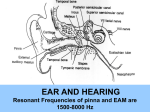
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Journal of Medical Microbiology (2010), 59, 377–383 Review DOI 10.1099/jmm.0.015792-0 Potential role for lipopolysaccharide in congenital sensorineural hearing loss A. L. Smit,1 R. J. Stokroos,1 S. G. H. Litjens,2 B. Kremer1,3 and B. W. Kramer3,4 Correspondence A. L. Smit [email protected] 1 Department of Otorhinolaryngology/Head and Neck Surgery, Maastricht University Medical Centre, PO Box 5800, Maastricht, The Netherlands 2 Faculty of Health, Medicine and Life Sciences, University of Maastricht, PO Box 616, Maastricht, The Netherlands 3 School of Oncology and Developmental Biology, University of Maastricht, PO Box 5800, Maastricht, The Netherlands 4 Department of Pediatrics, Maastricht University Medical Centre, PO Box 5800, Maastricht, The Netherlands Congenital sensorineural hearing loss (SNHL) is common. In the Western world, the incidence is 1–3 per 1000 live births. The aetiology encompasses genetic and non-genetic factors accounting for 55 % and 45 % of cases, respectively. Reports that describe the contribution of intrauterine infection to the occurrence of congenital SNHL are limited, and comparative analysis of the different pathogens is lacking. Lipopolysaccharide (LPS), a product of bacteriolysis, has been demonstrated to be associated with inner ear damage in experimental studies. To elucidate the potential role of this toxin in congenital SNHL and to identify the pathogenesis and transmission routes, we reviewed the literature. We speculate that different routes of exposure to LPS in utero may result in congenital inner ear damage. Introduction Congenital sensorineural hearing loss (SNHL) in childhood is common. In the Western world, the incidence amongst full-term live births amounts to 1–3 per 1000 (Davis et al., 1995; Erenberg et al., 1999; Mencher, 2000; Nance et al., 2006). In preterm infants, the incidence is even higher, with 2–4 per 100 live births (Erenberg et al., 1999). The aetiology of congenital SNHL is attributable to genetic and non-genetic factors in 55 % and 45 % of cases, respectively (Nance et al., 2006). Non-genetic causes are diverse, e.g. perinatal exposure to hypoxia (Abramovich et al., 1979), acidosis (Marlow et al., 2000), otoxicity of drugs such as aminoglycoside or loop diuretics (Young & Magnum, 2007) and viral or bacterial infections (Gross et al., 2000). In this review, we will focus on the infectious causes of congenital SNHL, of which infection with cytomegalovirus (CMV) is the most common (Nance et al., 2006). The estimated prevalence of CMV infection amounts to 0.2–2.2 % of all live births (Ludwig & Hengel, 2009; Mussi-Pinhata et al., 2009). Ten to twenty per cent of infants congenitally infected with CMV had varying degrees of hearing loss at birth (Katano et al., 2007). Besides CMV, other micro-organisms, e.g. Toxoplasma gondii, measles virus, mumps virus, varicella-zoster virus, parvovirus B19, herpes simplex virus, Treponema pallidum and rubella virus, have been identified to cause unilateral 015792 G 2010 SGM or bilateral congenital SNHL, which may be temporary or permanent, with or without loss of vestibular function (Gross et al., 2000; Wild et al., 1989). Bacterial products leading to SNHL are the subject of increasing interest. Lipopolysaccharide (LPS) is released from the outer wall of Gram-negative bacteria such as Escherichia coli and Neisseria spp. after bacteriolysis (Darrow et al., 1992). This bacterial endotoxin consists of a lipid A, a core polysaccharide and an O antigen and is known for its multiple effects on biological systems, e.g. induction of inflammation and the modulation of the immune responses (Darrow et al., 1992). It is also associated with functional and morphological inner ear damage in experimental animal models (Guo et al., 1994; Watanabe et al., 2001, 2002), which we will discuss in the next paragraph. We hypothesize that this toxin, by means of an intrauterine infection, can play a role in the pathogenesis of congenital SNHL. LPS-induced functional and morphological inner ear changes By discovering LPS in middle ear effusions in chronic otitis media (Bernstein et al., 1980; DeMaria et al., 1984; Iino et al., 1985), attention was drawn to its crucial role in the onset of SNHL. Subsequent functional studies did Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 16 Jun 2017 14:06:22 Printed in Great Britain 377 A. L. Smit and