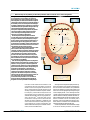
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
REVIEWS DOES THE RIBOSOME TRANSLATE CANCER? Davide Ruggero and Pier Paolo Pandolfi Ribosome biogenesis and translation control are essential cellular processes that are governed at numerous levels. Several tumour suppressors and proto-oncogenes have been found either to affect the formation of the mature ribosome or to regulate the activity of proteins known as translation factors. Disruption in one or more of the steps that control protein biosynthesis has been associated with alterations in the cell cycle and regulation of cell growth. Therefore, certain tumour suppressors and proto-oncogenes might regulate malignant progression by altering the protein synthesis machinery. Although many studies have correlated deregulation of protein biosynthesis with cancer, it remains to be established whether this translates directly into an increase in cancer susceptibility, and under what circumstances. NUCLEOLUS The nucleolus is a suborganelle of the nucleus where rRNA is transcribed, processed and assembled into ribosomal subumits. Ribosomal proteins are added to rRNAs within the nucleolus. Molecular Biology Program, Department of Pathology, Memorial Sloan-Kettering Cancer Center, Sloan-Kettering Institute, 1275 York Avenue, New York, New York 10021, USA. Correspondence to P. P. P. e-mail: [email protected] doi:10.1038/nrc1015 NATURE REVIEWS | C ANCER The production of mature ribosomes that are competent to translate cellular mRNAs requires a multistep process that is highly coordinated in eukaryotic cells. The ribosome, which is the central protein synthesis factory of the cell, is often viewed as a finely tuned machine that functions as a static, reliable component of higher-order cellular processes. In fact, the ribosome has the overwhelming burden of correctly and efficiently producing all proteins in the cell. Although it has long been known that, in cancer cells, components of the translation machinery are deregulated or misexpressed, their role in tumorigenesis has largely been overlooked. For example, as early as 1970, changes in the NUCLEOLUS were recognized as a reliable marker of cellular transformation1. However, these findings were dismissed as non-consequential to the overall transformation process. After all, how could the ribosome, which is required for such a crucial cellular function, be tweaked or perturbed without causing death to the cell? An exciting body of research is now challenging this concept. In the past, the protein synthesis machinery was viewed as a static entity, but it is now becoming clear that its activity is highly regulated. In fact, several proto-oncogenes and tumour suppressors have recently been shown to directly regulate ribosome production or the initiation of protein translation, or both (TABLE 1). These findings raise the possibility that alterations in components of the protein synthesis machinery and aberrant regulation of translation can promote cellular transformation. The goal now is to determine how and to what extent ribosome function is directly modulated by tumour suppressors and oncogenes, and whether this represents a cause or consequence of cancer progression. Mutations in genes that encode proteins that are directly involved in ribosome biogenesis have also been associated with cancer and human disease. The DKC1 gene is mutated in patients with dyskeratosis congenita, a disease that is characterized by premature ageing and an increased susceptibility to cancer2,3. DKC1 encodes dyskerin, a pseudouridine synthase that mediates posttranscriptional modification of ribosomal RNA. Mutations in the gene that encodes ribosomal protein S19 have been identified in another syndrome that is characterized by increased susceptibility to cancer — Diamond–Blackfan anaemia4. But do mutations in these genes directly result in impaired mRNA translation? Is this a quantitative or qualitative translation problem? How can defects in protein synthesis result in a specific disease phenotype or carcinogenesis? Although there are many questions to be answered, it is now becoming evident that regulation of ribosome function can be lost in cancer cells. VOLUME 3 | MARCH 2003 | 1 7 9 REVIEWS All roads lead to rRNA synthesis Cell growth and proliferation are associated with changes in the rate of ribosome production. During G1, an increase in rRNA synthesis and ribosome assembly is a prerequisite for increased protein synthesis during S phase5. Furthermore, downregulation in ribosome activity or ribosome formation, or both, could be required during M phase to ensure proper exit from the cell cycle6,7. Therefore, an important relationship exists between the cell cycle and ribosome production. This balance is maintained in the cell through key checkpoints, which ensure that translation of mRNAs occurs at appropriate levels and during a specific window of the cell cycle. As will be discussed below, in cancer cells this balance can be broken, resulting in deregulation of rRNA synthesis. The synthesis of rRNA is the first event in ribosome biogenesis. It is dependent on RNA polymerase I (Pol I) regulation of rDNA transcription in the nucleolus (BOX 1). The synthesis of rRNA in the cell can be induced by extracellular stimuli at moments when a cell needs to grow and proliferate. The concept of regulated rRNA synthesis was originally shown in cells in which deprivation of a single amino acid resulted in a rapid termination of rRNA synthesis8. Since then, many other reports have shown that the initiation of rRNA transcription is tightly linked to cell-cycle progression. Synthesis of Summary • Ribosome biogenesis and translation are regulated at multiple levels and are associated with accurate cell growth and proliferation. The loss of key checkpoints during protein synthesis might contribute to the initiation and progression of cancer. • During specific phases of the cell cycle, the synthesis of rRNA, as well as components of the protein synthesis machinery, is initiated by the phosphorylation of key transcription factors that regulate polymerase I (Pol I) and Pol III activity, respectively. This tight link between cell-cycle progression and protein synthesis exists to ensure accurate cell growth and proliferation, which might be lost in cancer cells. • p53 and retinoblastoma (RB) repress Pol I and Pol III transcription. In cancer cells, which harbour inactivating mutations in these tumour suppressors, deregulation of Pol I and Pol III activity might contribute to tumorigenesis. • Several ribosomal proteins are overexpressed in a variety of tumours. It remains to be determined whether this represents a cause or consequence of tumour formation. Increased phosphorylation of the S6 ribosomal protein is thought to result in enhanced translation of specific mRNAs. This raises the possibility that deregulation of ribosomal proteins in tumours might affect the translation of specific target mRNAs. • MYC and PTEN act as master regulators of ribosome biogenesis and translation control. Their deregulation in tumour cells increases the expression and activity of components of the translation apparatus. It remains to be determined which of the downstream targets of MYC and PTEN involved in controlling protein synthesis are directly responsible for tumour susceptibility. • Further investigation will be needed to clarify to what extent deregulation in total or specific translation of mRNAs contributes to tumorigenesis. Although mutations in genes that are directly responsible for ribosome biogenesis, such as those encoding the ribosomal protein S19 and DKC1 (the enzyme that modifies rRNA), have been found in cancer susceptibility syndromes, the molecular mechanisms by which these proteins cause cancer remain largely unknown. • Components of the translation machinery that are overexpressed or deregulated in cancer cells could represent targets for cancer therapy. The macrolide rapamycin, which affects the translation machinery, has already been used in clinical trials as a tumour inhibitory agent. 