Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Plant Science Letters, 22 (1981) 169--176
© Elsevier/North-Holland Scientific Publishers Ltd.
169
PRESENCE OF ALLHNASE IN ISOLATED VACUOLES AND OF ALKYL
CYSTEINE SULPHOXIDES IN THE CYTOPLASM OF BULBS OF ONION
(ALLIUM CEPA )
JANE E. LANCASTER* and H.A. COLLIN
Department o f Botany, University of Liverpool, P.O. Box 147, Liverpool L69 3BX
(United Kingdom)
(Received December 1st, 1980)
(Revision received February 23rd, 1981)
(Accepted March 3rd, 1981)
SUMMARY
Protoplasts were isolated from the inner scales of mature onion bulbs,
then the vacuoles liberated from the protoplasts by gentle osmotic shock.
Protoplasts and vacuoles were compared for alliinase activity (all'fin alkylsulphenate-lyase, EC 4.4.1.4). The vacuoles contained alliinase in amounts
which account for the alliinase found in protoplasts. Both vacuoles and
protopiasts were analysed for 8-alkyl substituted cysteine sulphoxides by
electrophoresis-thin-layer chromatography (TLC). Whereas the vacuoles
contained negligible amounts of alkyl cysteine sulphoxides, the cytoplasm
of the protoplasts contained a considerable a m o u n t of alkyl cysteine sulphoxides. It is concluded that alliinase and alkyl cysteine sulphoxides are
prevented from acting in the intact protoplast by compartmentation of the
alliinase in the vacuoles and alkyl cysteine sulphoxides in the cytoplasm.
INTRODUCTION
8-Alkyl substituted cysteine sulphoxides are the characteristic secondary
amino acids of the Amaryllidaceae and Cruciferae [ 1]. There are also present
in each plant family enzymes which degrade these amino acids to form
pyruvate, ammonia and a sulphur-containing product. In onions and garlic
these reactions are of economic importance because the volatile sulphur
products give rise to the characteristic flavour of these plants. It is generally
*To whom reprint requests should be addressed at: Applied Biochemistry Division, Department o f Scientific and Industrial Research, Private Bag, Christchurch, New Zealand.
Abbreviations: DTT, dithiothreitoi; FDA, fluorescein diaeetate; TLC, thin-layer chromato-
¢~phy.
170
considered that the enzyme and the amino acid substrate are separated in
vivo and only react on rupture or wounding of the cells.
Wagner and Siegelman [2] developed techniques for the isolation of large
numbers of intact vacuoles. These and similar techniques have been used to
study the localisation of the enzyme alliinase and the S-alkyl substituted
cysteine sulphoxides in onion bulb.
MATERIALS AND METHODS
Plant material
Mature bulbs of Allium cepa var. Rijnsburger were purchased from a local
market and dissected to give an inner scale, next to the first lateral bud. The
inner and outer epidermis of the scale were removed and any true stem (base
plate} and greening upper scale were excised. It was important to use an
inner scale of smaller, less mature, but uniform cells.
Protoplast isolation
Onion scale (7 g) was finely sliced (0.5--1.0 mm) into 70 ml of incubation
medium containing 0.7 M sorbitol, 0.5% cellulase O n o z u k a R-10, 0.0001%
pectolyase [3] at pH 4.8--5.0. Incubation was in the dark at 25°C, overnight
( 1 6 : 0 0 ~ 9 : 0 0 ) , b u t with no shaking. Increasing the shaking produced
ruptured protoplasts and considerable cellular debris. The digested bulb
scales were filtered through plancton net (pore size 200 × 250 ~M) into a
test tube.
The onion protoplasts were large and dense, but also very fragile. The
protoplasts were allowed to settle under gravity for 30 rain (microscopic
observation indicated when the majority had sedimented). The lighter
cellular debris remained in suspension and was sucked off from the loose
protoplast pellet. The pellet was resuspended in 0.7 M sorbitol (pH 5.0) to
wash away any remaining enzyme. Sedimentation was repeated as before.
The pellet was transferred to a graduated 10 ml centrifuge tube. Protoplast
numbers were c o u n t e d using an improved neubauer h a e m a t o c y t o m e t e r .
