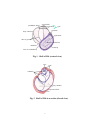
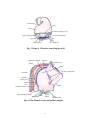
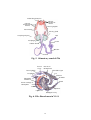
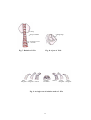
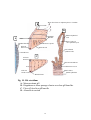
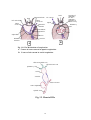
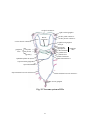
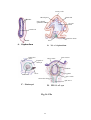
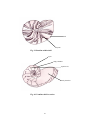
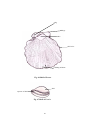
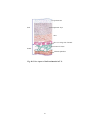
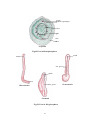
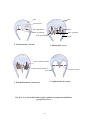
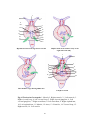
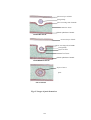
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Animal Diversity: (Non-Chordates) Phylum : Mollusca Ranjana Saxena Associate Professor, Department of Zoology, Dyal Singh College, University of Delhi Delhi e-mail: [email protected] 24th September 2007 CONTENT 1. GENERAL CHARACTERISTICS 2. PILA GLOBOSA a) Habit and Habitat b) Morphology c) Coelom d) Locomotion e) Digestive System f) Respiratory system g) Circulatory System h) Excretory System i) Nervous System j) Sense organs k) Reproductive System 3. SEPIA a) Habit and Habitat b) Morphology c) Shell d) Coelom e) Locomotion f) Digestive System g) Respiratory System h) Circulatory System i) Excretory System j) Nervous System k) Sense Organs l) Reproductive System 4. ANCESTRAL MOLLUSK 5. SHELL IN MOLLUSCA 6. FOOT AND ITS MODIFICATION 2 7. GILLS AND ITS MODIFICATION 8. MANTLE 9. TORSION IN MOLLUSCA 10. PEARL FORMATION 11. CLASSIFICATION 12. BIBLIOGRAPHY 13. SUGGESTED READING 3 PHYLUM MOLLUSCA The word Mollusca is derived from the latin word mollis which means soft bodied. GENERAL CHARACTERISTICS • It is the second largest phylum of invertebrates consisting of more than 80,000 living species and about 35,000 fossil species. • The adults are triploblastic, bilaterally symmetrical animals with a soft unsegmented body. However, the bilateral symmetry may be lost in some adult mollusc. Majority of them are enclosed in a calcareous shell. The shell may be external or in a few molluscs it may be internal, reduced or absent. They have a well marked cephalisation. The body is divisible into head, mantle, foot and visceral mass. The visceral mass is enclosed in a thick muscular fold of the body wall called mantle which secretes the shell. Body cavity is a haemocoel. True coelom is schizocoelic and reduced to cavities of pericardium, gonads and kidneys. The buccal cavity contains a rasping organ called radula. It helps the animal in feeding. In some of the molluscs, however, it is secondarily lost. A ventral muscular foot is an adaptation for locomotion, primarily for crawling. In some, it is secondarily modified for swimming or burrowing. Paired gills or ctenidia are the primary aquatic respiratory organs. Some of the molluscs may possess pulmonary sac for breathing atmospheric oxygen. Blood vascular system is of the open type with dorsal heart and a few blood vessels. However cephalopods show a tendency towards a closed system. Respiratory pigment is haemocyanin. Excretion is by a pair of kidneys or nephridia or organ of Bojanus. An olfactory organ, the osphradia is situated at the base of the gills. The osphradia tests the purity of water that enters the mantle cavity. Majority of them are dioceous but a few of them may be hermaphrodite. Development is generally indirect having larval stages but direct development is also found in some molluscs. Trocophore larva, veliger larva and glochidium larva are the characteristic larvae of mollusc that have an indirect development. • • • • • • • • • • • • • • Molluscs may be terrestrial or aquatic (freshwater or marine). They may be found in all habitats, deserts, forests, lakes, rivers, abysses of sea, coral reefs, underground or even as parasite in the body of other animals. They may be found clinging to the rocks, crawling, swimming, burrowing or even digging. They have a diversified feeding habit, may be herbivores, carnivores or omnivores. They vary in size from giant squids (nearly 2m long) to little snails, a millimeter long. Molluscan shell have always been economically important, having served as money in early days. They have been used in jewellery and buttons. Scallops, oysters, squids and octopus are important food items. Since phylum mollusca has wide range of structural diversities two animals namely pila and sepia will be described in details for a better understanding of the phylum. 4 PILA GLOBOSA Commonly known as apple snail or pond snail. HABIT AND HABITAT: Pila globosa is amongst the largest fresh water molluscs that have invaded various kinds of habitats. They live in fresh water pools, ponds, lakes and paddy fields where succulent water plants are in abundant. Sometimes they may be found in running streams also. They are herbivores and aquatic plants like Vallisnaria and Pistia constitutes its food. Pila is a voracious feeder and feeds on plant scrapings which are cut with the help of chitinous jaws and radular teeth. The snail creeps slowly with the help of ventral muscular foot. While feeding or moving, the animal does not leave the shell completely but carries it on its back. Whenever it senses danger, it retreats into its shell and closes the shell aperture with the operculum. When the snail is in a quiescent state, it retracts its body into the shell. But when active, the head and foot protrudes out of the shell and the animal creeps slowly. During unfavourable conditions, when the pond dries up as is seen in summers, it completely retracts their body within the shell and remain torpid. This period of inactivity is called aestivation or summer sleep. They can live for years in this condition. When the conditions are favourable, like in rainy season they once again become active. They are amphibious i.e. can respire both in water and on land. In water they respire with the help of ctenidia while on land they respire by pulmonary sac. MORPHOLOGY: The soft body of the animal is enclosed in a thick, calcareous shell composed of a single piece and hence the shell is called a univalve shell. The shell has a conical structure, spirally coiled around a central axis called the columella. The columella is hollow and its cavity opens to the outside by umbilicus. A shell with an umbilicus is called as umbilicate or perforate shell. Each revolution around the axis is called a whorl. The small rounded tip of the shell is called the apex and the whorl surrounding it is called the apical whorl (Fig. 1). The apex of the shell is the smallest and is regarded as the oldest part of the shell. The lower most whorl is the largest and is known as the body whorl. The whorl above the body whorl is called the penultimate whorl. All the whorls except the body whorl together are called spire (Fig. 2). Internally, all the whorls freely communicate with one another and there is no separating partition between them and thus the shell is known as unilocular. Externally, there is a line at the junction of the two successive whorls which is known as suture. The penultimate whorl and the body whorl are large enough to enclose most of the body parts. The outer surface of the shell is marked with numerous fine vertical lines called lines of growth. In addition to these lines of growth, there are few prominent deep vertical lines along the entire length of the shell. These are called varices (singular varix) and indicate fresh growth after every aestivation or hibernation. The body whorl opens to the outside by a wide aperture or mouth, the outer margin of which is called the outer lip while the inner one is called the columellar lip. The margin of the aperture is smooth and rolled out and is known as peristome. The aperture can be closed by a flat calcareous plate called the operculum (Fig. 1), which is attached to the dorsal side of the hind part of the foot. The operculum is secreted by the glandular cells in the foot. A distinct elliptical area is seen on the inner surface of the operculum and is called the boss. Opercular muscles are attached to the boss. The operculum shows many concentric rings of growth arranged around a small central axis called the nucleus. When the foot is withdrawn into the shell, the operculum fits into the mouth of the shell and closes it. 5 The whole body is enclosed within the whorls of the shell and is attached to the columella by columellar muscles. The columellar muscles arise from the foot and prevent the animal from extending out of the shell beyond a certain limit and also helps to withdraw the body into the shell. Beneath the shell is the soft body made up of head, foot, visceral mass and mantle or pallium (Fig.3). Head and foot together form the head-foot complex which retains the bilateral symmetry. The visceral mass and the pallium together form the visceropallium which has secondarily lost the bilateral symmetry. HEAD: The head is the muscular part of the body which hangs over the foot and projects out as a retractile snout. Head bears two pairs of contractile tentacles arising from behind the snout from the dorsal surface of the head.. The anterior pair of tentacles are called the cephalic tentacles or labial palps and are prolongations of the sides of the head. The posterior pair of tentacles are long, fleshy and contractile and are the true tentacles. The tentacles are hollow and are capable of extension and retraction. Eye is present on a small club shaped stalk called ommatophore just behind the true tentacles (Fig. 3 and 4). Mouth is a vertical slit like aperture lying ventrally between the bases of the cephalic tentacles. FOOT: A large more or less conical foot adapted for creeping movement is present below and behind the head. The anterior part of the foot is round and the posterior dorsal surface bears the operculum (Fig. 3). The foot is highly muscular and contains pedal glands, the secretions of which form a slime trail during locomotion. The head and foot together constitute the head-foot complex. The two are connected to the visceral mass through a slight constriction. 6 penultimate whorl apex of shell spire sutures peristome body- whorl operculum lines of growth nucleus umbilicus outer lip inner or columellar lip Fig. 1. Shell of Pila (ventral view) apex spire columella body chamber aperture of shell Fig. 2. Shell of Pila in a section (dorsal view) 7 visceral mass head mantle supra marginal groove right pseudopipodium left pseudopipodium operculum eye foot first tentacle second tentacle Fig. 3. Body of Pila after removing the shell pulmonary chamber intestine pulmonary sac mantle vagina opening of pulmonary sac ctenidium epitaenia vaginal pore rectum anus branchial chamber mantle eye Right pseudepipodium head 1st tentacle mouth 2nd true tentacle Fig. 4. Pila: Mantle cavity and pallial complex 8 VISCERAL MASS: It lies above the head-foot complex and contains the main organs of the body. It is spirally coiled like the shell and fills all the whorls of the shell. The visceral mass undergoes torsion (will be discussed later). The visceral mass is soft and is covered by a delicate mantle or pallium (Fig. 3). When the animal is in a retracted state, the mantle covers not only the visceral mass but also the head and its appendages. The edge of the mantle is thick and contains shell glands which secrete the shell. At the sides of the head and projecting above the foot, the mantle bears two fleshy lobes called nuchal lobes or pseudepipodia (Fig. 3). The left pseudepipodium is large and can be extended to form a long tubular respiratory siphon for aerial respiration. The right pseudepipodium is less developed. The mantle is free anteriorly and encloses a spacious cavity called the mantle cavity or the pallial cavity. The mantle cavity contains some of the important organs of the animal and together constitute the pallial complex. The mantle cavity is imperfectly divided into smaller right branchial chamber and larger left pulmonary chamber by a longitudinal ridge known as epitaenium. The right chamber is called the branchial chamber and contains the ctenidium, rectum and the genital duct while the left chamber called the pulmonary chamber contains the pulmonary sac (Fig.4). Pulmonary sac is a bag-like organ which hangs downwards from the roof of the mantle cavity and communicates with the pulmonary chamber by an elongated opening. A comb- shaped structure called the osphradium is attached to the roof of the mantle cavity close to the left nuchal lobe, on the left side of the pulmonary chamber. It is olfactory in function and tests the purity of water current entering the mantle cavity. The ctenidium is attached to the branchial chamber on the right angle. The rectum is present on the left of the ctenidium on the floor of the branchial chamber and opens to the outside by the anus which is situated just near the base of the right nuchal lobe. Both mouth and anus are situated on the anterior side of the body. Anterior renal chamber projects as a reddish sac in the branchial chamber near the posterior end of the epitaenia and opens into the mantle cavity by an oblique slit. The genital duct is present on the left of the rectum and the genital aperture is present very close to the anus (Fig.4). FUNCTIONS OF MANTLE: 1. Protects the visceral mass and head. 2. Serves as an additional respiratory organ. 3. Secretes the shell. COELOM: The coelom is poorly developed in adults and is represented by the pericardial cavity and the cavities around the kidney and gonads. The general body cavity is a haemocoel. LOCOMOTION: Pila moves slowly by creeping over the substratum with the help of its foot. While creeping, the foot is protruded out of the shell by pumping the blood into its sinus. The pedal mucus glands in the foot produces slimy secretion that enables the animal to glide on the dry surface. The foot is also provided with longitudinal, vertical and transverse muscles. The main power for locomotion is provided by waves of contraction passing from the anterior to posterior end of the foot. The contraction of the transverse muscles drive the blood forward which causes the extension of the foot. The contraction of the vertical muscles raise the anterior end and allows it to move forward while the posterior end is pulled forward by the contraction of the longitudinal muscles. 9 DIGESTIVE SYSTEM: The digestive system consists of a long curved tube extending from the mouth to the anus. It is broadly differentiated into foregut, midgut and hindgut (Fig. 5). FORE GUT: The fore gut is ectodermal in origin and consists of mouth, buccal mass and oesophagus. Mouth is a median vertical slit situated at the anterior end of the snout that leads into a thick walled muscular buccal mass. The buccal mass encloses a cavity known as buccal cavity which contains two jaws and a radula. Projecting from the roof of the buccal cavity, at the entrance of the mouth is a pair of chitinous jaws (Fig. 8). Both jaws are connected together by a thin cuticular membrane. The jaws bear numerous small and two or three large teeth. The jaw divides the buccal cavity into an anterior smaller cavity called the vestibule and a posterior larger buccal cavity. Present on the floor of the buccal cavity is a rounded elevation called the odontophore or tongue mass (Fig.6). It divides the buccal cavity into anterior subradular cavity and posterior radular sac. The odontophore is made up of muscles with cartilaginous support. The anterior end of the odontophore is round and is known as subradular organ. Beneath the sub-radular organ is a small sac like part called the sub-lingual cavity. Jaws work against the sub-radular organ to cut the food into smaller bits. Along the upper surface of odontophore there is a chitinous ribbon shaped organ called radula. Posteriorly, it is enclosed in a sac called the radular sac that extends below the buccal mass (Fig. 6). The radula bears numerous backwardly directed horny teeth arranged in transverse rows (Fig. 7). In Pila each transverse row possesses seven teeth, a median rachidian teeth, on its either side one lateral teeth and two marginal teeth, giving the formula 2, 1, 1, 1, 2 (Fig. 9). The teeth of the radula are secreted by special cells called odontoblasts which are arranged in transverse rows at the posterior end of the radular sac. The teeth formed in the posterior part keep on moving forward replacing the broken and worn out teeth of the anterior part. Thus, the anterior most teeth are the oldest and the most worn out. The radula grows at the rate of one to five rows of teeth per day. The teeth are pointed with sharp edges and are used like a saw for cutting the food. The radula is moved forward and backward, up and down with the help of protractor and retractor muscles and act as a rasping organ. A sub-radular membrane is present below the radula. A pair of salivary glands are present on the sides of the buccal mass and their ducts open into the buccal cavity. The secretions of the salivary glands contain mucus and an enzyme which digests starch. The mucus lubricates the radula and helps in the transport of food. Dorsally, the buccal mass leads into a long narrow oesophagus. Just below the salivary glands, near the origin of the oesophagus are a pair of round oesophageal pouches. They are prolongations of the oesophagus and probably secrete digestive enzymes. Digestion begins in the oesophageal pouches, and they also serve as site for the temporary storage of food. A pair of buccal glands are present in the roof of the buccal cavity, a little in front of its junction with the oesophagus. They are probably accessory digestive glands. MID GUT: Mid gut is endodermal in origin and consists of stomach and intestine. Oesophagus opens into the stomach which lies on the left side of the visceral mass just below the pericardium. It is red in color. The stomach consists of two parts: 1. Saccular cardiac chamber and 2. Tubular pyloric chamber. The oesophagus opens into the cardiac chamber. The cardiac chamber constitutes the main part of the stomach and possesses longitudinal folds on its inner surface while the pyloric chamber has transverse folds on its inner surface. The pyloric 10 chamber lies parallel to the oesophagus and opens into the intestine. Intestine turns backward and lies as a coiled tube in the visceral mass. The intestine is long and forms 2 ½-3 coils. A short, round blind pouch called the caecum arises from the pyloric chamber and opens at the junction of the stomach and the intestine. Lying in the visceral mass is the digestive gland (often referred to as the hepatopancreas or the liver) which is dirty brown in color (Fig. 5). It has two lobes; one smaller in close contact with the stomach, and the other larger extending upto the apex of the spiral. A large duct comes out of each lobe and the two ducts unite into a common duct before opening into the stomach by a common aperture. The digestive gland contains three types of cells: 1. SECRETORY CELLS: They secrete cellulose digesting enzymes. 2. RESORPTIVE CELLS: They digest proteins intracellularly. 3. LIME CELLS: They store calcium phosphate. HIND GUT: The posterior part of the intestine is straight and thick walled and is called the rectum. It lies on the floor of the right side of the mantle cavity and terminates in an anus which is situated near the mouth on the right side of the head (Fig. 5). The hindgut is ectodermal in origin. 