others demonstrate an elevation of hearing level thresholds after LPS exposure. Comis et al. (1991) and Tarlow et al. (1991) revealed early onset SNHL within 2 h after LPS application into the guinea pig inner ear by cochlear potential measurement. In a similar model, high frequency losses were observed by auditory brainstem response measurement within 1 (Guo et al., 1994) or 2 (Darrow et al., 1992) days after LPS application. A gradual improvement of responses was seen after a 5 day interval to near baseline level by 28 days (Darrow et al., 1992). To explain the LPS-related functional impairment of the inner ear, several morphological temporal bone studies were designed using different animal models. These findings are summarized in Table 1. Comis et al. (1991) and Tarlow et al. (1991) showed loss of hair bundles of inner and outer hair cells as well as swelling of the tectorial membrane directly after LPS application into the inner ear of guinea pigs. Darrow et al. (1992) demonstrated in the same species a dosage-dependent inflammatory reaction with inflammatory cells in the perilymphatic spaces; lower LPS dosages resulted in minimal inflammation during the evaluation period of 7 days. Higher dosages showed a higher degree of inflammation after 2–4 days which resolved at day 7 (Darrow et al., 1992). Also the stria vascularis revealed LPS-related alterations in the first days after exposure (Guo et al., 1994). Watanabe et al. (1995) demonstrated similar results after LPS application into the middle ear. Moreover, neutrophil recruitment was seen by myeloperoxidase staining in multiple cochlear structures 48 h after LPS exposure (Watanabe et al., 2001). Other experimental animal models and application schemes demonstrated analogous patterns of inflammatory and structural changes within small variations of time of onset. Within 12–72 h post middle ear application of LPS in a chinchilla model, bleeding and inflammation in the perilymphatic spaces and spiral ligament, strial swelling and sensory cell degeneration were observed (Kawauchi et al., 1989; Lim et al., 1990). Repeated application of LPS at the round window niche of rats by Spandow et al. (1989) resulted in hair cell loss within 2 weeks. Nakai et al. (1980) observed similar results as well as a total disappearance of outer hair cells and nerve fibres in the spiral lamina after a period of 1.5 months (Nakai et al., 1980). Pathogenesis of LPS-induced inner ear dysfunction LPS is known for its diverse effects on the inflammatory cascade through activation of the Toll-like receptor 4 (TLR-4). These receptors are expressed in the inner ear by immune cells such as macrophages and B cells, and non-immune cells, including fibroblasts and epithelial cells (Wang et al., 2006). TLR-4 signalling activates nuclear factor kappa B, which is the ‘master switch’ for inflammation-mediated cytokine production and release of mediators such as nitric oxide (NO) (Mustafa et al., 1989; Ramilo et al., 1989; Spandow et al., 1989; Watanabe et al., 378 2000b). In the resulting network of pro- and antiinflammatory cytokines, interleukin-1 (IL-1), IL-2, tumour necrosis factor alpha and transforming growth factor beta are the most important cytokines and modulators for the inflammatory response as well as for homeostasis and tissue repair (Garcia Berrocal & Ramirez-Camacho, 2000; Harris et al., 1997). Recent studies provided new insights into the relationship between LPS and NO related potential inner ear damage (Shi et al., 2003). NO has a role in regulation of vascular tone, auditory transduction and cochlear and vestibular neurotransmission (Fessenden & Schacht, 1998; Hess et al., 1998; Morizono & Ikeda, 1990; Takumida & Anniko, 2004a, b; Zdanski et al., 1994). Besides this, NO has a dose-dependent action. In small amounts, it has a defensive action against invading organisms by inhibition of adhesion molecule expression, cytokine and chemokine synthesis and leukocyte adhesion and transmigration (Guzik et al., 2003; Liaudet et al., 2000; Takumida et al., 2000). When produced in larger amounts, NO can be injurious to healthy tissue such as cochlear cells (Amaee et al., 1997; Forstermann et al., 1994; Iadecola et al., 1995a, b; Jung et al., 2003; Schmidt & Walter, 1994; Takumida et al., 2000) due to formation of peroxynitrite (ONOO2) by reaction of NO with the superoxide anion (O{ 2 ) (Kastenbauer et al., 2001) (Fig. 1). NO is synthesized by NO synthase (NOS). This enzyme has