180 | MARCH 2003 | VOLUME 3 rRNA is maximal in S and G2 phases, repressed in mitosis and increased in G1 (REFS 7,9,10). These fluctuations in cell-cycle-mediated rRNA synthesis are dependent on the activity of Pol I. The transcription factor UBF (upstream binding factor) is a key regulator of rRNA synthesis owing to its ability to modulate Pol I transcriptional activity11–13. Furthermore, several proto-oncogenes and tumour suppressors directly regulate rRNA synthesis by potentiating or repressing UBF activity, respectively (FIG. 1). UBF binds to two regions of the rDNA promoter — the upstream control element (UCE) and the core — which are also recognized by Pol I (REFS 14–16). Phosphorylation of UBF upregulates rRNA synthesis in response to external stimuli (FIG. 1). The amount of prerRNA in cells that express a mutant non-phosphorylatable form of UBF is twofold lower than in cells that express wild-type UBF, with a concomitant reduction in cell growth17. Different kinases phosphorylate UBF at serine and threonine residues positioned at the carboxyterminal and internal regions of the protein. The modification of UBF predominately affects its ability to bind to DNA and/or to mediate protein–protein interactions that are required for assembly of an active initiation complex18–21. Importantly, phosphorylation of UBF is directly dependent on mitogen stimuli and cell-cycle progression, and, in quiescent cells, UBF is hypophosphorylated and transcriptionally inactive22–24. UBF phosphorylation therefore represents a point of convergence for many cell-cycle-regulated proteins in modulated rRNA synthesis. It could also represent a potential target for proto-oncogenes, some of which have been shown to directly regulate UBF activity. The first protein identified to regulate UBF activity was casein kinase II (CKII)23 — a serine threonine kinase that is overexpressed in many cancers, including leukaemias and solid tumours. CKII has been proposed to contribute to tumorigenesis through direct interactions with the cell-cycle machinery. In addition, it phosphorylates UBF at its carboxyl terminus and thereby regulates rDNA transcription. Subsequent studies have identified other UBF kinases that are also deregulated in cancer and similarly affect rRNA synthesis. The complexes that are formed by G1-specific cyclin-dependent kinases and cyclins — CDK4–cyclin-D1 and CDK2–cyclin-E — directly phosphorylate UBF at serine 484 and 388, facilitating the interaction between UBF and Pol I and so controlling rRNA synthesis in a cell-cycleregulated manner17,20. The CDK–cyclin complex is overexpressed by human neoplasias. Transgenic mice that express proteins that mimic the human mutant forms also develop tumours 25,26. Would alterations in rRNA synthesis have a consequence for cancer progression, and, if so, what would be the underlying mechanism? The above-mentioned kinases all mediate phosphorylation of UBF in a specific window of the cell cycle. Although all these kinases are overexpressed in cancer, the exact implications of upregulation in rRNA synthesis on the transformation process are unclear. If cancer cells have lost cell-cycle control, upregulation in rRNA synthesis www.nature.com/reviews/cancer REVIEWS Table 1 | Translation control and cancer Gene Syndrome or tumour type Function in protein synthesis TP53 Wide range of tumours Regulates Pol I and Pol III activity RB Retinoblastoma; a wide range of tumours Regulates Pol I and Pol III activity NPM Haematological malignancies; overexpressed in a wide range of tumours Ribosome biogenesis PTEN Cowden disease; a wide range of tumours Regulates mTOR kinase; important for phosphorylation of S6 and 4EBP1 Ribosomal protein Overexpressed in a wide range of tumours Ribosome biogenesis S19 Diamond–Blackfan anaemia; haematological malignancies; myelodysplastic syndrome Ribosome biogenesis S6 NA Translation of 5′ TOP mRNAs DKC1 Dyskeratosis congenita; haematological malignancies; carcinomas Modification of rRNA mTOR, mammalian target of rapamycin; NA, not applicable; NPM, nucleophosmin; Pol, polymerase; RB, retinoblastoma; TOP, terminal oligopyrimidine tract. would simply be a readout of upregulated cellular proliferation. In neoplasia, however, upregulation of UBF activity has been shown to arise from direct extrastimulatory stimuli. This represents an instance in which upregulation of rRNA synthesis does not simply reflect cell-cycle progression, but, rather, UBF serves as a stimulus that might directly activate the translational machinery to initiate tumorigenesis. Little is known about the molecular mechanism by which an extracellular cue could directly regulate Pol I activity and augment rRNA synthesis. The first indication of the way in which specific growth factors could regulate rRNA synthesis came from the finding that epidermal growth factor (EGF) signalling via extracellular-signal-regulated kinase (ERK) activation could directly regulate UBF activity21. The ERK1/2 kinases were shown to phosphorylate UBF at two threonine residues, which resulted in an immediate upregulation of endogenous ribosomal transcription. This finding places ribosomal transcription under the direct control of extracellular growth signals. Given the finding that ERK is upregulated in many cancers27, the early cellular response to upregulate rRNA synthesis might prove to be a mechanism by which ERK mediates cellular transformation. The phosphorylation of UBF by ERK might be one mechanism by which rRNA synthesis is controlled by extracellular growth signals. However, given that shutdown of rRNA synthesis during mitosis and early G1 was shown to be due to the inactivation of UBF by its dephosphorylation10, the ability of kinases to upregulate rRNA synthesis would be limited to a specific window of the cell cycle. The determination of the phosphatase that is directly responsible for mediating the mitotic repression of UBF activity will undoubtedly shed light on an important checkpoint within the cell, as this phosphatase would prevent inappropriate ribosome biogenesis during cell division. NATURE REVIEWS | C ANCER Some evidence exists to indicate that the tumoursuppressor protein phosphatase (PP2A) mediates dephosphorylation of UBF10. Deregulation of this phosphatase in cancer