Protoplast viability was determined using fluorescein diacetate (FDA).
Intact cells fluoresce as fluorescein, an enzymic product of FDA, accumulates
inside the membrane [4].
Vacuole isolation
To 0.5 ml loose pellet of protoplasts in 0.7 M sorbitol (pH 5.0), 4.5 ml
0.45 M sorbit~ol (pH 7.5), 2 mM EDTA and 1 mM dithiothreitol (DTT)
were gently added. The tube was gently inverted several times to mix the
contents. The contents were allowed to stand for several minutes before
again inverting the tube to mix. This was repeated several times. After
5--10 min microscopic observation of an aliquot indicated whether or
n o t protoplast lysis was complete. The plasmalemma and cytoplasmic debris
aggregated to form a 'slime'. Vacuoles could be seen sedimenting as opal-
171
escent points. The tube contents were gently poured through 200 X 250 ~M
plancton net, which held the aggregated debris. The debris was washed with
0.7 M sorbitol (pH 5.0), to wash free any adhering vacuoles. It was important
at this point to add the 0.7 M sorbitol (pH 5.0), to the vacuole solution to
stabilise the vacuoles. Any further dilution of the vacuole solution resulted
in lysis of the vacuoles.
Vacuoles were counted using an haematocytometer and were allowed to
sediment under gravity for 10--15 min. The supernatant containing the soluble components of the cytoplasm and of burst vacuoles, a few very small
vacuoles and an occasional piece of debris was sucked off and spun at
500 × g for 10 rain. The supernatant thus produced was used in succeeding
assays as the cytoplasm fraction.
Alliinase determination
Alliinase was assayed using exogenous alkyl cysteine sulphoxide and
determining the pyruvate produced, by the method of Schwimmer and
Guadagni [5]. The protoplast sample and the vacuole sample were each
shaken on a rotomix to burst the particular organelle and made up to 5 ml
with 0.45 M sorbitol, 2 mM EDTA and 1 mM DTT (pH 7.5). The cytoplasm fraction was also made up to 5 ml.
The standard reaction mixture consisted of the following components:
0.2 ml of 0.5 mM pyridoxal phosphate, 0.4 ml of 0.1 M sodium pyrophosphate buffer (pH 9.0), 0.4 ml of 100 mM synthetic S-methyl-L-cysteine sulphoxide, at 2 concentrations of enzyme viz. 0.5 ml and 1.0 ml [6]. The
mixture was incubated at 25°C and 0.25-ml aliquots taken at 0-, 10-, 20- and
30-rain intervals to determine completeness of pyruvate production. The
reaction was terminated by adding the aliquot to 1 ml 0.0125% 2,4-dinitrophenylhydrazine in 2 N HC1, which was then ready for determination of
pyruvic acid.
Protein concentrations of the enzyme samples were determined at the
above mentioned concentrations of 0.5 and 1.0 ml using the procedure of
Lowry et al. [7]. Recrystallised bovine serum albumin was used as the
standard.
Determination of S-alkyl cysteine sulphoxides
The method used was an adaptation of that described by Bieleski and
Turner [8] in that electrophoresis was used as a preparative step to remove
sugars, organic acids and most amino acids. Alkyl cysteine sulphoxides were
eluted and separated by thin-layer chromatography (0.2 m m silica gel), in
methyl ethyl ketone/pyridine/water/acetic acid (70 : 15 : 15 : 2, v/v) then
(in the same direction) in n-propanol/water/n-propyl acetate/acetic acid/
pyridine (120 : 60 : 20 : 4 : 1, v/v). They were vimmlim~ by spraying with
polychromatic ninhydrin reagent [9] and heating 105°C 1.5--2 min. Cis~q(1-propenyl)-L-cysteine sulphoxide appeared as a pink-red spot Rf ffi 0.34;
$-methyl-L-cysteine sulphoxide and 8-propyl-L-cy~teine sulphoxide appeared
172
as yellow-red spots at R f = 0.28 and 0.47, respectively. Quantities of the
alkyl cysteine sulphoxides were estimated f r o m various amounts of the
appropriate standards using spot size and colour intensity.