different isoforms: constitutive and inducible NOS (cNOS and iNOS) (Takumida & Anniko, 2004a). The neuronal cNOS is always present and produces, depending on the intracellular calcium levels, small amounts of NO. In contrast, the iNOS isoform is usually absent, but can be synthesized by virtually any cell type when adequately stimulated by cytokines or LPS (Hess et al., 1999; Moncada et al., 1991; Takumida et al., 2000; Takumida & Anniko, 2004a; Watanabe et al., 2000a, b, 2001). This LPS-mediated elevation of iNOS was also demonstrated in the walls of blood vessels of the cochlear lateral wall and in different cells of the organ of Corti, including in stereocilia of inner and outer hair cells, pillar cells, hair cell nerve fibres and in marginal cells (Hess et al., 1999; Shi et al., 2003; Watanabe et al., 2000a). By this mechanism, the formation of NO and subsequent peroxynitrite could be responsible for the described functional and morphological cochlear alterations after LPS exposure of the inner ear. In utero lesion of the inner ear As discussed above, experimental studies have shown LPSinduced functional and structural inflammatory changes in the inner ear. However, several pathways must be considered to explain LPS-related in utero inflammatory changes of the inner ear and subsequent potential congenital SNHL. Fetal inflammation is the consequence of a chorioamnionitis (Kramer et al., 2005b). This is defined as an inflammation of the amniotic fluid and membranes (Goldenberg et al., 2000). This inflammatory process is Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 16 Jun 2017 14:06:22 Journal of Medical Microbiology 59 Potential role for LPS in congenital hearing loss Table 1. Effects of LPS on the inner ear in experimental animal models Reference Animal model LPS origin Methods* Evaluation time after application ResultsD Degeneration of OHCs and IHCs and stria vascularis. Vacuolization and degeneration of vestibular sensory cells; vacuolization of dark cells. OHC loss Nakai et al. (1980) Mice E. coli Repeated intradermal and intravenous injection 3 days/1.5 months/3 months Spandow et al. (1989) Kawauchi et al. (1989) Rat E. coli 2 weeks Chinchilla Salmonella typhimurium Instillation into RW niche Instillation into tympanic cavity Lim et al. (1990) Chinchilla Salmonella typhimurium Instillation into tympanic cavity 12 h–3 weeks Comis et al. (1991); Tarlow et al. (1991) Darrow et al. (1992) Guo et al. (1994) Guinea pig E. coli Hours Guinea pig Guinea pig Salmonella typhimurium E. coli Watanabe et al. (1995) Guinea pig E. coli Perfusion in scala tympani or injection in CSF Inoculation in scala tympani Perfusion into perilymphatic space via RWM Intraperitoneal injection and application at RW niche Watanabe et al. (2001) Guinea pig Not specified Transtympanical injection 2 days Watanabe et al. (2002) Guinea pig E. coli Transtympanical injection 2 days 12–72 h 1–7 days 1/3/5 days 2 days/1 day Bleeding and inflammatory cell recruitment in perilymphatic space including the spiral ligament, strial swelling and sensory cell degeneration Inflammatory infiltrates in the scala tympani, vestibule and spiral ligament, strial swelling, and sensory cell degeneration Damage cochlear hair cells, swelling of the tectorial membrane Inflammatory infiltrates in perilymphatic spaces Damaged stria vascularis Inflammatory cells in the endoand perilymphatic spaces, stria vascularis, spiral ligament and organ of Corti. Widened intercellular spaces in the stria vascularis. Bleeding in the endoand perilymphatic spaces. MPO in the spiral ligament, stria vascularis and supporting cells of the organ of Corti MPO in the sensory epithelium and dark cell area *CSF, cerebrospinal fluid; RW, round window; RWM, round window membrane. DIHCs, inner hair cells; OHCs, outer hair cells; MPO, myeloperoxidase. generally regarded as a continuum with early-stage maternal neutrophils involved progressing to later stages with a fetal neutrophil inflammatory reaction (Goldenberg et al., 2000). Micro-organisms responsible for chorioamnionitis can reach the chorioamnion by the abdominal cavity through the Fallopian tubes or inadvertent needle contamination at the time of amniocentesis or chorionicvillus sampling, by passage through the cervix from the vagina (Goldenberg et al., 2000; Romero & Mazor, 1988), or transplacentally due to a maternal systemic infection (Goldenberg et al., 2000) (Fig. 2). The micro-organisms or the microbial products, once