cells could have an even more profound effect in disrupting the coordination between rRNA synthesis and cell-cycle progression. Fluctuations in rRNA synthesis, which occur in various conditions that affect cell growth, also correlate with the activity of the transcription initiation factor TIF-IA. Mammalian TIF-IA serves as a bridge between Pol I and the pre-initiation complex on the rDNA promoter28,29 (FIG. 1). Remarkably, cellular lysates from cells that arrest in the cell cycle by confluent inhibition, and are starved of amino acids or treated with cyclohexamide, are incapable of efficiently transcribing rDNA. Addition of recombinant TIF-IA to cells, however, completely restores transcriptional activity29. Given that TIF-IA is phosphorylated at multiple sites by different extracellular signals29, the identification of the kinases that mediate its phosphorylation will be an exciting avenue for future research. It would shed light on how mitotic stimuli, which are relevant in cancer progression, directly regulate rRNA synthesis. Regulation of protein synthesis by RB and p53 The family of retinoblastoma proteins (RB) have also been shown to regulate rRNA synthesis through UBF (FIG. 1). The RB gene is frequently disrupted in human tumours. Its function as tumour suppressor has been attributed to its ability to regulate the cell cycle, which involves modulation of the transcriptional activity of E2F 30. In addition, several findings indicate that RB regulates cell growth and proliferation by restricting the production of rRNA11. It is accomplished by the direct association of RB with UBF. This interaction prevents UBF from recruiting other cofactors that are required for Pol I activity, such as TIF-IB/SL1(REFS 31,32).Another RB-family member, p130, also represses rRNA transcription through its ability to bind and inactivate UBF 33. Three lines of evidence have highlighted the potential importance of the interaction of UBF and RBfamily members in controlling cell proliferation. Immunofluorescence studies have shown that RB accumulates in the nucleolus when cells differentiate or downregulate Pol I transcription, or both 11,34. Overexpression of RB can repress the rDNA promoter both in vitro and in vivo. Furthermore, rRNA concentrations are abnormally raised in cells that lack the entire RB family of proteins, or a combination of RB and p130 (REF. 33). Cells that lack both RB and p130 do not show significant differences in cell-cycle progression. Therefore, an upregulation of rRNA in this case cannot be explained simply as a consequence of increased cell-cycle progression. These results show that the effects of RB on cell growth can be discriminated from its ability to regulate UBF activity. Similar to RB and p130, the tumour suppressor p53 has been shown to repress Pol I transcription by directly interfering with the assembly of a protein complex that is required for transcriptional initiation on the rRNA promoter35 (FIG. 1). Wild-type p53 can suppress Pol I VOLUME 3 | MARCH 2003 | 1 8 1 REVIEWS transcription in cotransfection assays, and p53-null cells display increased Pol I transcriptional activity compared with wild-type cells35. Because RB-inactivating mutations also occur in tumour cells that harbour p53 mutations36, the combined effect of both mutations on Pol I activity might have a synergistic effect on tumour progression. Although beyond the scope of this review, p53 and RB-family members have also been shown to control RNA polymerase III (Pol III) transcription37–39 (FIG. 1). Pol III is responsible for synthesizing various small stable RNAs that include components of ribosome, such as 5′SrRNA, as well as tRNA. In p53 and RB-null cells, Pol III transcription is increased with respect to wildtype cells. Both of these tumour suppressors negatively regulate Pol-III-mediated transcription through direct, inactivating interactions with TF-IIIB — a co-activator complex that is responsible for Pol-III-mediated transcription. Therefore, loss of RB and p53 in tumours could result in increased cell proliferation through aberrant upregulation in essential components of the protein synthesis machinery. Several tumour-associated RB mutations specifically interfere with the ability of RB to regulate Pol I or Pol III activity, or both. A RB mutation found in small-cell lung carcinoma that results in a single amino acid change prevents RB interaction with UBF and repression of Pol I transcription11,40. In addition, several mutations in RB lead to deletions of regions that control Pol III activity, and the ability of RB to repress Pol III activity was shown to be lost in these tumours41. So, the ability of RB to directly suppress the level of protein synthesis by controlling the availability of tRNA and rRNA might be compromised in several tumour lesions. How can increases in Pol I and Pol III activity be directly linked to cancer? It has been known for more than 25 years that the rate of cellular proliferation and growth is proportional to the rate of protein synthesis42,43. Translation is dependent on the availability of synthesized tRNA and rRNA. Both RB-family members and p53 could exert some aspects of their tumour-suppressor properties through their ability to suppress Pol I and Pol III transcriptional activity and, subsequently, the level of protein synthesis. In addition, the upregulation in cancer of the UBF kinases described above could stimulate rRNA synthesis and contribute to their oncogenic properties. Disruption of protein synthesis control could render the cells more susceptible to deregulated cell growth and proliferation. It remains to be determined to what extent deregulation of Pol I and Pol III transcription contributes to tumour initiation or progression, or both. Box 1 | Making ribosomes UCE Core ψ and CH3 modification SnoRNPs ψ ψ ψ ITS 1 ψ ITS 2 ψ 18S 5.8S 45S 5′ETS CH3 CH3 ψ ψ ψ 28S CH3 CH3 CH3 CH3 3′ETS 41S 36S 32S RNases 18S 5.8S ψ ψ ψ 28S S+ CH3 ψ ψ 18S CH3 CH3 ψ ψ ψ ψ ψ CH3 5.8S ψ 5S CH3 ψ L S ψ + CH3 28S CH 3 Ribosomal proteins L 12S ψ | MARCH 2003 | VOLUME 3 UBF TIF-IB/SL1 ψ 182 POL I TIF-IA 3 ψ CH All ribosomal RNAs, with the exception of the 5S, are transcribed as a polycistronic transcript known as preribosomal RNA (pre-rRNA or 45S) in the nucleolus46. Transcription of rDNA is dependent on three basal transcription factors: the ‘selectivity complex’ (SL1 or TIFIB), the HMG1 box architectural upstream binding factor (UBF) and the DNA-dependent RNA polymerase I (Pol I). Both UBF and SL1 make direct contacts with a 150-bp region of DNA in the rRNA gene promoter, which contains two sequence elements known as the upstream control element (UCE) and the core. In the case of UBF, it probably binds to these promoter elements as a dimer. UBF is thought to bind the promoter first, enabling subsequent recruitment of SL1 and Pol I. The initiation factor TIF-IA is also part of this complex. Concomitant with rRNA transcription, the rRNA sequences are extensively modified134. Specifically, a large family of small nucleolar RNAs (snoRNAs) guides the sitespecific conversion of uridine to pseudouridine (Ψ) in rRNA. This is accomplished through direct base-pairing of snoRNAs