11 medial dorsal muscles anterior dorsal-lateral muscle posterior dorsal-lateral muscles buccal ganglion buccal mass salivary gland anus Oesophageal pouch pouch oesophagus radular sheath rectum intestine stomach Fig. 5. Alimentary canal of Pila buccal cavity buccal glands muscles of odontophores subradular organ oesophagus jaw sphincter vestibule Subradular membrane mouth lateral cartilage of odontophore radula sphincter radular sac Sublingual cavity Fig. 6. Pila: Buccal mass in V.L.S. 12 flap strap of radula cutting edge transverse row of teeth Fig. 7. Radula of Pila outer Inner marginal marginal lateral Fig. 8. A jaw of Pila. median rachidian lateral Inner outer marginal marginal Fig. 9. A single row of radular teeth of Pila 13 FEEDING AND DIGESTION: Pila is herbivorous. The food consists of aquatic vegetation like Vallisnaria and Pistia and algae attached to rocks etc. With the help of the jaws, the food such as Vallisnaria and Pistia leaves are cut into smaller pieces and then pushed into the buccal cavity with the help of the forward and backward movement of the radula. On the other hand while feeding on algal materials, the protractor muscles of the odontophore pull the odontophore forward causing it to protrude slightly out of the mouth over the substratum. The protractor and the retractor muscles of the radula now contract alternately thereby moving the radula forward and backward over the odontophore. The radula thus moves against the substratum and the radular teeth scraps away the algae and other particles on the substratum into the buccal cavity. The contraction the retractor muscles of the odontophore returns the odontophore to its resting position thus drawing the scraped food into the buccal cavity. The food is cut and masticated in the buccal cavity. The salivary glands pour their secretions into the buccal cavity where it mixes with the food. The starch is converted into soluble sugars. The food is digested in the stomach by the secretions of the digestive gland. The digestive juices of the digestive glands bring about the extracellular digestion while the intracellular digestion takes place in the resorptive cells of the digestive glands. Cellulose is digested intracellularly in the digestive gland. The digested food is absorbed in the digestive gland and the intestine. The undigested food is voided out from the anus into the branchial chamber from where it is thrown out of the body with the outgoing water current through the right pseudopipodium. RESPIRATORY SYSTEM: Pila is unique in having double mode of respiration: 1. Aquatic by ctenidium 2. Aerial by pulmonary sac. AQUATIC RESPIRATION: Aquatic respiration takes place by a single ctenidium which lies in the branchial chamber attached to the dorso-lateral wall of the mantle on the right side. Ctenidium is an elongated structure made up of numerous thin and triangular leaflets or lamellae. The lamellae are arranged transversely in a single row one behind the other, running parallel along the ctenidial axis. The ctenidial axis remains attached to the mantle wall. This type of ctenidia are known as monopectinate type (Fig. 10 A). The lamellae are not of equal size, the ones in the middle of the ctenidia are the biggest and their size reduces towards the two ends. Each lamella has two free ends corresponding to two sides of the triangle. One of the end is longer than the other. The longer side is known as the efferent side while the shorter one is the afferent side (Fig. 10B). The ctenidium is supplied with blood vessels, an afferent and an efferent blood vessel (Fig. 10C). Ctenidium is innervated by nerves from the left pleural and supra-intestinal ganglia. Although the ctenidium is attached to the right side, its blood and nerve supply shows that it is the gill of the left side which has shifted to the right side due to the development of the large pulmonary cavity on the left side (due to torsion- discussed later). A vertical section passing through the lamellae and the mantle shows that each lamella is made up of double layer of epithelial cells enclosing a narrow space between them. The epithelial cells consists of ciliated columnar cells, non-ciliated columnar cells and a few glandular cells. The epithelial layer is supported by muscle fibres and connective tissue(Fig. 10D). 14 MECHANISM OF RESPIRATION: AQUATIC RESPIRATION: During aquatic respiration the two nuchal lobes expand to form shallow channels allowing the ingress (entry) and egress (exit) of water into the branchial chamber. The water current enters the mantle cavity through the left nuchal lobe and comes in contact with the osphradium which tests the nature of water. It then reaches the posterior part of the pulmonary chamber, crosses the epitaenia and reaches the posterior part of the branchial chamber (Fig.11A). As the water current moves forward in the branchial chamber it bathes the ctenidium. The carbondioxide of the blood contained in the blood vessels of the ctenidium is exchanged for the oxygen dissolved in water. After the exchange of gases the water is expelled out through the right nuchal lobe (Fig. 11A). A continuous water current is maintained by the beating of the cilia lining the ctenidium and by the alternate rise and fall of the floor of the mantle cavity brought about by the protrusion and retraction of the head. 15 Water flow between adjacent plates or lamellae B Afferent side D A Ctenidial lamellae Afferent vessel blood Ctenidial lamellae Transverse pleats Efferent blood vessel Space between two epithelial layers Efferent side Non-cilliated epithelial cells Ctenidial axis Afferent blood vessel Cilliated epithelial cells C Basement membrane Axis Connective tissue Muscle fibres Efferent blood vessel Pallial epithelium Blood to heart Fig. 10. Pila ctenidium A. Monopectinate gill B. Diagramme to show passage of water over four gill lamellae C. Flow of blood in a gill lamella D. A lamella in section 16 PULMONARY OR AERIAL RESPIRATION: The pulmonary sac is a modification of the mantle and is a closed cavity which hangs from the dorsal wall of the mantle in the pulmonary chamber. The pulmonary sac has thick vascular walls. The sac communicates with the pulmonary chamber by a large oval opening guarded by two unequal valves (Fig. 11B). MECHANISM OF RESPIRATION: The aerial respiration occurs either when the pond water is polluted and deficient in oxygen or when Pila comes on land. For aerial respiration, Pila comes up to the surface of water and thrusts out its left nuchal lobe which elongates and becomes rolled up into a tube or siphon. The siphon extends beyond the water level so that the atmospheric air is drawn into the pulmonary sac through it. The pulmonary sac now becomes greatly enlarged and occupies nearly two-thirds of the pulmonary chamber. The pulmonary chamber is completely cut off from the branchial chamber by the epitaenia which comes in contact with the roof of the mantle during this process (Fig. 11B). The alternate contraction and expansion of the mantle wall and the pulmonary sac helps in the process of inspiration and expiration of air. The expelled air goes out of the pulmonary chamber by the same route. The walls of the pulmonary sac are richly supplied with blood vessels. When Pila is on land, the respiratory tube is not formed and the air enters the pulmonary chamber directly through the expanded nuchal lobe. During aestivation the life activities of Pila are at low ebb and the air stored up in the pulmonary chamber seems to be the only source of oxygen for respiration. 17 head left nuchal lobe right nuchal lobe Water excurrent Water excurrent head Air current siphon Branchial chamber Pulmonary sac ctenidium epitaenia osphradium Bloobbb auricle ventricle Pulmonary chamber B A Fig. 11. Pila. Mechanism of respiration A. Course of water current in aquatic respiration B. Course of air current in aerial respiration efferent ctenidial vein efferent renal vein pulmonary vein auricle ventricle visceral aorta aortic ampulla cephalic aorta Fig. 12. Heart of Pila 18 Opening of pulmonary sac into pulmonary chamber of mantle cavity CIRCULATORY SYSTEM: The circulatory system is open and consists of heart, arteries, veins and the sinuses and has attained great complexity because of its amphibious nature. The heart is situated on the left side of the visceral mass very close to the posterior end of the ctenidium (Fig. 12 and 13). The heart is enclosed in a thin-walled coelomic cavity called pericardium. The pericardium is situated between the pulmonary chamber and the posterior renal sac on the left side of the body whorl (Fig.12). It extends anteriorly upto the stomach and digestive gland and communicates with the posterior renal chamber by a reno-pericardial aperture. Heart consists of two chambers: an auricle and a ventricle (Fig. 12). The auricle is thin walled, highly contractile and lies in the dorsal part of the pericardium. Ventricle is thick walled, spongy and muscular situated just below the auricle in the same vertical axis. Ventricle has a reduced cavity due to a coarse meshwork of muscular strands. The auricle communicates with the ventricle by a auriculo-ventricular aperture which is guarded by two semilunar valves to prevent the backflow of blood from the ventricle to the auricle(Fig. 13). The auricle receives oxygenated blood from the ctenidium and the pulmonary sac through the efferent ctenidial vein and the pulmonary vein. It also receives blood from the posterior renal chamber by efferent renal vein. The lower end of the ventricle gives rise to an aorta which immediately divides into two branches: 1. An anterior cephalic aorta 2. Posterior visceral aorta . The opening between the ventricle and aorta is guarded by two semilunar valves which prevent the backflow of blood into the ventricle. CEPHALIC AORTA: The cephalic aorta supplies blood to the head region. It is swollen into a thick walled contractile aortic ampulla at its base which helps in the propulsion of the blood. The cephalic aorta gives off three branches on its outer side: 1. A fine cutaneous artery to the skin. 2. 3. A thick oesophageal artery to the oesophagus. A thick pallial artery to the left side of the mantle, left nuchal lobe and the osphradium. The inner side of the cephalic aorta gives off a single pericardial artery to the pericardium which finally enters the posterior renal chamber and gives off branches to both the renal chambers and part of reproductive system. The cephalic aorta enters the perivisceral sinus (space surrounding the buccal mass and oesophagus) along the left side of the oesophagus and then crosses beneath the oesophagus and gives off many arteries to the buccal mass, oesophageal wall, right side of the mantle, right nuchal lobe, copulatory organ, eyes, tentacles etc. 19 tentacular artery optic artery pedal artery penial artery Radular sac artery right siphonal artery left siphonal artery right pallial artery osphradium ctenidium osphradial artery left pallial artery afferent ctenidial vein oesophageal artery cutaneous artery pericardial artery ventricle efferent ctenidial vein auricle aortic ampulla pericardial sinus visceral aorta efferent pulmonary vein branchio-renal sinus perivisceral sinus pericardial artery hepatic artery efferent renal vein afferent renal sinus renal artery gastric artery intestinal artery posterior renal chamber anterior renal chamber afferent renal vein Fig. 13. Heart and blood vascular system of Pila 20 The visceral aorta passes through the visceral mass and supplies blood to the visceral organs. It gives out: 1. 2. 3. 4. 5. A small pericardial artery to the pericardium. A large and stout gastric artery to the stomach. Intestinal arteries to the intestine. A hepatic artery to the digestive gland and the gonads. Renal arteries to the anterior and posterior renal chamber. Finally the visceral aorta terminates into branches that supply the rectum and the anus. The blood that is supplied to the various organs of the body by the arteries and its branches is finally collected in small spaces called the lacunae. These lacunae unite to form large sinuses (Fig. 13). Sinuses are spaces between the body wall and visceral organs and together with the lacunae form the haemocoel. They connect arteries to veins and serve as capillaries. There are four blood sinuses in Pila: 1. A peri-visceral sinus surrounding the anterior part of the alimentary canal. It contains deoxygenated blood. Three channels carry blood from this sinus to the renal chamber and the pulmonary sac. 2. Peri-intestinal sinus surrounding the terminal part of the intestine and the the genital duct. It collects blood from the digestive gland and the visceral organs. 3. Branchio-renal sinus lying outside the anterior renal chamber and leading into the afferent ctenidial vein. It receives blood from the roof of both the renal chambers. 4. Pulmonary sinus present in the wall of the pulmonary sac. It receives blood from the peri-visceral sinus. The blood from different organs of the body is collected by three main veins and poured into the auricle either directly or through the respiratory and excretory organs. 1. Afferent ctenidial vein: It collects deoxygenated blood from the peri- visceral sinus, the rectum and the terminal part of the genital ducts and carries it to the ctenidium where the blood is oxygenated. The efferent ctenidial vein collects the oxygenated blood from the ctenidium, mantle, and the copulatory organs and pours it into the auricle. 2. Afferent renal vein: It receives blood from the peri-intestinal sinus and carries it to the posterior renal chamber. The efferent renal vein then collects blood from the posterior renal chamber and conveys it to the auricle. 3. Pulmonary vein collects blood from the pulmonary sinus and sends it to the auricle. BLOOD: Blood is colorless because of the absence of haemoglobin. It contains the respiratory pigment haemocyanin which is dissolved in blood plasma. Haemocyanin contains copper and thus it becomes blue when it combines with oxygen and colorless when it combines with carbondioxide. Blood also contains some colorless stellate amoeboid cells which are phagocytic in nature and help in the removal of waste substances. Some intracellular digestion also takes place in these amoeboid cells. 21 COURSE OF CIRCULATION: The cephalic and visceral aorta supplies blood to different parts of the body. The blood is then collected from various parts of the body in two main sinuses, the peri-visceral and periintestinal. From these sinuses, the blood passes either into the ctenidium, pulmonary sac or the kidney (Fig. 13). During aerial respiration, the blood flows from the peri-visceral sinus into the pulmonary sac and after aeration comes to the auricle by the pulmonary vein. During aquatic respiration, the blood flows from the perivisceral sinus to the ctenidium and after aeration comes to the auricle by the efferent ctenidial vein. The blood from the peri- intestinal sinus takes two courses: it either goes to the anterior renal chamber from where it goes into the ctenidium where it is purified and finally through the efferent ctenidial vein enters the auricle, or from the peri-intestinal sinus, the blood goes to the anterior renal chamber and then into the posterior renal chamber. However, the blood can enter the posterior renal chamber directly from the peri-intestinal sinus. Whatever may be the case, the blood is finally carried from the posterior renal chamber by the efferent renal vein to the auricle. This blood is not aerated. Thus, the aerated and non-aerated blood gets mixed up in the auricle. This mixed blood then enters the ventricle and is distributed to the arteries. The renal chambers remove the excretory products from the blood. CEPHALIC AND VISCERAL AORTA ALL THE DIFFERENT ORGANS OF THE BODY VENTRICLE PERI-INTESTINAL SINUS AURICLE PULMONARY VEIN PULMONARY SAC PERI-VISCERAL SINUS AFFERENT AFFERENT EFFERENT CTENIDIAL VEIN CTENIDIUM AFFERENT CTENIDIAL VEIN EFFERENT RENAL VEIN 22 BRANCHIO RENAL SINUS RENAL RENAL SINUS VEIN ANTERIOR RENAL POSTERIOR RENAL CHAMBER CHAMBER EXCRETORY SYSTEM: Excretion is performed by a large thick walled muscular renal organ or kidney which consists of an anterior chamber and a posterior chamber, the two communicating with each other by an aperture guarded by a valve. The renal organ communicates with the exterior on one hand and with the pericardial cavity on the other (Fig.14). ANTERIOR RENAL CHAMBER: The anterior chamber is more or less ovoid, reddish in color, smaller than the posterior chamber, situated in front of the pericardium. It opens into the mantle cavity by an elongated aperture on the right side of the epitaenial ridge. On the other end it communicates with the posterior renal chamber through an internal opening. The internal cavity of the anterior chamber is reduced due to the presence of numerous lamellae projecting both from the roof and the floor. The lamellae are arranged on the floor on either side of a single median axis, the afferent renal sinus and also on the roof on either side of the median longitudinal axis, the efferent renal sinus (Fig. 14). POSTERIOR RENAL CHAMBER: The posterior renal chamber lies on the left side of the rectum, closely pressed against the pericardium and the digestive gland. Its cavity communicates with the pericardium by the reno-pericardial aperture at one end while the other end communicates with the anterior renal chamber by an aperture. The posterior renal chamber is broad, somewhat hook shaped, brownish to grey in color. Its large internal cavity encloses a part of the genital duct and a few coils of the intestine. The roof of the chamber is richly supplied by the branches of the afferent and efferent renal vessels (Fig. 14). 23 root of posterior renal chamber intestinal coils into cavity of posterior renal chamber efferent renal vein anterior renal chamber afferent renal sinus afferent renal vein auricle reno-pericardial aperture opening into mantle cavity inter-renal aperture connecting the two renal chamber ventricle auricle pericardium aortic ampulla Fig. 14. Excretory organ of Pila 24 MECHANISM OF EXCRETION: The walls of both the chambers are richly supplied with a network of blood vessels. The renal chambers separate the nitrogenous waste material from the blood. The excretory products are passed from the posterior renal chamber into the anterior renal chamber, from where it is discharged into the mantle cavity through the renal duct. The waste products are finally expelled out of the body from the mantle cavity through the right nuchal lobe along with the outgoing water current. The excretory fluid mainly contains ammonia, ammonium compounds, urea and uric acid. During the aquatic phase, Pila excretes out ammonia (ammonotelic) but during the terrestrial phase it excretes out uric acid (uricotelic). This is so because during the terrestrial phase it needs to conserve water. NERVOUS SYSTEM: The nervous system consists of a series of paired ganglia with commissures and connectives (Fig. 15). The term commissure is applied to a nerve cord connecting two similar ganglia, while connective is applied to a nerve cord connecting two dissimilar ganglia. • A pair of cerebral ganglia lies anteriorly on the dorso-lateral sides of the buccal mass, one on each side of the head. The two cerebral ganglia are connected by a thick cerebral commissure running transversely above the buccal mass. Each cerebral ganglia gives off nerves that innervates the eye, the snout, statocyst and the tentacles of its side. • A pair of buccal ganglia lies at the junction of the buccal mass and the oesophagus. They are connected to each other by a delicate transverse buccal commissure. Each buccal ganglion is also connected to the cerebral ganglion of its side by a cerebrobuccal connective, so that a nerve ring is formed anteriorly encircling the gut. The buccal ganglion gives off nerves to the buccal mass, radular sac, salivary glands, oesophagus and the oesophageal pouches. • A pair of pleuro-pedal ganglionic mass is present on the ventro-lateral side of the buccal mass, one on the right side and the other on the left side. Each pleuro-pedal ganglionic mass is formed by the partial fusion of outer pleural and inner pedal ganglia which lie very close to one another and are separated only by a notch. A small infra-intestinal ganglion is also fused with the right pleuro-pedal ganglionic mass, also called as pleuro-pedal infraintestinal ganglionic mass. The two pedal ganglia are connected by a broad pedal commissure lying under the buccal mass. The pedal ganglion gives out numerous nerves to the foot. The left pleural ganglion supplies the nerves to the body wall, mantle, osphradium, left nuchal lobe, columellar muscles, and anterior part of ctenidia while the right pleural ganglion innervates the parietal wall, epitaenia, right nuchal lobe, copulatory organs, columellar muscles and the rectum. The cerebral ganglion of each side is connected to the pleuro-pedal ganglionic mass of its side by two connectives called the cerebro-pedal (connecting the cerebral and the inner pedal ganglion) and cerebro-pleural connective (connecting the cerebral and the outer pleural ganglion). • A very slender nerve called the infra –intestinal nerve connects the pleural ganglion of the two sides. It forms a loop just behind the pedal commissure. 25 • An unpaired supra-intestinal ganglion lies a little behind the left pleuro-pedal ganglionic mass. The supra-intestinal ganglion is connected to the right pleuro-pedal ganglionic mass by a very fine oblique nerve lying just above the oesophagus called as the supra-intestinal nerve. The supra-intestinal ganglion supplies nerves to the ctenidium and the pulmonary sac. • The visceral ganglion is really a ganglionic mass formed by the fusion of two ganglia lying very close to anterior lobe of the digestive gland. The pleural ganglion is connected with the visceral ganglion by pleuro- visceral connective on each side. The right pleuro-visceral connective lies below the oesophagus and is called as infraintestinal visceral connective. The left pleuro-visceral connective lies above the oesophagus and is called as supra-intestinal visceral connective. • The visceral ganglion gives off nerves to the renal organs, genital organ, pericardium, intestine and the digestive glands. 26 cerebral commissure right cerebral ganglion Buccal commissure cerebro-buccal connective cerebro-pedal connective cerebro-pleural connective right buccal ganglion statocyst ganglionic mass right pedal right pleural infra intestinal left pedal left pleural ganglionic mass pallial nerve zygoneury parietal nerve pedal commissure infra-intestinal nerve osphradial-pallial nerve supra-intestinal ganglion supra-intestinal nerve Supra-intestinal-visceral connective infra-intestinal-visceral connective visceral ganglion Fig. 15. Nervous system of Pila 27 SENSE ORGANS: The sense organs include two pairs of tentacles, osphradium, a pair of eyes (ommatidia), and a pair of statocyst. TENTACLES: Two pair of tentacles are present in the head region. The first pair of tentacles are contractile and are known as labial palp while the second pair is longer, filamentous and retractile and is the true tentacle, present behind the first one. Both the tentacles are tactile in nature. Tentacles are supplied by nerves from the cerebral ganglia. OSPHRADIUM: There is only one osphradium that lies suspended from the roof of the mantle cavity on the left side. It is oval in shape with inner or right end bluntly rounded while the outer or left end pointed (Fig.16 A). It has a central axis to which two rows of thick, fleshy leaflets are attached. Leaflets are largest in the middle of the osphradium. In a transverse section, the osphradia is made up an outer layer of epithelial cells lined internally by a basement membrane (Fig. 16B). The epithelial cells are elongate and consists of three types of cells: 1. Sensory cells – are devoid of cilia and cover the osphradium. 