present in the amniotic fluid, can gain access to the fetus (Goldenberg et al., 2000) and evoke a local inflammatory response by entry via the http://jmm.sgmjournals.org respiratory tract, skin, gastrointestinal tract and ear (Lee et al., 2007). When neutrophils infiltrate the umbilical cord vessels (Sampson et al., 1997), this can be considered a progression of a chorioamnionitis to a funisitis, which results in a fetal systemic inflammatory response syndrome (Lee et al., 2007; Yoon et al., 2000). This is associated with increased morbidity and mortality of the fetus (Kramer et al., 2005b, 2009b). Once the inflammatory mediators or micro-organism enter the fetus, several routes are possible for reaching the labyrinth (Fig. 2). The upper respiratory tract as well as passage of the tympanic membrane gives entry to the middle ear. Subsequently, the oval and round window are a Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 16 Jun 2017 14:06:22 379 A. L. Smit and others Fig. 1. Mechanisms leading to an LPS-related inflammatory response of the inner ear and cell death through the action of cytokines and NO. porte d’entrée to the inner ear. The round window membrane can be passed easily by small molecular mass substances (Cureoglu et al., 2005; Goycoolea et al., 1988; Goycoolea, 2001; Kawauchi et al., 1989; Spandow et al., 1988; Trinidad et al., 2005) in contrast to large molecular mass substances such as albumin or LPS (Goldberg et al., 1981; Goycoolea et al., 1980; Spandow et al., 1988). However, previous studies have demonstrated passage of the toxin and cochlear inflammation by a suspected LPSrelated alteration of the membrane permeability or structure (Cureoglu et al., 2005; Darrow et al., 1992; Engel et al., 1995; Goycoolea et al., 1988; Juhn et al., 1989; Kim & Kim, 1995; Morizono & Ikeda, 1990; Paparella et al., 1970; Schachern et al., 1992; Spandow et al., 1988, 1989, 1990; Takumida & Anniko, 1998, 2004b; Trinidad et al., 2005). Another option is a haematogenous route via blood vessels of the inner ear (Watanabe et al., 2002), which are mainly located in the spiral limbus and the lateral cochlear wall (Kastenbauer et al., 2001; Li et al., 2008; Watanabe et al., 2002). Previous findings after LPS exposure-related damage in the endolymphatic compartment suggested blood–labyrinth barrier disruption by the inflammatory molecules and the capability of reaching the endolymph (Li et al., 2008). This barrier may be even more permeable at very early stages of development as shown in a rat model (Suzuki et al., 1998). Also the permeability to proteins of the blood–brain barrier is affected by LPS (Wang et al., 2006). This can result in a third, neurogenic, route, in which the infection could travel via meninges and along the eighth cranial nerve through the modiolus to the spiral ganglion and limbus (Klein et al., 2008; Li et al., 2008). Besides this, propagation of infections from the cerebrospinal fluid to the peri- and endolymphatic space via the cochlear aquaduct or endolymphatic duct, respectively, has been observed (Klein et al., 2008; Li et al., 2008). Role of LPS in congenital hearing loss Regarding the described LPS-related functional and morphological effects on the inner ear as well as the presumed transmission routes, an association between an LPS-induced chorioamnionitis and congenital inner ear changes or congenital SNHL seems plausible. This is supported by the findings of Suppiej et al. (2009), who recently described a positive association between a histological identified chorioamnionitis and SNHL in childhood after preterm delivery before 32 weeks gestational age. They postulated a relationship with worse neurological and cochlear development and maturation. Fig. 2. Inflammatory pathways from the uterine compartment to the fetal inner ear. 380 Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 16 Jun 2017 14:06:22 Journal of Medical Microbiology 59 Potential role for LPS in congenital hearing loss However, it was not possible to determine whether preterm histological chorioamnionitis in itself or the higher exposure to other risk factors such as neonatal hypotension had concomitant roles in this relationship with SNHL. The presented studies demonstrating LPS-induced functional or morphological inner ear changes were done using different experimental animal models and time intervals. To what extent these changes can be extrapolated to the human fetus has not been addressed so far. It