with specific rRNA sequences, leaving a single uridine exposed to the enzymatic activity of dyskerin, the pseudouridine synthase that mediates the modification of this residue. In addition, other snoRNAs also guide the formation of 2′-0-methylated nucleosides (-CH3) in rRNA. The pre-rRNA precursor is then cleaved at specific sites by RNases to produce a series of characteristic intermediates (41S), and finally mature rRNAs — 18S, 5.8S and 28S. During rRNA processing, the rRNA species must associate with more than 70 ribosomal proteins, as well as the 5S rRNA in the nucleolus, to form the small (S, 40S) and the large (L, 60S) ribosomal subunits, which are assembled and transported to the cytoplasm to initiate protein synthesis. CH3 www.nature.com/reviews/cancer REVIEWS 5S tRNA snRNAs POL III RB p53 TFIIIB TFIIIC2 P TIF-1A P POL I rRNA TIF-1B/SL1 UBF 18S 5.8S 28S P P ERK CDK2/4 CKII Figure 1 | Regulation of Pol I and Pol III activity by tumour suppressors and proto-oncogenes. The tumour suppressors retinoblastoma (RB) and p53 negatively regulate RNA polymerase (Pol I) and Pol III complex activity. Pol I activity transcribes rRNA, whereas Pol III transcribes a class of small RNAs that includes the 5S rRNA, small nuclear RNAs (snRNAs) and tRNA. Both RB and p53 interfere with the assembly of transcriptional machinery on the rRNA promoter by inhibiting the interaction between TIF-IB/SL1 and upstream binding factor (UBF). In the case of p53, the interaction of SL1 and UBF is destabilized by the ability of p53 to bind to SL1, whereas RB directly interacts with UBF. RB and p53 also bind and repress the TFIIIB transcription factor, thereby negatively regulating Pol III activity. Therefore, mutations that inactivate the tumoursuppressor activity of RB or p53, or both, result in aberrant upregulation in essential components of the protein synthesis machinery and increase ribosome biogenesis, leading to enhanced mRNA translation rates. UBF activity is also tightly regulated at the level of its phosphorylation (P). The carboxyterminal acidic region of UBF is phosphorylated by cyclindependent kinase 2/4 (CDK2/CDK4) and casein kinase II (CKII), and this has been correlated with enhanced binding of UBF to SL1. A rapid activation of endogenous rRNA by epidermal-growth-factor signalling is mediated by direct phosphorylation of UBF by the mitogen-activated protein (MAP)-kinase ERK (extracellular signal-regulated kinase). Given that the CDK and cyclin complex that phosphorylates UBF, as well as ERK, are overexpressed in human neoplasias, an increase in UBF modification could increase rRNA transcription and promote transformation. Phosphorylation of the initiation factor TIF-IA after stimulation by various extracellular stimuli correlates with an increase in growth-dependent regulation of Pol I activity, and the kinases that initiate this phosphorylation are likely to regulate rRNA synthesis on stimulation with mitogenic stimuli. CRYO-ELECTRON MICROSCOPY A technique that is used to study the three-dimensional structures of biomolecules. A sample is rapidly frozen in liquid helium to preserve its structure and then examined in the frozen, hydrated state in a cryoelectron microscope. NATURE REVIEWS | C ANCER Although compelling biochemical evidence indicates that many proto-oncogenes and tumour suppressors interact directly with components of the Pol I and Pol III transcription complex, the functional consequences of increased rRNA synthesis on translation or in the increased production of functional ribosomes is lacking. Are translation rates upregulated in all cancer cells? Does specificity exist in the type of mRNAs that show a potential increase in translation following an increase in Pol I and/or Pol III activity? Furthermore, given that many of the tumour suppressors and protooncogenes that regulate rRNA synthesis have additional cellular functions, further research is required to clarify the consequences of increased Pol I and Pol III activity in cancer cells. Some indication of the effects of increasing ribosome synthesis comes from recent characterization of the Drosophila tumour suppressor mutant brain tumor (brat). Homozygous brat mutant flies die of an enlarged brain that is up to eight times bigger than the normal size, and these tumours consist of malignant optic neuroblasts that have the potential to metastasize44. The brat gene encodes a protein that regulates rRNA synthesis in the fly. The Caenorhabditis elegans homologue of brat, ncl-1, has been shown to directly negatively regulate both Pol I and Pol III transcription45, and brat is able to rescue the phenotype of ncl-1 mutants. This indicates that the function of this protein is conserved during evolution. The brat-mutant tumour phenotype is associated with an increase in rRNA synthesis, although little is known about the precise mechanism by which Brat regulates Pol I transcription. The brat-mutant cells are larger than wild-type cells and they have enlarged nucleoli, which are associated with an increase in total rRNA production. These results provide evidence that the tumour phenotype of brat mutants could be caused by excessive cell growth and ribosome synthesis. The association of cell growth and increased ribosome function is commonly associated with overproduction of the translation apparatus (see below). It remains to be determined whether this is merely a correlation, as cells that produce more ribosomes are larger to enable them to accommodate their increased protein load, or whether increased cell size actually promotes tumour development. Ribosomal proteins and tumorigenesis More than 70 ribosomal proteins (r-proteins) must assemble with the rRNAs in the nucleolus to form the small and the large ribosomal subunits 40S and 60S46 (BOX 1). Several ribosomal proteins possess conserved domains that are important for their interaction with rRNA47. In the past, it was assumed that r-proteins stabilize specific rRNA structures in mature ribosomal subunits and promote correct folding of rRNAs during ribosome assembly. The recent solution of the ribosome structure by X-ray crystallography and CRYO-ELECTRON MICROSCOPY has improved our understanding of the specific roles of different regions of rRNA, as well as the function of r-proteins during the steps that lead to protein synthesis48,49. These data revealed that many r-proteins might function as RNA chaperones not only during the assembly of the ribosome particles, but also in the stabilization of important domains of the rRNA, such as the peptidyl transferase center of the large subunit. In addition, r-proteins could coordinate the interaction between the ribosome and mRNA, as well as with initiation and elongation factors. Why should the activity of r-proteins interest the cancer biologist? Alterations in the expression of r-proteins seem to be consistently associated with tumorigenesis. In fact, many r-proteins that belong to the small (S) and the large (L) ribosomal subunit are overexpressed in cancer cell lines, as well as in primary tumours50–52. R-proteins are markers for general cellular transformation, as single and multiple groups of proteins are overexpressed by solid