant activation of DCs. It is possible that the induction of MHC class II and co-stimulatory molecules achieves maximum intensity at different time points after infection with Brucella compared with E. coli. Consistent with this hypothesis, Billard et al. (2007) found lower levels of DC activation at 48 h postinfection with Brucella in contrast to E. coli. Alternatively, the differences between these two studies could be due to the method of preparation of human DCs or the different cell densities used. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved Because of the migratory properties of these cells, DCs may contribute to systemic dissemination of Brucella during infection. In mice, infected DCs were observed in the lung-draining mediastinal lymph nodes following intranasal inoculation with B. abortus (Archambaud et al., 2010). When the number of alveolar macrophages was artificially reduced prior to Brucella inoculation, slightly higher numbers of pulmonary DCs were infected, resulting in significant increase in replication within the lungs as well as dissemination to the liver and spleen. These results show that DCs are indeed an attractive niche for intracellular replication and that alveolar macrophages have an important role not only as a replicative niche but also in initial containment of the bacterial load in the lung. In the absence of alveolar macrophages, both CD11b- and CD103-positive lung DCs become activated following infection, and there is significant recruitment of iNOSand TNFa-producing monocyte-derived inflammatory DCs. Alveolar macrophages are then contributing to the containment of the inflammatory response and reduction of tissue damage in the lungs during Brucella pulmonary infection. Further work is necessary to understand how the interplay between Brucella and these two phagocytic cells, macrophages and DCs, determines disease outcome in vivo. Using a murine intestinal loop infection model, DCs in the subepithelial dome of ileal Peyer’s patches were also found to efficiently internalize Brucella and could therefore constitute a portal of entry in the gut. These are likely to be the recently identified LysoDCs, which secrete high levels of lysozyme and have been shown to very efficiently capture Salmonella and dead cells (Lelouard et al., 2010, 2011). Trophoblasts In animals, abortion is associated with a rapid proliferation of Brucella within the placenta. The presence of high bacterial loads within placental trophoblasts ultimately results in disruption of the placenta and infection of the foetus (Anderson & Cheville, 1986; Meador & Deyoe, 1989). Trophoblasts are therefore a primary cellular target for Brucella in the natural host. However, little is known about the infectious process in these cells. In ruminants, placental trophoblasts produce substantial amounts of erythritol in the third trimester, which is a favoured carbon source of Brucella. Erythritol may therefore contribute to exacerbated replication of Brucella and subsequent abortion or stillbirth of the infected foetus, predominantly occurring during the third trimester. Brucellainduced abortion in humans is not a frequent outcome of brucellosis but is of medical concern in endemic regions (Khan et al., 2001), suggesting that if Brucella can reach FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle the placenta in pregnant women, trophoblasts may also constitute an important cellular niche. In vitro studies are lacking, particularly regarding intracellular trafficking. One study has demonstrated significant differences in the ability of B. abortus to replicate within bovine trophoblastic cell lines (Samartino et al., 1994). Replication was observed in cells corresponding to middle and late gestation but not early gestational stages, consistent with abortion being predominant in the third trimester. It would be interesting to perform a comparative study with human trophoblasts as abortion in humans, in contrast to the situation in cattle, is mainly reported in the first trimester of pregnancy. Only a few studies have been carried out in the mouse model using murine placenta or placental-derived cells. In mice, transmission to foetus or high colonization of the placenta is mainly observed following high inocula doses. Brucella induces placental damage (Tobias et al., 1993) and targets mouse giant trophoblasts (Kim et al., 2005b). However, as there are important anatomical differences in placentation between ruminants and mice, caution should be taken in comparing to abortion in the natural host or humans. Other cellular niches Although poorly investigated to date, additional cell types may be relevant in brucellosis. A rich amount of literature describes the ability of Brucella to survive and replicate in nonphagocytic cells such as fibroblasts and epithelial cells. Human epithelial cell lines are often used for in vitro studies of Brucella interaction with host cells. For example, human alveolar and bronchial epithelial cell lines have been used in the context of understanding pulmonary brucellosis (Ferrero et al., 2009). In vivo, brucellae can be found in uterine epithelial cells of experimentally infected pregnant goats (Meador et al., 1988). Invasion of epithelial cells is likely to be a relevant step in crossing the mucosal barrier during initial infection but this remains to be demonstrated. Recent studies have looked at Brucella infection of more peculiar cells such as human osteoblastic cell lines (Delpino et al., 2009; Scian et al., 2011), which may be relevant cellular targets during joint and bone complications. Infection of primary cell cultures of mouse astrocytes and microglia has also been carried out (Garcia Samartino et al., 2010). These cells of the central nervous system may be relevant in the context of neurobrucellosis (McLean et al., 1992; Sohn et al., 2003). Moreover, Brucella has been shown to survive within bovine and human polymorphonuclear cells (PMNs) but data are still limited. Brucella inhibits PMN degranulation preventing release of antimicrobial hydrogen peroxide derivatives FEMS Microbiol Rev 36 (2012) 533–562 537 (Riley & Robertson, 1984; Orduna et al., 1991; Iyankan & Singh, 2002). Consistently, Brucella was shown to efficiently resist killing by rat and human PMNs as it fails to induce a strong respiratory burst and degranulation in these cells (Barquero-Calvo et al., 2007). Depletion of PMNs has no significant effect on Brucella proliferation in the mouse model of brucellosis (Barquero-Calvo et al., 2007). Brucella are generally regarded as intracellular pathogens. However, extracellular growth may also be of relevance during disease. Brucellae are well equipped to withstand attacks of the mammalian arsenal of humoral immunity including complement (Barquero-Calvo et al., 2007). At 21 days postinfection of C57BL/6 mice, approximately one third of B. abortus is present not inside but outside of spleen cells. In mice lacking functional B cells (Igh6 / ), the ratio is inverse and just one-third of brucellae are intracellular, yet the bacterial loads in the spleen in both mice strains are very similar (Rolan et al., 2009). These high percentages of extracellular bacteria suggest that bacterial growth outside of their cellular niches may be important during certain stages of infection, at least in the mouse model of brucellosis. However, the fact that Brucella mutants that fail to replicate inside mammalian cells or whose vacuoles become phagolysosomal are generally attenuated in animal models and the fact that extensive analysis of infected animal tissues finds them to be inside cells indicate the intracellular lifestyle to be a key to successful establishment and/or maintenance of infection. Brucella intracellular trafficking The early stages The mechanism for Brucella entry into host cells remains poorly characterized and some data are contentious. However, the early interactions between Brucella and host cells are decisive for intracellular survival. Both antibodyand complement-opsonized Brucella as well as nonopsonized Brucella survive and replicate within macrophages (Baldwin & Goenka, 2006). In contrast, opsonized Brucella fail to replicate in murine bone marrow-derived DCs (S.P. Salcedo and J.P. Gorvel, unpublished results) suggesting that in these cells, different uptake mechanisms result in a different intracellular compartmentalization and survival. Nonetheless, it is clear that Brucella can enter host cells in the absence of opsonin receptors as they can infect nonprofessional phagocytic cells such as trophoblasts, fibroblasts and epithelial cells. In the case of nonopsonized Brucella, entry into both murine macrophages and human monocytes is mediated by lipid rafts (Naroeni & Porte, 2002; Watarai et al., ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 538 2002; Fig. 2). This process is dependent on PI3-kinase and TLR4 (Guzman-Verri et al., 2001; Pei et al., 2008a) and does not result in a significant activation of these cells. Uptake of Brucella by human DCs was also shown to be partially dependent on lipid rafts (Billard et al., 2005). This mode of entry seems to contribute to intracellular survival within phagocytic cells. Engineered rough mutants (lacking the O-polysaccharide of the LPS, see below) that enter independently of lipid rafts fail to reduce macrophage activation and are subsequently killed (Porte et al., 2003; Rittig et al., 2003; Pei & Ficht, 2004). These results suggest that the Brucella O-polysaccharide (also known as O-antigen) mediates interaction with specific receptors at the cell surface and thereby dictates early events in the maturation of Brucella-containing vacuoles (BCVs). Alternatively, the O-polysaccharide could be K. von Bargen et al. modifying the fusogenic properties of the early BCV. However, mutations in LPS and outer membrane proteins are highly pleiotropic and affect multiple surface components making it difficult to interpret the specificity of the phenotypes observed with these mutants. For example, loss of the O-antigen may unmask other surface molecules that would act as PAMPs, which would trigger cell signalling and induce cell death irrespective of the LPS structure. Furthermore, uptake of the naturally rough mutants B. ovis and B. canis is dependent on lipid rafts (Martin-Martin et al., 2010) arguing against a role for the O-polysaccharide in mediating entry in macrophages. Brucella entry into host cells is also dependent on the two-component regulator BvrR/BvrS, which controls the expression of numerous genes including those affecting the acylation status of the LPS lipid A as well as several Fig. 2. Brucellae entry and intracellular trafficking in host cells. Eucaryotic molecules recruited to the BCV or necessary for its maturation are summarized. Green zooms indicate bacterial factors implicated at each step. Brucellae transiently interact with different compartments of the endocytic pathway. Avirulent mutants (e.g. lacking a functional VirB T4SS) are degraded in phagolysosomes, whereas wild-type Brucella strains are capable of controlling fusion with late endosomes and lysosomes and instead interact with the ER exit sites and efficiently replicate in an ERderived phagosome. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle outer membrane proteins, such as Omp3a and Omp3b (Guzman-Verri et al., 2002; Manterola et al., 2005; Lamontagne et al., 2007). Two receptors have been proposed to mediate lipid raft-dependent internalization of Brucella by macrophages: (1) the class A scavenger receptor (Kim et al., 2004b) and (2) the cellular prion protein PrPc (Watarai et al., 2003). The putative receptor PrPc was shown to bind the Brucella heat-shock protein Hsp60. However, the specific role of this receptor in Brucella infection remains controversial as a separate study failed to demonstrate its involvement in entry of B. suis and B. abortus in human monocytes (THP1), murine macrophages (J774 and BMDM) or in vivo (Fontes et al., 2005). In both the case of macrophages and epithelial cells, Brucella adhesion seems to be mediated by eucaryotic receptors containing sialic acid residues that interact with the Brucella surface protein 41 (SP41) encoded by the ugpB locus (CastanedaRoldan et al., 2006). This locus encodes for a glycerol-3phosphate binding ABC transporter present in the outer membrane of pathogenic and nonpathogenic bacteria. Interestingly, Brucella was shown to bind the extracellular matrix proteins fibronectin and vitronectin, which may contribute to tissue colonization (Castaneda-Roldan et al., 2004). Additional recent studies have implicated other Brucella proteins in adhesion/internalization into nonprofessional phagocytes. A null mutant of the BMEI0216 gene (annotation in B. melitensis) has a severe entry deficiency in HeLa cells (Hernandez-Castro et al., 2008). The B. abortus efp gene was also shown to have a defect in internalization but no attenuation in macrophages or in vivo (Iannino et al., 2012). In addition, the pathogenicity island Bab1_2009–2012 encoding a Brucella adhesin has been identified. Deletion of the entire island leads to a decrease in internalization in both HeLa cells and J774 macrophages and reduced adhesion to HeLa cells. More interestingly, the mutant was not attenuated in mice inoculated via the intraperitoneal route but displayed reduced spleen colonization in mice inoculated by oral gavage at 7 and 21 days postinfection (Czibener & Ugalde, 2012). This is the first adhesin identified for Brucella that also displays a role in virulence in vivo. In both phagocytic and nonphagocytic cells, Brucella entry relies on the actin cytoskeleton and partially on the microtubule network. The small GTPases Cdc42, Rac and Rho are necessary for invasion and Cdc42 is directly recruited and activated at the site of entry (Guzman-Verri et al., 2001). In murine trophoblast giant cells, bacterial entry was also dependent on ezrin (Watanabe et al., 2009), a member of the ezrin-radixin-moesin family of proteins that tether actin filaments to the plasma membrane. Once inside cells, Brucella resides in a vacuole, designated the BCV, which interacts with components of the FEMS Microbiol Rev 36 (2012) 533–562 539 endocytic and secretory pathways to ensure bacterial survival. The nascent BCVs undergo interactions with early endosomes (Fig. 2), acquiring markers such as the tethering protein early endosomal antigen EEA1 and the GTPbinding protein Rab5 (Pizarro-Cerda et al., 1998b; Chaves-Olarte et al., 2002; Celli et al., 2003). These interactions occur immediately after internalization and are very transient. Early BCVs are enriched in cholesterol and flotilin-1, a protein involved in lipid raft signalling associated with phagosome maturation and interaction with the endocytic pathway (Arellano-Reynoso et al., 2005). The Brucella cyclic ß-1,2-glucan has been proposed to modify cholesterol-rich lipid rafts present on the BCV membrane (Fig. 2) and control BCV maturation in both epithelial cells and macrophages (Arellano-Reynoso et al., 2005). Interestingly, it does not seem to play a role in virulence in DCs suggesting the composition of the vacuolar membrane in these cells may be different (Salcedo et al., 2008). Analysis of early stage trafficking, particularly in macrophages, is hampered by the fact that more than 90% of bacteria are killed within the first hours of infection. Therefore, the majority of BCVs will become phagolysosomal and bacteria will be degraded. In consequence, analysis of the remaining 10% that will actually successfully establish a replicative niche will be difficult and will likely require live cell imaging. An intermediate stage As BCVs lose early endosomal markers, they acquire the late endosomal/lysosomal membrane protein LAMP1 (Fig. 2). Although multiple studies have shown that BCVs (unlike latex-beads and heat-killed bacteria) do not acquire most of the markers of late endosomes and lysosomes, live imaging in infected epithelial cells has recently challenged this hypothesis (Starr et al., 2008). According to this study, a significant proportion of BCVs are accessible to fluid phase markers preloaded to lysosomes, although at a much reduced rate compared with heatkilled BCVs (Starr et al., 2008). Furthermore, BCVs acquire late endocytic markers such as Rab7 and its effector the Rab-interacting lysosomal protein (RILP) and this is necessary for further BCV trafficking. These results demonstrate that maturation of BCVs does involve controlled and limited fusion events with late endosomes and lysosomes, a step necessary for reaching the replicative niche. Consistently, numerous studies using fixed samples had previously shown that luminal lysosomal enzymes such as cathepsin D cannot be detected on wild-type BCVs, in contrast to heat-killed BCVs (Pizarro-Cerda et al., 1998a; Comerci et al., 2001; Celli et al., 2003; Arellano-Reynoso et al., 2005), suggesting they are not extensively fusing with lysosomes. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 540 An important phase in phagosome maturation is the acidification of BCVs, an essential step for Brucella survival within host cells (Porte et al., 1999; Boschiroli et al., 2002) and essential for diversion of intracellular trafficking (Starr et al., 2008). In fact, inhibition of BCV acidification at early stages of the phagosome maturation (within 1 h) completely abolishes intracellular replication. This is likely due to the fact that an acidic vacuolar pH induces the expression of Brucella genes that are required for virulence. For example, BCV acidification induces the expression of the virB operon that encodes a type IV secretion system (T4SS) crucial for the establishment of the Brucella replication niche (Boschiroli et al., 2002). The endoplasmic reticulum-derived vacuole: the Brucella safe haven Following transient fusion events with the endocytic pathway, BCVs undergo extensive interactions with the secretory pathway before finally fusing with the endoplasmic reticulum (ER) (Fig. 2). BCV interaction with endocytic compartments is followed by the acquisition of features of autophagosomes in epithelial cells but not in macrophages: they become multimembranous and positive for monodansylcadaverine (Pizarro-Cerda et al., 1998a, b), although this compound is likely rather a general marker for late endocytic/lysosomal and acidic compartments (Mizushima, 2004). This transient passage through an autophagosome-like compartment is probably a consequence of fusion events with autophagic vacuoles already present in the cell, perhaps induced upon infection as an innate immune response to clear the invading bacterial pathogen (Gorvel & de Chastellier, 2005). Virulent Brucella strains, however, efficiently escape these autophagosomal compartments to reach the ER. Interestingly, an RNAi screen carried out in Drosophila S2 cells identified 52 host factors that when inhibited either increased or reduced intracellular replication of Brucella, one of which has been related to autophagy (Qin et al., 2008). Drosophila S2 cells cannot be incubated above 30 °C and may therefore be problematic to study pathogens requiring 37 °C for maximal expression of genes involved in virulence. However, they have been shown to be a powerful system to study Brucella entry and replication, because the pathways involved are very similar to those described for mammalian cells, including dependency on Rho subfamily GTPases and PI3 kinase for entry and replication occurring within compartments positive for ER markers. Twenty-nine genes identified in this ER-directed RNAi screen had never been correlated to bacterial virulence. Interesting factors include specific SNAREs, kinases, proteins related to cytoskeleton, chaperª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. ones and biosynthetic or metabolic enzymes. Inhibition of the inositol-requiring enzyme 1 (IRE1a), an important kinase that regulates the host cell unfolded protein response (UPR) and also controls autophagosome biogenesis, resulted in a strong reduction in Brucella survival in both insect and mammalian cells. Because other kinases of the UPR were not involved in control of Brucella replication, it has been proposed that IRE1a activation by Brucella may induce the biogenesis of autophagosomes from ribosomal-free regions of the ER, which would interact with BCVs, modulate fusion events with lysosomes, and contribute to complete association with the ER (Qin et al., 2008). Further work is now required to test this hypothesis and determine the role of IRE1a during Brucella survival within phagocytic cells, for which no autophagosome-derived BCV has yet been described. Brucella replication in ER-derived vacuoles has been described for all cell types analysed with the exception of human monocytes, in which opsonized Brucella was shown to replicate in ER-negative LAMP1-positive large vacuoles containing multiple bacteria (Bellaire et al., 2005). In murine immortalized macrophages, murine bone marrow-derived macrophages and DCs as well as human epithelial cell lines, BCVs acquire numerous proteins of the ER such as the lectin chaperones calnexin and calreticulin as well as the ER protein translocator Sec61 (Pizarro-Cerda et al., 1998b; Celli et al., 2003). The vacuolar membrane presents numerous ribosomes visible by electron microscopy, and the ER lumen enzyme, glucose 6-phosphatase can be found within BCVs. At this stage, Brucella has reached a safe niche for intracellular persistence. Therefore, the ER is not only an important source of membrane for replicating bacteria but also creates an environment suited for Brucella replication. These late BCVs behave as ‘extensions’ of the ER illustrated by the fact that induction of ER vacuolation by treating cells with the toxin aerolysin also results in vacuolation of BCVs (Pizarro-Cerda et al., 1998b; Celli et al., 2003). Importantly, Brucella can be found in ER-associated compartments in placental trophoblasts from tissue obtained from B. abortus-infected cattle and goats (Anderson & Cheville, 1986), highlighting the importance of the ER-derived BCVs as an intracellular niche for infection in vivo. The molecular mechanisms that enable Brucella to initiate fusion with the ER still remain to be deciphered. The initial contact with the ER is established at ER exit sites, where BCVs interact with the Sar1/COPII complex. Inhibition of the Sar1 activity, which results in disruption of ER exit sites, blocks intracellular replication by preventing BCVs from fusing with the ER (Celli et al., 2005). The Brucella proteins that mediate these interactions are still unknown but there is clear evidence that the T4SS FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle encoded by the virB operon is necessary for sustained interactions with the ER (Comerci et al., 2001; Celli et al., 2003) (see below). A recent proteomic approach to characterize the composition of the BCV membrane has revealed the role of the small GTPase Rab2 in Brucella intracellular replication (Fugier et al., 2009). Rab2 was recruited to BCVs, and inhibition of Rab2 prevented the fusion of BCVs with ER-derived vesicles and instead BCVs retained LAMP1. Rab2 is known to interact with the glyceraldehyde-3phosphate dehydrogenase (GAPDH), the coat COPI complex and the protein kinase C (PKC ι/k) to control vesicular trafficking from the Golgi to the ER, via vesicular tubular clusters (Tisdale et al., 2004). All the members of the GAPDH/COPI/Rab2/PKC ι/k complex are required for Brucella intracellular replication, suggesting that maturing BCVs are interacting with vesicular tubular clusters and may intercept retrograde vesicular trafficking (Fig. 2). It is therefore likely that the final maturation of BCVs as well as their interaction with the ER is mediated by specific Brucella effectors translocated across the BCV membrane into host cells. Consistent with this hypothesis, the Brucella-translocated RicA was recently identified in a high-throughput yeast two-hybrid screen and shown to interact with Rab2 (de Barsy et al., 2011). Further work is now crucial to understand at the molecular level how this Brucella protein in conjunction with other uncharacterized effectors directs BCV interactions with the secretory pathway to sustain successful interactions with the ER and ensure intracellular replication. The knowledge of the trafficking events described above represents a summary of numerous studies using multiple cell lines infected with either B. abortus, B. melitensis or B. suis strains. No analysis of the intracellular compartment of any of the newly described Brucella strains has yet been carried out. Brucella microti, which is highly virulent in mice, has been shown to efficiently replicate within human and murine macrophages (Jimenez de Bagues et al., 2010). Three lines of evidence are indicative that the intracellular trafficking of B. microti phagosomes is equivalent to that of other described Brucella strains (Hanna et al., 2011): (1) intracellular replication is dependent on the VirB T4SS, (2) the VirB operon is induced in acidic media and (3) neutralization of acidification during early steps of infection blocks intracellular replication. However, the nature of B. microti BCVs remains uncharacterized. It would be interesting to investigate further the intracellular trafficking of these strains and other more clinically relevant Brucella strains such as B. ceti, B. pinnipedialis and B. inopinata. Once Brucella reach their replicative niche within host cells, extensive replication occurs without much disruption of host cell integrity. This is seen both in cell culture FEMS Microbiol Rev 36 (2012) 533–562 541 and in infected animals. It remains completely unknown what happens after reaching high bacterial numbers within host cells; how bacteria exit host cells and how cell-to-cell spread may occur. One possible hypothesis for cell-to-cell spread may be the induction of cell death by spontaneously occurring rough mutants (see below) but this remains to be verified. It is also not clear whether low replication observed in some cells may actually constitute an important reservoir for Brucella relevant during chronic stages of infection. These are all important new avenues of research that must be undertaken. The problem of defining virulence in Brucella and identifying factors required for intracellular trafficking and disease The mechanistic basis for Brucella trafficking diversion is so far poorly understood. Most studies screen for attenuated mutants based on intra-host replication/survival and few control for actual defects in intracellular trafficking. A major drawback in the search for factors involved in the establishment of the intracellular niche is the fact that killed brucellae are delivered to phagolysosomes. This renders the distinction between the actual effects of mutations rather difficult. The reason that the vacuole of a Brucella mutant is retained in lysosomes may be its deficiency in reaching the ER-derived intracellular niche. However, particularly because the conditions in the early BCV are acidic and proteolytic (Starr et al., 2008), it might just be more sensitive to environmental stresses such as reactive metabolites or, at later stages, auxotroph for an essential nutrient lacking in the bacterial vacuole. In such a scenario, the bacteria would be perfectly equipped to establish a replicative niche in their host cells, and the difference in trafficking to wild-type Brucella would reflect merely bacterial death because of nontrafficking-related events. For instance, the Brucella membrane components phosphatidylcholine and phosphatidylethanolamine have been described to be involved in intracellular survival (Conde-Alvarez et al., 2006; Bukata et al., 2008). BCVs of mutants of the phosphatidylcholine pathway show a higher acquisition of LAMP at early time points in bone marrow-derived macrophages which then progressively moves to wild-type levels (Conde-Alvarez et al., 2006). Similarly, vacuoles of a phosphatidylethanolamine-deficient mutant early acquire and maintain LAMP-1 (Bukata et al., 2008). In both cases, LAMP acquisition coincides with a more pronounced initial bacterial killing, which is followed by late stage replication. However, some mutants of the phosphatidylcholine pathway are more sensitive to killing by nonimmune serum and poly-L-ornithine (Conde-Alvarez ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 542 et al., 2006), and the B. abortus mutant of phosphatidylethanolamine is more sensitive to anionic detergents and polycationic peptides (Bukata et al., 2008), both of which seems to be related to general changes in membrane properties. This increased sensitivity to macrophage antimicrobial actions complicates a differentiation. The mutation could result in an actual trafficking defect leading to killing of the bacteria. Alternatively, their increased sensitivity to macrophage bactericidal mechanisms may lead to enhanced phagolysosome formation of vacuoles containing dead Brucella. Likewise, although Brucella strains found in phagolysosomes usually do not replicate, a lack of replication is not indicative for their incapability to avoid lysosomal degradation. For instance, a mutant for nicotinamidase/pyrazinamidase of B. abortus does not replicate in HeLa cells and macrophages. However, the mutant’s vacuole does not fuse with lysosomes and bacterial replication can be restored upon external addition of nicotinic acid to the medium (Kim et al., 2004a). A spoT mutant of B. abortus that is more sensitive to various stress conditions as compared with wild-type bacteria has a lower intracellular replication within macrophages but does not co-localize with either late endosomes or lysosomes (Kim et al., 2005a). Similar questions remain to be solved with respect to the correlation between intracellular trafficking and disease. Brucella mutants whose vacuoles become phagolysosomal are generally attenuated in a mouse model (see below) and factors involved in trafficking diversion crucial for bacterial pathogenicity therefore usually defined as virulence factors. However, although being widely used, this classical concept of ‘virulence factors’ (Falkow, 1988; Casadevall & Pirofski, 1999; Wassenaar & Gaastra, 2001) is problematic in the analysis of bacterial pathogens such as Brucella whose overall morphology and metabolism have seen a long-term evolutionary adaptation to their respective hosts (Seleem et al., 2008). Even though they can be grown in culture medium, brucellae are usually not found in a noninfectious context (Gorvel & Moreno, 2002), rendering it difficult to draw the line between genes required for (intra-host) survival in general and actual virulence genes. Unbiased screens for virulence determinants based on classical (Delrue et al., 2001; Kohler et al., 2002) or signature-tagged transposon mutagenesis (STM) (Lestrate et al., 2003) in the last decade have identified a multitude of genes involved in metabolism, nutrition and cell wall or inner and outer membrane constituents rather than those homologous to known virulence factors (in their various definitions including to cause host damage, not to be essential for bacterial fitness or to be present only in pathogenic species). This has led to the idea of Brucella being a pathogen ‘without classic virulence genes’, such as, for example, haemolytic toxins (Seleem et al., ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. 2008). Although many open reading frames identified to be crucial for intra-host survival have no homology to any known bacterial protein and may code for factors that match the concept of classical virulence factors (Kohler et al., 2002; Lestrate et al., 2003; Wu et al., 2006), adaptation processes might have led to overall optimization of morphology, metabolism and effector proteins for survival in the host. Particularities of the cell envelope or amino acid metabolism – although present in nonpathogenic species and vital for bacterial fitness – may likewise serve to promote host invasion. An evolutionary adaptation to its host ultimately leads to a bacterium whose morphology, physiology and potential to cause disease form an inseparable entity. Accordingly, rather than terming them virulence factors, for clarity, we will refer to bacterial factors in regard to their particular functions and effects such as on trafficking diversion or intracellular multiplication. The mutant as a mirror of intracellular requirements Although they may not serve to understand the mechanisms of how Brucella establish their intracellular niche, the multitude of transport and metabolism-associated genes (reviewed in Roop et al., 2009 and Barbier et al., 2011) that have been found to mediate Brucella intracellular/intra-host survival and replication collectively draft an indirect yet detailed picture of the environment encountered by the bacteria during infection. For instance, the high number of Brucella attenuated mutants that are affected in carbohydrate metabolism or transport suggests that these may serve as energy and/or carbon source for intracellular brucellae (Foulongne et al., 2000; Hong et al., 2000; Lestrate et al., 2000; Kohler et al., 2002). Kinetic studies of regulation of carbon metabolism genes suggest a constant process of adaptation to the environmental changes in the BCV: enzymes of sugar metabolism and uptake systems are downregulated early in infection, whereas an increase in amino acid catabolism might open up an alternative carbon source for anabolic processes; the sugar-based catabolism might then be reactivated once the replicative ER-derived niche is established (McKinney et al., 2000; Kohler et al., 2002; Barbier et al., 2011). The requirement for several genes involved in metabolism of different amino acids and nucleotide synthesis sketches the picture of a nutrient-poor environment allowing for few to no auxotroph defects (Hong et al., 2000; Foulongne et al., 2001; Kohler et al., 2002; Kim et al., 2003; Lestrate et al., 2003). Successful infection additionally requires the presence of several ion transporters such as the manganese transporter MntH (Anderson et al., 2009). FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle The BCV seems to display a low oxygen tension. Brucella mutants that are deficient in cytochromes that are particularly efficient at low O2 concentrations (the cbb3-type cytochrome c oxidase or cytochrome bd ubiquinol oxidase) are attenuated in cellular and animal infection models (Endley et al., 2001; Kohler et al., 2002), just like Brucella lacking components of the denitrification pathway (Kohler et al., 2002; Haine et al., 2006; LoiselMeyer et al., 2006) which may allow for the use of terminal electron