is unclear whether the described alterations are persistent or fluctuating in time or will recover in the long term with or without removal of the stimulus. Moreover, the ototoxic potential of LPS seems to be dependent on dosage, LPS type (Darrow et al., 1992) and application method. Intramuscular (Jobe et al., 2000), intraperitoneal (Newnham et al., 2002) and intravascular (Cock et al., 2002) administration show more severe effects on the fetus (Jobe et al., 2000; Nitsos et al., 2002) than intra-amniotic LPS injection (Moss et al., 2003). Previous studies in a sheep model did confirm the hypothesis of mechanisms of maturation of the fetal immune system in the case of repeated exposure to LPS (Kallapur et al., 2007; Kramer et al., 2005a, 2007, 2009a; Kramer, 2008). To what extent this will influence the immunological response of the fetal inner ear to prolonged in utero exposure to LPS is not clear. By designing a new experimental animal model we hope to answer some of these fundamental questions in the near future to elucidate the role of LPS in and mechanisms of LPS-related congenital SNHL after intrauterine fetal exposure. A better understanding of the fetal health risks can play a role in maternal screening programmes and could be a step forward in the design of preventive strategies for congenital SNHL or early clinical screening. Darrow, D. H., Keithley, E. M. & Harris, J. P. (1992). Effects of bacterial endotoxin applied to the guinea pig cochlea. Laryngoscope 102, 683– 688. Davis, A., Wood, S., Healy, R., Webb, H. & Rowe, S. (1995). Risk factors for hearing disorders: epidemiologic evidence of change over time in the UK. J Am Acad Audiol 6, 365–370. DeMaria, T. F., Prior, R. B., Briggs, B. R., Lim, D. J. & Birck, H. G. (1984). Endotoxin in middle-ear effusion from patients with chronic otitis media with effusion. J Clin Microbiol 20, 15–17. Engel, F., Blatz, R., Kellner, J., Palmer, M., Weller, U. & Bhadki, S. (1995). Breakdown of the round window membrane permeability barrier evoked by streptolysin O: possible etiologic role in development of sensorineural hearing loss in acute otitis media. Infect Immun 63, 1305–1310. Erenberg, A., Lemons, J., Sia, C., Trunkel, D. & Ziring, P. (1999). Newborn and infant hearing loss: detection and intervention. American Academy of Pediatrics. Task Force on Newborn and Infant Hearing, 1998-1999. Pediatrics 103, 527–530. Fessenden, J. D. & Schacht, J. (1998). The nitric oxide/cyclic GMP pathway: a potential major regulator of cochlear physiology. Hear Res 118, 168–176. Forstermann, U., Closs, E. I., Pollock, J. S., Nakane, M., Schwarz, P., Gath, I. & Kleinert, H. (1994). Nitric oxide synthase isozymes. Characterization, purification, molecular cloning, and functions. Hypertension 23, 1121–1131. Garcia Berrocal, J. R. & Ramirez-Camacho, R. (2000). Immune response and immunopathology of the inner ear: an update. J Laryngol Otol 114, 101–107. Goldberg, B., Goycoolea, M. V., Schleivert, P. M., Shea, D., Schachern, P., Paparella, M. M. & Carpenter, A. M. (1981). Passage of albumin from the middle ear to the inner ear in otitis media in the chinchilla. Am J Otolaryngol 2, 210–214. Goldenberg, R. L., Hauth, J. C. & Andrews, W. W. (2000). Intrauterine infection and preterm delivery. N Engl J Med 342, 1500–1507. Goycoolea, M. V. (2001). Clinical aspects of round window membrane permeability under normal and pathological conditions. Acta Otolaryngol 121, 437–447. Goycoolea, M. V., Paparella, M. M., Juhn, S. K. & Carpenter, A. M. (1980). Oval and round window changes in otitis media. Potential References pathways between middle and inner ear. Laryngoscope 90, 1387– 1391. Abramovich, S. J., Gregory, S., Slemick, M. & Stewart, A. (1979). Goycoolea, M. V., Muchow, D. & Schachern, P. (1988). Experimental Hearing loss in very low birthweight infants treated with neonatal intensive care. Arch Dis Child 54, 421–426. studies on round window structure: function and permeability. Laryngoscope 98, 1–20. Amaee, F. R., Comis, S. D., Osborne, M. P., Drew, S. & Tarlow, M. J. (1997). Possible involvement of nitric oxide in the sensorineural Gross, M., Finckh-Kramer, U. & Spormann-Lagodzinski, M. (2000). hearing loss of bacterial meningitis. Acta Otolaryngol 117, 329–336. Congenital hearing disorders in children. 