tumours VOLUME 3 | MARCH 2003 | 1 8 3 REVIEWS Growth factors, mitogen stimuli PI3K PTEN ? PDK1 X AKT 5′ TOP mRNA translation P S6 Elongation factors Ribosomal proteins Ribosome biogenesis factors mTOR S6K Rapamycin ? TSC1 TSC2 Figure 2 | Regulation of protein synthesis by the PI3K signal-transduction pathway. The activation of the phosphotidylinositol 3-kinase (PI3K) signal-transduction pathway by growth factors and mitogenic stimuli results in increased translation of a subset of mRNAs that contain a terminal oligopyrimidine tract (TOP) in their 5′ untranslated region (UTR). The 5′ TOP genes encode many components of the translation apparatus, including ribosomal proteins and elongation factors. Three different signal-transduction pathways downstream of PI3K activation have been proposed to account for regulation of 5′ TOP gene expression following mitogenic stimuli. In this first pathway, direct activation of the kinase mTOR (mammalian target of rapamycin) by AKT results in increased S6 kinase (S6K) activity and, subsequently, the phosphorylation of the ribosomal protein S6. Activation of S6 by phosphorylation leads to an upregulation in the translation of selective messengers, 5′ TOP genes, through an unknown mechanism. It has also been shown that a second signaltransduction pathway exists whereby PDK1 can directly phosphorylate S6K, leading to activation of S6. Deregulation of S6 phosphorylation could therefore contribute to tumorigenesis elicited by activation of the PI3K signal-transduction pathway in cancer. The tumour suppressor tuberous sclerosis complex-2 (TSC2), which exists as a heterodimeric complex with TSC1, acts as an antagonist of S6K activation. It has been proposed that TSC1/2 can either inhibit mTOR activity by preventing its phosphorylation or inhibit S6K phosphorylation through a poorly defined pathway. Furthermore, AKT can also inhibit the formation of the TSC1/2 complex by phosphorylating TSC2 and thereby disrupting the activity of this S6K antagonist on stimulation with mitogenic stimuli107,135,136. Cancer cells that lose TSC1/2 function therefore might also show increased 5′ TOP mRNA production, leading to increased uncontrolled protein synthesis. It has recently been shown that a third signaltransduction pathway could also exist, whereby the regulation of 5′ TOP mRNA translation would be dependent on the PI3K signal-transduction pathway, but independent of both S6K and phosphorylation of S6. This pathway involves unidentified protein components (X). Given that most 5′ TOP genes encode proteins that can upregulate protein synthesis, a feedback loop might exist by which an increase in S6 phosphorylation results in an upregulation in total protein synthesis in the cell. Rapamycin, a specific inhibitor of the mTOR kinase, is currently in clinical trials for its growth/tumour-suppressive activity. and by leukaemia cells53–57. How could alterations in r-protein expression lead to a defect in ribosome function, and, more importantly, to alterations in cellular proliferation? The synthesis of r-protein genes is a coordinated process, leading to the precise equimolar production of 78 proteins58. Investigations into the consequences of deletions in specific r-proteins have been conducted mainly on lower eukaryotic organisms such as yeast. Gene-targeting experiments in yeast have shown that the depletion of a single r-protein such as L16, results in a decrease of the 60S ribosomal 184 | MARCH 2003 | VOLUME 3 subunit, and that this is associated with a reduction in polyribosomes and a defect in cellular growth 59. Disrupting expression of only one r-protein can therefore shut down ribosome production. The control of ribosome biogenesis by single r-proteins has been validated for other r-proteins in addition to L16, and it has also been shown in other model organisms. Mutations in individual r-proteins in Drosophila lead to a class of mutants collectively known as minute. Minute flies are characterized by reductions in body size, diminished fertility and recessive lethality 60. Direct and indirect evidence support the fact that minute cells contain fewer ribosomes and so have a reduced capacity for protein synthesis61. Reductions in protein synthesis could reduce cell growth and proliferation. Verification of this hypothesis in higher organisms has been partially validated by experiments that involve the conditional deletion of the 40S r-protein, S6, in mice. This results in defective ribosome biogenesis and reduced cell proliferation62. The ribosomal protein S6 is of particular interest, due to the fact that its phosphorylation is tightly regulated by extracellular signalling pathways that are deregulated in cancer cells (see below). S6 phosphorylation has been associated with translation of a specific class of mRNAs termed TOP (a terminal oligopyrimidine tract in the 5′ untranslated region (UTR)) mRNA63–65. This class of mRNAs includes ribosomal proteins, elongation factors 1A1 and 2 (EEF1A1 and EEF2) and several other proteins that are involved either in ribosome biogenesis or in translation control (FIG. 2). TOP mRNAs are regulated at the translation level as they are shifted from polysomes (active translational ribosomes) in growing cells into mRNP (inactive translational particles) in quiescent cells66. The proteins encoded by TOP genes might themselves act as protooncogenes. For example, EEF2 — an isoform of EEFIA — is amplified in primary ovarian tumours, and its overexpression is oncogenic in fibroblast cell lines and in xenograft tumour models67. Therefore, deregulation in TOP gene expression might promote tumorigenesis. Furthermore, given that most known TOP genes encode proteins that can upregulate protein synthesis, a feedback loop might exist in which an increase in S6 phosphorylation results in upregulation in total protein synthesis in the cell (FIG. 2). S6 is phosphorylated when quiescent cells reenter the cell cycle in response to mitogens 68. Phosphorylation of S6 has been observed both in vivo, during liver regeneration, and in various cell lines 69,70. Because the level of S6 phosphorylation increases in proportion to the level of protein synthesis, it has been proposed that S6 could be an important regulator of cell growth, through the regulation of translation of TOP mRNAs. This has been proven, in part, in Drosophila, where the inactivation of the kinase that is responsible for the S6 phosphorylation (dS6K) results in a decrease in body size and a defect in cell growth71. There is no data to indicate that this is due to alterations in translation of TOP genes. www.nature.com/reviews/cancer REVIEWS RAPAMYCIN Rapamycin is a macrolide with immunosuppressant properties and tumour-inhibitory effects. To exert its effect, rapamycin binds an immunophilin FK506-binding protein 12, and this complex inhibits the cellular target of rapamycin, mTOR. NATURE REVIEWS | C ANCER In mammals, the regulation of S6 phosphorylation is more complex. This increased complexity is due to the presence of another kinase, S6K2, which is highly homologous to S6K1 and encodes a protein with redundant function72. Despite the presence of this homologue, S6 is not phosphorylated