RESULTS
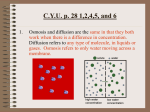
Protoplasts isolated by the above procedure ranged in size from 80 to 180
uM (Fig. 1). Staining with FDA and observation under a fluorescent microscope showed a viability of about 95%. Little cellular debris was present.
Lowering the sorbitol concentration of the m e d i u m and increasing the pHvalue caused protoplasts to lyse and vacuoles to emerge. The vacuoles were
thin-wailed and quite translucent under the microscope {Fig. 2). There was
hardly any contamination from cytoplasmic debris in the vacuole pellet.
Vacuoles were f o u n d to contain alliinase (Table I). The alliinase from
Fig. 1. Protoplasts from inner scale of mature onion bulb; isolated overnight in 0.7 M
sorbitol (pH 4.8), 0.5% cellulue; 0.0001% pectolyase, no shaking. (Scale bar ffi 100 #M).
173
Fig. 2. Vacuoles prepared from protoplasts of onion bulb, after sedimentation under
gravity from 0.45 M sorbitol (pH 7.5) 1 mM D ~ F , 2 rnM EDTA. (Scale bar ffi I 0 0 ~,M).
TABLE I
ALLIINASE ACTIVITY IN THE VACUOLES, CYTOPLASM AND PROTOPLASTS
OF ONION
Alliinase was assayed using exogenous alkyl cysteine sulphoxide and determining the
pyruvate produced, as described in Materials and Methods. Pyruvate levels were corrected
for endogenous pyruvate.
Organelle
Protoplast
Vacuole
CytoplIm
Number
12 X 107
5 x 107
__a
Protein/
organeUes
Pyruvate/organelles
(mg)
~mol
mg
Alliinase specific
activity
(~mol/mg protein)
6.2
2.6
3.5
30.9
10.9
16.3
3.4
1;2
1.8
4.9
4.2
4.6
aComprising contents of 12 X 107 protoplasts and 7 x I 0 ' burst vacuoles.
174
TABLE II
AMOUNT OF S-ALKYL SUBSTITUTED CYSTEINE SULPHOXIDES IN THE
VACUOLES, CYTOPLASM AND PROTOPLASTS OF ONION
S-Alkyl substituted cysteine sulphoxides were determined as described in Materials and
Methods.
Organelle
Number
Alkyl cysteine sulphoxide
g/organelles
Protoplast
Vacuole
Cytoplasm
40 × 10"
8 × 10 ~
a
880
Trace (<2)
80
in p r o t o p l ast %
100
0.5--1.0
10
aComprising contents of 40 × 10 ~ protoplasts and 32 × 107 burst vacuoles.
12 × 107 protoplasts produced 3.4 mg pyruvate from exogenous methyl
cysteine sulphoxide; alliinase from the same number of vacuoles would
have produced 2.9 mg pyruvate from exogenous substrate. Specific activity
of the alliinase in the protoplasts and vacuoles was similar.
The soluble cytoplasmic fraction, formed from the cytoplasm of all the
lysed protoplasts and some burst vacuoles (about 50--60% in this example)
also contained alliinase, of a similar specific activity to the protoplasts and
vacuoles. Protein levels of the cytoplasm and vacuoles, added together, were
equal to those of the intact protoplasts. Similarly the total pyruvate formed
by the action of cytoplasmic and of vacuole alliinase on exogenous substrate
(3.0 mg) was almost equal to the pyruvate formed from the alliinase of
intact protoplasts (3.4 mg).
TLC-electrophoretic analysis of extracts of intact protoplasts showed 3
spots only at Rf-values of 0.27, 0.32 and 0.46; which corresponded in
colour and Rf to the 3 alkyl cysteine sulphoxide standards. Propenyl cysteine
sulphoxide was the predominant compound in the protoplasts, methyl
cysteine sulphoxide and propyl cysteine sulphoxide being much less. Vacuoles were found to contain a negligible amount of alkyl cysteine sulphoxides,
about 0.5- -1.0% of that found in the extract of intact protoplasts (Table II).
Very small vacuoles spun down from the crude cytoplasmic fraction contained no alkyl cysteine sulphoxides. The cytoplasmic fraction contained
a greater quantity of the alkyl cysteine sulphoxides than did the vacuoles,
but only about 10% of the amount in the extract of intact protoplasts.