1: Acquired hearing disorders. HNO 48, 879–886. Bernstein, J. M., Praino, M. D. & Neter, E. (1980). Detection of Guo, Y., Wu, Y., Chen, W. & Lin, J. (1994). Endotoxic damage to the endotoxin in ear specimens from patients with chronic otitis media by means of the limulus amebocyte lysate test. Can J Microbiol 26, 546–548. stria vascularis: the pathogenesis of sensorineural hearing loss secondary to otitis media? J Laryngol Otol 108, 310–313. Cock, M. L., Dalitz, P. A. & Harding, R. (2002). Reduced placental perfusion following endotoxin administration to the preterm ovine fetus. Pediatr Res 51, 450. Comis, S. D., Osborne, M. P. & Tarlow, M. J. (1991). The effect of bacterial endotoxin upon the morphology of the tectorial membrane and stereocilia in the guinea pig cochlea. Scanning Microsc 5, 1129– 1133. Cureoglu, S., Schachern, P. A., Rinaldo, A., Tsuprun, V., Ferlito, A. & Paparella, M. M. (2005). Round window membrane and labyrinthine pathological changes: an overview. Acta Otolaryngol 125, 9–15. http://jmm.sgmjournals.org Guzik, T. J., Korbut, R. & Adamek-Guzik, T. (2003). Nitric oxide and superoxide in inflammation and immune regulation. J Physiol Pharmacol 54, 469–487. Harris, J. P., Heydt, J., Keithley, E. M. & Chen, M. C. (1997). Immunopathology of the inner ear: an update. Ann N Y Acad Sci 830, 166–178. Hess, A., Bloch, W., Arnhold, S., Andressen, C., Stennert, E., Addicks, K. & Michel, O. (1998). Nitric oxide synthase in the vestibulocochlear system of mice. Brain Res 813, 97–102. Hess, A., Bloch, W., Huverstuhl, J., Su, J., Stennert, E., Addicks, K. & Michel, O. (1999). Expression of inducible nitric oxide synthase Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 16 Jun 2017 14:06:22 381 A. L. Smit and others (iNOS/NOS II) in the cochlea of guinea pigs after intratympanical endotoxin-treatment. Brain Res 830, 113–122. Iadecola, C., Xu, X., Zhang, F., el-Fakahany, E. E. & Ross, M. E. (1995a). Marked induction of calcium-independent nitric oxide synthase activity after focal cerebral ischemia. J Cereb Blood Flow Metab 15, 52–59. (2009b). Intravenous lipopolysaccharide-induced pulmonary maturation and structural changes in fetal sheep. Am J Obstet Gynecol 200, 195.e1–195.e10. Lee, S. E., Romero, R., Jung, H., Park, C. W., Park, J. S. & Yoon, B. H. (2007). The intensity of the fetal inflammatory response in Iadecola, C., Zhang, F., Xu, S., Casey, R. & Ross, M. E. (1995b). intraamniotic inflammation with and without microbial invasion of the amniotic cavity. Am J Obstet Gynecol 197, 294.e1–294.e6. Inducible nitric oxide synthase gene expression in brain following cerebral ischemia. J Cereb Blood Flow Metab 15, 378–384. Li, L., Kosugi, I., Han, G. P., Kawasaki, H., Arai, Y., Takeshita, T. & Tsutsui, Y. (2008). Induction of cytomegalovirus-infected labyrinthi- Iino, Y., Kaneko, Y. & Takasaka, T. (1985). Endotoxin in middle ear tis in newborn mice by lipopolysaccharide: a model for hearing loss in congenital CMV infection. Lab Invest 88, 722–730. effusions tested with Limulus assay. Acta Otolaryngol 100, 42–50. Jobe, A. H., Newnham, J. P., Willet, K. E., Sly, P., Ervin, M. G., Bachurski, C., Possmayer, F., Hallman, M. & Ikegami, M. (2000). Liaudet, L., Soriano, F. G. & Szabo, C. (2000). Biology of nitric oxide Effects of antenatal endotoxin and glucocorticoids on the lungs of preterm lambs. Am J Obstet Gynecol 182, 401–408. Lim, D. J., Kawauchi, H. & DeMaria, T. F. (1990). Role of middle ear Juhn, S. K., Hamaguchi, Y. & Goycoolea, M. (1989). Review of round window membrane permeability. Acta Otolaryngol Suppl 457, 43–48. Jung, T. T., Llaurado, R. J., Nam, B. H., Park, S. K., Kim, P. D. & John, E. O. (2003). Effects of nitric oxide on morphology of isolated signaling. Crit Care Med 28, N37–N52. endotoxin in inner ear inflammatory response and hydrops: longterm study. Ann Otol Rhinol Laryngol Suppl 148, 33–34. Ludwig, A. & Hengel, H. (2009). Epidemiological impact and disease burden of congenital cytomegalovirus infection in Europe. Euro Surveill 14, 26–32. cochlear outer hair cells: possible involvement in sensorineural hearing loss. Otol Neurotol 24, 682–685. Marlow, E. S., Hunt, L. P. & Marlow, N. (2000). Sensorineural hearing Kallapur, S. G., Jobe, A. H., Ball, M. K., Nitsos, I., Moss, T. J., Hillman, N. H., Newnham, J. P. & Kramer, B. W. (2007). Pulmonary and Mencher, G. T. (2000). Challenge of epidemiological research in the systemic endotoxin tolerance in preterm fetal sheep exposed to chorioamnionitis. J Immunol 179, 8491–8499. loss and prematurity. Arch Dis Child Fetal Neonatal Ed 82, F141–F144. developing world: overview. Audiology 39, 178–183. Kastenbauer, S., Klein, M., Koedel, U. & Pfister, H. W. (2001). Moncada, S., Palmer, R. M. & Higgs, E. A. (1991). Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacol Rev 43, 109–142. Reactive nitrogen species contribute to blood-labyrinth barrier disruption in suppurative labyrinthitis complicating experimental pneumococcal meningitis in the rat. Brain Res 904, 208–217. Morizono, T. & Ikeda, K. (1990). Effect of Escherichia coli endotoxin on cochlear potentials following its application to the chinchilla middle ear. Eur Arch Otorhinolaryngol 247, 40–42. Katano, H., Sato, Y., Tsutsui, Y., Sata, T., Maeda, A., Nozawa, N., Inoue, N., Nomura, Y. & Kurata, T. (2007). Pathogenesis of Moss, T. J., Nitsos, I., Newnham, J. P., Ikegami, M. & Jobe, A. H. (2003). Chorioamnionitis induced by subchorionic endotoxin cytomegalovirus-associated labyrinthitis in a guinea pig model. Microbes Infect 9, 183–191. Kawauchi, H., DeMaria, T. F. & Lim, D. J. (1989). Endotoxin permeability through the round window. Acta Otolaryngol Suppl 457, 100–115. infusion in sheep. Am J Obstet Gynecol 189, 1771–1776. Mussi-Pinhata, M. M., Yamamoto, A. Y., Moura Brito, R. M., de Lima Isaac, M., de Carvalho e Oliveira, P. F., Boppana, S. & Britt, W. J. (2009). Birth prevalence and natural history of congenital cyto- Kim, C. S. & Kim, H. J. (1995). Auditory brain stem response changes megalovirus infection in a highly seroimmune population. Clin Infect Dis 49, 522–528. after application of endotoxin to the round window membrane in experimental otitis media. Otolaryngol Head Neck Surg 112, 557–565. Mustafa, M. M., Ramilo, O., Olsen, K. D., Franklin, P. S., Hansen, E. J., Beutler, B. & McCracken, G. H., Jr (1989). Tumor necrosis factor in Klein, M., Koedel, U., Kastenbauer, S. & Pfister, H. W. (2008). mediating experimental Haemophilus influenzae type B meningitis. J Clin Invest 84, 1253–1259. Nitrogen and oxygen molecules in meningitis-associated labyrinthitis and hearing impairment. Infection 36, 2–14. Kramer, B. W. (2008). Antenatal inflammation and lung injury: Nakai, Y., Morimoto, A., Chang, K. C., Yamanaka, M. & Nishisato, K. (1980). Inner ear damage induced by bacterial endotoxin. Arch prenatal origin of neonatal disease. J Perinatol 28, S21–S27. Otorhinolaryngol 229, 209–220. Kramer, B. W., Ikegami, M., Moss, T. J., Nitsos, I., Newnham, J. P. & Jobe, A. H. (2005a). Endotoxin-induced chorioamnionitis modulates Nance, W. E., Lim, B. G. & Dodson, K. M. (2006). Importance of innate immunity of monocytes in preterm sheep. Am J Respir Crit Care Med 171, 73–77. Kramer, B. W., Kaemmerer, U., Kapp, M., Herbst, D., Marx, A., Berg, D., Groneck, P. A. & Speer, C. P. (2005b). Decreased expression of angiogenic factors in placentas with chorioamnionitis after preterm birth. Pediatr Res 58, 607–612. Kramer, B. W., Joshi, S. N., Moss, T. J., Newnham, J. P., Sindelar, R., Jobe, A. H. & Kallapur, S. G. (2007). Endotoxin-induced maturation of monocytes in preterm fetal sheep lung. Am J Physiol Lung Cell Mol Physiol 293, L345–L353. Kramer, B. W., Kallapur, S., Newnham, J. & Jobe, A. H. (2009a). Prenatal inflammation and lung development. Semin Fetal Neonatal Med 14, 2–7. Kramer, B. W., Ladenburger, A., Kunzmann, S., Speer, C. P., Been, J. V., van Iwaarden, J. F., Zimmermann, L. J., Gantert, M. & Garnier, Y. 382 congenital cytomegalovirus infections as a cause for pre-lingual hearing loss. J Clin Virol 35, 221–225. Newnham, J. P., Moss, T. J., Kramer, B. W., Nitsos, I., Ikegami, M. & Jobe, A. H. (2002). The fetal maturational and inflammatory responses to different routes of endotoxin infusion in sheep. Am J Obstet Gynecol 186, 1062–1068. Nitsos, I., Moss, T. J., Cock, M. L., Harding, R. & Newnham, J. P. (2002). Fetal responses to intra-amniotic endotoxin in sheep. J Soc Gynecol Investig 9, 80–85. Paparella, M. M., Brady, D. R. & Hoel, R. (1970). Sensori-neural hearing loss in chronic otitis media and mastoiditis. Trans Am Acad Ophthalmol Otolaryngol 74, 108–115. Ramilo, O., Mustafa, M. M., Saez-Llorens, X., Mertsola, J., Ohkawara, S., Yoshinaga, M., Hansen, E. J. & McCracken, G. H., Jr (1989). Role of interleukin 1-beta in meningeal inflammation. Pediatr Infect Dis J 8, 909–910. Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 16 Jun 2017 14:06:22 Journal of Medical Microbiology 59 Potential role for LPS in congenital hearing loss Romero, R. & Mazor, M. (1988). Infection and preterm labor. Clin Obstet Gynecol 31, 553–584. Trinidad, A., Ramirez-Camacho, R., Garcia-Berrocal, J. R., Verdaguer, J. M. & Daza, R. (2005). Labyrinthitis secondary to Sampson, J. E., Theve, R. P., Blatman, R. N., Shipp, T. D., Bianchi, D. W., Ward, B. E. & Jack, R. M. (1997). Fetal origin of amniotic fluid Wang, X., Rousset, C. I., Hagberg, H. & Mallard, C. (2006). polymorphonuclear leukocytes. Am J Obstet Gynecol 176, 77–81. Schachern, P. A., Paparella, M. M., Hybertson, R., Sano, S. & Duvall, A. J., III (1992). Bacterial tympanogenic labyrinthitis, meningitis, and sensorineural damage. Arch Otolaryngol Head Neck Surg 118, 53–57. experimental otitis media. Am J Otolaryngol 26, 226–229. Lipopolysaccharide-induced inflammation and perinatal brain injury. Semin Fetal Neonatal Med 11, 343–353. Watanabe, K., Naito, N. & Tanaka, Y. (1995). Morphological changes Schmidt, H. H. & Walter, U. (1994). NO at work. Cell 78, 919–925. in the inner ear induced by bacterial endotoxin. Med Electron Microsc 28, 80–87. Shi, X., Dai, C. & Nuttall, A. L. (2003). Altered expression of inducible Watanabe, K., Hess, A., Bloch, W. & Michel, O. (2000a). Inhibition of nitric oxide synthase (iNOS) in the cochlea. Hear Res 177, 43–52. Spandow, O., Anniko, M. & Moller, A. R. (1988). The round window as access route for agents injurious to the inner ear. Am J Otolaryngol 9, 327–335. Spandow, O., Anniko, M. & Hellstrom, S. (1989). Inner ear disturbances following inoculation of endotoxin into the middle ear. Acta Otolaryngol 107, 90–96. Spandow, O., Hellstrom, S. & Anniko, M. (1990). Structural changes in the round window membrane following exposure to Escherichia coli lipopolysaccharide and hydrocortisone. Laryngoscope 100, 995–1000. Suppiej, A., Franzoi, M., Vedovato, S., Marucco, A., Chiarelli, S. & Zanardo, V. (2009). Neurodevelopmental outcome in preterm histological chorioamnionitis. Early Hum Dev 85, 187–189. Suzuki, M., Yamasoba, T. & Kaga, K. (1998). Development of the blood-labyrinth barrier in the rat. Hear Res 116, 107–112. Takumida, M. & Anniko, M. (1998). Lipopolysaccharide-induced expression of nitric oxide synthase II in the guinea pig vestibular end organ. Eur Arch Otorhinolaryngol 255, 184–188. Takumida, M. & Anniko, M. (2004a). Functional significance of nitric oxide in the inner ear. In Vivo 18, 345–350. Takumida, M. & Anniko, M. (2004b). Localization of endotoxin in the inner ear following inoculation into the middle ear. Acta Otolaryngol 124, 772–777. inducible nitric oxide synthase lowers the cochlear damage by lipopolysaccharide in guinea pigs. Free Radic Res 32, 363–370. Watanabe, K., Hess, A., Zumeger, C., Stennert, E., Bloch, W., Addicks, K. & Michel, O. (2000b). Changes of the compound action potential (CAP) and the expression of inducible nitric oxide synthase (iNOS/NOS II) in the cochlea under the inflammatory condition. Hear Res 145, 149–155. Watanabe, K., Jinnouchi, K., Pawankar, R. & Yagi, T. (2001). Expression of myeloperoxidase and cochlear dysfunction in the lipopolysaccharide-treated guinea pig. Eur Arch Otorhinolaryngol 258, 164–167. Watanabe, K., Jinnouchi, K. & Yagi, T. (2002). Immunoreactivity for myeloperoxidase (MPO) in the vestibule after the injection of bacterial lipopolysaccharide into the middle ear. Auris Nasus Larynx 29, 241–245. Wild, N. J., Sheppard, S., Smithells, R. W., Holzel, H. & Jones, G. (1989). Onset and severity of hearing loss due to congenital rubella infection. Arch Dis Child 64, 1280–1283. Yoon, B. H., Romero, R., Park, J. S., Kim, M., Oh, S. Y., Kim, C. J. & Jun, J. K. (2000). The relationship among inflammatory lesions of the umbilical cord (funisitis), umbilical cord plasma interleukin 6 concentration, amniotic fluid infection, and neonatal sepsis. Am J Obstet Gynecol 183, 1124–1129. Young, T. E. & Magnum, O. B. (2007). Neofax: a Manual of Drugs used Takumida, M., Anniko, M., Popa, R. & Zhang, D. M. (2000). in Neonatal Care, pp. 46–58. Montvale, NJ: Thomson. Lipopolysaccharide-induced expression of inducible nitric oxide synthase in the guinea pig organ of Corti. Hear Res 140, 91–98. Zdanski, C. J., Prazma, J., Petrusz, P., Grossman, G., Raynor, E., Smith, T. L. & Pillsbury, H. C. (1994). Nitric oxide synthase is an active Tarlow, M. J., Comis, S. D. & Osborne, M. P. (1991). Endotoxin induced damage to the cochlea in guinea pigs. Arch Dis Child 66, 181–184. enzyme in the spiral ganglion cells of the rat cochlea. Hear Res 79, 39– 47. http://jmm.sgmjournals.org Downloaded from www.microbiologyresearch.org by IP: 88.99.165.207 On: Fri, 16 Jun 2017 14:06:22 383