in S6K1−/− embryonic stem cells73. Furthermore, serum stimulation of starved wildtype cells causes increased amounts of the mRNA for the TOP gene EEF1A1 to associate with the polysome fraction, whereas this did not occur in S6K1−/− cells. Overexpression of a dominant-negative form of S6K1 also suppressed the mitogen-induced translation of chimeric transcripts that contained the 5′ TOP sequence at the 5′ UTR65. In addition, RAPAMYCIN, a macrolide that inhibits cellular proliferation by inhibiting the mammalian target of rapamycin (mTOR), reduces S6K1 activity and inhibits the upregulation of TOP mRNAs after mitogen stimulation65,73 (FIG. 2). A recent study has challenged these findings, and has proposed that TOP gene expression is dependent on the phosphotidyl inositol 3-kinase (PI3K) signalling pathway, but is independent of S6K1 activity74. Further experiments need to be performed to reconcile these data — for example, by studying cells or mice that carry gene disruptions in both S6K1 and S6K2 kinases. Importantly, the activity of S6K is markedly upregulated in tumours that carry mutations in PTEN and tuberous sclerosis 1/2 (TSC1/2), and therapeutic agents that disrupt S6K activity were found to slow tumour growth in mice and in humans (FIG. 2). The upregulation of r-proteins in cancer cells fits nicely with their involvement in ribosome production. Similar to the increase in Pol I transcriptional activity that results in an increase in rRNA synthesis, r-proteins could also regulate the number of functional ribosomes in the cell. In both cases, cells that contain more ribosomes would have an increased translation rate, which would promote transformation. Although many studies have shown that several r-proteins are overexpressed by cancer cells, no experiments have been performed to determine the consequences of this overexpression on ribosome production and translation. It is clear that deletion of a single r-protein has a direct effect on cellular growth in many model organisms, but the consequences of r-protein overexpression are not as well characterized. In one study, overexpression of the r-protein S3a was able to induce transformation of NIH 3T3 cells and induce formation of tumours in nude mice75. The ability of S3a to induce transformation is dependent on its role in suppressing programmed cell death, and therefore S3a might cause an upregulation of anti-apoptotic proteins. However, in these studies, the effects of S3a on protein translation have not been investigated. Furthermore, it is difficult to envision how S3a could specifically induce translation of anti-apoptotic proteins, so extra-ribosomal functions have been attributed to S3a by default. The advent of proteomics will help to identify proteins that are affected by r-protein overexpression. It is possible that r-protein overexpression is simply the result of rapid cellular proliferation. Cancer cells, which show an increase in cell-cycle progression, might require more ribosomal components to sustain increased proliferation. In this case, upregulation of r-proteins could be an indirect consequence of transformation that simply maintains the increased proliferation of cancer cells. However, this has not been proven and it is an unlikely scenario, given that no data exist to correlate tumorigenic cells that possess high proliferative rates with overexpression of specific r-proteins. Certain r-proteins could also have additional functions. For example, it has been shown that the r-protein S3 also functions as a DNA-repair enzyme — ultra-violet endonuclease III — that cleaves DNA in response to ultraviolet irradiation76. Although the extra-ribosomal function of these proteins might be important for some aspects of cancer development, the fact that there are no specific r-proteins associated with specific tumour types makes this hypothesis unlikely. This issue becomes particularly difficult to address when several different combinations of r-proteins are deregulated in cancer cells. Ultimately, given the overwhelming number and combination of r-proteins that are overexpressed in a wide array of cancer types, it is highly unlikely that all of their effects will be attributed to extra-ribosomal function. MYC and PTEN Both the proto-oncogene product MYC and the tumour suppressor PTEN have been shown to directly regulate ribosome biogenesis through the transcription of r-proteins and the regulation of S6K activity, respectively (FIGS 2,3). c-MYC encodes a transcription factor that is deregulated through genomic aberrations in a wide array of B-cell-specific malignancies and several other types of neoplasias77. How is MYC function linked to ribosome biogenesis? This question has been answered by the finding that r-proteins, as well as factors involved in ribosome assembly, are MYC gene targets78–80 (FIG. 3). Transgenic mice that constitutively expressed c-MYC under the control of the immunoglobulin heavy-chain enhancer show increases in cell size. This effect on cell size is prevalent during all stages of B-cell differentiation and is independent of the cell cycle, as cells in G0/G1 or G2 are bigger in size compared with the control cells81. As discussed above, overexpression of a single r-protein has not been conclusively shown to affect protein synthesis. However, MYC-overexpressing cells upregulate r-proteins, resulting in an increase in cell size and in S35 incorporation. MYC can therefore regulate total protein synthesis in the cell. Overexpression of human c-MYC in the liver resulted in hepatocyte enlargement82. This increase in size occurred in the absence of cell proliferation and was associated with enlarged nucleoli. Expression of several r-proteins and nucleolar proteins that are involved in ribosome biogenesis were also increased, with respect to control cells. These findings are in agreement with results from SERIAL ANALYSIS OF GENE EXPRESSION (SAGE) studies in neuroblastoma cell lines. In these studies, these cells were transfected with another member of the MYC oncogene family, NMYC 79. Most genes upregulated in these cells VOLUME 3 | MARCH 2003 | 1 8 5 REVIEWS SERIAL ANALYSIS OF GENE EXPRESSION (SAGE). A technique that is used to identify and quantitate genes that are differentially expressed between two different samples. SAGE libraries are generated by adding a short nucleotide sequence tag (10 base pairs) to cDNAs from different RNA samples that are being analysed. The abundance of a given tag within the SAGE library corresponds to the expression level of the corresponding gene, which can easily be identified by sequencing directly from the transcript tag. were r-proteins associated with the large and small ribosomal subunits (FIG. 3). In addition, analysis of primary neuroblastoma samples with an amplification of endogenous NMYC revealed an overexpression of genes that encode components of the protein synthesis machinery79. A second group of genes upregulated in NMYCtransfected cells consist of nucleolar proteins such as B23 (nucleophosmin or NPM) and nucleolin78–80,82. These two gene products were also found to be upregulated in c-MYC-overexpressing hepatocytes. These nucleolar proteins function in