2. Ciliated cells 3. Glandular cells are found scattered among the sensory cells. Osphradium is innervated by a nerve from the left pleural ganglion. The osphradium is an olfactory organ that lies in the path of water current entering the mantle cavity. It enables the animal to test the physical and chemical nature of the incurrent water and also assists in the selection of food. If the incurrent water is foul then it is prevented from entering the mantle cavity by the closure of the left nuchal lobe. STATOCYST: A pair of small, pyriform, cream colored statocyst lie one on either side of the pleuro-pedal ganglionic mass. Each statocyst is a hollow capsule surrounded by thick, tough, leathery covering of connective tissue. The statocyst is ectodermal in origin, filled with a fluid containing calcareous particles known as statolith or statoconia (Fig. 16C). Statocyst are balancing organs and regulate the position in water and on land. It gets its nerve supply from the cerebral ganglion. 28 sensory cells Blood spaces gland cells lamellae Non-ciliated epithelial cells nervous mass central axis connective tissue nerve mantle A. Osphradium B. T.S. of Osphradium Connective tissue outer cornea inner cornea Capsule of statocyst lens retina nerve statoconia optic vesicle connective tissue optic nerve C. Statocyst D. H.L.S. of eye Fig.16 Pila 29 EYES: A pair of short, black and stalked simple eyes are present one on either side of the head at the base of each true tentacle. The stalk is known as ommatophore. Each eye is covered in front by the transparent cornea which is actually the part of the general epithelium of the body. In section, eye is in the form of a cup like invagination called the optic vesicle lined by photosensitive or pigmented retinal cells. The cavity of the vesicle is filled with an oval hyaline structure called the lens. The anterior part of the optic vesicle in front of the lens is transparent and forms the inner cornea (Fig. 16D). The optic vesicle is surrounded by a firm sheath of connective tissue. The optic nerve enters the retina from the posterior end of the vesicle. Although, Pila has an elaborate visual organ with all the components for photoreception, it is poor in sight and cannot distinguish objects. It can only respond to variations in light intensity. The eye is chiefly adapted to work in dim light. REPRODUCTIVE SYSTEM: Sexes are separate and sexual dimorphism is distinct. The shell of the female is larger and more globular than the male. Males have a less swollen body whorl, and a more developed copulatory organ than the female. MALE REPRODUCTIVE SYSTEM: A single cream colored testis lies in the upper two or three whorls in close contact with the digestive gland. The testis is a flat plate-like structure, more or less triangular in shape. Many fine ducts known as vasa efferentia originate from different parts of the testis. The vasa efferentia unite together to open into a common vas deferens (Fig. 17). The vas deferens is divisible into three distinct parts: 1. The proximal thin walled tubular part 2. The middle slightly swollen seminal vesicle which stores the sperms 3. Terminal glandular part lying left of the rectum and opening into the mantle cavity close to the anus The male copulatory organ is the penis which arises from the edge of the mantle in front of the anus and is enclosed in a penis sheath. The penis sheath is a simple outgrowth from the inner surface of the mantle. At the base of the penis sheath is a glandular thickening- the hypobranchial gland of unknown function. The spermatozoa of Pila are of two types: 1. Hair shaped or eupyrene sperm 2. Worm-like oligopyrene sperm Only the eupyrene sperms are functional and are capable of fertilizing the eggs. Also, the eupyrene sperm show a distinct head, a middle piece and a tail. The head and middle piece cannot be distinguished in oligopyrene sperm and also the tail bears four or five cilia (Fig. 17). The oligopyrene sperms are non-motile and cannot fertilize the eggs. FEMALE REPRODUCTIVE SYSTEM: A much branched orange colored ovary is situated in the upper two or three whorls, attached to the inner surface of the digestive gland. The oviduct leads from the ovary and extends downwards along the digestive gland. It is differentiated into an upper tubular part and the lower glandular part that remains on the floor of the mantle cavity parallel to the rectum. The glandular part is distinguishable into an anterior yellow colored albumen gland that secretes albumen and a posterior part called as uterus (Fig.18). A bean-shaped receptaculum 30 seminalis or the spermatheca is present at the junction of the tubular and glandular portion of the oviduct. The sperms received from the male are temporarily stored in the receptaculum seminalis after copulation. The terminal part of the uterus is differentiated into vagina. The vagina opens into the mantle cavity close to the anus by a female genital aperture situated on a small papilla (Fig. 18). The male intromittent organ, the penis is rudimentary and useless in females. Existence of rudimentary penis in female is suggestive of the fact that this group had hermaphroditic ancestor. A rudimentary hypobranchial gland is present as a glandular thickening. 31 penis sheath testis digestive gland penis vas deferens head rectum seminal vesicle Male genital pore glandular part of vas deferens middle piece tail Fig. 17. Male Reproductive system of Pila penis sheath ovary digestive gland rudimentary penis oviduct albumen gland vagina glandular part uterus female genital pore seminal receptacle cluster of eggs Fig. 18. Female Reproductive system of Pila 32 COPULATION : Pila reproduces only during the rainy season. Copulation takes place on land, on the moist ground of the banks. During copulation, the male and female Pila come close together in such a way that the right nuchal lobe of one is opposed to the other. The copulation lasts for three to four hours at the end of which sperms are transferred from the seminal vesicle of the male to the receptaculum seminalis of the female where the sperms are temporarily stored. FERTILIZATION AND DEVELOPMENT: Fertilization is internal and oviposition commences one or two days after copulation at a suitable place, such as moist earth near ponds. Fertilized eggs are laid in masses of 200-800. Development takes place outside the body of the female and is direct without any larval stage. During development, the shell of the embryo become spirally coiled and undergoes torsion resulting in asymmetry of the body. A young snail emerges from the fertilized egg. The young ones resemble the adult in form. 33 SEPIA COMMONLY KNOWN AS CUTTLE FISH HABIT AND HABITAT: Sepia is a marine mollusc found in shallow coastal water. They are active swimmers and usually swim at night and rest flat at the bottom during day time. The lateral fins, the funnel and the arms help in swimming. They can make burrows with the help of funnel and can use fins as shovel to cover the body with sand. They have a flattened body which indicates their sand and mud dwelling habit. They usually move in groups. At the time of danger by its enemy, it discharges a dark ink from the ink sac which forms a dark screen and the cuttlefish escapes the enemy under this dark cover. Sepia is carnivorous, feeds on crustaceans, small fishes and other small animals. The cuttlefish can change its color to blend with its surrounding and may exhibit bioluminescence. MORPHOLOGY: The body is divisible into a head and an elongated dorso-ventrally flattened trunk. The head and the trunk are joined by a narrow neck. HEAD: The head bears a pair of well developed eyes laterally and mouth at its free end. The mouth is surrounded by five pairs of arms. Of the five pairs, four pairs are short and stout and bear suckers on their inner side. The suckers are arranged on a short stalk in longitudinal rows. Each sucker is a muscular cup having a horny rim with minute teeth and help the animal to seize the prey or secure attachment to the substratum by creating a partial vacuum. The fifth pair of arm is long and narrow. It ends in a club-shaped extremity with suckers and are known as the tentacles (Fig. 19). The tentacles can be retracted into large depressions at their bases. During breeding season, the fourth arm of the left side is modified into an intromittent organ called the hectocotylized arm in the males which is used to transfer the spermatophores into the mantle cavity of the female. The oral arms are considered as the modification of the foot. TRUNK: The trunk is flat and shield shaped, with its narrow end directed away from the oral end. The trunk is slightly convex from above and is called the anterior or antero-dorsal surface. It is pigmented and hard due to the presence of shell under the integument. The side opposite to it is flat and soft and is the posterior or postero-ventral surface. The margins of the trunk are fringed with thin, lateral, muscular folds called the fins. The fins assist in swimming. MANTLE: Trunk is enclosed in a thick tegument, the mantle. The mantle encloses the mantle cavity on the posterior side of the trunk. The visceral organs occupy a major portion of the mantle cavity. The mantle cavity communicates to the exterior by a tube-like funnel or siphon present on the mid-ventral side of the oral end. The other end of the funnel opens into the mantle cavity by a wide internal aperture. Present on the ventral surface of the funnel are two oval cartilaginous depressions called the funnel or infundibular cartilages. 34 oral arms suckers tentacle eye head collar trunk suckers Lateral fin Fig. 19. Dorsal view of Sepia (Male) 35 At the oral border, on the inner surface of the mantle are two oval cartilaginous projections called mantle cartilages (Fig. 20 and 21). The mantle and funnel cartilages interlock with each other as a result of which the direct exit of water from the mantle cavity to the exterior is prevented. Thus, the water is allowed to flow out from the mantle cavity through the funnel. The funnel is provided by a semilunar valve that opens outwards and allows the water to flow out from the mantle cavity but prevents the entry of water into the mantle cavity from outside. The water can enter the mantle cavity by a wide cleft present between the free edge of the mantle and the neck. The funnel is innervated by nerves from the pedal ganglia and is therefore regarded as the modification of the foot. A longitudinal incision of the mantle through the mid-ventral line exposes the internal organs which occupy the major part of the mantle cavity (Fig.20 and 21). The internal organs consist of: • A pair of ctenidia one on each side of the mantle cavity. • • A tubular rectum present in the middle line that opens into the mantle cavity by anus. A renal sac is present on either side of the rectum. It opens into the mantle cavity by a renal pore. The ink sac with its duct opening into the rectum close to the anus. Gonads are located dorsal to the ink sac. The testis in males is partly covered by the ink sac. In females, a pair of nidamental glands and a pair of accessory nidamental glands are present between the ctenidia (Fig. 20). Genital duct opens on the left side near the left renal aperture by an external opening called the genital aperture. A pair of stellate ganglia is present on either side of the mantle cavity close to the oral end. • • • • 36 head funnel opening anus funnel valve eye rectum funnel cartilage stellate ganglion female genital opening mantle cartilage renal pore gland of oviduct ctenidium accessory nidamental gland mantle cavity nidamental gland visceral mass oviduct Ink-sac stomach ovary cut open parapodia Fig. 20. Mantle cavity of a female Sepia: Visceral sac partly dissected mouth tentacle Oral arms Funnel opening lip funnel cartilage head eye stellate ganglion anus male genital opening rectum renal pore penis ctenidium ink-duct mantle cavity Lateral fin testis cut open branchial heart visceral mass ink sac Fig. 21. Mantle cavity of a male Sepia 37 SHELL: The shell is internal. It is bilaterally symmetrical and is called the cuttle bone. The shell is calcareous in nature, flat and ovoidal in shape, enclosed in a shell sac. Its oral end is slightly broad and round, while the aboral end narrow and slightly pointed, terminating in a sharp spine or rostrum. The shell is secreted by the epithelial lining of the shell sac. The shell is a non- living structure formed of thin parallel layers or lamellae of calcium carbonate. The lamellae enclose air-spaces between them to make the shell light. Thus, the shell not only acts as an endoskeleton giving rigidity to the trunk but also acts as a float. As the shell is dorsally present , it helps in maintaining equilibrium of the body. The shell of the dead cuttlefishes is called as sea foam. As the shell of cuttle fish is light, it floats in water and during the monsoon months a large number of these are drifted ashore and hence the name sea foam. Sepia can change its color because of the presence of flat, contractile, pigment containing cells called the chromatophores. The chromatophores are present in the dermis of the integument. The chromatophores contain three kind of pigments: dark, yellow brown and orange. These cells have contractile radiating fibres attached to their walls at one end and to the integument at the other end. Contraction of the fibres dilates the cells and the pigment become diffused. This disperses the pigment over a larger surface, making it more visible. On the contrary, relaxation of the fibres causes the contraction of the cell and the pigment becomes concentrated in a smaller area, rendering it less visible. This results in the color changes of the skin. Lying beneath the chromatophores are iridocytes which are transparent cells that impart iridescence to the skin of Sepia. The coloration of the skin at a particular time is due to the combined effect of the chromatophores and the iridocytes. The color changes are under the nervous control. COELOM: True coelom is represented by viscero-pericardial coelom and the cavities of the renal sac. The viscero-pericardial coelom is divided into two parts: an anterior pericardial cavity which encloses the heart and the posterior gonocoel that encloses the gonads. The visceropericardial cavity communicates with the renal sacs by a pair of reno- pericardial aperture. LOCOMOTION: Sepia is a good swimmer. It makes use of its fins and funnel in swimming. It swims about gently forwards and backwards by the undulating movements of the fins bordering the lateral sides of the trunk. During swift backward darting movements it makes use of its funnel. Sepia darts quickly by forcing out jets of water through the funnel by the rhythmic contraction of the muscular mantle. The mantle cavity enlarges or contracts by the alternate contraction and relaxation of the mantle. When the mantle relaxes, the mantle cavity enlarges and the water from outside is freely drawn in. When the mantle contracts, the pallial aperture closes due to the interlocking of the mantle cartilages with the funnel cartilage. This results in a flow of water through the funnel like a jet and the animal darts back quickly. DIGESTIVE SYSTEM: The mouth lies amidst the arms. The mouth is surrounded by a membranous circular lip provided with many papillae. Within the circular lip lies a pair of strong, curved horny jaws. Mouth opens into a thick walled buccal cavity which possesses the odontophore and the radula ( refer Pila). The radula however, is comparatively weak. The buccal cavity leads into a straight, narrow tube, the oesophagus which in turn opens into a rounded, thick walled, 38 muscular sac, the stomach. The stomach leads into a short, tubular intestine which runs parallel to the oesophagus towards the oral side and opens into the rectum (Fig. 22). The rectum finally opens into the mantle cavity by the anus. The duct of the ink sac is connected with the rectum. A spirally coiled pouch, the caecum opens into the alimentary canal at the junction of the stomach and intestine. Two pairs of salivary glands are present in Sepia. The anterior pair of salivary glands lies inside the buccal mass and their ducts open on either side of the radula. A pair of posterior salivary glands is situated close to the oesophagus. The ducts of the two salivary glands unite into a common, median duct which opens into the buccal cavity. The secretion of both the pair of salivary glands seem to be of poisonous nature and paralyses the prey instead of digesting the food. Two sets of digestive glands are present in Sepia. One of them is a large brownish gland extending from near the posterior salivary gland close to the aboral end of the body and is called the liver. The ducts from the liver open into the stomach near the point of origin of the caecum. The other gland is clustered around the ducts of the liver and open into them. It is small, wedge shaped, creamy in color and is called the pancreas (Fig. 22). FEEDING AND DIGESTION: Sepia is carnivorous and feeds on crustaceans, other molluscs and small fishes. The tentacles are extended out and attached to the prey with the help of the suckers. The tentacles then shorten, bringing the prey within the reach of the arms. The arms then bend over the prey and hold it firmly with their suckers. The prey is paralysed by the secretions of the salivary glands and cut into small pieces with the jaws. The small food pieces are passed into the buccal cavity, from where it is passed into the oesophagus by radula. From oesophagus, the food is pushed by peristaltic action into the stomach. Digestion takes place in the stomach. The liver and pancreas pour their secretions into the stomach for digesting the food. The semi-digested food is passed into the caecum where digestion is completed. Absorption of food takes place in the caecum and intestine. The undigested food is passed out into the mantle cavity through the anus to be finally expelled out from the body via siphon alongwith the outgoing water current. Digested food is stored in the liver and the pancreas. 39 Dorsal Posterior Anterior jaw muscle digestive gland oesophagus digestive duct buccal cavity upper jaw pancreas lips rectum posterior anus salivary gland stomach ink sac duct ink sac anterior salivary gland radula jaw muscle odontophore ink gland caecum lower jaw Ventral Fig. 22. Lateral view of digestive tract of Sepia 40 INK-SAC: The ink sac is a pear shaped organ that lies on the anterior wall of the mantle cavity (Fig. 23). It opens into the rectum by a cylindrical ink duct. The inner glandular mass secretes a blackish ink. When cuttlefish is in danger of attack by an enemy the ink is forced out to produce a colored surrounding under the cover of which it escapes from the enemy. anus sphincter muscles ampulla rectum ink-duct cavity of ink-sac Orifice of ink-gland ink-gland Fig. 23. Sepia. Ink-sac in M. L. S. RESPIRATORY SYSTEM: Respiratory organs are a pair of ctenidia one on each side of the mantle cavity. Each ctenidium is bipectinate consisting of a central axis and numerous delicately folded lamellae on either side. The central axis is attached throughout the greater part of its length to the mantle wall by a thin, muscular fold which keeps the ctenidium in place. The blood flows into the ctenidium by afferent branchial vessels and returns to the auricle by efferent branchial vessels. MECHANISM OF RESPIRATION: Rhythmic contraction and relaxation of the mantle enlarges and reduces the mantle cavity. When the mantle cavity is enlarged, current of water containing oxygen enters the mantle cavity through the pallial aperture. The water then passes over the ctenidia where exchange of gases takes place in the vascular lamellae. The foul water containing carbondioxide passes out through the funnel when the mantle cavity is reduced. 