acceptors different from O2. Alternatively (or additionally), these proteins could be involved in scavenging and detoxification of host-derived reactive oxygen or nitrogen species, respectively. Several attenuated mutants suggest that Brucella is exposed to these bactericidal compounds in the course of infection such as a strain defective in a Cu–Zn superoxide dismutase (Tatum et al., 1992; Gee et al., 2005) or with a double deletion in catalase (KatE) and peroxiredoxin (AhpC) (Steele et al., 2010). Exposure to reactive intermediates may also account for the requirement of DNA repair systems such as RecA for survival in mice (Tatum et al., 1993). These mutant-based indications of the Brucella intrahost environment have been complemented in recent years by the determination of Brucella intracellular gene expression profiles. The quorum sensor regulator BvrR/ BvrS controls intracellular expression of several genes such as genes involved in carbon metabolism and denitrification and seems to be critically involved in the switch from an extracellular to an intracellular lifestyle (Viadas et al., 2010). Particularly, the analysis of early time points of infection and in vivo has recently been improved by an optimized methodology to obtain pure RNA from limited amounts of intracellular Brucella (Rossetti et al., 2010). Brucella factors involved in establishment of the intracellular niche The type IV secretion system VirB Perhaps one of the most studied factors required for Brucella trafficking diversion is the T4SS encoded by the VirB operon (de Jong & Tsolis, 2012). The VirB genes are homologous to those of type IV DNA transfer systems known from other Gram-negative bacteria such as A. tumefaciens or Legionella pneumophila (Alvarez-Martinez & Christie, 2009). T4SS are classified in two subgroups, IVA and IVB. The latter is closely related to conjugation systems and includes those of pathogens such as L. pneumophila and Coxiella burnetii. The Brucella VirB T4SS apparatus, which belongs to the subgroup IVA, was first identified in B. suis and is encoded by 12 open reading frames (virB1 to virB12) on chromosome II (O’Callaghan et al., 1999). It is essential to the virulence of all FEMS Microbiol Rev 36 (2012) 533–562 543 Brucella strains investigated in their respective models including the more recently described species B. microti (Hanna et al., 2011). As mentioned earlier, the VirB apparatus is crucial for the intracellular trafficking of Brucella within professional phagocytes and nonprofessional phagocytes including macrophages, DCs and epithelial cells (Fig.1, O’Callaghan et al., 1999; Foulongne et al., 2000; Sieira et al., 2000; Comerci et al., 2001; Delrue et al., 2001; Celli et al., 2003; Kim et al., 2003; Billard et al., 2005; Rajashekara et al., 2006; den Hartigh et al., 2008; Salcedo et al., 2008). BCVs of VirB deficient strains are unable to sustain interaction with the ER. The vacuoles of most of these mutants eventually fuse with lysosomes where they are degraded (Sieira et al., 2000; Comerci et al., 2001; Delrue et al., 2001; Celli et al., 2003; Salcedo et al., 2008; Starr et al., 2008). Interestingly, a polar mutant of virB10 that lacks transcription of downstream genes is degraded in phagolysosomes, whereas a nonpolar mutant of virB10 is recycled to the cell surface (Comerci et al., 2001), suggesting that the escape from the degradative pathway and establishment of the ER-derived niche for intracellular replication are two distinct events mediated by different mechanisms (Comerci et al., 2001). The inability of virB mutants to establish an intracellular replicative niche reflects their attenuation in the mouse infection model (Hong et al., 2000; Lestrate et al., 2000; Sieira et al., 2000; Kahl-McDonagh & Ficht, 2006; Rajashekara et al., 2006) and in goats (Kahl-McDonagh et al., 2006; Zygmunt et al., 2006). After infection by gavage, virB-deficient B. melitensis present lower bacterial numbers in liver, spleen and intestinal tissues (Paixao et al., 2009). Bypassing the digestive tract using intraperitoneal injection, virB-deficient strains are able to disseminate to lymph nodes, liver and spleen (Rajashekara et al., 2005; Rolan & Tsolis, 2007; Paixao et al., 2009). However, after the initial phase, Brucella virB mutant strains are cleared much faster than wild-type bacteria, suggesting defects in persistence to maintain chronic infection. The difference in survival of virB mutants within infected cells in vitro (eliminated in 24 h) and in vivo (persistence equivalent to wild type for the first few days) could reflect different bacterial localization during mouse infections (e.g. intra- vs. extracellular) or that cells that are able to kill in vitro cannot do so in vivo because, for example, they are regulated by other cells or a particular cytokine environment. In contrast to wild-type brucellae, virB mutants with a deficient T4SS induce essentially no transcriptional changes in spleen cells. They do not induce inflammation-related genes and the infection remains quiescent, suggesting that T4SS function triggers innate immune responses (Roux et al., 2007). However, the level of ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. 544 response is much lower than that induced by pathogens such as Salmonella (Barquero-Calvo et al., 2007), underlining the immune evasive strategies of Brucella. The regulation of the virB operon expression correlates to its virulence functions. The B. suis virB operon is expressed maximally in minimal medium (rather than rich medium) at early exponential phase, at temperature of 37 °C and inside of the host cell (Boschiroli et al., 2002). In all Brucella species investigated, an induction of VirB protein expression is observed in response to an acidic environment, and this environmental stimulus seems to account for the major part of induction observed intracellularly (Boschiroli et al., 2002; Rouot et al., 2003). The initial acidification of phagosomes which is essential for B. suis intramacrophage replication (Porte et al., 1999) is required to induce VirB expression. Interestingly, there seem to be some differences in the regulation of VirB expression among Brucella strains: whereas B. abortus, B. melitensis and B. ovis express VirB at neutral pH in a rich medium, B. suis, B. canis and the vaccine strains S19, RB51 (both derived from B. abortus) and Rev1 (derived from B. melitensis) show little to no VirB protein expression in these conditions (Rouot et al., 2003). Transcriptional regulators of VirB synthesis are the quorum-sensing regulators VjbR (Delrue et al., 2005) and BlxR (Rambow-Larsen et al., 2008), the histidine utilization regulator HutC (Sieira et al., 2010), the transcription factors DeoR, AraC8, AraC2, GntR4 and NolR (Haine et al., 2005), the two-component system (TCS) BvrR/BvrS (Martinez-Nunez et al., 2010), the stringent response regulator encoded by Rsh (Dozot et al., 2006) (see below) and the integration host factor that specifically interacts with the virB promoter during intracellular and vegetative growth (Sieira et al., 2004). Recent findings suggest that apart from being subject to regulation, mutation of VirB may itself affect expression of other genes. Relative transcription levels of several genes including the quorum-sensing regulator VjbR are downregulated in a B. melitensis VirB mutant during growth in culture and inside macrophages (Wang et al., 2009). Differential expression at transcriptional and posttranscriptional levels can also be observed with several outer membrane proteins, which may explain higher sensitivity of virB mutants to polymyxin B and several environmental stresses (Wang et al., 2010). Substrates of the T4SS Bacterial proteins (‘effectors’) translocated into the host cell are important elements of T4SS in the intracellular survival of pathogens. Although the VirB Brucella T4SS was first described more than 10 years ago, few potential effector proteins have been identified and only one for which a function has been ascribed (Table 1). Two proteins that are translocated into the macrophage cytoplasm in a T4SS-dependent manner are VceA and VceC (de Jong et al., 2008). Translocation of VceA and VceC fused with the TEM1 b-lactamase (N-terminus) into mouse macrophage-like J774 cells can be detected from 7 h onwards and is dependent on the last C-terminal 20 amino acids, which contain a motif reminiscent of the secretion signal identified in Agrobacterium T4SS substrates. The open reading frames of VceA and VceC are conserved in all Brucella species sequenced, including B. suis, canis, ovis, abortus and melitensis (de Jong et al., Table 1. Brucella proteins translocated into host cells during infection Name ORF* VceA BAB1_1652/BMEI0390 VceC BAB1_1058/BMEI0948 RicA BAB1_1279/BMEI0736 BPE123 BAB2_0123/BMEII1111 BPE043 BPE005 BPE275 BPE865 BPE159 BAB1_1043/BMEI0961 BAB1_2005/BMEI0067 BAB1_1275/BMEI0739 BAB1_1865/BMEI0198 BAB2_0159/BMEII1079 Signals required for secretion/translocation T4SS translocation Tag used Function Reference C terminus required for translocation N-terminus Sec signal present C terminus required for translocation nd Yes TEM1 Unknown de Jong et al. (2008) Yes TEM1 Unknown Yes TEM1 Yes CyA and 3FLAG Interacts with Rab2 Unknown Yes Yes Yes No No CyA CyA CyA CyA CyA Unknown Unknown Unknown Unknown Unknown N-terminus required for translocation (Sec signal) nd nd nd nd nd de Barsy et al. (2011) Marchesini et al. (2011) *ORF nomenclature is given for the genomes of Brucella abortus 2308 (BAB) and B. melitensis 16M (BME). ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle 2008). VceA and VceC are co-regulated with the T4SS as they belong to the VjbR regulon (de Jong et al., 2008), which controls expression of several virulence-associated factors including VirB. The cellular targets and the functions of VceA and VceC remain unknown. The third Brucella protein found to be translocated into the cytoplasm of RAW264.7 macrophages is RicA (de Barsy et al., 2011). It has been identified because of its interaction with the guanosine diphosphate (GDP)-bound form of the eucaryotic protein Rab2, a small GTPase that is crucial for Brucella intracellular replication (Fugier et al., 2009). A B. abortus ricA deletion mutant replicates faster in HeLa cells and shows an accelerated loss of LAMP1 from its BCVs than wild-type bacteria. The ricA mutant resides in BCVs, which recruit less GTP-locked Rab2 (de Barsy et al., 2011) suggesting that RicA plays a role in control of BCV maturation. However, the fact that inhibition of Rab2 blocks intracellular replication but the ricA mutant is not attenuated in virulence suggests other effector proteins are involved in the control of Rab2 function. At this stage, it is not clear what is the activity of RicA on Rab2. Results indicate that RicA does not act as a guanine nucleotide exchange factor catalysing the replacement of GDP by GTP on Rab GTPases. Because RicA interacts with the GDP-bound form of Rab2, it is unlikely to be a GTPase-activating protein (GAP) that stimulates GTP hydrolysis by converting the GTPase back to its GDP-bound form. It is possible that RicA is functioning as a guanine nucleotide dissociation inhibitor (GDI) that stabilizes the inactive Rab form by preventing GDP dissociation or GDI displacement factor that removes GDI and allows membrane insertion of Rab2 by its geranylgeranyl anchor. Alternatively, RicA could be interacting with Rab2 without having a direct control on its activity. Translocation of TEM1-RicA is observed in the wildtype strain but not in the virB mutant and only 24 h after infection of RAW264.7 macrophages. At this time point, there is strong attenuation of virB mutants, which are in very different compartments than the virulent strain, as they are degraded in lysosomes. This could account for the lack of translocation of TEM1-RicA in the virB mutant. Interestingly, there is no obvious C-terminal motif like for VceA and VceC, and secretion of RicA into the culture media was independent of the T4SS. It is possible that the in vitro secretion assay used in this study induces an alternative pathway for effector proteins to cross the bacterial membranes. Another explanation could be that the secretion across the bacterial membranes is independent of VirB and uncoupled from the translocation across the vacuolar membrane. This has recently been shown to occur in the case of T3SS (Akopyan et al., 2011). Alternatively, RicA may not be a T4SS effector. Additional work is now necessary to test these hypotheses. FEMS Microbiol Rev 36 (2012) 533–562 545 An in silico screen has recently identified four additional substrates of the VirB T4SS (Marchesini et al., 2011). Furthermore, it identified two Brucella proteins that are translocated into macrophages in a VirB-independent manner suggesting that Brucella has another secretion system yet to be identified. Because of the structural similarities of bacterial export machineries and flagella, it has been speculated that in Brucella (normally considered nonmotile), the flagella genes may serve as a secretion apparatus rather than as an organelle mediating bacterial movement (Lestrate et al., 2003). Moreover, the co-regulation of flagellum components with the VirB secretion system and the attenuation of a flagellum mutant during persistence of B. melitensis in the mouse model of infection suggest a role for this organelle in Brucella pathogenicity (Delrue et al., 2005; Fretin et al., 2005). However, it remains to be demonstrated if these proteins are translocated via the flagella system. Translocation of all proteins identified in the screen was assayed 5 h after infection of murine macrophages, using C-terminal fusions with the adenylate cyclase reporter CyA from Bordetella pertussis. In the case of the T4SS substrate BPE123-CyA, a protein with no predicted function, the concentration of cAMP peaked between 2 and 5 h postinfection consistent with maximal activation of VirB (Sieira et al., 2004). In murine bone marrow-derived macrophages, a 3xFLAG-tagged BPE123 was found in the proximity of BCVs at 4 h after infection. In contrast with VceA and VceC, translocation of BPE123 was dependent on the N-terminal 25 amino acids, which contain a Sec secretion signal. Although no obvious C-terminal motif was identified, BPE123 contains several positively charged amino acids that could constitute a signal for the T4SS. It is curious that for some substrates (e.g. BPE123), translocation by the VirB T4SS would be coupled to Sec-dependent secretion but not for all effectors (e.g. VceA and VceC). T4SS substrates of other bacteria have been shown to depend on a two-step translocation, including the pertussis toxin which contains a sec-dependent signal to cross the inner membrane (Gauthier et al., 2003) and some substrates in Agrobacterium that lack a signal peptide but form a soluble complex with VirJ in the periplasm, which then enables further interaction with T4SS components and translocation across the bacterial outer membrane and host cell membrane (Pantoja et al., 2002). Other potential translocated effectors Although Brucella induces a minimal inflammatory response in the host, it has acquired proteins that help modulate host innate and adaptive immune response mechanisms and which may therefore be important in the development of a chronic infection. One such interesting ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. 