the processing of rRNA precursors, and have been implicated in the regulation of ribosome assembly or the nucleo-cytoplasmatic transport of mature ribosomal subunits. Nine translation initiation and elongation factors were also induced in these cells. These results support a role for MYC-family members as key regulators of ribosome biogenesis and translation control. The ability of MYC to regulate the transcription of several components of the protein synthesis machinery has been validated in several different cell types and corresponds with both MYC loss and gain of function. Furthermore, the ability of MYC to regulate the expression of this class of genes is dependent on its ability to regulate transcription. This has been shown in cells that overexpress MAD1, a transcriptional repressor of MYC target genes. Cell growth is inhibited in these cells, and genes that are involved in ribosome biogenesis and translation control are repressed83. It remains to be determined which of these genes have MYC DNA-binding sites within their promoters and which, by contrast, are in pathways indirectly controlled by MYC. MYC Ribosomal proteins: L3, L15, L39, L37, S2, S3a, S6, etc Nucleolar proteins: B23, nucleolin, fibrillarin, etc Translation factors: elF-4A, eIF-4E, elF3, EEF1α1, etc Ribosome biogenesis and translation control Figure 3 | MYC regulates the expression of proteins involved in ribosome biogenesis and control of translation. The proto-oncogene MYC encodes a transcription factor that is deregulated through genomic aberrations in a wide array of B-cell-specific malignancies and several other types of neoplasias. Most genes that are found to be upregulated in MYC-overexpressing cells are ribosomal proteins associated with the large and small ribosomal subunits. In addition, several nucleolar proteins are found upregulated, including B23, which is overexpressed in a wide array of tumours and might have direct consequences for tumour progression that is initiated by MYC. The translation factor eIF-4E is also itself a proto-oncogene and is one of many translation factors found upregulated in MYCoverexpressing cells. The ability of MYC to regulate the expression of these classes of mRNAs corresponds to an increase in protein synthesis observed in MYC-overexpressing cells. Regulation of protein synthesis could therefore be an important mechanism by which MYC regulates cell growth and initiates tumorigenesis. 186 | MARCH 2003 | VOLUME 3 It will be essential to determine which of the MYC target genes involved in regulation of protein synthesis also promote transformation, and whether an increase in ribosome biogenesis and translation of specific mRNAs directly mediates MYC’s oncogenic effects. Overexpression of MYC has been shown to increase cell size — possibly to accommodate the expanded proteinsynthesis capacity of these cells81–84. Whether the expansion in cell size per se has an affect on the susceptibility of these cells to undergo transformation remains unknown. Although beyond the scope of this review, much research is currently focused on whether the ability of proto-oncogenes, such as MYC, to regulate cell size would make these cells more susceptible to acquire secondary mutations involved in cancer progression (see REFS 85,86 for reviews). The tumour suppressor PTEN87,88 has also been shown to regulate cell size in association with its ability to regulate ribosome biogenesis89,90. PTEN is a phosphatase that downregulates the PI3K pathway by dephosphorylating the lipid phosphatidylinositol-3,4, 5-trisphosphate to phosphatidylinositol-4,5-bisphosphate (REFS 91,92). PI3K signalling leads to an increase in S6K activity, concomitant with hyperphosphorylation of the S6 ribosomal protein86 (FIG. 2). These events are positively regulated by the kinase mTOR (a target of rapamycin), although it is unclear whether mTOR directly phosphorylates S6K in physiological conditions93–96. PTEN acts upstream mTOR and has a negative effect on the phosphorylation of S6 (REFS 97,98). In fact, PTEN-null embryonic stem cells show an increase in S6K activity and display constitutive phosphorylation of S6. A linear model has been proposed for S6 activation, whereby AKT signals to S6K through the activation of mTOR, and PTEN negatively regulates this process99,100. Additionally, it was shown that the plekstrin homology (PH)-domain containing protein, phosphoinositidedependent kinase (PDK1), which phosphorylates AKT at the cell membrane, also directly phosphorylates S6K in an AKT-independent manner101 (FIG. 2). PTEN is mutated in many different types of tumour, but how important is its ability to regulate mTOR activity in coordinating tumour suppression? Recent reports have highlighted the importance of mTOR in PI3Kdependent oncogenesis. Transformation by PI3K or AKT directly correlates with mTOR activation and activation of its downstream target S6K in primary cultures of chicken embryo fibroblasts102. Rapamycin, which inhibits mTOR, prevents induction of transformed cell foci by PI3K and AKT, but does not have an effect on cells transformed with other oncogenes such as v-Jun or v-Src102. Pten+/− mice are more prone to developing tumours of different histological origins, which are associated in some cases with PTEN loss of heterozygosity 87,88,103,104. Data from different reports indicate loss of PTEN function in tumours with increases in AKT and S6K kinase activity. Pharmacological inactivation of mTOR by rapamycin reduces tumour-cell proliferation and tumour size in Pten+/− mice98. This tumour regression is associated with the downregulation of S6K activity to wild-type levels. In tumours derived from www.nature.com/reviews/cancer REVIEWS murine Pten−/− embryonic stem cells, or in human tumour cell lines in which PTEN is lost, rapamycin treatment led to a reduction in cell size97. The drug has no effect on tumour cells without PTEN mutations. This supports the role of PTEN in growth regulation through S6K activation. The mTOR pathway and the downstream effector S6K seem to be involved in tumorigenesis, and rapamycin and its analogues might therefore be developed as chemotherapeutic agents. Another tumour suppressor that was recently shown to regulate S6K activity is the heterodimeric complex TSC1/2, which is mutated in patients with tuberous sclerosis105. This disease is associated with cancer susceptibility, including haemartoma growth. The TSC1/2 complex in Drosophila, as well as in mammalian cells, acts as an inhibitor of S6K activity106–110 (FIG. 2). TSC2 mutations are also associated with tumour development in lymphangioleiomyomatosis (LAM) in primary tumours, as well as in cell lines derived from these tumours, which display constitutive activation of S6K. The hyperphosphorylation of S6 is associated with an increase in cell proliferation111. So, does the TSC1/2 complex directly inactivate S6K activity, or does regulation of S6 phosphorylation by TSC1/2 depend on other components of PI3K pathway, such as mTOR? Although there is more data to support the model that the TSC1/2 complex inhibits mTOR106–109, and this in turn results in inactivation of S6K activity, other data place mTOR in a parallel