41 CIRCULATORY SYSTEM: Sepia has a well developed closed circulatory system. The heart lies in the middle of the visceral mass enclosed in the pericardium and consists of a large, median thick walled ventricle and two small, lateral, thin walled auricles. Anteriorly, the ventricle gives off a stout anterior or oral or cephalic aorta and posteriorly a smaller aboral or posterior aorta (Fig. 24). The anterior aorta gives off branches (arteries) to the mantle, alimentary canal, the funnel and the head. The posterior aorta gives off branches (arteries) to the viscera, the renal sacs and the gonads. The arteries divide and redivide to form fine capillaries which in turn reunite to form larger vessels, the veins. Blood from different parts of the body is collected into a large vena cava. The vena cava bifurcates anteriorly into lateral vena cava which pass through the kidney and continue as afferent branchial vein at the base of the ctenidium. The afferent branchial vein at the base of the ctenidium dilates to form the contractile branchial heart (Fig. 24,25). Attached to the branchial heart is a glandular appendage (Fig. 24). A pair of pallial vein brings blood from the mantle, and a pair of abdominal vein from the posterior region of the body and open into the branchial heart of their side. The genital vein collects blood from the gonads while the ink sac vein collects the blood from the ink sac and opens into the right afferent branchial vein. Thus, the blood from different parts of the body is brought to the branchial heart from where the deoxygenated blood is pumped into the ctenidia by afferent branchial vein. The blood is then oxygenated in the ctenidia and returned to the auricle by efferent branchial vein. Heart is guarded by semi-lunar valves which prevent the backflow of blood from the ventricle to the auricle or from the aorta to the ventricle (Fig. 24, 25). Like Pila the respiratory pigment is haemocyanin. The blood is colorless when deoxygenated and light-blue when oxygenated. EXCRETORY SYSTEM: The excretory system consists of a pair of thin walled renal sacs or kidneys which communicate with the mantle cavity by two separate apertures, the excretory or renal pores, one on each side. The right and the left renal sac communicate with one another both anteriorly and posteriorly. The afferent renal vein passes through the corresponding renal sacs, where they are surrounded by masses of glandular tissue (Fig 24 and 26). The waste products are extracted by the glandular walls from the blood in the form of guanin and passed into the renal sac. From here the waste products are finally discharged into the mantle cavity to be thrown out of the body with the outgoing water. 42 anterior vena cava afferent branchial vessel Renopericardial canal gills branchial heart branchial heart appendage abdominal vein branchial heart pericardial cavity renal appendage lateral vena cava renal sac auricle efferent branchial vessel cephalic aorta systematic heart Fig. 24. Cephalopod hemal and excretory system. The arrow indicate the flow of blood and urine 43 tissue capillary anterior vena cava gill aorta afferent branchial vessel branchial heart efferent branchial vessel auricle systematic heart Fig. 25. Diagram showing a simplified pattern of blood flow in Sepia 44 gills efferent branchial vessel vena cava branchial heart branchial pericardial cavity afferent branchial vessel renal appendages mantle cavity Renopericardial canal with primary urine final urine renal sac nephridiopore Fig. 26. Excretory system of Sepia. Solid arrows indicate the flow of blood and dashed arrows show urine movement 45 NERVOUS SYSTEM: The nervous system is highly developed in Sepia and consists of a series of paired ganglia with their commissures and connectives. However, the cerebral , pedal, pleural and visceral ganglia are relatively large, protected by the cranial cartilage and concentrated in the head around the oesophagus due to the shortening of their connectives. The cerebral ganglia or supra-oesophageal ganglia are fused to form a single rounded mass, the brain, situated dorsal to the oesophagus. A pair of thick optic nerves are given off laterally from the cerebral ganglia which expand into optic ganglia near the eyes. The pedal ganglia, like the cerebral ganglia, is also fused into a single mass below the oesophagus. It is anteriorly differentiated into a brachial ganglion that gives out ten brachial nerves to the arms and posteriorly into the infundibular ganglion supplying nerves to the funnel and statocyst. The brachial nerves are connected with one another by a ring commissure. The pedal ganglion is connected to the cerebral ganglia by cerebro-pedal connective (Fig. 27). The pleuro-visceral ganglia are also fused into a single ganglionic mass and lie posterior to the infundibular ganglia behind the oesophagus. They give out two stout pallial nerves posteriorly to the mantle where each one expands into a large, flat pallial or stellate ganglion in front of the ctenidium. The stellate ganglion gives off nerves to the mantle. Swimming and respiratory contractions result from impulses conveyed by small motor neurons given out from the stellate ganglion, one on each side of the mantle wall. A pair of visceral nerves is also given out from the pleuro-visceral ganglia which supplies nerves to the various internal organs. A pair of these branches innervate the ctenidia and are called branchial nerves. The branchial nerve expands into a ganglion known as branchial ganglion at the base of the ctenidium. A pair of nerves called the cerebro-buccal connective run forward from the cerebral ganglia to a pair of superior buccal ganglia lying close to the buccal mass, dorsal to the oesophagus. The superior buccal ganglia are also connected to the inferior buccal ganglia lying ventral to the oesophagus. Nerves are given off from the superior buccal ganglion to the lips and from the inferior buccal ganglion to the buccal mass. A pair of sympathetic nerves arise from the inferior buccal ganglia and run along the sides of the oesophagus to the stomach where it ends in a gastric ganglion (Fig. 27). Gastric ganglion supplies nerves to the stomach, caecum and intestine. A small olfactory ganglion lies close to the optic ganglion and sends a fine olfactory nerve to the olfactory pit. SENSE ORGANS: Sense organs of Sepia are better developed than Pila. They consist of paired eyes, paired statocyst, paired olfactory pits and an unpaired gustatory organ. EYES: The eyes resemble the vertebrate eye. They are large and conspicuous, enclosed in a cartilaginous optic capsule. A transparent cornea covers the eye laterally. The eye ball has a stout outer wall called sclerotic layer which is provided with sclerotic cartilage. The sclerotic layer extends in front as the iris, with pupil in the centre. The pupil can be widened and narrowed by the muscles of the iris. A large, spherical, transparent lens lies behind the iris (Fig. 28). The lens is formed of two plano-convex halves with a layer of cells, the cornea in between. The lens is held in position by ciliary process that project inward from the sclerotic layer. The lens and the ciliary process divide the cavity of the eye ball into two: 46 1. A small anterior chamber in front of the lens filled with a watery aqueous humour 2. A larger posterior chamber containing the jelly-like vitreous humour. The posterior chamber is lined by sensory layer called the retina which contains rods and cones. The retina consists of a single layer of photosensitive retinula cells. The eye is innervated by the optic nerve which dilates into the optic ganglion just behind the eye ball. Nerve fibres pierce through the sclerotic wall and terminate in the retinal cells. Close to the optic ganglion is a small gland of unknown function known as optic gland. A fold of the skin from the head extends in front of the eye and serves as the eye-lid. The eye is capable of forming images. It can adjust to changes in the light intensity by modifications in the size of the pupil and by migration of the pigment in the retina. In bright light the pupil is constricted and screening pigment in the retinula cells move distally to isolate adjacent cells. In dim light the pupil dilates and the screening pigment is withdrawn so that more light reaches the sensory cells. STATOCYST: Situated very close to the pleura-visceral ganglion are two statocyst. The statocyst are spherical bodies enclosed in a cartilaginous capsule. The inner cavity of the statocyst has rounded elevation called the crista statica and macula statica. The crista statica is lined by flattened epithelial cells and the macula statica is composed of elongated cylindrical cells with hair like processes at the free end. A large statolith lies within the statocyst attached to the macula. Statocyst are balancing organs. OLFACTORY PITS: A pair of ciliated olfactory pits are present behind the eyes. They are olfactory in function. GUSTATORY ORGAN: This organ for taste is present as an elevation on the floor of the buccal cavity in front of the odontophore. They are covered with papillae. Arms and tentacles are also regarded as tactile organs. 47 inter-brachial commissure brachial nerve tentacular nerve brachio-buccal nerve superior-buccal ganglion cerebro-buccal connective cerebral ganglion eye muscle eye olfactory ganglion olfactory loop optic ganglion post funnel nerve visceral ganglion pallial nerve stellate ganglion oesophagus visceral nerve nerves to mantle sympathetic nerves branchial nerve posterior visceral commissure branchial ganglion cerebro-buccal nerve fin nerves gastric ganglion Fig. 27. Nervous system of Sepia (dorsal view) 48 secondary or false cornea outer opening of anterior chamber iris-fold rods cilliary process lens retina sclerotic cartilage optic ganglion posterior chamber optic nerve orbital cartilage Fig. 28. Sepia: Eye in section 49 REPRODUCTIVE SYSTEM: Sexes are separate. Sexual dimorphism is present. The males are distinguishable by the presence of hectocotylized arm. The males are smaller with slightly longer arms. MALE REPRODUCTIVE SYSTEM: The male reproductive system consists of a large oval testis situated in the aboral region of the mantle cavity enclosed in a capsule. The capsule leads into a narrow coiled tube, the vas deferens or spermiduct. The vas deferens opens into an elongated seminal vesicle having grooves and ridges on its inner surface. The seminal vesicle expands into a wide sperm sac or Needham’s sac (Fig. 29). A glandular prostrate gland is connected with the terminal part of the seminal vesicle close to the Needham’s sac. The sperm sac leads into a narrow, cylindrical penis which opens into the mantle cavity by the genital aperture situated on a papilla. The sperms which are formed in the testis pass through the vas deferens into the seminal vesicle where they are rolled up into narrow bundles called spermatophores by the action of grooves and ridges (Fig. 30). The spermatophore is enclosed in a chitinous capsule. One end of the spermatophore contains a spring apparatus for rupturing the chitinous wall and liberating the sperms. All the spermatophores are stored in the Needham’s sac. The hectocotylized arm takes out the spermatophores , and keeps them for sometime in the suckers and places them on the body of the female during copulation. FEMALE REPRODUCTIVE SYSTEM: A single, large, ovoid ovary is situated in the aboral region of the mantle cavity enclosed in a capsule also called the coelomic sac. The capsule leads into a wide, thin walled tubular oviduct which runs orally and opens into the mantle cavity by the genital aperture to the left of the anus. A pair of large, flat, oval bodies, the nidamental glands are present on the sides of the ink duct. Each nidamental gland is oval in shape and their secretion helps the eggs to adhere together. A pair of smaller accessory nidamental glands of unknown function is present around the anterior ends of the nidamental gland proper. They open into the mantle cavity by numerous pores present on their surface. COPULATION, FERTILIZATION AND OVIPOSITION: During copulation, the male inserts one of its arms (the fourth left arm) into its own mantle cavity, and the spermatophores from the genital duct adhere to its suckers. This arm is then inserted into the mantle cavity of the female and the spermatophores are deposited in the part of the funnel modified as bursa copulatrix. This arm modified for the transference of sperms is known as hectocotylized arm. The spermatophores (Fig. 30) now burst by the spring apparatus and sperms are released. At the same time ripe ova are passed out of the ovary one by one and as they pass the funnel and fertilization takes place. Fertilization is thus, internal. The secretions of the nidamental glands cover the eggs and cements them into a mass. Eggs are laid singly. The eggs laid by the female Sepia are large, pear shaped and heavily laden with yolk (Fig. 31). Yolk provides nourishment for the developing embryo. They get attached to weeds or any suitable substratum by a stalk. 50 DEVELOPMENT: The development of Sepia is direct. The young ones that emerge out of the egg resemble the adult except that they are smaller in size and are not sexually mature. genital orifice vas deferens seminal vesicle penis caecum genital coelomic capsule testis sperm-sac prostrate Fig. 29. Sepia: Male reproductive system spring apparatus mass of sperms Fig. 30. A spermatophore of Sepia eggs Fig. 31. Sepia : A cluster of eggs 51 ANCESTRAL MOLLUSC The mollusc are thought to be pre-cambrian in origin, but their ancestors or animal group from where they have descended is still debatable. The main reason being their highly diverse classes with virtually no converging form. It is thus presumed that they have descended from a hypothetical ancestor known as archimollusc which were probably very primitive and from which all the present and extinct groups can be derived with some modification. The archimollusc were bilaterally symmetrical, sluggish animals that crawled about on rocks and hard surfaces. They had a dorsoventrally flattened and ovoid body with an anterior mouth and a posterior anus. A distinct head with a pair of tentacles and a pair of eyes was anteroventrally situated. The muscular ventral surface was flattened to form a creeping sole or foot (Fig. 32). The foot was abundantly supplied with mucus gland cells that produced mucus over the hard substratum. The animal attached itself to the substratum and moved over the slime secreted by it. The dorsal surface was covered by an oval, convex, shield like shell that acted as a support for the mantle cavity. The shell also protected the underlying internal organs or visceral mass and acted as an anchor for the shell muscles which entered the foot and allowed the shell to be pulled down over it for protective purposes. The primitive shell was probably made up of conchiolin (a protein), but lacked calcium carbonate (modern shell of mollusc is made up of conchiolin and calcium carbonate). The underlying epidermis called the mantle or pallium secreted the animals shell and the most active secretion was located mainly in the mantle edge. The primitive mantle cavity may have been a marginal groove between the mantle and the foot. Within this mantle cavity was a pair of respiratory organs, sensory osphradia and openings from a pair of nephridia and a median anus (Fig. 32). 52 shell ventricle mucous cord gonad digestive gland aorta auricle mantle mantle nerve ring nephrostome nephridium mouth anus radula mantle cavity nephridiopore osphradium intestine style sac pedal nerve cords protostyle foot visceral nerve cords Fig. 32. A generalized mollusc 53 gills Respiratory organs consisted of several pairs of gills or ctenidia (ctene means a comb). Each ctenidium consisted of a central axis, bearing a row of triangular leaflets or filaments on each side and were provided with mucus cells and cilia(Fig. 34). Each filament had an upstream frontal margin on one side and a downstream abfrontal margin on the opposite side (Fig. 35). The ctenidia was referred to as bipectinate in these archimollusc because of the presence of two rows of gill filaments. The ctenidia divided the mantle cavity into two chambers (Fig.33): 1. Ventral inhalant or infrabranchial chamber. 2. Dorsal exhalant or suprabranchial chamber. The exposed gill surfaces were known as lamellae- the frontal lamellae facing the inhalant chamber and the abfrontal lamellae facing the exhalant chamber(Fig. 35) Each ctenidium was held in place by a long efferent membrane attached to the floor of the mantle cavity and a short afferent membrane attached to the roof. Water entered the lower part of the mantle cavity from the posterior, passed upward through the gills and then moved back posteriorly to be finally given out of the mantle cavity(Fig. 35). The propulsion of water current through the mantle cavity was brought about by the beating of the cilia present on the ctenidial filaments. There were three types of cilia present on the gill filaments: 1. Lateral cilia 2. Frontal cilia present on the frontal margin of the filament. 3. Abfrontal cilia present on the abfrontal margin of the filament. It was the lateral cilia which generated the respiratory current through the mantle cavity for the exchange of gases. The frontal and abfrontal cilia moved mucus and particles and kept the ctenidia free of particulate matter like sediment and detritus that might otherwise would have interfered with the respiratory flow of water between the filaments. Sediments brought in by respiratory current were trapped on the gills by mucus secreted by the gill epithelium. The frontal cilia generated a current moving away from the gill axis towards the tip of the filament and there by carrying these sediments along the edge of the filament to be finally swept out by the exhalant current as pseudofeces. Pseudofeces were thus strings of sediments entangled in mucus. The abfrontal cilia helped in the transportation of these pseudofeces to the exhalant water current. Two, large mucous secreting glands called the hypobranchial glands were present on the roof of the mantle (Fig. 33). Their function was to further trap the particles before they left the mantle cavity and prevent the clogging of the ctenidia. The ctenidium was richly supplied by blood vessels – an upper afferent and a lower efferent blood vessel (Fig.33). The afferent blood vessel carried unoxygenated blood into the gills and the efferent vessel carried oxygenated blood from the gills to the heart. Like the present day mollusc, the archimolluscs were also grazer of fine algae and other small sessile organisms growing on rocks. 54 rectum mantle cavity Hypobranchial glands shell gill filament mantle abfrontal margin afferent branchial vessel frontal margin inhalent chamber efferent branchial vessel foot exhalent chamber Fig. 33 T. S. through the body of the generalized mollusc 55 gill axis interfilamentary space filaments skeletol rods Fig. 34. Gill of generalised mollusca abfrontal cilia abfrontal lamellae filament axis frontal lamellae lateral cilia frontal cilia afferent vessel efferent vessel skeletol rod axis Fig. 35. T. S. through the gill of primitve mollusc 56 The digestive system consisted of an anteriorly placed mouth. The mouth opened into a buccal cavity lined by chitin. The posterior wall of the buccal cavity evaginated to form the radular sac which contained the unique molluscan feeding organ, the radula on its floor. The radula had numerous chitinous teeth on a membranous belt called the radula proper. The radula apparatus consisted of an elongated, cartilaginous base called the odontophore(Fig. 36). The odontophore was a broad tongue like ribbon which could be thrust out of the mouth and pulled back by complex musculature (the protractor and the retractor muscles). Not only the odontophore, but also the radula could move over the odontophore to some extent. The movement of the odontophore and the radula over the substratum brought about the scraping of the food particles, bringing it back into the buccal cavity. Radula thus functioned as a scraper and a collector. However, there was a wear and tear of the radular teeth caused by the scraping. This gradual loss of the radular teeth and membrane at the anterior end of the ribbon was compensated by a continuous secretion of the new ones at the posterior end (refer Pila for a detailed account). A pair of salivary glands opened into the anterior dorsal wall of the buccal cavity. These glands secreted mucus which lubricated the radula and entangled the ingested food particle in the mucus string and passed it into the oesophagus from where it moved posteriorly into the stomach. The stomach was complex, pear shaped with a broad anterior hemispherical part into which opened the oesophagus and a tapered posterior end called the style sac leading into the intestine. A pair of highly branched digestive glands also called digestive caeca, or hepatopancreas, or digestive diverticula, or liver opened into the anterior part of the stomach by ducts. The anterior region of the stomach was lined with chitin, except for a ciliated, ridged sorting region and the entrance point of the ducts of the digestive glands. The contents of the stomach were rotated by the style sac cilia which pulled the mucus food string from the oesophagus into the stomach. The rotating mucus mass was called the protostyle. The size and consistency of the particles within the mucus string varied greatly. The chitinous lining of the anterior part of the stomach also called the gastric shield protected the wall from damage by sharp surfaces (Fig. 37). The rotating protostyle brought its contents repeatedly in contact with the ciliated stomach wall, where the acidity of the stomach (pH 5-6) lowered the viscosity of the stomach thereby freeing the contained particles. These particles were then swept against the large sorting region where they were graded on the basis of their size. Lighter fine food particles were passed into the digestive gland while the heavier, large rejected particles were either passed into the intestine directly or remained in the ptotostyle. The long coiled intestine functioned largely in the formation of faecal pellets which were then passed into the rectum, to be finally expelled out into the posterior margin of the mantle cavity. Anus was dorsally present in the mantle cavity near the exhalant current. The waste was finally swept out of the body through the exhalant current. Digestion was both extracellular and intracellular. The digestive enzymes secreted by the gastrodermal cells of the digestive glands were transported to the stomach, where extracellular digestion occurred. The partially digested food was then passed into the digestive glands where intracellular digestion took place. Absorption of the digested food also took place in the digestive glands. 