DISCUSSION
Gentle overnight enzymic digestion of cell walls gave large numbers of
viable protoplasts with very little cellular debris. Shaking, higher enzyme
concentrations and the use of strong enzymes such as macerozyme gave
fewer intact protoplasts and large amounts of cellular debris. Protoplasts
could not be isolated from bulb scales with higher enzyme concentrations
and short incubation times of a few hours.
175
It was observed that protoplasts formed from different cell types had
slightly different optimal conditions for lysis.Thus bulb scales were removed
of epidermis, base plate and any greening portions to give as uniform a
population of protoplasts as possible. It was noteworthy in this respect that
young green leaf cylinders gave good protoplasts in the enzyme conditions
described above, but that those protoplasts were from several cell types and
thus good yields of vacuoles were not obtained, some protoplasts bursting
completely, others producing vacuoles and some not lysing at all.
Yields of vacuoles obtained from the lysis conditions described above
varied from 20% to 50% (the remainder of the vacuoles bursting after or
during protoplast lysis);this compares favourably with the figure of about
10% obtained by Wagner and Siegelman [2]. Vacuole preparations usually
contained a few intact protoplasts, although the contamination of the tonoplast by cytoplasmic membranes observed by Saunders and Conn [10] was
not observed in the vacuoles isolated herein (Fig. 2). Thus the cytoplasm
fraction reported in the tables comprised the water soluble contents of the
cytoplasm and the water soluble contents of any burst vacuoles. Aggregated
plasmalemma components were removed by filtration through plancton net.
Vacuoles isolated by the above procedures contained alliinase but not
alkyl cysteine sulphoxides. (The trace of alkyl cysteine sulphoxides was
probably the result of the 1% or so contamination by intact protoplasts).
The amount of alliinase in the vacuoles was sufficient to account for the
alliinase in a similar number of intact protoplasts, suggesting that in the intact
cell all the alliinase is localised in the vacuole. The cytoplasmic fraction also
contained alliinase, an expected occurrence in view of the contamination of
this cellular component by the contents of the burst vacuoles. However, the
amount of alliinase in the cytoplasmic fraction can be accounted for by the
number of burst vacuoles liberating their alliinase.
Alkyl cysteine sulphoxides, absent from the vacuoles, were found in the
cytoplasmic fraction, but not as much as in intact protoplasts. It is likely
that the alliinase liberated into the cytoplasm-osmoticum from burst vacuoles
had already begun to degrade the alkyl cysteine sulphoxides from the cytoplasm. Both alkyl cysteine sulphoxides and alliinase were readily soluble in
the aqueous osmoticum. Extracting the cytoplasm (minus intact vacuoles)
into methanol/chloroform/water immediately denatured the alliinase and
prevented further degradation of the alkyl cysteine sulphoxides. Thus the
cellular compartmentation maintains the intactness of the alliinase and
alkyl cysteine sulphoxides in the undamaged cell.
ACKNOWLEDGEMENT
We wish to thank Dr. I. McKenzie, Nottingham University, for the gift of
the pectolyase.
REFERENCES
1 L. Fowden, Ann. Rev. Biochem., 33 (1964) 173.
176
2 G.J. Wagner and H.W. Siegelman, Science, 190 (1975) 1298.
3 T. Nagata and S. Ishii, Can. J. Bot., 57 (1979) 1820.
4 P.J. Larkin, Planta, 128 (1976) 213.
5 S. Schwimmer and D.G:Guadagni, J. F o o d Sci., 27 (1962) 94.
6 C. Selby and H.A. Collin, Ann. Bot., 40 (1976) 911.
7 0 . H . Lowry, N.J. Rosebrough, A.L. Farr and R.J. Randall, J. Biol. Chem., 193 (1951)
265.
8 R.L. Bieleski and N.A. Turner, Anal. Biochem., 17 (1966) 278.
9 E.D. Moffatt and R.I. Lytle, Anal. Chem., 31 (1959) 926.
10 J.A. Saunders and E.E. Conn, Plant Physiol., 61 (1978) 154.