546 protein contains a Toll/interleukin-1 receptor domain (TIR), which is essential in TLR signalling. It has been designated Btp1 for B. abortus (Salcedo et al., 2008) and TcpB for B. melitensis (Cirl et al., 2008) but for clarity we will refer to it as Btp1 (Brucella TIR-containing protein 1). This nomenclature is in our opinion more appropriate because of the presence of a second TIR-containing protein in the genome of Brucella (Btp2; Salcedo and Gorvel, unpublished results). Btp1 does not have a role in trafficking but is seems to contribute to infection by manipulating intracellular host pathways. Btp1 has been shown to interfere with TLR4- and TLR2-mediated NF-jB activation as well as cytokine secretion (Cirl et al., 2008; Salcedo et al., 2008; Radhakrishnan et al., 2009). Although one study suggested that Btp1 interacted with Myd88 (Cirl et al., 2008), there is also good evidence that it targets the adaptor protein TIRAP/MAL required for both TLR2 and TLR4 signalling. A more recent study describes a stronger interaction between Btp1 and Myd88 when compared with TIRAP and that Btp1 specifically targets the death domain of Myd88 (Chaudhary et al., 2011). Differences observed by various research groups regarding the eucaryotic target of Btp1 may simply reflect the different methodology used, and it is now essential to determine what is the ‘real’ target of Btp1 during infection. Btp1 binds phosphoinositides similarly to TIRAP, so it may be targeted to the plasma membrane and interfere with the Myd88-TIRAP complex (Radhakrishnan et al., 2009) where it can induce ubiquitination and degradation of TIRAP (Sengupta et al., 2010). More recently, Btp1 was shown to modulate microtubule dynamics. Btp1 stabilizes polymerized microtubules and it enhances their rate of nucleation and polymerization (Radhakrishnan et al., 2011). However, most of the work regarding Btp1 has been performed in vitro, by ectopically expressing the protein in host cells. It is now important to determine the cellular localization of translocated Btp1 and its molecular interactions during infection to confirm these hypotheses. During Brucella infection, Btp1 has been shown to contribute to the modulation of TLR2-dependent activation of murine bone marrow-derived DCs (Salcedo et al., 2008) and to decrease TIRAP degradation in macrophage-like J774 cells (Sengupta et al., 2010). Although the btp1 mutant is attenuated neither in any of the cell systems tested so far (cultured macrophages, epithelial cells and DCs) nor in immune competent intraperitoneally inoculated mice, it plays a role in control of the immune response during infection. In intranasally infected mice, specific subsets of lung DCs showed higher maturation levels when infected with the btp1 mutant compared with the wild-type strain (C. Archambaud, S.P. Salcedo and J.P. Gorvel, unpublished results). It will be interesting to determine whether Btp1 is a substrate for the VirB T4SS and whether it has a ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved role in control of microtubules or microtubule-dependent vesicular trafficking during infection. One poorly characterized candidate effector is BvfA, a small protein of 11 kDa which is unique to the genus Brucella (Lavigne et al., 2005). It has been identified in a random screen using the Yersinia YopP as a reporter system, which induces apoptosis when the fusion protein is translocated into the host cytosol. Although the function of BvfA is unknown, it is necessary for intracellular survival within both human and murine macrophages, and bvfA mutants are highly attenuated in mice. Interestingly, the promoter of BvfA is induced intracellularly upon acidification but it is unclear whether this small protein predicted to be periplasmic is translocated into host cells. It would be worth re-analysing the translocation of BvfA into host cells using the TEM1 or CyA reporter system as it may be another substrate for the VirB T4SS. Its specific role during intracellular trafficking needs to be investigated. Brucella LPS Being a Gram-negative bacterium, Brucella is enclosed by an outer membrane containing LPS. LPS molecules consist of three major subcomponents (Fig. 3): (1) a lipid A, which serves to anchor the molecule in the outer membrane, (2) a sugar-based outer and inner core and (3) a chain of several sugar molecules, the O-polysaccharide that reaches into the extracellular space (Haag et al., 2010). Brucella strains naturally occur as strains with Smooth LPS that contains a terminal O-polysaccharide or Rough strains with LPS that is lacking the O-polysaccharide (Haag et al., 2010). Brucella strains found in human infections are generally characterized by a Smooth LPS phenotype. The outer membrane is the first site of contact between eucaryotic cell and bacterium. In the constant co-evolution of host and pathogen, LPS compounds have become a major pathogen-associated pattern (PAMP) recognized by innate immune cells to mount an efficient antibacterial response. The LPS of Brucella, however, has evolved to avoid these mechanisms. Brucella LPS reduces binding of complement to the bacterial surface, it does not induce pro-inflammatory responses in mice as indicated by no change in blood leucocyte numbers, absence of recruitment of polymorphonuclear neutrophils to the site of infection, low levels of pro-inflammatory cytokines and low cytotoxic activity (Moreno et al., 1981; BarqueroCalvo et al., 2007) which might represent a specific adaption to the intracellular lifestyle of this pathogen. The structure of Brucella LPS differs from the canonical pattern (Moreno & Moriyon, 2006). Its nonclassic lipid A possesses a diaminoglucose backbone and very long chain FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle 547 Fig. 3. Brucella LPS. The O-polysaccharide of Brucella abortus is composed of homopolymers of a-1,2-linked 4,6-dideoxy-4-formamido-a-Dmannopyranosyl (formyl-perosamine) subunits with an average chain length of 96 to100 subunits. The oligosaccharide core is still poorly characterized. It contains glucose, mannose, quinovosamine, glucosamine, 3-deoxy-D-manno-2-octulosonic acid (Kdo) and several unidentified sugars. Unlike other members of a-Proteobacteria, it lacks heptose, phosphates, galucturonic and glucuronic acid. The lipid A of Brucella is considered nonclassical because of the following particular features: (1) backbone is composed of 2,3-diamino-2,3-dideoxy-D-glucose (diaminoglucose) instead of glucosamine present in classical lipid A; (2) all acyl substitutions are in amide linkages; (3) sugar backbone is modified with unusually very long chain hydroxylated fatty acids. These remarkable features are highlighted in red boxes in the figure. fatty acid groups (Iriarte et al., 2004; Fig. 3). These particular features seem to be crucial for Brucella-specific infection responses. In contrast to the lipid A of other Gram-negative bacteria, that of Brucella is not efficiently recognized by TLR4 (Lapaque et al., 2006). Infection with a Brucella bacA mutant that is deficient in the very long chain fatty acid content of lipid A results in higher inflammation and the mutant is attenuated in mice (Ferguson et al., 2004; Parent et al., 2007). Likewise, the increased amounts of underacylated lipid A species in a Brucella bvrRS mutant (Manterola et al., 2005) may partially account for their attenuation in the mouse model of infection (Sola-Landa et al., 1998). Brucella LPS interferes with the MHCII-dependent antigen processing machinery of macrophages by clustering with MHCII molecules, likely resulting in a downregulated T cell activation (Forestier et al., 1999, 2000). The LPS O-polysaccharide crucially impacts outcome of infection with B. abortus, B. melitensis and B. suis. Strains bearing mutations in several genes involved in FEMS Microbiol Rev 36 (2012) 533–562 various stages of the LPS biosynthesis pathway such as the mannosyltransferase wbdA (Kohler et al., 2002) or the phosphomannomutase manB (Allen et al., 1998; Foulongne et al., 2000; Kohler et al., 2002) and whose mutation generally result in a ‘Rough’ phenotype are avirulent in cellular or animal infection models (Roop et al., 1991; Cheville et al., 1992; Winter et al., 1996; Allen et al., 1998; Elzer et al., 1998; Godfroid et al., 1998; McQuiston et al., 1999; Foulongne et al., 2000; Hong et al., 2000; Ugalde et al., 2000; Kohler et al., 2002; Lestrate et al., 2003). The degree of this attenuation depends on the particular gene and pathway affected. For instance, mutants with defective O-polysaccharide are able to replicate to varying levels, whereas those defective in synthesis of the LPS core are more sensitive to macrophage killing (Gonzalez et al., 2008). However, it has been suggested that the decrease in intra-macrophage survival of Rough mutants is not because of general defects in survival and persistence abilities but because of their cytotoxic potential which results in destruction of their replicative niche. The ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. 548 attenuation in vivo may therefore result from Rough strain cytotoxicity as well as several changes in strain infection biology. Rough Brucella strains induce more cytokine and chemokine production than their Smooth counterparts (Rittig et al., 2003). They induce maturation of human DCs (Billard et al., 2007), and their BCVs fuse more frequently with lysosomes (Porte et al., 2003) but also with neighbour phagosomes to form large communal vacuoles which cannot be observed with Smooth strain brucellae (Rittig et al., 2003). These differences in trafficking may result from a different mode of attachment and subsequent uptake. Brucella strains with Rough LPS show an increased attachment to and uptake into monocytes and macrophages (Rittig et al., 2003; Pei & Ficht, 2004), indicating that they may enter cells upon interaction with receptors different from those utilized during uptake of Smooth strains. A more efficient uptake into cells can also be observed in vivo: Smooth Brucella remain sensitive to Gentamicin treatment up to 24 h postintraperitoneal injection, whereas Rough strains are insensitive to the antibiotic and therefore completely internalized into cells as early as 1 h postinfection (Turse et al., 2011). Brucella strains with a Smooth LPS are thought to be internalized upon interaction with lipid rafts which essentially determines their ability for short-term intracellular survival, whereas short-term survival of Rough B. suis is not dependent on interaction with lipid rafts (Naroeni et al., 2001; Porte et al., 2003). The spontaneous appearance of Rough mutants derived from Smooth Brucella strains has been known for a long time. This conversion from Smooth to Rough LPS occurs in culture but also during animal infection, likely due to integration of the unstable genetic element genomic island 2 (Mancilla et al., 2010). A spontaneous mutation resulting in a Rough phenotype can be observed 100–1000 times more frequently than spontaneous Brucella pyrimidine auxotrophs (Turse et al., 2011). The fact that Rough strains are eliminated by the host quickly and efficiently raises the question as to why, over time and under these selective pressures, Brucella maintains an intrinsic instability of one of its most essential means of intra-host survival. One explanation would be that LPS conversion actually serves a function during infection that, in the long run, favours bacterial shortterm survival and/or persistence (e.g. as described below: Brucella and host cell death). However, because so far there exist no experimental means to suppress LPS conversion, its actual role during animal infection remains to be investigated. Although a Smooth LPS seems to be a major determinant of intra-host survival for most Brucella strains, the two species B. canis and B. ovis naturally lack the O-polysaccharide yet cause infection in their respective hosts ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved (dogs or sheep and rams). As with in vitro generated Rough mutants, these naturally Rough strains are internalized more efficiently into host cells than Smooth brucellae without the requirement of opsonization (Detilleux et al., 1990; Fernandez-Prada et al., 2003; Rittig et al., 2003; Chen & He, 2009; Ferrero et al., 2009). Their intracellular multiplication is reduced compared with that of Smooth Brucella strains (Detilleux et al., 1990; Rittig et al., 2003; Ferrero et al., 2009; Martin-Martin et al., 2009) and in the infection models used; phagosomes containing B. canis and B. ovis fuse more frequently with lysosomes than those of Smooth Brucella strains (Porte et al., 2003; Rittig et al., 2003). In contrast to Rough mutants of B. abortus or melitensis, the naturally Rough strains B. ovis or canis have no cytopathic effects (Pei & Ficht, 2004). Cyclic glucan Like their relatives Agrobacterium and Rhizobium, Brucella strains produce periplasmic cyclic ß-1,2-glucans (CßG) (Ugalde, 1999). In plant pathogens, CßG has been described as a factor essential for endosymbiontic invasion of nodules by Rhizobium meliloti or tumour induction by A. tumefaciens, respectively (Breedveld et al., 1994). A central enzyme in Brucella CßG synthesis is cyclic glucan synthetase (Cgs) (Inon de Iannino et al., 1998). Brucella strains defective in CßG synthesis have a decreased survival in mice and do not replicate in HeLa cells and mouse peritoneal macrophages (Briones et al., 2001). Interestingly, cyclic glucan is not required for Brucella replication in DCs (Salcedo et al., 2008). The functions of cyclic glucan in bacterial physiology and during infection are not yet fully understood. Cyclic b-1,2-glucan mutants of Agrobacterium have multiple altered cell surface properties including loss of motility as a result of defective flagellum assembly (Breedveld et al., 1994). Likewise, Brucella cgs mutants seem to have general defects in their membranes, as suggested by their higher sensitivity to surface-active molecules (Briones et al., 2001). However, although a cgs mutant of the B. abortus wild-type strain or of the attenuated vaccination strain B. abortus S19 shows the same sensitivity against surfaceactive reagents, respectively, the former is much less attenuated than the latter, suggesting that membrane alterations are not the main cause for the decreased persistence (Briones et al., 2001). During infections, cyclic glucan might be released from periplasmic space via outer membrane vesicles (Briones et al., 2001), allowing its interaction with host cell components. It has been proposed that it targets cholesterolrich lipid rafts found on the BCV membrane to control the vacuole’s interactions with the endocytic pathway FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle 549 (Arellano-Reynoso et al., 2005). Consistent with this hypothesis, purified Brucella CßG was shown to extract cholesterol from eucaryotic membranes. Furthermore, CßG enhanced recruitment of flotillin-1 to BCVs, a signalling molecule characteristic of lipid rafts involved in control of phagosome maturation. Brucella CßG mutants reside in vacuoles that fuse with lysosomes where the bacteria are degraded (Arellano-Reynoso et al., 2005). This trafficking defect is restored when the mutant strain is grown with purified CßG prior to infection (ArellanoReynoso et al., 2005). Brucella regulation of gene expression Bacterial genes mediating intra-host survival and intracellular trafficking diversion are usually subject to complex regulatory networks orchestrating gene expression in the required amount at the appropriate phase of infection. Several gene regulatory systems of Brucella belonging to different protein families have been correlated to infection success: • Two-component regulatory systems (TCS). These systems represent a common bacterial mechanism to adapt to environmental changes. A transmembrane sensor kinase is reacting by autophosphorylation to a particular extracellular signal, for example, a drop in pH or an increase in temperature TCS; the phosphate group is then transferred to one or several response regulators that mediate the change in transcription (Casino et al., 2010). TCS mediating Brucella intra-host replication include FeuQ, NtrY, VsrB, OmpR and BvrR/BvrS. FeuQ encodes the putative sensor kinase of the FeuP/FeuQ system (Lestrate et al., 2003) that mediates the regulation of high affinity iron uptake in Rhizobium leguminosarum (Yeoman et al., 1997). However, a B. suis mutant of the corresponding feuP response regulator replicates on wild-type levels in mice and macrophages (Dorrell et al., 1998), suggesting either species-specific differences (melitensis vs. suis) or that FeuP is not the only regulator responding to feuQ signalling (Lestrate et al., 2003). Additional TCS regulating Brucella virulence are the putative nitrogen responsive factor NtrY (B. suis, Foulongne et al., 2000), VsrB (B. melitensis, Lestrate et al., 2000) and OmpR (B. melitensis, Wu et al., 2006). One of the best studied Brucella TCS is the BvrR/BvrS system which was also one of the first global gene expression regulator systems involved in Brucella virulence identified (Sola-Landa et al., 1998). Mutants of this TCS have a Smooth type LPS yet they do not persist in mouse spleens, they poorly invade and do not FEMS Microbiol Rev 36 (2012) 533–562 • replicate in macrophages and HeLa cells, their vacuoles show enhanced fusion with lysosomes and they have a decreased resistance to antimicrobial peptide polymyxin B (Sola-Landa et al., 1998) which might result from changes in the outer membrane (Manterola et al., 2005). Interestingly, there is a small but significant difference between the residual persistence of the bvrR and the bvrS mutant in mice (Sola-Landa et al., 1998), suggesting that the lack of the sensor protein results in a more severe phenotype. In contrast to wild-type bacteria, bvrS mutants do not recruit the small GTPases of the Rho subfamily essential for actin polymerization and they do not activate Cdc42 Brucella that are all crucial for Brucella uptake into cells (Guzman-Verri et al., 2001). BvrR/BvrS regulates expression of more than 100 genes including those involved in cell envelope modulation, carbon and nitrogen metabolism, and factors that have been shown to be involved in virulence such as the VirB T4SS and the LuxR-type regulator VjbR (Lamontagne et al., 2007; MartinezNunez et al., 2010; Viadas et al., 2010). Quorum-sensing regulatory systems. Quorum sensing is based on signalling molecules that can transmit messages between individual bacteria and their corresponding regulators. It is an essential bacterial mechanism to adequately respond to environmental changes and also for virulence of many bacterial pathogens (Antunes et al., 2010) including Brucella (Rambow-Larsen et al., 2009). Brucella species synthesize a C12-homoserine lactone quorum-sensing signal (Taminiau et al., 2002), and five putative Brucella LuxR-like transcriptional regulators have been identified, though only two (VjbR and BlxR) have been investigated more closely and only one (VjbR) seems to be crucially involved in virulence (Delrue et al., 2005; Rambow-Larsen et al., 2008; Weeks et al., 2010). The LuxR-type quorum-sensing regulator VjbR (Delrue et al., 2005) controls more than 100 genes including outer membrane proteins, the VirB operon, the cyclic β-1,2-glucan synthetase Cgs and genes encoding flagella-related gene products such as FliC (Uzureau et al., 2007; Weeks et al., 2010). VjbR mutants are attenuated in macrophages, HeLa cells and the mouse model of infection (Delrue et al., 2005; ArenasGamboa et al., 2008). The LuxR-type regulator BlxR (Rambow-Larsen et al., 2008) also affects expression of the virB operon and flagella-related genes. BlxR and VjbR are positively autoregulated and they cross-regulate each other (Rambow-Larsen et al., 2008). However, compared to VjbR, BlxR mutation has negligible effects on B. melitensis 16M virulence (Rambow-Larsen et al., 2008; Weeks et al., 2010) suggesting an overlap but not redundancy in regulated genes. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. 550 • • • • The stringent response. Brucella expresses the 3′,5′-bispyrophosphate (ppGpp) synthetase (Rsh for RelA/SpoT homologue). This enzyme produces ppGpp in response to starvation to induce global changes of adaptive responses to cope with low nutrient situations. The trigger is thought to be an amino acid-free tRNA molecule binding to the ribosome, leading to guanosine pentaphosphate (pppGpp) production by the RelA protein. pppGpp is hydrolysed to guanosine tetraphosphate (ppGpp) which is believed to associate with the RNA polymerase to modify its promoter specificity (Wells & Long, 2003). In correspondence with the BCV being a low nutrient environment, Brucella rsh mutants are less resistant to starvation and are attenuated in macrophages, HeLa cells and mice (Kohler et al., 2002; Kim et al., 2005a; Dozot et al., 2006). Starvation also induces virB operon genes (Boschiroli et al., 2002) which are targets of Rsh regulation (Dozot et al., 2006). LysR-type transcription regulators. Members of the family of LysR-type of transcriptional regulators (LTTRs) are the most abundant type of transcriptional factors in prokaryotes (Maddocks & Oyston, 2008). Several LTTRs regulate Brucella intra-host survival although none of them has been fully investigated (Foulongne et al., 2000; Wu et al., 2006). Additional transcriptional regulators. Several other regulators of Brucella belonging to different protein families have been found to be involved in virulence. A mutant of a GntR family transcription regulator is attenuated both in mice and in cellular infection models (Haine et al., 2005). Several more transcription regulators of this family are involved in virulence in the mouse, although interestingly, most of them are not attenuated in cellular infection models (Lestrate et al., 2003; Haine et al., 2006). Attenuated Brucella mutants of transcription regulators include those disrupted in DeoR (Kohler et al., 2002; Wu et al., 2006), MerR (Wu et al., 2006) and MucR (Wu et al., 2006). More recently, HutC, a transcriptional repressor of histidine utilization genes (Sieira et al., 2010), was also implicated in virulence as well as a light-responsive histidine kinase carrying a light, oxygen or voltage (LOV) domain (Swartz et al., 2007). Nonlight responsive mutants of this protein are attenuated in mouse macrophages (Swartz et al., 2007). Small regulatory RNAs. In addition to regulation by protein factors, control of bacterial gene expression can include small regulatory RNAs (sRNAs) which may bind to mRNA to regulate their translation or degradation (Zhou & Xie, 2011). Some of these RNAs require the chaperone hfq for interaction with their targets. Factors whose expression is controlled by Hfq ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved include the quorum-sensing regulator BlxR and the VirB T4SS (Caswell et al., 2012). A B. abortus hfq mutant is more sensitive to several environmental stresses, including H2O2, acid and starvation (Robertson & Roop, 1999), and is avirulent in murine and human macrophages as well as in mice (Robertson & Roop, 1999; Bellaire et al., 2005). A corresponding mutant of B. melitensis is attenuated in goats (Roop et al., 2002). Although the functions of several of these regulatory elements seem to converge in control of expression of the factors such as the T4SS, deletions in these regulators result in more or less pronounced attenuation. This lack of redundancy suggests individual roles for these transcriptional factors and supports the notion of a versatile and complex regulatory network of Brucella gene expression. Brucella and host cell death The manipulation of programmed death of cells is one of the key mechanisms of pathogens to promote their intrahost survival (Lamkanfi & Dixit, 2010). Several bacterial pathogens such as C. burnetii (Luhrmann et al., 2010) or Salmonella enterica serovar Typhimurium (Lindgren et al., 1996) interfere with the cell’s programmed death pathways in promoting or suppressing them which may support infection stage specific bacterial actions such as dissemination or long-term intracellular persistence. Brucellae seem to belong to the category of cell death inhibiting pathogens. Brucella strains with a Smooth LPS phenotype (see below) inhibit the programmed cell death (Fig. 4) in murine and human macrophages (Gross et al., 2000; Tolomeo et al., 2003; He et al., 2006). Infection with a Smooth B. suis strain suppresses spontaneously occurring as well as interferon-gamma or Fas-induced apoptosis in human monocytes (Gross et al., 2000). Likewise, serum-deprived human monocytes are protected from apoptosis by B. melitensis infection (Fernandez-Prada et al., 2003). A delayed apoptosis has been shown for lymphocytes and monocytes of naturally Brucella-infected cattle compared with healthy or vaccinated controls (Galdiero et al., 2000). An increased resistance to spontaneous or induced apoptosis has also been shown for monocytes and lymphocytes of brucellosis patients (Tolomeo et al., 2003). Interestingly, this resistance is reversed more efficiently in acute brucellosis patients compared with chronic patients following antibiotic therapy (Tolomeo et al., 2003). These protective effects require bacterial survival (Gross et al., 2000), they are not mediated by Brucella LPS alone (Gross et al., 2000) and might be induced by a soluble factor as suggested by protection from apoptosis of cells FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle 551 Fig. 4. Brucella manipulation of macrophage cell death. (a) (1) Macrophages/monocytes infected with virulent smooth Brucella strains are protected from spontaneous cell death or from death induced by interferon-c treatment, serum deprivation or Fas. This protection might be mediated by inhibition of cytochrome C and reactive oxygen species release by mitochondria, upregulation of the anti-apoptotic Bcl-2 family member A1, downregulation of pro-apoptotic Bcl-2 family members and caspase-3 (2) and depends on bacterial viability. Uninfected cells of the same sample are likewise protected from cell death. (3) However, with overexpression of the VirB secretion system, infection with smooth brucellae becomes cytotoxic and results in macrophage cell death. (4) Cytopathic effects also occur during infection with high bacterial numbers. This cytotoxicity can be reduced by (5) deletion of the VirB secretion system. (b) (1) Infection with several Rough mutant derivatives of Brucella strains induces a caspase-2-dependent cell death that involves secretion of proinflammatory cytokines and NFkB activation. Induction of cell death requires bacterial protein biosynthesis, (2) the uptake of bacteria into the cell, (3) bacterial viability and (4) the VirB secretion system. Noninfected neighbour cells are not affected from Rough strain cytotoxicity. which are not themselves infected (Gross et al., 2000; Tolomeo et al., 2003). The mechanisms could involve increased expression of the survival-promoting protein of the bcl-2 family A1 and/or suppression of pro-apoptotic proteins of the same family as well as several other proapoptotic proteins including caspase-3 (Gross et al., 2000; Eskra et al., 2003). Another study based on microarray analysis of B. melitensis infected macrophages has found that bacteria may inhibit apoptosis by blocking mitochondrial release of cytochrome c as well as the production of reactive oxygen species which may prevent caspase activation (He et al., 2006). In contrast to their Smooth counterparts, various Rough mutants of these strains have been found to be cytotoxic (Fernandez-Prada et al., 2003; Pei & Ficht, 2004; Pei et al., 2006; Chen & He, 2009; Chen et al., 2011; Fig. 4). Because they are spontaneously generated during animal infection, Rough mutants could provide an exit strategy for their Smooth parental bacteria in the same intracellular vacuole to leave their niche for cell-to-cell spread (Turse et al., 2011). The type of cell death induced by live, Rough B. abortus and B. melitensis strains includes necrotic features (Pei & Ficht, 2004; Pei et al., 2006) as well as those of the mitochondrial apoptosis pathway (Chen & He, 2009) and seems to be specific for macrophages whereas epithelial cells are not affected (Pei & Ficht, 2004). A major difference of this particular cell FEMS Microbiol Rev 36 (2012) 533–562 death from classical apoptosis is the involvement of a robust pro-inflammatory response including TNF-a and IL-1b secretion and activation of NF-jB (Chen et al., 2011). It is also different from what has been described as ‘pyroptotic cell death’ because it is not mediated by caspase-1 but by caspase-2 activity (Chen & He, 2009; Chen et al., 2011), which occurs at very early time points of infection with Rough Brucella strains (Chen & He, 2009). The cell death induced by Rough strains seems to be dominant compared with the protective effects of Smooth Brucellae: preinfection of cells with Smooth Brucella strains or pretreatment with their LPS does not protect from the cell death induced by their Rough derivatives (Pei et al., 2006). Whereas the anti-apoptotic effects of their wildtype parental strains also protect noninfected cells, Rough strain-induced cytotoxicity is likely not mediated by a soluble factor (such as TNF-a or nitric oxide), but rather requires the direct contact of host cell and bacterium (Pei et al., 2006). Furthermore, it requires bacterial uptake, viability and protein biosynthesis (Pei et al., 2006). One of the factors that has been related to interference with host cell death is the VirB secretion system. Although Smooth brucellae generally inhibit programmed cell death in murine and human macrophages, infection with B. melitensis becomes increasingly cytotoxic for macrophages with increasing multiplicities of infection (Zhong et al., 2009). These Smooth strain cytopathic effects can be ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 552 strongly reduced by deletion of the VirB secretion system (Zhong et al., 2009). Correspondingly, overexpression of VirB considerably enhances the cytotoxicity and results in a decrease in virulence for mice (Zhong et al., 2009). Likewise, the T4SS seems to be involved in Rough strain cytotoxicity. In a screen for factors involved in mediating these cytopathic effects by transposon mutation of a Rough B. melitensis strain, almost two-third of the open reading frames whose disruption resulted in a loss of cytotoxicity induction were part of the VirB operon (Pei et al., 2008b). At the same time, elimination of T4SS expression in Rough mutants results in enhanced replication and persistence (Pei et al., 2008b). All of these findings are consistent with the hypothesis that the cytotoxic effects of Rough Brucella mutants may result from enhanced secretion by the VirB system (Zhong et al., 2009). It has been shown that T3SSmediated secretion of effectors is increased in Shigella with shorter LPS (West et al., 2005). This enhanced secretion is thought to result from increased efficiency of the secretion system because of steric effects. A similar effect could lead to an uncontrolled effector secretion in Rough Brucella mutants, resulting in an effector ‘overdose’ and host cell death. However, different Rough mutants show variable levels of cytotoxicity; some are not cytotoxic at all (Pei & Ficht, 2004; Pei et al., 2006), and there are no cytopathic effects during cell infection with the naturally Rough strain B. ovis (Martin-Martin et al., 2008), suggesting that the correlation of LPS phenotype, VirB-mediated secretion and cytotoxic potential are characterized by strain-specific differences. The attenuated phenotype of a Rough mutant strain may not just result from a difference in intracellular trafficking, but also from the cytopathic effects of Rough brucellae on infected cells which might counteract their effective colonization of the Brucella intracellular niche inside of their hosts. The inhibition of Rough straininduced cell death promotes bacterial intracellular survival (Chen & He, 2009). Likewise, the stability in and, at later time points, decline of Rough strain bacterial numbers in macrophages might not result from bacterial killing by macrophage microbicidal actions, but rather macrophage death and release of bacteria into the extracellular media, where they are not quantified in gentamicin-protection assays (Pei & Ficht, 2004). A similar effect might take place inside the host, exposing bacteria from their protective environment inside of cells and rendering them vulnerable to humoral antibacterial mechanisms (Pei & Ficht, 2004). Future directions The understanding of the mechanisms that underlie the pathogenesis of Brucella has greatly advanced in the past ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. 10 years. The availability of genome sequences has contributed to this success, in particular, in view of the growing number of newly isolated Brucella species, some of which also have pathogenic potential for humans. Some recent exciting studies are highlighted in Table 2. These include use of large-scale methodology such as the classical STM, allowing the identification of key factors of Brucella intra-host survival (Lestrate et al., 2003) and transcriptomic approaches that enable a global view on Brucella gene expression profiles throughout specific stages of the infection (Rossetti et al., 2010; Viadas et al., 2010). Proteomic characterization of the BCV membrane composition revealed important host factors involved in Brucella intracellular replication (Fugier et al., 2009). This study was strengthened by the identification of a Brucella effector protein involved, via a high-throughput yeast two-hybrid screen that defined the interactome map between predicted Brucella proteins and human phagosomal proteins (de Barsy et al., 2011). Directed RNAi screens are powerful tools to identify novel eucaryotic molecules relevant during a specific step of the infection, as demonstrated by the ER-specific RNAi screen carried out in Drosophila S2 cells (Qin et al., 2008). This screen has identified a very extensive and interesting list of eucaryotic proteins, including numerous proteins involved in vesicle transport along the secretory pathway, that are likely targeted by Brucella and should therefore be further characterized. The recent identification of several Brucella-translocated effector proteins (de Jong et al., 2008; de Barsy et al., 2011; Marchesini et al., 2011) will greatly advance the study of how Brucella manipulates and interacts with host cell pathways at the molecular level. It will enable identification of the eucaryotic targets, which may include additional Rab GTPases or microtubule-interacting proteins that might enable establishment of the ER-derived compartment where Brucella replicates. For instance, the small GTPases Rab3d and Rab9 as well as the kinesin motor proteins Kif1 and Kif4 have been shown to be downregulated in Brucellainfected macrophages (Eskra et al., 2003). Perhaps some of these effectors are dedicated to inhibiting host cell death or controlling the fusion events between BCVs and late endosomes and lysosomes (Starr et al., 2008). These studies will be reinforced by use of advanced imaging technology, including live imaging of Brucella-infected cells (Starr et al., 2008) as well as the use of bioluminescent strains to follow the infection in vivo (Rajashekara et al., 2005). Integration of in vitro studies in the context of in vivo models and in natura brucellosis taking into account the complex interplay of cellular and immune factors that contribute to disease development may help to close the gap separating results of clinical and fundamental research. Several target cells have been identified and have FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle 553 Table 2. Techniques that provided important advances in understanding Brucella intracellular pathogenesis Technique Objective Advantages Disadvantages Reference (1) Transposon mutagenesis and (2) Signaturetagged transposon mutagenesis Microarray technology Identification of virulence determinants (1) and (2) Unbiased screen; (2) reduced number of mice possible preselection of mutant pools (e.g. by growth in minimal media to exclude auxotrophs) (1) Kohler et al. (2002); Delrue et al. (2001) (2) Foulongne et al. (2000); Lestrate et al. (2003) Characterization of host cell transcriptional responses to Brucella infection Thorough analysis of induction/suppression of eucaryotic pathways following Brucella infection Bioluminescent Brucella strains in vivo Kinetic profiles of bacterial