57 odontophore radular sac salivary gland oesophagus buccal cavity radular membrane radular retractor radular teeth mouth odontophore retractor radular protractor odontophore protractor Fig. 36. Archimolluscan radula 58 salivary glands oesophageal gland oesophagus digestive caecum duct duct of digestive caecum gastric shield sorting shield minor typhlosole major typhlosole style sac intestinal groove intestine Fig. 37. A primitive molluscan stomach 59 Coelom was represented by pericardium and the cavity of the gonads. The primitive blood system was probably similar to the modern molluscs i.e. of the open type. The heart consisted of a single muscular median anterior ventricle and a pair of posterior auricle. A single aorta was given off from the ventricle. The auricles drained blood from the ctenidia and passed it to the ventricle, which pumped it anteriorly through a single aorta. The aorta branched into smaller blood vessels that delivered the blood into tissue spaces and sinuses of the hemocoel. The blood after passing through the nephridia and the gills was returned back to the auricles from these sinuses. The respiratory pigment in these archimollusc was haemocyanin. Associated with the pericardium was a pair of renal organs. A pair of tubular metanephridia called kidneys originated from the pericardium and discharged the waste into the mantle cavity through the nephridiopore. The waste was finally given out of the body along with the exhalant current. The ducts of the renal organs acted as reno-genital ducts discharging both renal and gonadial products. The nervous system was very simple, consisting of a central circumoesophageal ring (formed by cerebral and pedal ganglia along with their connectives and commissures), from the underside of which two pairs of longitudinal nerve cords extended posteriorly. The ventral pair called the pedal cords innervated the foot while the dorsal pair called the visceral cords ran laterally and innervated the visceral mass and the pallial organs. The nervous system was said to be tetraneurous because of the presence of four nerve cords. In these primitive molluscs, there were transverse connectives between the nerve cords, giving a ladderlike appearance to the system. Sense organs included tentacles, a pair of statocyst in the foot, a pair of eyes, and osphradia. Osphradia were chemoreceptors present on the posterior margin of each afferent gill membrane. They monitored the water entering the mantle cavity. The archimollusc had a pair of anterior dorsolateral gonads on each side of the coelom. It is not clear whether the sexes were separate or not. When ripe, the sperms or eggs broke off into the coelomic cavity from where they were transported to the outside by the ducts of the nephridia which also served as gonoducts. Fertilization was external in sea water. Spiral and holoblastic cleavage was a characteristic feature of these archimollusc. The gastrula developed into a free swimming, planktonic trochophore larva (refer Annelida). The trochophore larva was similar to the trochophore larva of annelids. In most of the classes of mollusc, the trochophore larva developed into a much advanced larva known as veliger larva. The veliger larva possessed two lateral ciliated lobes known as velum which helped the larva in swimming and feeding. Foot, shell and mantle made their appearance after which the larva settled at the bottom and metamorphosed into an adult. 60 SHELL IN MOLLUSCA The shell is one of the most important diagnostic feature of the phylum mollusca. It is present in almost all the molluscs but varies greatly. The shell may be present outside the body as in gastropods, bivalves and chiton. Internal shell is present in most cephalopods. Some gastropods like Aplysia, have an internal shell which is thin, flexible and plate-like, reduced, and completely covered by mantle.. In some molluscs like Nacticidae, the shell is partly external and partly internal. In some cephalopods and Nudibranchia, the shell is totally absent in adult stage. The Aplacophorans too do not have a shell. Their mantle contains tiny calcareous spicules that hardens to form a protective covering. Shell is even absent in some of the gastropods like the Eolis, the sea slug; Doris, the sea lemon and some others. Limax. the grey slug too has a thin, reduced shell, partly horny and partly calcareous in nature. Class Monoplacophora: Monoplacophorans have a single bilaterally symmetrical shell that varies in shape from a flattened, shield like plate to a short cone. Class Polyplacophora: Shell of Polyplacophorans like Chiton is composed of eight overlapping shell plates or shell valves (Fig. 38). The shell valves are either completely covered by mantle as seen in Chiton or partially covered as is found in Katharina. The eight shell plates are encircled by the mantle margin like a girdle. The shell is held in position by a large wing like insertion plate which extends laterally and anteriorly from each valve into the surrounding mantle below the preceding valve. Each plate except for the first and the last are somewhat triangular and similar in structure. The first and the last ones are smaller and hemispherical in outline. A pair of pedal retractor muscles extend from the foot to each of the valves. Each shell plate is made up of two layers: an outer tegmentum composed of protein conchiolin and calcium carbonate and a lower, thicker layer called the articulamentum composed of only calcium carbonate. The tegmentum contains the sense organs called the esthetes. The articulamentum forms the insertion plates and keeps the shell in position. It is the tegumentum which is visible, the articulamentum remains hidden either by the tegumentum or the mantle. Class Gastropoda: The shell in most gastropods is made up of a single piece (univalve shell). The univalve shell is an elongated cone which becomes twisted, the spires descending in a screw-like manner from the apex to the aperture. The shell was originally a simple cone e.g. Patella, the true limpet, but due to the increase in size of the visceral hump, the shell becomes twisted in order to accommodate the visceral organs. Instead of an elongated cone, the shell can either form a flattened spire as in Polygyrantia or a globular spire as in Pila. Usually the spire is more or less obliquely coiled around a central axis. There are two types of spirals encountered in gastropods. • DEXTRAL OR CLOCKWISE: The spiral is right handed round the central axis. When the shell of a mollusc like Pila is held in the observers hand with the apex upwards and the aperture turned towards the observer, then the aperture of Pila will be towards his right. This is found in majority of gastropods. • SINISTRAL OR ANTICLOCKWISE OR COUNTERCLOCKWISE: It does not occur very commonly, but may be regarded as the normal type of shell in some of the molluscs. The spiral is left handed around the central axis. In this type of spiral, when the shell of a mollusc is held in the same manner as above, then the aperture will be towards the left of the observer. The direction of spiral does not show uniformity. In some of the molluscs, the sinistral type of shell may be present in the embryonic stages while the adults may show dextral type of 61 shell. Abnormal growth of shell does occur in nature which may change the normal organization of the animal. In many gastropods, the spiral nature of the shell is present only in the juvenile stages. In Haliotis, the shell uncoils, but remains asymmetrical. A typical gastropod shell has been discussed in details in the type study of Pila (Fig. 1). Class Bivalvia: The shell of bivalves is made up of two valves. The dorsal margin (hinge lines ) of the two valves are usually united by a hinge ligament made up of conchiolin. The ligament is secreted by the mantle. Two adductor muscles extend from one valve to the other and the space between them is called the gape. The two valves can be opened or closed by adductor muscles and the hinge ligament. The hinge ligament is responsible for keeping the valves open. When the valves are open, the ligaments are relaxed. Contraction of the adductor muscles are responsible for closing the valves. When the adductor muscles contract, the ligaments are under tension and are compressed thus causing the valves to close. When the adductor muscles relax, the ligaments return to their original shape thereby once again opening the valves. The two valves of the shell may be alike, or nearly so, and is said to be an equivalve shell e.g. Unio, Mytilus etc.(Fig. 42, 45). Solen has a long, narrow, straight and thin equivalved shell (Fig.43). However, in some forms the two valves are markedly dissimilar and is said to be inequivalve shell e.g. Pecten and Oyster. In Pecten the right valve is flat while the left one is convex. In Oysters (Fig. 44), the right valve is concavo-convex and permanently attached to the rocks while the left valve forms a flat lid. The valves may be inequilateral (a line drawn from the umbo to the ventral border divides them into two unequal parts) or equilateral as in Pecten(Fig.46). The valves of the shell vary greatly in color and form. They may have smooth, rough or ribbed surface with wavy margins. The inner surface of each shell valve bears two sharp blade like teeth. The teeth are separated from each other by grooves. The teeth of one valve fits into the groove of the other valve and thus preventing them from slipping out of their position. The hinge teeth keep the shell valves properly closed, and the edges of the shell fix tightly to keep the animal safe within it. The shape, number, and development of these teeth varies in different species. In Anodonta, Pecten and Mytilus they are totally absent. The hinge teeth may have been secondarily lost in Anodonta. The outer surface of the shell bears a number of concentric lines running parallel to the free margins of the shell and are called the lines of growth. The lines start from a small elevation, the umbo situated near the anterior end of the hinge line. Thus, umbo is the part of the shell first formed in the young animal and is the oldest part of the shell. As the animal grows, new layers are secreted beneath it. Each new layer projects a little beyond its predecessor, thus giving rise to the lines of growth. Thus, the shell is thickest at the umbo and becomes progressively thin towards the margins. The distantly spaced lines show rapid growth while the closely placed lines represent slow growth of the valves. The age of the animal can be found out by counting these lines (Fig. 42). However, the shell is greatly modified in burrowers. The shell may be weak and brittle with additional calcareous plates in between the two valves e.g. Pholas. The Pholas (rock borers) have an elongated shell having broad anterior serrated margins for drilling (Fig.52). Teredo, the shipworm burrows in wood and have very small shell-valves at the anterior end of the body. They are used for making burrows in the wood. Class Scaphopoda: The shell of scaphopoda is an elongate cylindrical tube usually shaped like an elephants tusk. Both ends of the tube are open; possessing a wide anterior aperture 62 and a narrow posterior aperture. The anterior aperture is larger than the posterior one. Some scaphopod shells are slightly curved towards the anterior end (Fig.53). Class Cephalopoda: Cephalopods have a single shell that varies in form. The shell may be internal (endocochleate cephalopods) or external (ectocochleate cephalopods), divided or undivided into chambers by septa. Nautilus has an external shell coiled like a watch spring and internally divided into a series of chambers by septa. It is perforated in the middle by a spiral tube known as siphuncle (Fig.39). The oldest and the smallest chambers are at the posterior end. The animal is present in the largest anteriormost chamber known as the body chamber or dermiciliary chamber. The chambers contain air so that the shell can float and the animal can swim easily. The siphuncle functions as an osmotic pump to remove liquid from the chambers and replace it with gas. The shell chambers and siphuncle enables the animal to maintain neutral buoyancy (Fig.40). The shell of Nautilus is planospiral with the spire coiled anteriorly above the head (epigastric coiling). All living cephalopods except Nautilus are endocochleate coleoids. The ancestral coleoids had an internal shell which was made up of three parts: 1. Phragmocone which was septate and had a siphuncle. It had a hydrostatic function, providing buoyancy to the body. 2. Rostrum which partially surrounded the phragmocone giving support and protection to it. 3. Proostracum, an anterior extension of the phragmocone that provided protection to the visceral mass and provided a base for the mantle and muscle attachment. The present day coleoids have been derived from this ancestor. For example: Spirula has a spirally twisted internal shell, completely covered by mantle and divided into chambers by septa. The rostrum and proostracum are lost in them. The phragmocone became coiled and the siphuncle is peripheral. Sepia (refer type study) has an internal shell in the form of a flat plate (Fig.41). The major part of the shell is the phragmocone. The rostrum and proostracum are reduced in Sepia. True siphuncle is lacking. It is replaced by a vascularized epithelium that functions for buoyancy control. In squids (Loligo), the shell is reduced to a chitinous pen located dorsally along the midline. The pen is derived from the proostracum and is responsible for stiffening the body. The phragmocone and rostrum have disappeared. Siphuncle too is absent. Octopus has a vestigeal shell while it is absent in male Argonauta. Female Argonauta has a thin external shell. It is delicately coiled and is not divided into chambers. Unlike the other mollusc, the shell of female Argonauta is also not secreted by the mantle, but by a pair of specially modified arms. Shell in females is used as a brood chamber for the protection of the eggs. The evolution of internal shell is an adaptation for powerful swimming. 63 shell plates Fig. 38 Shell of Chiton 64 shell septum Fig. 39 Nautilus within shell septa empty Chamber septal necks body chamber Fig. 40 Nautilus shell in section 65 pro-ostracum phragmo-cone shell margin spine Fig. 41 Shell of Sepia hinge ligament umbo lines of growth foot shell valve Fig. 42 Shell of Unio foot shell valve Fig. 43 Shell of Solen 66 hinge umbo shell valve finger like projections Fig. 44 Pearl Oyster shell valve hinge ligament lines of growth umbo mantle edges foot byssus Fig. 45. Mytilus 67 wing hinge umbo shell valve radiating striations Fig. 46 Shell of Pecten shell aperture of shell Fig. 47 Shell of Cowrie 68 MICROSCOPIC STRUCTURE OF SHELL (Fig.48): Shell is composed of three layers: 1. OUTER THIN PERIOSTRACUM: It is a translucent horny layer composed of a chitinous substance, the conchiolin. It is a protective layer that protects the underlying layers from erosion by the carbonic acid of water. The periostracum is worn away at the umbo, leaving the ostracum exposed. It is only this layer that is externally visible. 2. OSTRACUM OR PRISMATIC LAYER: The middle thick layer made up of calcium carbonate or aragonite crystals altering with thin layers of conchiolin. The layers are transversely arranged, at right angles to the margin of the mouth of the shell. This layer gives strength to the shell. 3. HYPOSTRACUM OR NACREOUS LAYER OR MOTHER OF PEARL: The innermost shining layer of shell made up of alternating layers of conchiolin and calcium carbonate or aragonite arranged parallel to the surface. The nacreous layer has a smooth iridescent lustrous surface and is of economic importance, used for the manufacturing of buttons. The periostracum and the ostracum are secreted by the margins of the mantle while the nacreous layer is secreted by the entire surface of the mantle. The ostracum and the hypostracum form the insertion plates. The shell may be of different colors. The color is because of the pigments present in the periostracum or pigments deposited in the calcareous layers during the secretion of the shell. COMPOSITION OF SHELL: The shell is mainly composed of carbonate of lime with traces of phosphate of lime and an organic base allied to chitin known as conchiolin. The proportion of carbonate of lime is about 88-99% and of phosphate of lime is 1-10%. In addition, carbonate of magnesium and traces of silica have also been detected in the shell. Conchiolin constitutes a membranous framework for the shell. 69 periostracum shell prismatic layer nacre nacre secreting cells of mantle connective tissue mantle cilliated epithelium Fig. 48. Unio. A part of shell and mantle in T. S. 70 FORMATION OF SHELL: Opinions differ regarding the formation of shell. Two main views have bee putforth: Bowerbank and Carpenters view: According to them the formation of shell is an organic process. It grows in the same manner as the teeth and bones of higher animals. Reaumur- Eisig view: According to them shell is a by-product of excretion and is deposited like the cuticle of Arthropods. The shell is formed by a number of calcareous particles held together by some kind of animal glue. The margin of the mantle is the main source of the deposition of the shell, while the rest of the mantle helps to thicken the innermost layer. The shell depositing cells are present in every part of the mantle. The epithelial cells of the margins of the mantle separate out the carbonate of lime from the circulating blood which becomes crystalline or granular and hard on exposure and is deposited as the shell substance. Deposition of shell substance is not a continuous process. The umbo part is secreted first and the subsequent layers are added in slow and rapid manner alternately in concentric lines, the lines of growth. FOOT AND ITS MODIFICATIONS Foot is a characteristic feature of the phylum Mollusca adapted for locomotion, but can take over the function of reproduction, defence and capturing of the prey. It is regarded as the remnant of the dermo-muscular tube of the ancestral form whose dorsal side becomes degenerated and the ventral side becomes greatly modified for creeping. Foot is innervated by the nerves of the pedal ganglion. Molluscs occupy different niches and thus the foot becomes greatly modified in shape, form and function. The simplest and primitive form of foot in molluscs is considered to be a broad, ventral flat foot adapted for creeping movement. However, foot is absent in some of the molluscs e.g. Ostrea ( class Bivalvia). It is very much reduced in Teredo. True molluscan foot is also absent in aplacophorans. In Neomenia (class Aplacophora), a ventral groove with ciliated ridge serves as a locomotory organ (Fig.50). Reduction or complete loss of foot is also seen in parasitic gastropods. In sedentary and sessile gastropods e.g. Bathysciadium, Vermetus etc. foot is reduced and the ventral sole becomes an effective sucker for attachment to the substratum. FOOT AS A CREEPING OR CRAWLING ORGAN: Neopilina is one of the very few living members of the class Monoplacophora that possess a disc-like foot with a flat creeping sole that occupies the entire ventral side (Fig.49). Polyplacophorans like Chiton (Fig.57) too have a broad, muscular flat foot on the ventral side that helps the animal to adhere to the substratum and creep. The foot in Gastropoda usually has an elongated, flat, ventral sole for creeping (Fig.51). Very fine bundles of muscles called as the tarsos muscle are present on the sole of the foot and are responsible not only for locomotion but also for capturing the prey. In higher gastropods, the foot is differentiated into three parts by transverse grooves: a small anterior propodium, large, middle mesopodium and small posterior metapodium(Fig.51). The mesopodium is the locomotory region. Metapodium bears the operculum. However, the propodium is either reduced or absent in most prosobranchs. The foot is variously modified in different gastropods e.g. anterior margin of the foot may form a small tactile process which helps in creeping as is seen in Trochus or the foot may project as a small fleshy process e.g. Turbonilla(Fig.51). Triton has a contractile foot. 71 Patella, the true limpet (prosobranch) is a sluggish animal which creeps slowly when required. It remains firmly attached to the rocks and other objects on the sea shore with the help of the foot. Thus, Patella has a well developed ventral foot with a broad, flat creeping sole to move over the rocks. The foot is surrounded by horse-shoe shaped muscle band known as the pallial muscles for attachment (Fig.51). Acteon and Cypraea possess a foot with very large, broad creeping sole. They move by producing waves of contraction on the foot. Limax, the grey slug is a terrestrial gastropod and bears a long foot on the ventral side with a creeping sole and a pair of longitudinal grooves. The pedal gland just behind the head secretes slime or mucus on which the animal creeps. Like the Limax, Helix, the land snail, too, is a terrestrial gastropod found in moist shady places like the gardens. It creeps very slowly on a self secreted tract of mucus. Haliotis is a herbivorous gastropod that lives attached to rocks and moves very fast, 5m. in a minute. They possess a very large foot that dorsally bears a fold of the skin, the epipodium. The epipodium is an appendage of the foot that gives off numerous sensory epipodal tentacles. Monodonta, and Trochus are also examples of gastropod possessing epipodia as appendages of the foot and beset with tactile papillae that assist them in creeping (Fig.51). Creeping commonly occurs by waves of muscular contraction that sweeps over the foot. The sole of the foot is firmly attached to the substratum by the mucus which is sticky and gelatinous. In most of the gastropods, mucus is secreted by the gland cells present in abundance on the sole of the foot. However, in pulmonates, mucus is secreted by the pedal gland situated on the ventral foot. As the muscular wave of contraction is passed forward, the gelatinous mucus liquefies in this region thus allowing the foot to move forward. The muscular contractions may be direct, i.e. passing forwards along the foot from behind or may be retrograde, i.e. passing backwards (anterior to posterior) as is seen in most gastropods, particularly in prosobranchs (refer Pila). The direct waves involve the contraction of the longitudinal and the dorsoventral muscles (from the posterior to the anterior end), while the retrograde waves involve contraction of the transverse muscles followed by the contraction of the longitudinal muscles (from the anterior to the posterior end). The waves may be monotaxic, i.e. a single series of waves traversing the foot, or ditaxic, i.e. distinct coordinated waves on the right and left halves of the foot is responsible for the snails movement. The amplitude of the waves however, is small. In some Gastropods, the foot shows partial regional modification e.g. in Pirulus only the left part of the foot acts as the creeping organ. In Atlanta, the posterior part of the foot is altered into a sucker. In Bullia, the foot is very peculiar as it encircles whole of the body. In Natica, the propodium forms a semicircular flap and is demarcated from rest of the foot by deep transverse grooves. The metapodium is provided with lateral parapodia in Natica (Fig.51). The highly glandular foot of Conus with a long backwardly bent siphon acts as an efficient creeping organ. Caecum is a peculiar gastropod that creeps entirely by the action of the cilia present on the sole of the foot. Bivalves like Anodonta and Unio have a laterally compressed triangular and ploughshare like or hatchet shaped muscular foot for crawling and ploughing through mud (Fig.42). Foot is actually the extension of the visceral mass. It is antero-ventral in position and ends ventrally in a keel. During locomotion, Unio protrudes out its foot between the two valves of the shell and burrows like a plough-share through the mud or sand by the contraction of the pedal protractor muscles. This results in the flow of blood into the foot which swells up and becomes turgid and anchors the foot in sand or mud. The blood is prevented from returning to the body by sphincter muscle. Now the retractor muscles contract and the body of Unio 72 moves forward. During this action, the blood is forced out of the foot and it becomes narrow. Thus, by alternate contraction and extension of the foot, the animal slowly creeps or burrows. In Bivalves like Nucula, the foot shows a primitive form, possessing a flat ventral sole which is not for creeping. The foot is laterally compressed and directed downward. The two sides of the foot can be folded together producing blade-like edge which is thrust into the sand or mud. The ventral sole then opens to serve as an anchor and the body is drawn down into the substratum. In lepton and some other Bivalves, the foot secondarily acquires a large free extremity with a creeping surface. FOOT AS A BURROWING ORGAN: Dentalium ( class Scaphopoda) has a small conical, highly extensible foot adapted for digging and burrowing. The foot in Dentalium and other scaphopods is equipped with pedal haemocoel which acts as a hydrostatic skeleton for extending the foot into the sand. In Siphonodentalium, the foot terminates in a retractile disc having papillae on the margins and is well adapted for digging like Dentalium (Fig.53). The foot In Pholas (class Bivalvia) foot assumes a short and blunt form, adapted for its burrowing habit (Fig.52). The mantle is closed except for a small pedal aperture through which the small foot can be protruded out to grip the end of the burrow. The foot in Anodonta and Unio can perform the function of burrowing in addition to creeping (already discussed above). Solen and Ensis (class Bivalvia) have a large, cylindrical and powerful foot that protrudes out from the anterior end of the shell and is used as a burrowing organ (Fig.43,52). The Solen burrows like Unio, but very quickly. They can dig about 30cm. in sand in a few seconds. They can also swim forward in jerks by suddenly withdrawing the foot and squirting out water through the siphons. In fast burrowers like Tellina, the foot is large and can be stretched out into a wide sheet of muscle as thin as a blade of a knife as it burrows in the sand. In Mya (soft-shell clam) the foot is feebly developed and is used as a weak burrowing organ (Fig.52). Sigaretus (class Gastropoda) is a burrower and the propodium is well developed and sharply marked off from the mesopodium to act as a digging organ (Fig.51). A dorsal flap-like fold of foot covers the head as a protective, fleshy shield. Tenebridae, a burrowing gastropod extends the anterior extremity of the foot with flow of blood, thus forming an anchor as is seen in bivalves and scaphopoda. In gastropods which crawl about in wet sand e.g. Oliva, the propodium is circular or disc-like modified for burrowing. FOOT AS THE LEAPING ORGAN: In Trigonia and Cardium (class Bivalvia) the foot is bent upon itself and when the foot suddenly straightens it violently lifts the animal off the ground in a powerful leap(Fig.52). In Trigona the foot is compressed antero-posteriorly as an elongated keel. Mytilus (class Bivalvia) has a cylindrical foot that acts as a spring tail. Mytilus remains attached to the substratum by a bunch of byssus thread (Fig.45). In gastropods like Rostellaria, the foot is reduced, and the operculum becomes sharp like a dragger. The metapodium is not sharply marked off from the rest of the foot. As the animal digs into the sand, the reduced hook-like foot becomes flexed pulling the animal forward in short sharp leaps (Fig.51). 73 FOOT AS A LOOPING ORGAN: In Pedipus, the propodium is sharply marked off from the rest of the foot by a groove that helps in the looping movement. FOOT AS A SWIMMING ORGAN: In many Molluscs, the foot becomes modified as a swimming organ. The foot of the swimming gastropods show several adaptive modifications. The common adaptation for swimming is the development of parapodia as lateral lobes of the foot which acts as fins, for example, in Oxygyus and Atlanta (class gastropoda), the propodium is fin like, the mesopodium bears a sucker and the metapodium is produced into a laterally flattened swimming lobe. The shell is laterally compressed and produced into a sharp keel. Carinaria too is a pelagic, marine gastropod where the foot is modified into a fin like swimming organ, its sole forming an adhesive sucker. The shell and metapodium are reduced in Carinaria (Fig.51). In Pterotrachea, the metapodium loses the operculum and becomes reduced to a short, filament like tail for swimming. Creseis , the sea butterfly is a small marine gastropod adapted for pelagic life. The foot develops a pair of large lateral parapodia that functions as fins for swimming. Spiratella is a pelagic gastropod that swims by flapping the parapodia. The foot develops anteriorly into highly muscular wing like projections called the parapodia while the posterior part of the foot is greatly reduced. Aplysia possesses a broad, muscular and ventral foot that bears a pair of lateral fleshy fan-like outgrowths, the parapodia which help in swimming. The posterior part of the foot is usually adhesive and gives rise to a very short and distinct tail. Swimming in Aplysia is achieved in short bursts by the rhythmic waves which pass along the parapodia (Fig.51). In shelled pteropods or sea-butterflies, broad wing like or oar-like parapodia confined to the anterior part of the body, form the largest part of the foot. For example, Limacina swims spirally upwards using the parapodia as oars and drops down by holding them motionless over the head (Fig.51). Naked pteropods are the fastest swimmers among gastropods and rapid sculling motion is brought about by the parapodia which are present on the ventral side. The foot is greatly reduced in Pecten, a marine bivalve. It swims by clapping of the shellvalves. The most noticable transformation of the foot as swimming organ is observed in Cephalopods. Foot is partly modified into a funnel lying below the head and partly into sucker bearing arms or into tentacle bearing lobes surrounding the mouth. The number of oral arms varies in different forms. They are eight in Octopus, ten in Loligo and Sepia. The fifth pair of arms in Loligo and Sepia are long, slender and prehensile and are called tentacles. The tentacles have expanded tips and are partly (Loligo) or fully (Sepia) retractile within special sacs present at their bases. They bear suckers only at their expanded ends (Fig.54). Loligo is very active and can swim both forward and backward with the help of fins and funnel. Octopus, the devil-fish inhabits all the seas and closely resembles squids and cuttlefish. They possess eight arms which are alike and slightly webbed at the bases. Each arm bears two rows of sessile suckers. Fins are absent in them. Both Sepia and Octopus dart quickly by forcing out jets of water through the funnel (Fig.54). Sepia also swims about gently by undulating movements of the fins (refer locomotion in Sepia). 74 Amphitretus is an inhabitant of deep sea and the arms are united by a web-like fold, the interbrachial membrane. Argonauta, the paper nautilus inhabits warm sea. The female Argonauta usually keeps her trunk in the shell, and when disturbed completely withdraws into it. She sails in the shell, propelling it with her arms. She has one of her arms flattened and expanded at the ends for the secretion and support of the shell. The male Argonauta is a coinhabitant of the females shell. FOOT AS AN ORGAN HELPING REPRODUCTION: In some cephalopods, during the breeding season, one of the arms in males is modified into an intromittent organ for the transference of sperm and is known as the hectocotylised arm (refer Sepia). In male Octopus, the third right arm is modified into a spoon-shaped hectocotylized arm (Fig.54). ACCESSORY ORGANS ASSOCIATED WITH THE FOOT: The foot in Molluscs is also a highly glandular organ. Some glands are present on the foot that help in locomotion. The secretions of the glands lubricate the passage during movement e.g. the pedal glands and the mucus secreting glands of the gastropods. In some bivalves, the adult or even the larva possesses a byssus apparatus which helps the animal in adhesion to the substratum. The foot in these animals is much reduced. The byssus gland may be considered homologous to the pedal mucus secreting glands of the gastropods. Mytilus is a common bivalve which remains attached to the rocks by byssus threads. These byssus threads are secreted by the byssus glands in the posterior groove of the much reduced foot. As the secretion of the gland passes out through the aperture, it hardens on contact with the water forming tough, bundle of silky threads which anchors the animal firmly to the substratum. The organ of Valenciennes in some cephalopod females and Van der Hoeven in Nautilus males are some of the accessory organs associated with the foot. 75 mouth palp like appendages foot gills anus shell Neopilina Fig.49 Foot in Monoplacophora mouth mouth foot groove mouth anus foot groove Chaetoderma Neomenia Fig.50 Foot in Polyplacophora 76 Proneomenia metapodium propodium mesopodium propodium metapodium Natica Sigaretus parapodium epipodial tentacles Limacina foot parapodium Trochus Aplysia metapodium foot propodium foot sucker Carinaria Bullia Patella pedal tentacles metapodium propodium operculum Rostellaria epipodial tentacles Monodonta Fig.51 Foot in gastropoda 77 operculum Valvata foot foot Cardium Tellina Ensis Pholas foot Mya Axinus Fig.52 Foot in Bivalves 78 foot foot Dentalium Siphonodentalium Fig.53 Foot in Scaphopoda 79 oral arms oral arms fin fin Loligo Sepia oral arms Octopus Fig.54 Foot modified into arms in Cephalopoda 80 GILLS AND ITS MODIFICATION Majority of the Molluscs respire by ctenidia or gills which remain well protected in the mantle cavity. However, gills are absent in some molluscs like scaphopods. In all the Molluscs, gill is a homologous organ, derived from the gill of ancestral type (archimollusc). A typical ctenidium consists of numerous trilangular leaflets or lamellae arranged in a single row running parallel to one another along the ctenidial axis. The ctenidial epithelium is usually covered by cilia which are responsible for maintaining a current of water over the gills. Ctenidium receives the venous blood from the body through the afferent branchial vein. The blood passes through the gill filaments and becomes oxygenated. The oxygenated blood is then sent back to the heart through the efferent branchial vein(refer ancestral mollusc) (Fig.35). Ctenidia can be of different types depending on their position. HOLOBRANCHIATE TYPE OF CTENIDIA: Found in Polyplacophorans. The gills are present throughout the length of the body along the margin except the head and anus. MEROBRANCHIATE TYPE OF CTENIDIUM: The ctenidia remain restricted to a particular area of the body. Depending upon the arrangement of the leaflets the merobranchiate type of ctenidia can be further subdivided into: • PLICATE TYPE: This type of gill consists of simple, flat, transversely folded projecting integumentary lamellae. In Chaetoderma, two large plume- like ctenidia arise from the cloacal wall. • MONOPECTINATE TYPE: The ctenidia consists of flattened gill filaments arranged in a single row on one side of the ctenidial axis e.g. Neopilina, Pila, Triton etc. (Fig. 10A, andFig.57) • BIPECTINATE TYPE: The ctenidium has flattened gill filaments arranged in two rows, one on each side of the central axis. The bipectinate type of ctenidium can either be equal or unequal. UNEQUAL BIPECTINATE TYPE OF CTENIDIUM: The ctenidium has flattened gill filaments arranged in two rows but the right one is smaller e.g. Haliotis, Fissurela etc. EQUAL BIPECTINATE TYPE OF CTENIDIUM: The flattened gill filaments of the ctenidium are arranged in two rows but both of them are of equal size. They become modified in bivalves. Nucula possesses short and flat leaflets. Long filamentous leaflets are present in some bivalves. These filaments may be free as in Arca, or may be joined by ciliary connectives as in Mytilus. In Unio, the ciliary junction is replaced by a membrane. In Poromya, the ctenidium becomes degenerated and is represented by a transverse partition. • FEATHERED TYPE: Characteristic of cephalopods and has been described in details in Sepia. MODIFICATIONS OF CTENIDIA: In certain Molluscs, true ctenidium is absent. The ctenidia are replaced by secondary gills or adaptive gills. 81 ANAL GILLS: Delicate leaflets form a rosette around the anus e.g. Doris (Fig.55). CLOACAL GILLS: They are present in Solenogasters and a gradual degradation of the gills is encountered. In Chaetoderma, a pair of gills are symmetrically present one on either side of the cloaca. Gills in Proneomenia is represented by a few folds of the cloacal wall. In Pterotrachea, the filamentous branchial leaflets project freely into the mantle as the mantle fold is absent in them. In most cases the mantle may act as the respiratory organ. SECONDARY GILLS OR CERATA: Highly vascular secondary gills are present on the dorsal surface of the body in Aeolis( Fig.55). They are capable of breaking off easily and also quickly regenerating. PALLIAL GILLS: Ctenidia are absent and series of adaptive gills occur in a row on each lateral side in the pallial groove e.g. Patella (Fig.56). Pallial gills may be present along with the true ctenidia e.g. Pneumoderma, Lottia. A brief description of the ctenidia in various classes will be discussed here. The structure of ctenidia and its working has been discussed at length in the type studies of Pila and Sepia. 82 rhinophore penis anus branchia Doris oral tentacle rhinophore eye spot anus cerata Aeolis Fig.55 Gastropods having Adaptive or secondary gills 83 head eye mouth foot shell gills mantle Fig. 56 Patella, Pallial gills (secondary gills) present in the pallial groove 84 CLASS MONOPLACOPHORA: The pallial groove contains three, five or six pairs of monopectinate gills. The posterior gills have lamellae usually on one side, while the anterior ones have lamellae on both the sides. The gills are supplied with blood by the afferent gill sinus to be returned by efferent gill sinus. They are not the true gills (Fig.49, 57). CLASS POLYPLACOPHORA: The bipectinate gills or ctenidia are arranged in the mantle groove. The mantle groove is divided into two narrow chambers (the inhalant and the exhalant chamber) by the presence of gills on either side. Each gill has two series of flat and oval lamellae arranged on a central axis. The lamellae are lined by ciliated epithelium. Their number is variable, varies from 688 pairs (Fig.58). The number of gills is not a constant feature of a species but increases with the increase of size of the body. Anteriorly, the girdle (mantle edge) can be raised at any point to form an inhalant aperture. Water enters through this opening, flows posteriorly in the inhalant chamber and then moves into the exhalant chamber through the gills. As the water flows between the gill filaments, exchange of gases takes place between the water and the blood. The water is finally expelled out of the exhalant chamber through a median exhalant aperture. The exhalant aperture is formed by the locally raised girdle. The structure and the blood flow of the gills are similar to the generalized mollusc. The gills may be holobranchiate type e.g. Chiton or may be merobranchiate (gills are restricted towards the posterior end of the body)e.g. Schizochiton. CLASS APLACOPHORA: Gills are either absent e.g. Neomenia or reduced to two large plume-like bipectinate ctenidia one on either side of the anus e.g. Chaetoderma. The ctenidia are supplied with blood vessels- an efferent and an afferent (Fig.59). 