organ invasion over time Live imaging of infected cells Observation of trafficking events following individual bacteria Kinetic analysis in the same individual; thorough in vivo studies of bacterial dissemination Avoid artifacts because of fixation; information on the dynamics of the events; kinetic analysis of the fate of single bacteria (2) Possible complementation of attenuated mutants by nonattenuated strains; masking of subtle effects by the high bacterial loads required to obtain individual signals for each mutant May yield results different from changes on proteomic level because of post-transcriptional regulation Limited resolution and sensitivity with respect to bacterial load observed RNAi screen in host cells such as S2 Drosophila cells Genome-wide loss-offunction screen in infected cells Proteomics of BCV membrane Determination of eucaryotic and prokaryotic proteins on vacuole Amplification of Brucella mRNA from infected samples Generation of intracellular Brucella gene expression profiles High-throughput yeast two-hybrid screen Identify potential interactions between subset of eucaryotic proteins and Brucella proteins Identification of effector molecules Bioinformatics Possibility of directed screens (e.g. ER-directed) relevant for Brucella; identification of conserved eucaryotic proteins involved in Brucella virulence Thorough and unbiased information on the recruitment of eucaryotic proteins to the vacuole Analysis of whole genome or multiple gene sets transcripts in same experiment; global picture of regulation Directed screening of whole bacterial ORFeome against specific baits Exhaustive analysis of potential effectors including those that would not be found in mutant screenings because of mild effects or functional redundancies been extensively studied in vitro, including macrophages, trophoblasts, DCs and epithelial cells but their relevance and particular role in the course of animal and even more human infection is poorly understood. Although macrophages seem to be essential to reduce the initial bacterial load of infection (Archambaud et al., 2010), they seem to serve as permanent residence for Brucella during persistence of disease. In contrast to other cell types, macro- FEMS Microbiol Rev 36 (2012) 533–562 Eskra et al. (2003) Rajashekara et al. (2005) Over-expression of markers may affect their location and/or function; time-consuming to analyse statistical significant number of bacteria and events Dependent on the efficiency of the knock-down; temperature limitation for S2 cells; Starr et al. (2008) Difficult to obtain good purity and enough quantity material Fugier et al. (2009) Only transcript level (not protein); only relative quantification Rossetti et al. (2010); Viadas et al. (2010) Nonspecificity of some interactions; de Barsy et al. (2011) High number of candidate effectors to test for translocation in infected cells de Jong et al. (2008); Marchesini et al. (2011) Qin et al. (2008) phages are able to initially kill a high percentage of internalized brucellae which are otherwise perfectly adapted to intra-host survival, raising the question whether this killing phase may serve a function during Brucella infection. Little is known as to where and how the bacteria cross the epithelial barriers of their hosts, which cells are targeted first, if/how a switch of host cells takes place and how these processes relate to the particular ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 554 intracellular trafficking of Brucella. The mouse model of brucellosis should be further exploited with all the new tools available to decipher in greater detail the cellular players and the immune response during infection, maybe also using the recently described rodent strain B. microti. How the intracellular lifestyle is contributing to Brucella persistence during the chronic stage of disease is an important question that merits further investigation. Intracellular replication to high numbers that has been observed in vitro and described, for example, in trophoblasts of infected pregnant animals may enable cell-to-cell spread and disruption of specific tissue or barriers. As discussed above, control of cell death may also regulate bacterial escape from cells and dissemination. Perhaps, in some cells, Brucella is able to survive but restrain its intracellular growth in a way that would establish a bacterial reservoir. Consistent with this possibility, cells with only a few bacteria are often observed in cultured cells even at very late time points after infection. Analysis of the viability of these bacteria needs to be undertaken to determine whether they could represent a latent virulent Brucella. Although Brucella induces a minimal inflammatory response in the host, it is not silent and has therefore acquired proteins that help modulate host innate and adaptive immune response mechanisms that may be important in the development of a chronic infection. Besides Btp1, another of these factors is PrpA, which belongs to the proline-racemase family. It is required for chronic infection in mice. It does not seem specifically associated with intracellular survival but instead, it is involved in immune modulation in the host, by eliciting B-lymphocyte polyclonal activation and IL-10 secretion (Spera et al., 2006). Additional factors are likely to be involved in persistence of Brucella within the host. As summarized in this review, great advances in the field of Brucella intracellular trafficking have been made in recent years. To integrate the results from studies at the cell and organism levels will open new perspectives and eventually allow for a deeper understanding of the complex mechanisms of brucellosis. Acknowledgements This work was supported by the CNRS (J.P.G.), INSERM (S.S.) and ANR BruTir (J.P.G., K.V.B. and S.S.). We thank Steve Garvis for critical reading of the manuscript. References Akopyan K, Edgren T, Wang-Edgren H, Rosqvist R, Fahlgren A, Wolf-Watz H & Fallman M (2011) Translocation of surface-localized effectors in type III secretion. P Natl Acad Sci USA 108: 1639–1644. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. Allen CA, Adams LG & Ficht TA (1998) Transposon-derived Brucella abortus rough mutants are attenuated and exhibit reduced intracellular survival. Infect Immun 66: 1008–1016. Alvarez-Martinez CE & Christie PJ (2009) Biological diversity of prokaryotic type IV secretion systems. Microbiol Mol Biol Rev 73: 775–808. Anderson TD & Cheville NF (1986) Ultrastructural morphometric analysis of Brucella abortus-infected trophoblasts in experimental placentitis. Bacterial replication occurs in rough endoplasmic reticulum. Am J Pathol 124: 226–237. Anderson ES, Paulley JT, Gaines JM, Valderas MW, Martin DW, Menscher E, Brown TD, Burns CS & Roop RM II (2009) The manganese transporter MntH is a critical virulence determinant for Brucella abortus 2308 in experimentally infected mice. Infect Immun 77: 3466–3474. Antunes LC, Ferreira RB, Buckner MM & Finlay BB (2010) Quorum sensing in bacterial virulence. Microbiology 156: 2271–2282. Archambaud C, Salcedo SP, Lelouard H, Devilard E, de Bovis B, Van Rooijen N, Gorvel JP & Malissen B (2010) Contrasting roles of macrophages and dendritic cells in controlling initial pulmonary Brucella infection. Eur J Immunol 40: 3458–3471. Arellano-Reynoso B, Lapaque N, Salcedo S, Briones G, Ciocchini AE, Ugalde R, Moreno E, Moriyon I & Gorvel JP (2005) Cyclic beta-1,2-glucan is a Brucella virulence factor required for intracellular survival. Nat Immunol 6: 618–625. Arenas-Gamboa AM, Ficht TA, Kahl-McDonagh MM & RiceFicht AC (2008) Immunization with a single dose of a microencapsulated Brucella melitensis mutant enhances protection against wild-type challenge. Infect Immun 76: 2448–2455. Ariza J, Bosilkovski M, Cascio A et al. (2007) Perspectives for the treatment of brucellosis in the 21st century: the Ioannina recommendations. PLoS Med 4: e317. Audic S, Lescot M, Claverie JM & Scholz HC (2009) Brucella microti: the genome sequence of an emerging pathogen. BMC Genomics 10: 352. Baldwin CL & Goenka R (2006) Host immune responses to the intracellular bacteria Brucella: does the bacteria instruct the host to facilitate chronic infection? Crit Rev Immunol 26: 407–442. Barbier T, Nicolas C & Letesson JJ (2011) Brucella adaptation and survival at the crossroad of metabolism and virulence. FEBS Lett 585: 2929–2934. Barquero-Calvo E, Chaves-Olarte E, Weiss DS, Guzman-Verri C, Chacon-Diaz C, Rucavado A, Moriyon I & Moreno E (2007) Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PLoS ONE 2: e631. Barrionuevo P, Delpino MV, Velasquez LN, Garcia Samartino C, Coria LM, Ibanez AE, Rodriguez ME, Cassataro J & Giambartolomei GH (2011) Brucella abortus inhibits FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle IFN-gamma-induced FcgammaRI expression and FcgammaRI-restricted phagocytosis via toll-like receptor 2 on human monocytes/macrophages. Microbes Infect 13: 239–250. Bellaire BH, Roop RM II & Cardelli JA (2005) Opsonized virulent Brucella abortus replicates within nonacidic, endoplasmic reticulum-negative, LAMP-1-positive phagosomes in human monocytes. Infect Immun 73: 3702– 3713. Bergagna S, Zoppi S, Ferroglio E, Gobetto M, Dondo A, Di Giannatale E, Gennero MS & Grattarola C (2009) Epidemiologic survey for Brucella suis biovar 2 in a wild boar (Sus scrofa) population in northwest Italy. J Wildl Dis 45: 1178–1181. Billard E, Cazevieille C, Dornand J & Gross A (2005) High susceptibility of human dendritic cells to invasion by the intracellular pathogens Brucella suis, B. abortus, and B. melitensis. Infect Immun 73: 8418–8424. Billard E, Dornand J & Gross A (2007) Brucella suis prevents human dendritic cell maturation and antigen presentation through regulation of tumor necrosis factor alpha secretion. Infect Immun 75: 4980–4989. Boschiroli ML, Ouahrani-Bettache S, Foulongne V, MichauxCharachon S, Bourg G, Allardet-Servent A, Cazevieille C, Liautard JP, Ramuz M & O’Callaghan D (2002) The Brucella suis virB operon is induced intracellularly in macrophages. P Natl Acad Sci USA 99: 1544–1549. Breedveld MW, Yoo JS, Reinhold VN & Miller KJ (1994) Synthesis of glycerophosphorylated cyclic beta-(1,2)-glucans by Rhizobium meliloti ndv mutants. J Bacteriol 176: 1047– 1051. Briones G, Inon de Iannino N, Roset M, Vigliocco A, Paulo PS & Ugalde RA (2001) Brucella abortus cyclic beta-1,2glucan mutants have reduced virulence in mice and are defective in intracellular replication in HeLa cells. Infect Immun 69: 4528–4535. Bukata L, Altabe S, de Mendoza D, Ugalde RA & Comerci DJ (2008) Phosphatidylethanolamine synthesis is required for optimal virulence of Brucella abortus. J Bacteriol 190: 8197– 8203. Caron E, Gross A, Liautard JP & Dornand J (1996) Brucella species release a specific, protease-sensitive, inhibitor of TNF-alpha expression, active on human macrophage-like cells. J Immunol 156: 2885–2893. Casadevall A & Pirofski LA (1999) Host-pathogen interactions: redefining the basic concepts of virulence and pathogenicity. Infect Immun 67: 3703–3713. Casino P, Rubio V & Marina A (2010) The mechanism of signal transduction by two-component systems. Curr Opin Struct Biol 20: 763–771. Castaneda-Roldan EI, Avelino-Flores F, Dall’Agnol M, Freer E, Cedillo L, Dornand J & Giron JA (2004) Adherence of Brucella to human epithelial cells and macrophages is mediated by sialic acid residues. Cell Microbiol 6: 435–445. Castaneda-Roldan EI, Ouahrani-Bettache S, Saldana Z, Avelino F, Rendon MA, Dornand J & Giron JA (2006) Characterization of SP41, a surface protein of Brucella FEMS Microbiol Rev 36 (2012) 533–562 555 associated with adherence and invasion of host epithelial cells. Cell Microbiol 8: 1877–1887. Caswell CC, Gaines JM & Roop RM II (2012) The RNA chaperone Hfq independently coordinates expression of the VirB Type IV secretion system and the LuxR-type regulator BabR in Brucella abortus 2308. J Bacteriol 194: 3–14. Celli J, de Chastellier C, Franchini DM, Pizarro-Cerda J, Moreno E & Gorvel JP (2003) Brucella evades macrophage killing via VirB-dependent sustained interactions with the endoplasmic reticulum. J Exp Med 198: 545–556. Celli J, Salcedo SP & Gorvel JP (2005) Brucella coopts the small GTPase Sar1 for intracellular replication. P Natl Acad Sci USA 102: 1673–1678. Chaudhary A, Ganguly K, Cabantous S, Waldo GS, MichevaViteva SN, Nag K, Hlavacek WS & Tung CS (2011) The Brucella TIR-like protein TcpB interacts with the death domain of MyD88. Biochem Biophys Res Commun 417: 299– 304. Chaves-Olarte E, Guzman-Verri C, Meresse S, Desjardins M, Pizarro-Cerda J, Badilla J, Gorvel JP & Moreno E (2002) Activation of Rho and Rab GTPases dissociates Brucella abortus internalization from intracellular trafficking. Cell Microbiol 4: 663–676. Chen F & He Y (2009) Caspase-2 mediated apoptotic and necrotic murine macrophage cell death induced by rough Brucella abortus. PLoS ONE 4: e6830. Chen F, Ding X, Ding Y, Xiang Z, Li X, Ghosh D, Schurig GG, Sriranganathan N, Boyle SM & He Y (2011) Proinflammatory caspase-2-mediated macrophage cell death induced by a rough attenuated Brucella suis strain. Infect Immun 79: 2460–2469. Cheville NF, Jensen AE, Halling SM, Tatum FM, Morfitt DC, Hennager SG, Frerichs WM & Schurig G (1992) Bacterial survival, lymph node changes, and immunologic responses of cattle vaccinated with standard and mutant strains of Brucella abortus. Am J Vet Res 53: 1881–1888. Cirl C, Wieser A, Yadav M et al. (2008) Subversion of Tolllike receptor signaling by a unique family of bacterial Toll/ interleukin-1 receptor domain-containing proteins. Nat Med 14: 399–406. Cloeckaert A, Verger JM, Grayon M, Paquet JY, Garin-Bastuji B, Foster G & Godfroid J (2001) Classification of Brucella spp. isolated from marine mammals by DNA polymorphism at the omp2 locus. Microbes Infect 3: 729–738. Comerci DJ, Martinez-Lorenzo MJ, Sieira R, Gorvel JP & Ugalde RA (2001) Essential role of the VirB machinery in the maturation of the Brucella abortus-containing vacuole. Cell Microbiol 3: 159–168. Conde-Alvarez R, Grillo MJ, Salcedo SP, de Miguel MJ, Fugier E, Gorvel JP, Moriyon I & Iriarte M (2006) Synthesis of phosphatidylcholine, a typical eukaryotic phospholipid, is necessary for full virulence of the intracellular bacterial parasite Brucella abortus. Cell Microbiol 8: 1322–1335. Cvetnic Z, Spicic S, Toncic J, Majnaric D, Benic M, Albert D, Thiebaud M & Garin-Bastuji B (2009) Brucella suis infection ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 556 in domestic pigs and wild boar in Croatia. Rev Sci Tech 28: 1057–1067. Czibener C & Ugalde JE (2012) Identification of a unique gene cluster of Brucella spp. that mediates adhesion to host cells. Microbes Infect 14: 79–85. de Barsy M, Jamet A, Filopon D et al. (2011) Identification of a Brucella spp. secreted effector specifically interacting with human small GTPase Rab2. Cell Microbiol 13: 1044–1058. de Jong MF & Tsolis R (2012) Brucellosis and type IV secretion. Future Microbiol 7: 47–58. de Jong MF, Sun YH, den Hartigh AB, van Dijl JM & Tsolis RM (2008) Identification of VceA and VceC, two members of the VjbR regulon that are translocated into macrophages by the Brucella type IV secretion system. Mol Microbiol 70: 1378–1396. Delpino MV, Fossati CA & Baldi PC (2009) Proinflammatory response of human osteoblastic cell lines and osteoblastmonocyte interaction upon infection with Brucella spp. Infect Immun 77: 984–995. Delrue RM, Martinez-Lorenzo M, Lestrate P, Danese I, Bielarz V, Mertens P, De Bolle X, Tibor A, Gorvel JP & Letesson JJ (2001) Identification of Brucella spp. genes involved in intracellular trafficking. Cell Microbiol 3: 487–497. Delrue RM, Deschamps C, Leonard S et al. (2005) A quorumsensing regulator controls expression of both the type IV secretion system and the flagellar apparatus of Brucella melitensis. Cell Microbiol 7: 1151–1161. den Hartigh AB, Rolan HG, de Jong MF & Tsolis RM (2008) VirB3 to VirB6 and VirB8 to VirB11, but not VirB7, are essential for mediating persistence of Brucella in the reticuloendothelial system. J Bacteriol 190: 4427–4436. Detilleux PG, Deyoe BL & Cheville NF (1990) Entry and intracellular localization of Brucella spp. in Vero cells: fluorescence and electron microscopy. Vet Pathol 27: 317–328. Dorrell N, Spencer S, Foulonge V, Guigue-Talet P, O’Callaghan D & Wren BW (1998) Identification, cloning and initial characterisation of FeuPQ in Brucella suis: a new sub-family of two-component regulatory systems. FEMS Microbiol Lett 162: 143–150. Dozot M, Boigegrain RA, Delrue RM, Hallez R, OuahraniBettache S, Danese I, Letesson JJ, De Bolle X & Kohler S (2006) The stringent response mediator Rsh is required for Brucella melitensis and Brucella suis virulence, and for expression of the type IV secretion system virB. Cell Microbiol 8: 1791–1802. El-Tras WF, Tayel AA, Eltholth MM & Guitian J (2010) Brucella infection in fresh water fish: evidence for natural infection of Nile catfish, Clarias gariepinus, with Brucella melitensis. Vet Microbiol 141: 321–325. Elzer PH, Enright FM, McQuiston JR, Boyle SM & Schurig GG (1998) Evaluation of a rough mutant of Brucella melitensis in pregnant goats. Res Vet Sci 64: 259–260. Endley S, McMurray D & Ficht TA (2001) Interruption of the cydB locus in Brucella abortus attenuates intracellular survival and virulence in the mouse model of infection. J Bacteriol 183: 2454–2462. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. Eskra L, Mathison A & Splitter G (2003) Microarray analysis of mRNA levels from RAW264.7 macrophages infected with Brucella abortus. Infect Immun 71: 1125–1133. Eze MO, Yuan L, Crawford RM, Paranavitana CM, Hadfield TL, Bhattacharjee AK, Warren RL & Hoover DL (2000) Effects of opsonization and gamma interferon on growth of Brucella melitensis 16M in mouse peritoneal macrophages in vitro. Infect Immun 68: 257–263. Falkow S (1988) Molecular Koch’s postulates applied to microbial pathogenicity. Rev Infect Dis 10(suppl 2): S274– S276. Ferguson GP, Datta A, Baumgartner J, Roop RM II, Carlson RW & Walker GC (2004) Similarity to peroxisomalmembrane protein family reveals that Sinorhizobium and Brucella BacA affect lipid-A fatty acids. P Natl Acad Sci USA 101: 5012–5017. Fernandez-Prada CM, Zelazowska EB, Nikolich M, Hadfield TL, Roop RM II, Robertson GL & Hoover DL (2003) Interactions between Brucella melitensis and human phagocytes: bacterial surface O-Polysaccharide inhibits phagocytosis, bacterial killing, and subsequent host cell apoptosis. Infect Immun 71: 2110–2119. Ferrero MC, Fossati CA & Baldi PC (2009) Smooth Brucella strains invade and replicate in human lung epithelial cells without inducing cell death. Microbes Infect 11: 476–483. Fontes P, Alvarez-Martinez MT, Gross A, Carnaud C, Kohler S & Liautard JP (2005) Absence of evidence for the participation of the macrophage cellular prion protein in infection with Brucella suis. Infect Immun 73: 6229– 6236. Forestier C, Moreno E, Pizarro-Cerda J & Gorvel JP (1999) Lysosomal accumulation and recycling of lipopolysaccharide to the cell surface of murine macrophages, an in vitro and in vivo study. J Immunol 162: 6784–6791. Forestier C, Deleuil F, Lapaque N, Moreno E & Gorvel JP (2000) Brucella abortus lipopolysaccharide in murine peritoneal macrophages acts as a down-regulator of T cell activation. J Immunol 165: 5202–5210. Foster G, Osterman BS, Godfroid J, Jacques I & Cloeckaert A (2007) Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int J Syst Evol Microbiol 57: 2688– 2693. Foulongne V, Bourg G, Cazevieille C, Michaux-Charachon S & O’Callaghan D (2000) Identification of Brucella suis genes affecting intracellular survival in an in vitro human macrophage infection model by signature-tagged transposon mutagenesis. Infect Immun 68: 1297–1303. Foulongne V, Walravens K, Bourg G, Boschiroli ML, Godfroid J, Ramuz M & O’Callaghan D (2001) Aromatic compounddependent Brucella suis is attenuated in both cultured cells and mouse models. Infect Immun 69: 547–550. Franco MP, Mulder M & Smits HL (2007) Persistence and relapse in brucellosis and need for improved treatment. Trans R Soc Trop Med Hyg 101: 854–855. FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle Fretin D, Fauconnier A, Kohler S et al. (2005) The sheathed flagellum of Brucella melitensis is involved in persistence in a murine model of infection. Cell Microbiol 7: 687–698. Fugier E, Salcedo SP, de Chastellier C, Pophillat M, Muller A, Arce-Gorvel V, Fourquet P & Gorvel JP (2009) The glyceraldehyde-3-phosphate dehydrogenase and the small GTPase Rab 2 are crucial for Brucella replication. PLoS Pathog 5: e1000487. Galdiero E, Romano Carratelli C, Vitiello M, Nuzzo I, Del Vecchio E, Bentivoglio C, Perillo G & Galdiero F (2000) HSP and apoptosis in leukocytes from infected or vaccinated animals by Brucella abortus. New Microbiol 23: 271. Galindo RC, Munoz PM, de Miguel MJ, Marin CM, Labairu J, Revilla M, Blasco JM, Gortazar C & de la Fuente J (2010) Gene expression changes in spleens of the wildlife reservoir species, Eurasian wild boar (Sus scrofa), naturally infected with Brucella suis biovar 2. J Genet Genomics 37: 725–736. Garcia Samartino C, Delpino MV, Pott Godoy C et al. (2010) Brucella abortus induces the secretion of proinflammatory mediators from glial cells leading to astrocyte apoptosis. Am J Pathol 176: 1323–1338. Gauthier A, Thomas NA & Finlay BB (2003) Bacterial injection machines. J Biol Chem 278: 25273–25276. Gee JM, Valderas MW, Kovach ME, Grippe VK, Robertson GT, Ng WL, Richardson JM, Winkler ME & Roop RM II (2005) The Brucella abortus Cu,Zn superoxide dismutase is required for optimal resistance to oxidative killing by murine macrophages and wild-type virulence in experimentally infected mice. Infect Immun 73: 2873–2880. Godfroid F, Taminiau B, Danese I, Denoel P, Tibor A, Weynants V, Cloeckaert A, Godfroid J & Letesson JJ (1998) Identification of the perosamine synthetase gene of Brucella melitensis 16M and involvement of lipopolysaccharide O side chain in Brucella survival in mice and in macrophages. Infect Immun 66: 5485–5493. Gonzalez L, Patterson IA, Reid RJ et al. (2002) Chronic meningoencephalitis associated with Brucella sp. infection in live-stranded striped dolphins (Stenella coeruleoalba). J Comp Pathol 126: 147–152. Gonzalez D, Grillo MJ, De Miguel MJ et al. (2008) Brucellosis vaccines: assessment of Brucella melitensis lipopolysaccharide rough mutants defective in core and O-polysaccharide synthesis and export. PLoS ONE 3: e2760. Gorvel JP & de Chastellier C (2005) Bacteria spurned by selfabsorbed cells. Nat Med 11: 18–19. Gorvel JP & Moreno E (2002) Brucella intracellular life: from invasion to intracellular replication. Vet Microbiol 90: 281– 297. Gross A, Terraza A, Ouahrani-Bettache S, Liautard JP & Dornand J (2000) In vitro Brucella suis infection prevents the programmed cell death of human monocytic cells. Infect Immun 68: 342–351. Guzman-Verri C, Chaves-Olarte E, von Eichel-Streiber C, Lopez-Goni I, Thelestam M, Arvidson S, Gorvel JP & Moreno E (2001) GTPases of the Rho subfamily are FEMS Microbiol Rev 36 (2012) 533–562 557 required for Brucella abortus internalization in nonprofessional phagocytes: direct activation of Cdc42. J Biol Chem 276: 44435–44443. Guzman-Verri C, Manterola L, Sola-Landa A, Parra A, Cloeckaert A, Garin J, Gorvel JP, Moriyon I, Moreno E & Lopez-Goni I (2002) The two-component system BvrR/BvrS essential for Brucella abortus virulence regulates the expression of outer membrane proteins with counterparts in members of the Rhizobiaceae. P Natl Acad Sci USA 99: 12375–12380. Haag AF, Myka KK, Arnold MF, Caro-Hernandez P & Ferguson GP (2010) Importance of lipopolysaccharide and cyclic beta-1,2-glucans in Brucella-mammalian infections. Int J Microbiol 2010: 124509. Haine V, Sinon A, Van Steen F, Rousseau S, Dozot M, Lestrate P, Lambert C, Letesson JJ & De Bolle X (2005) Systematic targeted mutagenesis of Brucella melitensis 16M reveals a major role for GntR regulators in the control of virulence. Infect Immun 73: 5578–5586. Haine V, Dozot M, Dornand J, Letesson JJ & De Bolle X (2006) NnrA is required for full virulence and regulates several Brucella melitensis denitrification genes. J Bacteriol 188: 1615–1619. Hanna N, de Bagues MP, Ouahrani-Bettache S, El Yakhlifi Z, Kohler S & Occhialini A (2011) The virB operon is essential for lethality of Brucella microti in the Balb/c murine model of infection. J Infect Dis 203: 1129–1135. He Y, Reichow S, Ramamoorthy S, Ding X, Lathigra R, Craig JC, Sobral BW, Schurig GG, Sriranganathan N & Boyle SM (2006) Brucella melitensis triggers time-dependent modulation of apoptosis and down-regulation of mitochondrion-associated gene expression in mouse macrophages. Infect Immun 74: 5035–5046. Hernandez-Castro R, Verdugo-Rodriguez A, Puente JL & Suarez-Guemes F (2008) The BMEI0216 gene of Brucella melitensis is required for internalization in HeLa cells. Microb Pathog 44: 28–33. Hong PC, Tsolis RM & Ficht TA (2000) Identification of genes required for chronic persistence of Brucella abortus in mice. Infect Immun 68: 4102–4107. Iannino F, Ugalde JE & Inon de Iannino N (2012) Brucella abortus efp gene is required for an efficient internalization in HeLa cells. Microb Pathog 52: 31–40. Inon de Iannino N, Briones G, Tolmasky M & Ugalde RA (1998) Molecular cloning and characterization of cgs, the Brucella abortus cyclic beta(1-2) glucan synthetase gene: genetic complementation of Rhizobium meliloti ndvB and Agrobacterium tumefaciens chvB mutants. J Bacteriol 180: 4392–4400. Iriarte M, González D, Delrue RM, Monreal D, Conde R, López-Goñi I, Letesson J-J & Moriyón I (2004) Brucella lipopolysaccharide: structure, biosynthesis and genetics. Brucella: Molecular and Cellular Biology (López-Goñi I & Moriyón I, eds.), pp. 152–183. Horizon Bioscience, Norfolk. Iyankan L & Singh DK (2002) The effect of Brucella abortus on hydrogen peroxide and nitric oxide production by ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 558 bovine polymorphonuclear cells. Vet Res Commun 26: 93– 102. Jiang X & Baldwin CL (1993) Effects of cytokines on intracellular growth of Brucella abortus. Infect Immun 61: 124–134. Jimenez de Bagues MP, Ouahrani-Bettache S, Quintana JF et al. (2010) The new species Brucella microti replicates in macrophages and causes death in murine models of infection. J Infect Dis 202: 3–10. Kahl-McDonagh MM & Ficht TA (2006) Evaluation of protection afforded by Brucella abortus and Brucella melitensis unmarked deletion mutants exhibiting different rates of clearance in BALB/c mice. Infect Immun 74: 4048– 4057. Kahl-McDonagh MM, Elzer PH, Hagius SD et al. (2006) Evaluation of novel Brucella melitensis unmarked deletion mutants for safety and efficacy in the goat model of brucellosis. Vaccine 24: 5169–5177. Khan MY, Mah MW & Memish ZA (2001) Brucellosis in pregnant women. Clin Infect Dis 32: 1172–1177. Kim S, Watarai M, Kondo Y, Erdenebaatar J, Makino S & Shirahata T (2003) Isolation and characterization of miniTn5Km2 insertion mutants of Brucella abortus deficient in internalization and intracellular growth in HeLa cells. Infect Immun 71: 3020–3027. Kim S, Kurokawa D, Watanabe K, Makino S, Shirahata T & Watarai M (2004a) Brucella abortus nicotinamidase (PncA) contributes to its intracellular replication and infectivity in mice. FEMS Microbiol Lett 234: 289–295. Kim S, Watarai M, Suzuki H, Makino S, Kodama T & Shirahata T (2004b) Lipid raft microdomains mediate class A scavenger receptor-dependent infection of Brucella abortus. Microb Pathog 37: 11–19. Kim S, Watanabe K, Suzuki H & Watarai M (2005a) Roles of Brucella abortus SpoT in morphological differentiation and intramacrophagic replication. Microbiology 151: 1607–1617. Kim S, Lee DS, Watanabe K, Furuoka H, Suzuki H & Watarai M (2005b) Interferon-gamma promotes abortion due to Brucella infection in pregnant mice. BMC Microbiol 5: 22. Kohler S, Foulongne V, Ouahrani-Bettache S, Bourg G, Teyssier J, Ramuz M & Liautard JP (2002) The analysis of the intramacrophagic virulome of Brucella suis deciphers the environment encountered by the pathogen inside the macrophage host cell. P Natl Acad Sci USA 99: 15711– 15716. Lamkanfi M & Dixit VM (2010) Manipulation of host cell death pathways during microbial infections. Cell Host Microbe 8: 44–54. Lamontagne J, Butler H, Chaves-Olarte E et al. (2007) Extensive cell envelope modulation is associated with virulence in Brucella abortus. J Proteome Res 6: 1519–1529. Lapaque N, Takeuchi O, Corrales F, Akira S, Moriyon I, Howard JC & Gorvel JP (2006) Differential inductions of TNF-alpha and IGTP, IIGP by structurally diverse classic and non-classic lipopolysaccharides. Cell Microbiol 8: 401– 413. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. Lavigne JP, Patey G, Sangari FJ, Bourg G, Ramuz M, O’Callaghan D & Michaux-Charachon S (2005) Identification of a new virulence factor, BvfA, in Brucella suis. Infect Immun 73: 5524–5529. Lelouard H, Henri S, De Bovis B, Mugnier B, Chollat-Namy A, Malissen B, Meresse S & Gorvel JP (2010) Pathogenic bacteria and dead cells are internalized by a unique subset of Peyer’s patch dendritic cells that express lysozyme. Gastroenterology 138: 173–184, e171–173. Lelouard H, Fallet M, de Bovis B, Méresse S & Gorvel JP (2011) Peyer’s Patch denditric cells sample antigens by extending dendrites through M cell-specific transcellular pores. Gastroenterology 142: 592–601. Lestrate P, Delrue RM, Danese I, Didembourg C, Taminiau B, Mertens P, De Bolle X, Tibor A, Tang CM & Letesson JJ (2000) Identification and characterization of in vivo attenuated mutants of Brucella melitensis. Mol Microbiol 38: 543–551. Lestrate P, Dricot A, Delrue RM, Lambert C, Martinelli V, De Bolle X, Letesson JJ & Tibor A (2003) Attenuated signaturetagged mutagenesis mutants of Brucella melitensis identified during the acute phase of infection in mice. Infect Immun 71: 7053–7060. Lindgren SW, Stojiljkovic I & Heffron F (1996) Macrophage killing is an essential virulence mechanism of Salmonella typhimurium. P Natl Acad Sci USA 93: 4197–4201. Loisel-Meyer S, Jimenez de Bagues MP, Basseres E, Dornand J, Kohler S, Liautard JP & Jubier-Maurin V (2006) Requirement of norD for Brucella suis virulence in a murine model of in vitro and in vivo infection. Infect Immun 74: 1973–1976. Luhrmann A, Nogueira CV, Carey KL & Roy CR (2010) Inhibition of pathogen-induced apoptosis by a Coxiella burnetii type IV effector protein. P Natl Acad Sci USA 107: 18997–19001. Maddocks SE & Oyston PC (2008) Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 154: 3609–3623. Mancilla M, Lopez-Goni I, Moriyon I & Zarraga AM (2010) Genomic island 2 is an unstable genetic element contributing to Brucella lipopolysaccharide spontaneous smooth-to-rough dissociation. J Bacteriol 192: 6346–6351. Manterola L, Moriyon I, Moreno E, Sola-Landa A, Weiss DS, Koch MH, Howe J, Brandenburg K & Lopez-Goni I (2005) The lipopolysaccharide of Brucella abortus BvrS/BvrR mutants contains lipid A modifications and has higher affinity for bactericidal cationic peptides. J Bacteriol 187: 5631–5639. Maquart M, Zygmunt MS & Cloeckaert A (2009) Marine mammal Brucella isolates with different genomic characteristics display a differential response when infecting human macrophages in culture. Microbes Infect 11: 361–366. Marchesini MI, Herrmann CK, Salcedo SP, Gorvel JP & Comerci DJ (2011) In search of Brucella abortus type IV secretion substrates: screening and identification of four FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle proteins translocated into host cells through VirB system. Cell Microbiol 13: 1261–1274. Martinez-Nunez C, Altamirano-Silva P, Alvarado-Guillen F, Moreno E, Guzman-Verri C & Chaves-Olarte E (2010) The two-component system BvrR/BvrS regulates the expression of the type IV secretion system VirB in Brucella abortus. J Bacteriol 192: 5603–5608. Martin-Martin AI, Caro-Hernandez P, Orduna A, Vizcaino N & Fernandez-Lago L (2008) Importance of the Omp25/ Omp31 family in the internalization and intracellular replication of virulent B. ovis in murine macrophages and HeLa cells. Microbes Infect 10: 706–710. Martin-Martin AI, Caro-Hernandez P, Sancho P, Tejedor C, Cloeckaert A, Fernandez-Lago L & Vizcaino N (2009) Analysis of the occurrence and distribution of the Omp25/ Omp31 family of surface proteins in the six classical Brucella species. Vet Microbiol 137: 74–82. Martin-Martin Al, Vizcaino N & Fernandez-Lago L (2010) Cholesterol, galglioside GM1 and class A scavenger receptor contribute to infection by Brucella ovis and Brucella canis in murine macropgaes. Microbes Infect 12: 246–251. Martirosyan A, Moreno E & Gorvel JP (2011) An evolutionary strategy for a stealthy intracellular Brucella pathogen. Immunol Rev 240: 211–234. McDonald WL, Jamaludin R, Mackereth G et al. (2006) Characterization of a Brucella sp. strain as a marinemammal type despite isolation from a patient with spinal osteomyelitis in New Zealand. J Clin Microbiol 44: 4363– 4370. McKinney JD, Honer zu Bentrup K, Munoz-Elias EJ, Miczak A, Chen B, Chan WT, Swenson D, Sacchettini JC, Jacobs WR Jr & Russell DG (2000) Persistence of Mycobacterium tuberculosis in macrophages and mice requires the glyoxylate shunt enzyme isocitrate lyase. Nature 406: 735–738. McLean DR, Russell N & Khan MY (1992) Neurobrucellosis: clinical and therapeutic features. Clin Infect Dis 15: 582–590. McQuiston JR, Vemulapalli R, Inzana TJ et al. (1999) Genetic characterization of a Tn5-disrupted glycosyltransferase gene homolog in Brucella abortus and its effect on lipopolysaccharide composition and virulence. Infect Immun 67: 3830–3835. Meador VP & Deyoe BL (1989) Intracellular localization of Brucella abortus in bovine placenta. Vet Pathol 26: 513–515. Meador VP, Hagemoser WA & Deyoe BL (1988) Histopathologic findings in Brucella abortus-infected, pregnant goats. Am J Vet Res 49: 274–280. Memish ZA & Balkhy HH (2004) Brucellosis and international travel. J Travel Med 11: 49–55. Mizushima N (2004) Methods for monitoring autophagy. Int J Biochem Cell Biol 36: 2491–2502. Moreno E & Moriyon I (2006) The genus Brucella. The Prokaryotes, Vol. 5 (Dworkin M, Falkow S, Rosenberg E, Schleifer K & Stackebrandt E, eds), pp. 315–456. Springer, New York. Moreno E, Speth SL, Jones LM & Berman DT (1981) Immunochemical characterization of Brucella FEMS Microbiol Rev 36 (2012) 533–562 559 lipopolysaccharides and polysaccharides. Infect Immun 31: 214–222. Moreno E, Cloeckaert A & Moriyon I (2002) Brucella evolution and taxonomy. Vet Microbiol 90: 209–227. Naroeni A & Porte F (2002) Role of cholesterol and the ganglioside GM(1) in entry and short-term survival of Brucella suis in murine macrophages. Infect Immun 70: 1640–1644. Naroeni A, Jouy N, Ouahrani-Bettache S, Liautard JP & Porte F (2001) Brucella suis-impaired specific recognition of phagosomes by lysosomes due to phagosomal membrane modifications. Infect Immun 69: 486–493. Nymo IH, Tryland M & Godfroid J (2011) A review of Brucella infection in marine mammals, with special emphasis on Brucella pinnipedialis in the hooded seal (Cystophora cristata). Vet Res 42: 93. O’Callaghan D, Cazevieille C, Allardet-Servent A, Boschiroli ML, Bourg G, Foulongne V, Frutos P, Kulakov Y & Ramuz M (1999) A homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl type IV secretion systems is essential for intracellular survival of Brucella suis. Mol Microbiol 33: 1210–1220. Olsen SC (2010) Brucellosis in the United States: role and significance of wildlife reservoirs. Vaccine 28(suppl 5): F73– F76. Orduna A, Orduna C, Eiros JM, Bratos MA, Gutierrez P, Alonso P & Rodriguez Torres A (1991) Inhibition of the degranulation and myeloperoxidase activity of human polymorphonuclear neutrophils by Brucella melitensis. Microbiologia 7: 113–119. Paixao TA, Roux CM, den Hartigh AB, Sankaran-Walters S, Dandekar S, Santos RL & Tsolis RM (2009) Establishment of systemic Brucella melitensis infection through the digestive tract requires urease, the type IV secretion system, and lipopolysaccharide O antigen. Infect Immun 77: 4197– 4208. Pantoja M, Chen L, Chen Y & Nester EW (2002) Agrobacterium type IV secretion is a two-step process in which export substrates associate with the virulence protein VirJ in the periplasm. Mol Microbiol 45: 1325– 1335. Pappas G (2010) The changing Brucella ecology: novel reservoirs, new threats. Int J Antimicrob Agents 36(suppl 1): S8–S11. Parent MA, Goenka R, Murphy E, Levier K, Carreiro N, Golding B, Ferguson G, Roop RM II, Walker GC & Baldwin CL (2007) Brucella abortus bacA mutant induces greater pro-inflammatory cytokines than the wild-type parent strain. Microbes Infect 9: 55–62. Pei J & Ficht TA (2004) Brucella abortus rough mutants are cytopathic for macrophages in culture. Infect Immun 72: 440–450. Pei J, Turse JE, Wu Q & Ficht TA (2006) Brucella abortus rough mutants induce macrophage oncosis that requires bacterial protein synthesis and direct interaction with the macrophage. Infect Immun 74: 2667–2675. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 560 Pei J, Turse JE & Ficht TA (2008a) Evidence of Brucella abortus OPS dictating uptake and restricting NF-kappaB activation in murine macrophages. Microbes Infect 10: 582–590. Pei J, Wu Q, Kahl-McDonagh M & Ficht TA (2008b) Cytotoxicity in macrophages infected with rough Brucella mutants is type IV secretion system dependent. Infect Immun 76: 30–37. Pizarro-Cerda J, Moreno E, Sanguedolce V, Mege JL & Gorvel JP (1998a) Virulent Brucella abortus prevents lysosome fusion and is distributed within autophagosome-like compartments. Infect Immun 66: 2387–2392. Pizarro-Cerda J, Meresse S, Parton RG, van der Goot G, SolaLanda A, Lopez-Goni I, Moreno E & Gorvel JP (1998b) Brucella abortus transits through the autophagic pathway and replicates in the endoplasmic reticulum of nonprofessional phagocytes. Infect Immun 66: 5711–5724. Porte F, Liautard JP & Kohler S (1999) Early acidification of phagosomes containing Brucella suis is essential for intracellular survival in murine macrophages. Infect Immun 67: 4041–4047. Porte F, Naroeni A, Ouahrani-Bettache S & Liautard JP (2003) Role of the Brucella suis lipopolysaccharide O antigen in phagosomal genesis and in inhibition of phagosomelysosome fusion in murine macrophages. Infect Immun 71: 1481–1490. Qin QM, Pei J, Ancona V, Shaw BD, Ficht TA & de Figueiredo P (2008) RNAi screen of endoplasmic reticulumassociated host factors reveals a role for IRE1alpha in supporting Brucella replication. PLoS Pathog 4: e1000110. Radhakrishnan GK, Yu Q, Harms JS & Splitter GA (2009) Brucella TIR domain-containing protein mimics properties of the Toll-like receptor adaptor protein TIRAP. J Biol Chem 284: 9892–9898. Radhakrishnan GK, Harms JS & Splitter GA (2011) Modulation of microtubule dynamics by a TIR domain protein from the intracellular pathogen Brucella melitensis. Biochem J 439: 79–83. Rajashekara G, Glover DA, Krepps M & Splitter GA (2005) Temporal analysis of pathogenic events in virulent and avirulent Brucella melitensis infections. Cell Microbiol 7: 1459–1473. Rajashekara G, Glover DA, Banai M, O’Callaghan D & Splitter GA (2006) Attenuated bioluminescent Brucella melitensis mutants GR019 (virB4), GR024 (galE), and GR026 (BMEI1090-BMEI1091) confer protection in mice. Infect Immun 74: 2925–2936. Rambow-Larsen AA, Rajashekara G, Petersen E & Splitter G (2008) Putative quorum-sensing regulator BlxR of Brucella melitensis regulates virulence factors including the type IV secretion system and flagella. J Bacteriol 190: 3274–3282. Rambow-Larsen AA, Petersen EM, Gourley CR & Splitter GA (2009) Brucella regulators: self-control in a hostile environment. Trends Microbiol 17: 371–377. Riley LK & Robertson DC (1984) Ingestion and intracellular survival of Brucella abortus in human and bovine polymorphonuclear leukocytes. Infect Immun 46: 224–230. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. Rittig MG, Kaufmann A, Robins A, Shaw B, Sprenger H, Gemsa D, Foulongne V, Rouot B & Dornand J (2003) Smooth and rough lipopolysaccharide phenotypes of Brucella induce different intracellular trafficking and cytokine/chemokine release in human monocytes. J Leukoc Biol 74: 1045–1055. Robertson GT & Roop RM Jr (1999) The Brucella abortus host factor I (HF-I) protein contributes to stress resistance during stationary phase and is a major determinant of virulence in mice. Mol Microbiol 34: 690–700. Rolan HG & Tsolis RM (2007) Mice lacking components of adaptive immunity show increased Brucella abortus virB mutant colonization. Infect Immun 75: 2965–2973. Rolan HG, Xavier MN, Santos RL & Tsolis RM (2009) Natural antibody contributes to host defense against an attenuated Brucella abortus virB mutant. Infect Immun 77: 3004–3013. Roop RM II, Jeffers G, Bagchi T, Walker J, Enright FM & Schurig GG (1991) Experimental infection of goat fetuses in utero with a stable, rough mutant of Brucella abortus. Res Vet Sci 51: 123–127. Roop RM II, Robertson GT, Ferguson GP, Milford LE, Winkler ME & Walker GC (2002) Seeking a niche: putative contributions of the hfq and bacA gene products to the successful adaptation of the brucellae to their intracellular home. Vet Microbiol 90: 349–363. Roop RM II, Gaines JM, Anderson ES, Caswell CC & Martin DW (2009) Survival of the fittest: how Brucella strains adapt to their intracellular niche in the host. Med Microbiol Immunol 198: 221–238. Rossetti CA, Galindo CL, Garner HR & Adams LG (2010) Selective amplification of Brucella melitensis mRNA from a mixed host-pathogen total RNA. BMC Res Notes 3: 244. Rouot B, Alvarez-Martinez MT, Marius C, Menanteau P, Guilloteau L, Boigegrain RA, Zumbihl R, O’Callaghan D, Domke N & Baron C (2003) Production of the type IV secretion system differs among Brucella species as revealed with VirB5- and VirB8-specific antisera. Infect Immun 71: 1075–1082. Roux CM, Rolan HG, Santos RL, Beremand PD, Thomas TL, Adams LG & Tsolis RM (2007) Brucella requires a functional Type IV secretion system to elicit innate immune responses in mice. Cell Microbiol 9: 1851–1869. Salcedo SP, Marchesini MI, Lelouard H et al. (2008) Brucella control of dendritic cell maturation is dependent on the TIR-containing protein Btp1. PLoS Pathog 4: e21. Samartino LE, Traux RE & Enright FM (1994) Invasion and replication of Brucella abortus in three different trophoblastic cell lines. Zentralbl Veterinarmed B 41: 229–236. Sathiyaseelan J, Jiang X & Baldwin CL (2000) Growth of Brucella abortus in macrophages from resistant and susceptible mouse strains. Clin Exp Immunol 121: 289–294. Scholz HC, Hubalek Z, Nesvadbova J et al. (2008a) Isolation of Brucella microti from soil. Emerg Infect Dis 14: 1316–1317. Scholz HC, Hubalek Z, Sedlacek I et al. (2008b) Brucella microti sp. nov., isolated from the common vole Microtus arvalis. Int J Syst Evol Microbiol 58: 375–382. FEMS Microbiol Rev 36 (2012) 533–562 Brucella intracellular lifestyle Scholz HC, Nockler K, Gollner C et al. (2010) Brucella inopinata sp. nov., isolated from a breast implant infection. Int J Syst Evol Microbiol 60: 801–808. Scian R, Barrionuevo P, Giambartolomei GH, Fossati CA, Baldi PC & Delpino MV (2011) Granulocyte-macrophage colony-stimulating factor- and tumor necrosis factor alphamediated matrix metalloproteinase production by human osteoblasts and monocytes after infection with Brucella abortus. Infect Immun 79: 192–202. Scurlock BM & Edwards WH (2010) Status of brucellosis in free-ranging elk and bison in Wyoming. J Wildl Dis 46: 442–449. Seleem MN, Boyle SM & Sriranganathan N (2008) Brucella: a pathogen without classic virulence genes. Vet Microbiol 129: 1–14. Sengupta D, Koblansky A, Gaines J, Brown T, West AP, Zhang D, Nishikawa T, Park SG, Roop RM II & Ghosh S (2010) Subversion of innate immune responses by Brucella through the targeted degradation of the TLR signaling adapter, MAL. J Immunol 184: 956–964. Sieira R, Comerci DJ, Sanchez DO & Ugalde RA (2000) A homologue of an operon required for DNA transfer in Agrobacterium is required in Brucella abortus for virulence and intracellular multiplication. J Bacteriol 182: 4849–4855. Sieira R, Comerci DJ, Pietrasanta LI & Ugalde RA (2004) Integration host factor is involved in transcriptional regulation of the Brucella abortus virB operon. Mol Microbiol 54: 808–822. Sieira R, Arocena GM, Bukata L, Comerci DJ & Ugalde RA (2010) Metabolic control of virulence genes in Brucella abortus: HutC coordinates virB expression and the histidine utilization pathway by direct binding to both promoters. J Bacteriol 192: 217–224. Silva TM, Costa EA, Paixao TA, Tsolis RM & Santos RL (2011) Laboratory animal models for brucellosis research. J Biomed Biotechnol 2011: 518323. Sohn AH, Probert WS, Glaser CA, Gupta N, Bollen AW, Wong JD, Grace EM & McDonald WC (2003) Human neurobrucellosis with intracerebral granuloma caused by a marine mammal Brucella spp. Emerg Infect Dis 9: 485–488. Sola-Landa A, Pizarro-Cerda J, Grillo MJ, Moreno E, Moriyon I, Blasco JM, Gorvel JP & Lopez-Goni I (1998) A twocomponent regulatory system playing a critical role in plant pathogens and endosymbionts is present in Brucella abortus and controls cell invasion and virulence. Mol Microbiol 29: 125–138. Spera JM, Ugalde JE, Mucci J, Comerci DJ & Ugalde RA (2006) A B lymphocyte mitogen is a Brucella abortus virulence factor required for persistent infection. P Natl Acad Sci USA 103: 16514–16519. Starr T, Ng TW, Wehrly TD, Knodler LA & Celli J (2008) Brucella intracellular replication requires trafficking through the late endosomal/lysosomal compartment. Traffic 9: 678– 694. Steele KH, Baumgartner JE, Valderas MW & Roop RM II (2010) Comparative study of the roles of AhpC and KatE as FEMS Microbiol Rev 36 (2012) 533–562 561 respiratory antioxidants in Brucella abortus 2308. J Bacteriol 192: 4912–4922. Swartz TE, Tseng TS, Frederickson MA et al. (2007) Bluelight-activated histidine kinases: two-component sensors in bacteria. Science 317: 1090–1093. Taminiau B, Daykin M, Swift S, Boschiroli ML, Tibor A, Lestrate P, De Bolle X, O’Callaghan D, Williams P & Letesson JJ (2002) Identification of a quorum-sensing signal molecule in the facultative intracellular pathogen Brucella melitensis. Infect Immun 70: 3004–3011. Tatum FM, Detilleux PG, Sacks JM & Halling SM (1992) Construction of Cu-Zn superoxide dismutase deletion mutants of Brucella abortus: analysis of survival in vitro in epithelial and phagocytic cells and in vivo in mice. Infect Immun 60: 2863–2869. Tatum FM, Morfitt DC & Halling SM (1993) Construction of a Brucella abortus RecA mutant and its survival in mice. Microb Pathog 14: 177–185. Tiller RV, Gee JE, Frace MA, Taylor TK, Setubal JC, Hoffmaster AR & De BK (2010a) Characterization of novel Brucella strains originating from wild native rodent species in North Queensland, Australia. Appl Environ Microbiol 76: 5837–5845. Tiller RV, Gee JE, Lonsway DR, Gribble S, Bell SC, Jennison AV, Bates J, Coulter C, Hoffmaster AR & De BK (2010b) Identification of an unusual Brucella strain (BO2) from a lung biopsy in a 52 year-old patient with chronic destructive pneumonia. BMC Microbiol 10: 23. Tisdale EJ, Kelly C & Artalejo CR (2004) Glyceraldehyde-3phosphate dehydrogenase interacts with Rab2 and plays an essential role in endoplasmic reticulum to Golgi transport exclusive of its glycolytic activity. J Biol Chem 279: 54046– 54052. Tobias L, Cordes DO & Schurig GG (1993) Placental pathology of the pregnant mouse inoculated with Brucella abortus strain 2308. Vet Pathol 30: 119–129. Tolomeo M, Di Carlo P, Abbadessa V, Titone L, Miceli S, Barbusca E, Cannizzo G, Mancuso S, Arista S & Scarlata F (2003) Monocyte and lymphocyte apoptosis resistance in acute and chronic brucellosis and its possible implications in clinical management. Clin Infect Dis 36: 1533–1538. Turse JE, Pei J & Ficht TA (2011) Lipopolysaccharide-deficient Brucella variants arise spontaneously during infection. Front Microbiol 2: 54. Ugalde RA (1999) Intracellular lifestyle of Brucella spp. Common genes with other animal pathogens, plant pathogens, and endosymbionts. Microbes Infect 1: 1211– 1219. Ugalde JE, Czibener C, Feldman MF & Ugalde RA (2000) Identification and characterization of the Brucella abortus phosphoglucomutase gene: role of lipopolysaccharide in virulence and intracellular multiplication. Infect Immun 68: 5716–5723. Uzureau S, Godefroid M, Deschamps C, Lemaire J, De Bolle X & Letesson JJ (2007) Mutations of the quorum sensingdependent regulator VjbR lead to drastic surface ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved 562 modifications in Brucella melitensis. J Bacteriol 189: 6035– 6047. Viadas C, Rodriguez MC, Sangari FJ, Gorvel JP, Garcia-Lobo JM & Lopez-Goni I (2010) Transcriptome analysis of the Brucella abortus BvrR/BvrS two-component regulatory system. PLoS ONE 5: e10216. Wang Y, Chen Z, Qiao F et al. (2009) Comparative proteomics analyses reveal the virB of B. melitensis affects expression of intracellular survival related proteins. PLoS ONE 4: e5368. Wang Y, Chen Z, Qiao F et al. (2010) The type IV secretion system affects the expression of Omp25/Omp31 and the outer membrane properties of Brucella melitensis. FEMS Microbiol Lett 303: 92–100. Wassenaar TM & Gaastra W (2001) Bacterial virulence: can we draw the line? FEMS Microbiol Lett 201: 1–7. Watanabe K, Tachibana M, Kim S & Watarai M (2009) Participation of ezrin in bacterial uptake by trophoblast giant cells. Reprod Biol Endocrinol 7: 95. Watarai M, Makino S, Fujii Y, Okamoto K & Shirahata T (2002) Modulation of Brucella-induced macropinocytosis by lipid rafts mediates intracellular replication. Cell Microbiol 4: 341–355. Watarai M, Kim S, Erdenebaatar J, Makino S, Horiuchi M, Shirahata T, Sakaguchi S & Katamine S (2003) Cellular prion protein promotes Brucella infection into macrophages. J Exp Med 198: 5–17. Weeks JN, Galindo CL, Drake KL, Adams GL, Garner HR & Ficht TA (2010) Brucella melitensis VjbR and C12-HSL regulons: contributions of the N-dodecanoyl homoserine lactone signaling molecule and LuxR homologue VjbR to gene expression. BMC Microbiol 10: 167. Wells DH & Long SR (2003) Mutations in rpoBC suppress the defects of a Sinorhizobium meliloti relA mutant. J Bacteriol 185: 5602–5610. West NP, Sansonetti P, Mounier J et al. (2005) Optimization of virulence functions through glucosylation of Shigella LPS. Science 307: 1313–1317. ª 2012 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved K. von Bargen et al. Whatmore AM, Dawson CE, Groussaud P, Koylass MS, King AC, Shankster SJ, Sohn AH, Probert WS & McDonald WL (2008) Marine mammal Brucella genotype associated with zoonotic infection. Emerg Infect Dis 14: 517–518. Winter AJ, Schurig GG, Boyle SM, Sriranganathan N, Bevins JS, Enright FM, Elzer PH & Kopec JD (1996) Protection of BALB/c mice against homologous and heterologous species of Brucella by rough strain vaccines derived from Brucella melitensis and Brucella suis biovar 4. Am J Vet Res 57: 677– 683. World Health Organization (2005) The Control of Neglected Zoonotic Diseases. World Health Organization, Geneva. Wu Q, Pei J, Turse C & Ficht TA (2006) Mariner mutagenesis of Brucella melitensis reveals genes with previously uncharacterized roles in virulence and survival. BMC Microbiol 6: 102. Yeoman KH, Delgado MJ, Wexler M, Downie JA & Johnston AW (1997) High affinity iron acquisition in Rhizobium leguminosarum requires the cycHJKL operon and the feuPQ gene products, which belong to the family of twocomponent transcriptional regulators. Microbiology 143(Pt 1): 127–134. Zhong Z, Wang Y, Qiao F et al. (2009) Cytotoxicity of Brucella smooth strains for macrophages is mediated by increased secretion of the type IV secretion system. Microbiology 155: 3392–3402. Zhou Y & Xie J (2011) The roles of pathogen small RNAs. J Cell Physiol 226: 968–973. Zwerdling A, Delpino MV, Barrionuevo P, Cassataro J, Pasquevich KA, Garcia Samartino C, Fossati CA & Giambartolomei GH (2008) Brucella lipoproteins mimic dendritic cell maturation induced by Brucella abortus. Microbes Infect 10: 1346–1354. Zygmunt MS, Hagius SD, Walker JV & Elzer PH (2006) Identification of Brucella melitensis 16M genes required for bacterial survival in the caprine host. Microbes Infect 8: 2849–2854. FEMS Microbiol Rev 36 (2012) 533–562