85 lamellae anterior edge of gill stem Fig.57 Ventral view of the gill of Monoplacophoran 86 mouth palps pallial groove gills or ctenidia girdle Exhalent chamber Inhalent chamber Fig.58 Chiton: Mantle groove showing the position of gills and the direction of water current 87 Neomenia cloaca or mantle cavity ctenidia or gills Chaetoderma Fig.59 Ctenidia in class Aplacophora 88 CLASS GASTROPODA: In the majority of aquatic gastropods, the respiratory organs are ctenidia which are present in the mantle cavity. The ctenidium may be bipectinate or it may be monopectinate. Most prosobranch archaeogastropoda possess a single pair of gills (bictenidial), the right side is usually smaller in size. The gills have a primitive bipectinate condition of the ancestral mollusc e .g. Fissurella and Haliotis. In mesogastropoda and neogastropoda due to torsion, the right gill completely disappears. A single monopectinate gill develops on the left side but is shifted to right side due to torsion. It has a single row of leaflets e.g. Pila, Triton. The structure of a typical ctenidia has been explained in the type study of Pila. In Opisthobranchs, due to detorsion and reduction of the shell, the mantle cavity becomes shorter and moves back to its posterior position, resulting in the decreased importance of ctenidia. There has been a tendency towards loss of original gill and development of secondary gills e.g. Doris and Aeolis have lost the true ctenidium and secondary gills namely anal gills and cerata respectively have developed in them (Fig.55). However, Aplysia retains a single ctenidium on the right side which is probably the left ctenidium of the higher prosobranchs. CLASS BIVALVIA: They have the most complex gills. Besides respiration, gills are also organs of nutrition and act as brood pouches. A pair of bipectinate and equal ctenidia are present one on each side of the foot. The ctenidia consist of a horizontal vascular axis, with two rows of hollow lamellate filaments, which are actually the outgrowth of the axis. However, the structure of ctenidia varies greatly in different lamellibranchiates. In the simplest state, gills are relatively smaller and situated at the back of the mantle cavity. The filaments are short, relatively few, borne transversely on the axis. The leaflets are arranged in two opposite rows. The ends of the leaflets of each gill bear tufts of loosely interlocking cilia , connecting them to the sides of the foot and to the inner surface of the mantle. The arrangement divides the mantle cavity into a large inhalant chamber below each gill and a small exhalant chamber above the gill. The ctenidial axis bears a dorsal afferent and a ventral efferent blood vessel. The function of ctenidia is primarily respiration. The food is collected by the labial palps and not the ctenidia. For example Nucula (Fig.60-1). Unlike the higher bivalves, the respiratory current generated by the beating of the lateral cilia are weak and the water enters the mantle cavity from the anterior end and leaves from the posterior end. In higher bivalves the respiratory current is stronger and enters from the posterior end. The ctenidia in these higher bivalves not only acts as a respiratory organ but also performs the function of food collection. A much developed ctenidia is found in filibranchia and eulamellibranchia. There is an increase in the number of filaments and their length. In Filibranchia, e.g. Amusium the filaments are long, thread-like and hang down ventrally from the axis into the mantle cavity. A more complex ctenidia are present in Arca (filibranchia). Each gill filament is bent upon itself to form a V- shaped demibranchs or gill laminae (Fig.60-2). Each demibranch consists of an ascending limb and a descending limb like the letter V. The adjacent filaments in the same row in both the ascending and descending limbs are held together loosely by interlocking cilia so as to form a lamella. The ascending and descending limbs of the same filament are also held together loosely by ciliary connections. Thus, the ctenidial axis bears a 89 double lamillated gill plate instead of two rows of simple filaments on each side as found in Nucula. A food groove is present on the undersurface of the gill. Mytilus possesses paired gills which are plate-like and filiform with ciliary interfilamentary junctions. The interlamellar junctions are absent in them. The adjacent filaments of each demibranch are loosely united to each other by stiff interlocking cilia. In pseudolamellibranchiata, the gills are W-shaped and folded vertically. The interfilamentary junction may be ciliary or vascular. The gills have a greater cohesion than in filibranchs. The distal tips of filaments become coalesced laterally with the mantle e.g. Pecten, Ostrea. In Eulamellibranchiata, the interfilamentary ciliary junctions are replaced by vascular crossconnections with narrow openings (ostia) between them. Interlamellar junctions also become large and vascular and partition the interlamellar space into vertical water tubes. Inhalant water enters the water tubes through minute pores or ostia in the lamellae. Gaseous exchange takes place as water moves upwards both outside and inside the water tubes and reach dorsal suprabranchial chamber from where it flows out of the exhalant opening (Fig.60-3). There are five groups of cilia: 1. Two groups of latero-frontals 2. Two groups of laterals 3. One group of frontals The constant beating of the lateral cilia generates the water current and the water enters the infrabranchial chamber through the inhalant siphon. From the infrabranchial chamber, water enters the gills through the ostia. As the water passes through the gill tubes present within the gills, exchange of gases takes place. The water finally reaches the exhalant chamber and is expelled out through the exhalant siphon. The latero-frontals are large compound cilia. They sieve out the large food particles from the inhalant current. The trapped particles mixed with mucus (secreted by the mucus secreting cells) are then carried by the frontal cilia to the ventral food groove of each demibranch. The constant beating of the cilia further carries the mucus trapped food particle to the palps where the food is further sorted out before being finally carried to the mouth In most specialized forms, Septibranchiata, the gills are reduced and represented by a muscular septum extending from the base of the foot to the mantle. The septa divides the mantle cavity into an inhalant and exhalant chambers. Septa moves up and down so that the water current enters through the inhalant siphon. Respiration takes place by the mantle in these septibranchs e.g. Poromya (Fig.60-4). 90 shell shell Visceral mass foot Outer gill filament Inner gill lamina Inner gill filament Outer gill lamina 1 . Protobranchia ( Nucula) mantle 2. Filibranchia (Arca) supra branchial chamber Inner lamellar junctions branchial septum perforations 3. Eulamellibranchia (Anodonta ) 4. Septibranchia (Poromya) Fig. 60 T. S. to show chief kinds of gills and their arrangement in different groups of bivalves 91 Inter lamellar junction Vertical gill filament ostia Chitinous rods Inner lamella Outer lamella Water tubes blood vessels Fig.61 Unio: Gill lamina in section 92 CLASS CEPHALOPODA: The members of this class possess simple, bipectinate gills on either side of the anus. The ctenidia consists of a central axis with delicate leaf-like lamellae arranged on either side of the axis in a linear manner. The cilia on the gills are absent in them and the water is pumped in and out by the coordinated activities of the mantle, funnel and the inlet valves. A typical ctenidium and its functioning has already been described in the type study of Sepia. The number of gills may vary, e.g. dibranchiates possess two gills (Sepia, Octopus etc.) while the tetrabranchiates (Nautilus) have four. MANTLE Mantle or pallium is a thin delicate covering formed by the fold of the body wall which secretes the shell. The active secretory centre is the mantle edge. Mantle encloses a cavity called the mantle cavity which contains the visceral organs. The mantle performs three functions: 1. Protects the visceral mass and the head. 2. Secretes the shell. 3. Serves as the additional respiratory organ in some molluscs. A brief description of the mantle in various classes of mollusca is given below: MONOPLACOPHORA: The mantle lies just within the edge of the shell. A mantle groove is present between the foot and the mantle edge. APLACOPHORA: The dorsal body wall extends laterally to cover the whole animal except for the foot (when present) and is called the mantle. The epidermis of the mantle secretes the cuticle. The cuticle is made up of glycoprotein and contains calcareous spicules of different shapes. POLYPLACOPHORA: The mantle covers the dorsal surface. Like the aplacophorans, the mantle of the polyplacophorans also secretes the cuticle. The cuticle may be smooth, or bear scales, bristles, or calcareous spicules. The mantle is a thick and heavy dorsal layer extending right upto the lateral margins of the body, covering partially or entirely the shell valves. The mantle extends well beyond the lateral margins of the shell valves. The outer thick and fleshy mantle edge is sometimes called as the girdle. SCAPHOPODA: The mantle has two large lateral folds that extend the entire length of the body. The lateral folds reach the ventral midline and fuse to form a tube, open at both ends. The tubular mantle encloses the whole body. The mantle extends posteriorly on the dorsolateral side and is equipped with sensory receptors. The mantle cavity is large and ventral, extending from one aperture to the other. GASTROPODA: The outer layer of the visceral mass forms a thin, delicate covering called the mantle or the pallium. The free edge of the mantle, in some gastropods, project beyond the shell and is provided with appendages like the tentacles. In primitive gastropods, the mantle edge has a 93 deep longitudinal slit which serves to eliminate the faecal matter. In Haliotis, the mantle bears a series of marginal slits while in Fissurella an apical hole is present in the mantle. In Pila, the free border of the mantle is attached to the margin of the shell and is in contact with the region above the head. The mantle is produced into highly contractile process called the nuchal lobes or pseudepipodium, the left being longer than the right and forms a respiratory siphon during aerial breathing. The mantle lining is thickened and differentiated into a glandular structure called the pallial mucus gland present between the gills and the rectum. BIVALVIA: The mantle consists of two lobes corresponding to the two valves of the shell and encloses the whole body. The two mantle lobes are fused dorsally as the mantle isthmus underlying the hinge ligament, but their margins are not fused. The two lobes are formed from the dorsal part of the body-wall. Each mantle lobe is attached to the corresponding shell valve by pallial muscles. The bivalves differ in the degree of fusion of the two mantle lobes which plays an important role in the evolution of the adult body form and its habit. The simplest fusion is by the temporary apposition of the mantle edges of the two lobes. However, in bivalves where permanent fusion of the mantle lobes occur, it involves first the union of the inner folds, followed by the middle and finally by the inner surface of the outer fold of the mantle edges. This fusion is brought about by the cilia and the tissues. In the fresh water mussel like Unio, the mantle lobes are free ventrally and anteriorly, but are fused posteriorly in the middle to form a dorsal and ventral siphon. Water current is constantly passing in by the ventral siphon and going out by the dorsal siphon, so the siphons are called as the inhalant and the exhalant siphon respectively. The inhalant siphon is wider, with papillated margin. It is formed by the coming together of the two mantle lobes. On the contrary, the exhalent siphon is narrower, smooth and is formed by the fusion of the two mantle lobes. This is an adaptation for burrowing deep in the soil. Some fresh water mussels like Anodonta have an additional dorsal opening called the supra-anal aperture above and anterior to the exhalent siphon. However, the function of this aperture is obscure. In some bivalves, like Nucula, Arca, Anomia and Trigonia, the edges of the mantle are free from each other, so that the siphons are absent in them. There is only a temporary fusion of the mantle edges, leaving two gaps: one anteriorly and the other posteriorly. The incurrent water enters through the anterior gap and traverses up in between the gill filaments into the dorsal exhalant chamber and finally leaves the mantle cavity through the posterior gap. In others, the edges of the two mantle folds are fused with each other at one or two places. Many bivalves like oyster Ostrea and the scallop Pecten, the inner fold of the mantle edge is well developed and is known as the velum or pallial curtain. Velum bears marginal sensory tentacles. These sensory structures are a common feature of the mantle border of bivalves. In the sedentary bivalves like Mytilus, the inhalant siphon is not properly differentiated. The bilobed mantle forms the exhalant siphon posteriorly. This lack of fusion of the mantle edge is because of the presence of a well developed byssus apparatus. In Solen, the mantle lobes are united ventrally to form an elongated tubular mantle cavity. Teredo has very long siphon fused together, and the mantle forms a closed tube. The siphon is non-retractile. In Mya, the elongate siphon is covered with chitinous plates and is incompletely retractile. In a few bivalves, in addition to the formation of the siphon, the ventral edges are fused to a greater or lesser extent leaving an opening for the foot to protrude. 94 CEPHALOPODA: In cephalopods the thick, muscular mantle covers the trunk. The free oral edge of the mantle fits loosely around the neck to form a collar, and thus leaves a circular opening. A muscular, conical tube called the siphon or funnel projects beyond the collar just beneath the head. The funnel represents the molluscan foot. It is through this funnel that the water is expelled out of the mantle cavity. One end of the funnel opens into the mantle cavity by a wide aperture, while the other end opens to the outside by a narrow aperture. The mantle encloses the large mantle cavity posteriorly and ventrally. The visceral mass occupies most of the space of the mantle cavity. Like any other mollusc, the epithelial lining of the mantle secretes the shell which in most of them is internal. TORSION IN MOLLUSCA The most significant feature of the prosobranchiate gastropods is torsion or twisting of the internal organs of the body. What is torsion? Torsion is a process in larval gastropods whereby the visceropallium is rotated anticlockwise through 180° from its initial position on the head foot complex bringing the posterior mantle cavity with its pallial complex, anus, rectum etc to the front of the body behind the head. Torsion takes place because of the retardation of growth on one side and active extension on the other. As has been already stated that it is the right side where the growth gets retarded and therefore the right side begins to move forward (Fig.62). It is a developmental event that occurs in larva and not in adult. Before torsion the larva is: • Bilaterally symmetrical. • The mantle cavity opens ventrally and posteriorly. • It possesses a simple straight gut with mouth placed anteriorly and anus at the posterior end. • A pair of ctenidia, osphradia and nephridiopore are posteriorly situated. • Auricle is situated behind the ventricle. • The nervous system is bilaterally symmetrical. • The bilaterally symmetrical larva undergoes torsion as a result of differential growth. After torsion: • The mantle cavity and its associated parts shift forward to take the anterior position. • Ctenidia, osphradia, and the two nephridiopores come to lie in the anterior part of the body behind the head. • Digestive system becomes U-shaped so that anus comes to lie in front near the mouth. • Auricle now becomes anterior to the ventricle. • The original symmetry of the body organisation is lost and organs of the left side move to the right side after torsion and persist. The original organs of the right side atropy and finally disappear. These include ctenidia, auricle, osphradium, hypobranchial gland and kidney. This leads to asymmetry in gastropods. • The nervous system is twisted to a figure of 8, so that the nerve ring comes to lie on top of the 8 and the visceral ganglion occupies a posterior position by crossing of 95 pleuro- visceral nerve connective. The right pleuro-visceral connective with its ganglion passes over the intestine and becomes the supra-intestinal nerve while the left connective passes under the intestine and becomes infra-intestinal nerve. This is known as chiastoneury. However, the head and foot retained the original bilateral symmetry and the shell retained the symmetrical spiral (Fig.63). In the majority of gastropods, torsion takes place in two steps: Step I: The contraction of the larval retractor muscles account for 90°rotation of the visceral hump. The mantle cavity at the end of this stage comes to lie on the right side with the foot projecting on the left side. This stage lasts for only a few hours. Step II: The torsion now is the result of only differential growth and is longer in duration. However, the actual mechanism of torsion is not known. Thomson in 1958 has suggested five possible ways in which torsion has taken place in gastropods. 1. The rotation of visceral hump by 180°is achieved by muscular contraction only. He regarded this as the original way of torsion. This mechanism was seen in Acmaea. 2. The rotation of the visceral hump by 180°is achieved in two steps: • The initial 90°rotation is caused by the larval retractor muscle. • The remaining 90°is by differential growth. The first step is however faster than the second. This is the most common way of torsion and is encountered in Haliotis, Patella etc. 3. In some gastropods like Vivipara, the complete rotation of 180°is achieved by differential growth alone. 4. In Aplysia, the differential growth is responsible for torsion, and the position of anus is halted at a region appropriate to the adult stage. 5. In Adalaria, torsion of the visceropallium is not recognisable. The different organs appear as in the post-torsional position. 96 left mesodermal band midgut cavity right mesodermal band shell Fig. 62 T.S. of veliger larva showing disproportionate growth of mesodermal cells 97 1 2' 2 4 16 6 11 10 8 3 13 15 5 14 9 12 7 Hypothetical ancestral stage before torsion Displacement of the mantle cavity to the right side of the body 1 2 3 4 7 9 8 10 12 16 5 15 14 13 Intermediate stage showing 90° torsion Complete torsion Fig.63 Torsion in Gastropods. 1. Mouth; 2. Right tentacle; 2‘. Left tentacle; 3. Right visceral loop; 4. left visceral loop; 5. Right visceral ganglion; 6. Left visceral ganglion; 7. Right ctenidium; 8. Left ctenidium; 9. Right osphradium; 10. Left osphradium; 11. Mantle; 12. Anus; 13. Ventricle; 14. Visceral loop; 15. Right auricle; 16. Left auricle. 98 SIGNIFICANCE OF TORSION: It is not clear whether torsion is of an advantage or not to the animal, or if it has any evolutionary significance, but it does take place during embryological development of gastropods. Many zoologists have postulated that that torsion represents a larval adaptation for the protection of the head. This view was first putforth by Garstang in 1928 and was supported by Ghiselin in 1966. Before torsion, the larva was an easy victim of the predators because of the posterior position of the mantle cavity in which case the foot would be withdrawn first and them the delicate vital head. The anterior position of the mantle cavity in the larva, after torsion, resulted in greater protection of the head and its associated parts by providing them with a cavity into which first the vulnerable parts and then the foot can be withdrawn at the time of danger. Ghiselin (1966) suggested some evolutionary changes to account for torsion: The shield like shell of the ancestral mollusc was of little protective value for the planktonic larva. Thus, a conical shell with a small aperture evolved which exposed only a small part of the body and at the same time served as a retreat into which the larva could withdraw its body at the time of danger. A long, straight conical shell was difficult to carry while swimming and thus the shell became spirally coiled. This planospiral shell was advantageous for the free swimming larval life but was of disadvantage for the adult crawling mode of existence. According to him, torsion was an adaptation to correct this difficulty. Rotation of the visceral mass by 180°displaced the coils of the shell to a trailing position behind the animal, thus preparing the animal for settling down for its adult mode of existence. Crofts (1937,1955), Youge (1947) and Eales (1949, 1950) also supported Garstang view. However, there were many objections to this theory: 1. In case of Heliotis, the pelagic larva undergoes torsion only through 90°, so that the head cannot be retracted first at the time of danger. Complete torsion of 180° occurs only when the larva settles down at the bottom. 2. Many pelagic larvae of lamellibranchs donot undergo torsion, but still they survive. 3. The velar cilia on some gastropod larva are under nervous control and they can be stopped even without being forcibly withdrawn into the mantle cavity. 4. The anus and nephridiopore are also anteriorly placed after torsion, thus discharging their waste on the animals head. If the larva is not benefitted from torsion, then the adult must have been benefitted as has been proposed by Morton in 1958. • • • The anterior position of the mantle cavity after torsion might facilitate the ventilation of the mantle cavity and the gills. Also, the anterior position provides the mantle cavity with water free of sediments. Torsion places the sensory osphradium anteriorly where it can sample the water entering the animal rather than sampling the one leaving the animal. Torsion may shift the shell and the visceral mass to a more advantageous position. However, the most important disadvantage of torsion is that the feces and excretory matter can pollute the respiratory current and fall in front of the mouth. To solve this problem the gastropods have only a single gill and one kidney and the inhalant current is 99 drawn from this side and after bathing the gill, it leaves by exhalant current on the other side. Also, the anus and renal aperture are located far back in the mantle cavity. The excretory waste matter and the feces are carried away by the exhalant current. DETORSION Detorsion is the complete reversal of the changes that take place during torsion. It is a characteristic feature of opisthobranchiate gastropods. A very good example where detorsion occurs is Aplysia. Detorsion occurs when the shell is reduced or lost. The visceral hump during detorsion gets completely untwisted and the pallial complex shifts back to its posterior position. The ctenidia once again take a posterior position. Their anterior position is of no advantage to them. The visceral loop of the nervous system becomes untwisted and symmetrical. However, the organs lost during torsion are not replaced as a result of which the opisthobranchs also possess only one ctenidium, one kidney, and one auricle. In some opisthobranch mollusc like Acteon, and Bulla, detorsion is partial. The visceral loop remains partly twisted in them and the ctenidium and anus are laterally directed. COILING Coiling is a post larval development. It is believed that the early gastropods had planospiral shell where each coil is located completely outside the coil of the preceding one but in the same plane. The shell is exogastrically coiled, i.e. the direction of coiling is anterior over the head. The centre of gravity of exogastric shell would cause the shell to fall forward in front of the head so that the animal would have to push the shell forward over the vegetation. Torsion and endogastric coiling (direction of coiling is posterior over the foot) solved this problem. PEARL FORMATION Pearls are formed in some bivalves. It is a natural secretion deposited by the mantle epithelium as a protection against foreign bodies such as sand grain, debris, any microscopic organism or even a parasite. The foreign body enters the bivalve and occupies a position in between the shell and mantle thereby causing irritation to the animal (Fig.64, 65). This induces the epithelial cells of the mantle to secrete concentric layers of nacre around the foreign particle in defence and finally pearl is formed (Fig.65). The foreign particle inside the pearl is known as nucleus and the concentric layers of nacre around it as mother of pearl. If the foreign particle is enfolded within the mantle and moved about during secretion, the pearl becomes spherical or ovoid. Finest natural pearls are produced by marine pearl oysters of Eastern Asia, of the genus Pinctada namely Pinctada margaritifera, Pinctada mertensi and Pinctada vulgaris. Unio and Anodonta also produce pearl but of inferior quality and rarely of any use. 100 hinge pearl gill mantle shell Fig.64 Site of pearl formation in Mytilus 101 nacreous layer of shell foreign body nacre secreting cells of mantle connective tissue ciliated epithelium of mantle PRIMARY STAGE nacreous layer of shell nacre secreting cells of mantle foreign body layers of nacre connective tissue ciliated epithelium of mantle PROGRESSIVE STAGE layers of nacre pearl FINAL STAGE Fig. 65 Stages of pearl formation 102 PEARL CULTURE Today pearls are artificially produced and Japan is one of the largest countries in the world to produce the bulk of pearls by pearl culture technique. Mr. Mikimoto of Japan has discovered a method of stimulating the pearl oysters to form pearls by artificially introducing foreign particles between the shell and mantle. Small windows are made on the shell through which sand grains, shell pieces etc. are introduced. The treated oysters are kept in perforated cages and lowered in the sea till pearls are formed. The main difficulty encountered in this method was that the partially exposed pallial membrane was prone to infection and damage. The latest technique used is the grafting method or nuclear insertion method. In this method, a calcareous substance (nucleus) is grafted with a piece of mantle into the body of pearl oyster. The entire technique is performed very systematically which can be summarised in steps: 1. COLLECTION OF SPAT OR BABY PEARL OYSTER: The oysters are collected from the bottom of the sea by throwing spat collectors into the water. The spat collectors consist of lines of plastic mesh bags filled up with discarded fish net to give them volume. These will stay in the water for 2-3months collecting the spat or baby pearl oyster. 2. REARING OF OYSTERS: The baby oysters as small as 3mm are segregated from each bag and put in special rearing cages known as pearl net. These are small pyramid shaped mesh cages of small mesh size. The small mesh size helps to keep the predators away. The baby oysters start growing very fast, feeding from the nutrient rich waters. The oysters become juveniles in 6-8 months and are then ready to be transferred to the next type of culture cage, the lantern cage. The lantern nets are cylindrical in shape, with 5 layers and have a wider mesh size. This is done to improve the water flow within the cage, thus allowing the oysters to feed properly. The oysters can grow faster and healthier by this process. The juvenile is kept in this cage until they become young adults attaining a size of 4-5inches (at an age of 12-18months). This is the minimum size of the oyster required to start growing a cultured pearl. The pearl seeding operation or grafting operation can now be initiated. 3. SEEDING OR GRAFTING OPERATION: The young adult oysters are taken out of their cages and scrubbed clean of the marine life growing on their shells like algae, sponges, barnacles etc. They are then transferred onto plastic trays and kept in clean and hygienic conditions in a pearl laboratory. There are two different grafting techniques and thus two different kinds of pearls can be made. a. Maybe pearls also known as half pearls. b. Loose pearls or round, and keshis pearls. 4. MAYBE IMPLANT: A minor surgery is performed on the organism to get the maybe pearl. With the help of a spatula, the mother of pearl shell, found under the mantle tissue is exposed. The maybe implant which is usually made up of plastic is then inserted into the shell and the animal is returned to its home in the water. The plastic mimics the action of the parasite drilling 103 through the shell, and the oyster’s mantle then starts secreting layers of nacre around it in defence, resulting in the formation of pearl. THE MISE-NISHIKAWA GRAFTING TECHNIQUE: i. PREPARATION OF NUCLEUS: Molluscan shell is generally used to prepare the nucleus (the foreign particle inside the pearl is called the nucleus), as it has been found that the secretion of nacre is more around the nucleus. Shell of Xancus is generally used for this purpose. The shell is cut into small cubes which are then grinded into spherical form by placing them in between two sheets of iron. The iron sheets are then revolved and the spheres smoothened. This is used as the nucleus for insertion into the oyster body which acts as the host. ii. PREPARATION OF GRAFT TISSUE: A donor oyster is needed to prepare the graft. A small strip of mantle tissue is taken from the edge of the mantle and cut into small squares. These small pieces of the mantle tissue are known as grafts. They are then kept in sea water at 22°C for 3-4hours. The outer surface of the mantle tissue is kept in contact with the nucleus to be inserted as the nacre secreting cells are found on the outer surface of the mantle. iii. IMPLANTATION OF NUCLEUS AND THE GRAFT TISSUE: The graft tissue is inserted along with the artificially prepared nucleus into the living tissue of another oyster. The recipient oyster shell is induced to open by placing it in a tray containing sea water with their dorsal portion directed downward. In a short time the shell opens. Shell openers or bamboo wedges are used to keep the valves open. An incision is made in the soft body of the oyster through which a long and slender steel probe is inserted and a tunnel is made in the soft body mass, taking utmost care and avoiding the delicate organs like the heart, liver, pancreas, kidney, stomach and intestine. At the end of the tunnel is the pearl sac made up of gonads and connective tissues. It has been experimentally found that gonads secrete nacre at a rapid rate. This is also the ideal place within the animal body to grow the cultured pearl, because it is big enough to allocate the nucleus (and future cultured pearl) and also receives nutrients from the body. The nucleus is inserted into the pearl sac. Finally, the mantle pieces i.e. the graft tissue is placed alongside the nucleus. Bamboo peg is removed and the oyster shell is automatically closed. In a fully grown oyster 4-5 nuclei can be successfully implanted. However, the operation time should not exceed 30 minutes as the oysters cannot survive more than one hour of operation time. POST OPERATIONAL CARE: The operated oysters are placed in wire cages and suspended in water from wooden rafts. The healing of the operated tissue takes about 6-7 days during which time the cages are left undisturbed. Then they are transferred to culture cages known as pocket nets and suspended at a depth of 2-3m. in water. The oysters take nearly 3-6 years for pearl formation. The little pieces of mantle tissue will graft itself onto the surrounding tissues, grow around the sphere and then begin to coat it up with microscopic layers of nacre. The culture cages are periodically checked, all encrusting organisms from the shell are removed and the oysters are protected from the parasite attack. PEARL HARVESTING: Harvesting of pearl is done from December to February, but it varies with the climatic conditions of the area. After 3years the oysters are brought to the dock, cleaned and 104 harvested. From each oyster we may get many products: loose pearls, mabe, mother-of-pearl shell (to make buttons or sell as souvenirs), and a few natural pearls. The pearls are then cleaned under freshwater and patted dry. Natural and good quality pearls need a minimum processing, but mabe pearl does need to be processed in order to be set in jewellery. The best quality pearl is Lingha pearl which is obtained from the marine oysters. CLASSIFICATION OF MOLLUSCA Molluscs are classified into seven classes according to their symmetry, shell, mantle, gills, nervous system and radula. The classification that has been followed here is from Invertebrate Zoology by Robert D. Barnes 5th edition. CLASS MONOPLACOPHORA: Gr. Monos means one; plax means plate; pherein means bearing. 1. The class consists of a small group of living species which are marine. 2. They possess a single, bilaterally symmetrical shell which varies in shape from a flattened, shield-like plate to a short cone. 3. The repetition of gills, retractor muscles, auricles and nephridia is a characteristic feature of living species. 4. They possess a broad, flat foot with pedal retractor muscles. 5. The fossil species show three to eight pairs of muscle scar. Example: Neopilina, Vema CLASS POLYPLACOPHORA: 1. Includes Chitons, about 500 living species and 350 fossil species. 2. 3. 4. 5. Bilaterally symmetrical, dorsoventrally flattened, ovoid body, with a heavy mantle. Indistinct head, with no cephalic eyes or tentacles. The mantle cavity is posterior. Broad and flat foot present for attachment to the hard substratum. Example: Cryptochiton CLASS APLACOPHORA: 1. They are found in oceans at a depth of 9000 meters. Some live on the bottom, and others creep on hydroids and corals. 2. They have a poorly developed head. 3. Shell, foot and mantle are absent. 4. Vermiform body, with a midventral, longitudinal groove containing one or more ridges which are probably homologous to the foot of other molluscs. 5. They have calcareous spicules buried in the cuticle. 6. The class comprises of about 180 species of worm like molluscs. Example: Solenogasters. 105 CLASS GASTROPODA: Gr. Gaster means belly; podos means foot 1. The class Gastropoda is the largest class of molluscs including about 35,000 existing species and 15,ooo fossil species. Mostly marine, some freshwater or terrestrial. 2. 3. 4. 5. 6. Torsion of the visceral mass takes place at some stage during development. Well developed head with eyes and tentacles. Radula present. Presence of large and flat foot. Shell usually present, but may be absent in some, univalve and usually coiled. SUBCLASS: Prosobranchia 1. Marine, freshwater and terrestrial forms in which the mantle cavity and the internal organs are anteriorly located. 2. 3. 4. 5. Shelled animals, operculum usually present. Head with a single pair of tentacles. The aquatic species possess one or two gills within the mantle cavity. Mostly dioecious. ORDER: Archaeogastropoda 1. Primitive forms possessing two bipectinate ctenidia, two auricles and two nephridia. 2. The right ctenidia may be reduced or absent in some. 3. Coiled shell. 4. Osphradium simple, ridge-like. Example: Haliotis, Patella, Fissurella, Perotrochus, Acmaea etc. ORDER: Mesogastropoda 1. Chiefly marine but freshwater and terrestrial forms are also present. 2. 3. 4. 5. Single monopectinate gill, one auricle, and one nephridium. Radula bears seven teeth in a transverse row (taenioglossate radula) Simple osphradium, ridge-like. Complex reproductive system, usually with a penis. Example: Pila, Atlanta, Carinaria, Pleurocera etc. ORDER: Neogastropoda 1. All are marine. 2. Similar to Mesogastropods in having a single monopectinate gill, one auricle and one nephridium,and in possessing a complex reproductive system. 3. Shell with a short to very long siphonal canal. 4. Radula bears three teeth in a transverse row (rachiglossate radula). 5. Complex osphradia with bipectinate folds. 6. Example: Murex, Mitra, Voluta, Oliva etc. 106 SUBCLASS: Opisthobranchia 1. Mostly marine. 2. 3. 4. 5. 6. 7. 8. 9. One ctenidia, one auricle, one nephridium and one gonad present. Ctenidium posterior to the heart. Many of them are secondarily bilaterally symmetrical. Reduction and loss of shell and mantle cavity common. They display detorsion. Head bears two pairs of cephalic tentacles. Buccal cavity with a pair of jaws. Hermaphrodite. ORDER: Cephalaspidea 1. Bubble shells. Shell generally present but reduced, absent in some. 2. Dorsal surface of head shield- like. 3. Lateral parapodial lobes prominent. Example: Acteon, Bulla etc. ORDER: Pyramidellacea 1. Ectoparasites of polychaetes and bivalves. 2. Proboscis contains a stylet instead of a radula. 3. Spirally coiled shell and operculum present. 4. Ctenidia absent. Example: Pyramidella, Brachystomia etc. ORDER: Acochlidioidea 1. Small naked species with no shell. 2. Visceral mass sharply set off from rest of the body and is covered with spicules. 3. Ctenidia absent. Example: Acochlidium, Unela ORDER: Anaspidea 1. Reduced shell, buried in the mantle. 2. Mantle cavity and ctenidia present. 3. Foot bears lateral parapodia. Example: Aplysia, Bursatella. ORDER: Notaspidea 1. Shelled or naked opisthobranchs. 2. Gills present. 3. Bipectinate osphradium on the right side. Example: Pleurobranchus 107 ORDER: Sacoglossa 1. Shelled animals with radula bearing a single row of teeth adapted for suctorial feeding. 2. Slug like opisthobranchs. 3. Herbivores. Example: Elysia, Alderia, Oxynoe. ORDER: Thecosomata 1. Sea butterflies. 2. Shelled pelagic species with large parapodia. 3. Well developed mantle cavity. Example: Spiratella, Limacina. ORDER: Gymnosomata 1. Naked pteropods 2. Pelagic species with the absence of shell and mantle cavity. 3. Parapodial fins present. Example: Pneumoderma ORDER: Nudibranchia 1. Sea slugs where the shell and mantle cavity both are absent. 2. Body is secondarily bilaterally symmetrical. Example: Doris, Tritonia, SUBCLASS: Pulmonata 1. Ctenidia absent. 2. Only one auricle and one nephridium present. 3. Mantle cavity is on the right side and is converted into a vascularised chamber for the exchange of gases. 4. Symmetrical nervous system. 5. Shell usually present but without an operculum. 6. Hermaphrodite. ORDER: Systellommatophora 1. Slugs with anus present in the posterior end of the body, instead of laterally as in other pulmonates. 2. Onchidiidae possesses a posterior pulmonary sac. Lung is lost in Veronicellidae. ORDER: Basommatophora 1. Primarily freshwater forms, some are marine. 2. Pulmonates with one pair of tentacles, eyes located near tentacle base. Example: Freshwater forms are Ferrissia, Ancylus, Lymnaea, Planorbis, Bulinus. Marine forms are Siphonaria, Otina. 108 ORDER: Stylommatophora 1. All are terrestrial. 2. Pulmonates with two pairs of tentacles, the posterior pair of tentacles bear eyes at its tip. Example: Helix, Achatina, Limax, Deroceras, Phylomycus, Partula CLASS BIVALVIA OR PELECYPODA OR LAMELLIBRANCHIA: Gr. Pelekus means batchet; podos means foot 1. The class includes clams, oysters, and mussels. 2. The members of this class are soft bottom burrowers. They burrow in soft mud and sand. 3. They have a laterally compressed body. 4. The shell has two valves, hinged dorsally and encloses the complete body. 5. Possesses a laterally compressed foot, hatchet-shaped, and extending between the mantle lobes. 6. Have large gills which have also taken over the food collecting function in addition to the gas exchange in some of the bivalves. SUBCLASS: Palaeotaxodonta 1. Shell valves equal and shell structure nacreous or cross lamellar. 2. Row of short teeth are present along the hinge margin. 3. Protobranchiate gills. ORDER: Nuculoida 1. Characteristics same as for Palaeotaxodonta. Example: Nucula, Nuculana etc. SUBCLASS: Cryptodonta 1. Thin, equal, and somewhat elongate valves. 2. Hinge teeth absent. 3. Protobranchiate gills. ORDER: Solemyoida 1. Characteristics same as Cryptodonta. Example: Solemya SUBCLASS: Pteriomorphia 1. Epibenthic bivalves attached to the substratum by byssus thread, but some are secondarily free. 2. They have unfused mantle margin. 109 ORDER: Arcoida 1. Straight hinge. 2. Filibranchiate gills. Example: Arca, Glycymeris etc. ORDER: Mytiloida 1. Valves equal, except in oysters and some scallops. 2. Inequilateral valve. Example: Mytilus, Pecten, Spondylus etc. SUBCLASS: Palaeoheterodonta 1. Equivalve, with a few hinge teeth. 2. The lateral teeth, when present, are not separated from the cardinal teeth. ORDER: Unionoida 1. Freshwater bivalves. Example: Mutela, Anodonta etc. ORDER: Trigonioida Example: Trigonia SUBCLASS: Heterodonta 1. Equivalve. Shell without nacreous layer. 2. Eulamellibranchiate gills. 3. Mantle edges usually united posteriorly at one or more points leading to the development of siphons. ORDER: Veneroida 1. Usually equivalve. Example: Solen, Mactra etc. ORDER: Myoida 1. Thin shelled burrowers with well developed siphons. 2. Shell without nacreous layer. 3. They have either one or no cardinal teeth. Example: Pholas, Mya, Teredo etc. SUBCLASS: Anomalodesmata 1. Equivalve, with a single hinge tooth or no teeth. 2. Hinge margin thickened. 3. Mantle margin fused. 4. Hermaphrodite. ORDER: Pholadomyoida Characteristics same as Anomalodesmata Example: Poromya, Pandora etc. 110 CLASS SCAPHOPODA: Gr. Scapha means boat; podos means foot 1. The class scaphopoda contains about 350 species, popularly known as tusk or tooth shells. 2. They are burrowing marine molluscs that burrow in sand in water depths greater than 6 meters. The animal lies buried with a larger anterior end downward, and the posterior end near the surface of the substratum. 3. They have an elongated cylindrical tube like body shaped like an elephants tusk. Both ends of the tube are open. 4. The head and foot project from the larger anterior aperture of the shell. 5. They have paired kidneys but a single gonad. Example: Dentalium, Cadulus, Pulsellum CLASS CEPHALOPODA: Gr. Kephale means head; podos means foot 1. Members of this class are adapted for swimming except for a few bottom dwelling forms like Octopus. 2. The body is elongated along the dorso- ventral axis which is the functional anterior posterior axis. 3. Shell may be external, internal or absent. 4. The tentacles are present at the anterior end of the body and are homologous to the foot of other molluscs. 5. Distinct large head with well-developed eyes. 6. Radula present. SUBCLASS: Nautiloidea 1. External shell present, which may be straight or coiled. 2. Living forms possess many slender, suckerless tentacles. 3. The members of this class possess two pairs of gills and two pairs of nephridia. Example: Nautilus SUBCLASS: Ammonoidea 1. Fossil forms with coiled external shells having complex septa and sutures. Example: Ceratites, Scaphites etc. SUBCLASS: Coleoidea 1. Internal shell, reduced, or absent. 2. The arms bear suckers. 3. One pair of gills and one pair of nephridia present. ORDER: Belemnoidea 1. Extinct species. 2. Internal shell. Example: Belemnites ORDER: Sepioidea 1. Possesses eight arms and two tentacles 111 2. Shell with septa, or shell greatly reduced or lost. 3. Body mostly short and broad or saclike. Example: Sepia, Spirula, Sepiola etc. ORDER: Teuthoidea 1. Mostly elongate body with eight arms and two long tentacles. 2. Internal shell lacks a rostrum. Example: Loligo, Abralia etc. ORDER: Vampyromorpha 1. Small, deep water dwellers with eight arms united by a web. 2. Large terminal body fins. 3. A pair of small retractile, coiled filaments present. Example: Vampyroteuthis ORDER: Octopoda 1. Globular body with eight equal arms. 2. Lacks internal shell. Example: Octopus, Argonauta, Amphitretus etc. 112 BIBLIOGRAPHY Afferent vessel: A vessel leading towards a given position. Bivalve: Molluscs with two valves. Calcareous: Composed of calcium salt. Efferent vessel: A vessel leading away from the given point. Ganglion: A mass of cell bodies of nerve cells located outside the central nervous system. Haemocoel: Coelom filled with blood. Pelagic: Living in open water, as in the mid-ocean. Univalve: Mollusc with a single shell. Veliger: Larva of molluscs. Suggested Reading 1. Robert D. Barnes : Invertebrate Zoology , Publisher: Saunders College International Edition (5th Edition) 2. Ruppert, Fox and Barnes : Invertebrate Zoology, Publisher: Saunders College International Edition (7th Edition) 3. Parker And Haswel: Text Book Of Zoology, Invertebrates, Volume 1 Edited by Marshall And Williams (7th Edition), A.I.T.B.S. Publishers And Distributors 4. Biology Of Animals, by Ganguly, Sinha and Adhikari, Publisher: New Central Book Agency 5. Modern text book of Zoology, Invertebrates by R. L. Kotpal, Publisher : Rastogi Publications 113