pathway112. If the former model is correct, then PTEN and TSC1/2 could cooperate to repress cellular growth and proliferation, and concomitant deregulation of both of these tumour suppressors might have an additive function during tumorigenesis. Regardless of the precise mechanisms by which S6K activation is perturbed in tumours, the fact remains that mutations in PTEN and TSC1/2, as well as amplification of the AKT gene, all have a common downstream r-protein target, which is S6. Unfortunately, although emphasis has been placed on how TSC1/2 and PTEN regulate SK6, the downstream effects of S6K hyperactivation in cancer remain less characterized. The fact that S6 is phosphorylated makes it a highly regulated r-protein, and is constitutively activated in several tumours. Whether this results in upregulation of TOP gene expression or an increase in total protein synthesis needs to be determined. It will also be important to establish whether rapamycin treatment also has an effect on the initiation factor of translation, eIF4E, whose activity is regulated by mTOR99. Because eIF4E has been shown to act as a proto-oncogene113–115, it might also be involved in deregulated protein synthesis that is elicited by the loss of PTEN and TSC1/2 tumour suppressors. Further experimental evidence is required to link the activity of ribosomal proteins such as S6 to translation defects and cancer. Crosses between Pten-mutant mice and mice deficient in components of the mTOR signal-transduction pathway will clarify to which extent activation of mTOR is a prerequisite for neoplastic transformation. In addition, it will be worthwhile to NATURE REVIEWS | C ANCER investigate in PTEN-null cells whether protein translation is impaired as a result of constitutive activation of the mTOR pathway. Given that S6K is involved in the control of TOP gene translation, it will be particularly informative to study the expression of this subset of proteins in Pten-mutant mice and cells. Cancer and ribosome biogenesis Oncogenes and tumour suppressors can contribute to cancer progression through alterations in ribosome biogenesis. Genetics studies have shown that components of the protein synthesis machinery can be directly mutated and are responsible for disease syndromes that are characterized by an increased susceptibility to cancer. Could disruption of ribosome function be the first oncogenic event in a multi-hit tumour-formation process? Dyskeratosis congenita. DKC1 encodes a putative pseudouridine synthase, which mediates post-transcriptional modification of ribosomal RNA (rRNA) through the site-specific conversion of uridine (U) to pseudouridine (ψ)116,117 (BOX 1). Mutations in DKC1 are associated with dyskeratosis congenita (DC), a disease that is characterized by features of premature ageing, including bone-marrow failure and hyperkeratosis of the skin2,118. Importantly, patients with DC show an increased susceptibility to tumour formation. In yeast and fly, mutations in DKC1 result in impaired rRNA modifications associated with a reduction in ribosome biogenesis119,120. It is therefore tempting to speculate that the molecular pathogenesis of DC and the tumour susceptibility observed in DC patients result from defects in ribosome modification. However, the possibility of a direct involvement in ribosome biogenesis as a cause of the disease has remained controversial, given that DKC1 is also associated with the RNA component of the telomerase complex121. Whether ribosome dysfunction is the underlying cause of DC pathogenesis has been tested in animal models3. The mouse represents an ideal model organism for addressing whether DKC1 mutations result in defects in rRNA modification, telomere maintenance, or both, as defects in telomerase function leading to telomere attrition would only be manifest in late generations122. Hypomorphic Dkc1 mutant (Dkc1m) mice3 recapitulate, in first generations, the clinical features of DC. Dkc1m cells from early-generation mice are impaired in rRNA pseudouridylation before disease onset. These results indicate that deregulated ribosome function is involved in the initiation of DC. More than 50% of Dkc1-mutant mice develop tumours during their lifespan, indicating that Dkc1 is an important tumoursuppressor gene. Dkc1m mice also show defects in rRNA processing (BOX 1), and cells from these mice are hypersensitive to translation inhibitors, which are used to quantify ribosome function. How can alterations in rRNA modification promote neoplastic transformation? DKC1 has been shown to mediate pseudouridylation. Although the specific function of the pseudouridine rRNA modification VOLUME 3 | MARCH 2003 | 1 8 7 REVIEWS remains to be determined, it has been shown that modification sites are strategically placed within specific domains of the ribosome123. For example, modifications have been found in regions of the ribosome that are important for tRNA and mRNA binding. The reduction in modified uridine residues in the ribosome could result in impaired translation of specific mRNAs that are important for cellular transformation. For example, genes encoding tumour suppressors such as p27 and p58 have been shown to contain 5′ leader sequences known as internal ribosome entry sites (IRES), which facilitate interaction of the ribosome with mRNA to promote translation124,125. Translation of these types of mRNAs requires more direct contacts with the ribosome and would therefore be more sensitive to alterations in ribosome folding. Given that co-crystallization of Escherichia coli DKC1 with RNA revealed a potential role for pseudouridine synthases as RNA chaperones that are important for proper ribosome folding126, DKC1-mutant cells might possess misfolded ribosomes. These mutant ribosomes could be impaired in the translation of mRNAs with IRES sequences124,125,127. The identification of genes that are deregulated in Dkc-mutant mice will improve our understanding of how pseudouridylation might affect ribosome function and contribute to cancer. Diamond–Blackfan anaemia. Another cancer susceptibility syndrome that is directly associated with defects in ribosome biogenesis is Diamond–Blackfan anaemia, which is characterized by anaemia and an increased susceptibility to haematopoietic malignancies128. This disease has been associated with mutations in the r-protein S19, providing direct evidence that loss of r-protein function can cause cancer4. Although the molecular mechanisms by which a reduction in the expression or activity of S19 causes the cancer predisposition are unknown, several features of the disease state indicate that patients are impaired in ribosome function. As in Drosophila, in which mutations in a single r-protein result in the minute phenotype, patients with Diamond–Blackfan anaemia have hypotrophy at birth and show severe growth retardations128. As r-proteins are highly conserved between Drosophila and humans, is it likely that defects in human r-proteins would result in overall ribosomal dysfunction, and that this could lead to growth defects. However, as stated above, the possibility for an extraribosomal function for S19 cannot be excluded. Because r-protein expression is upregulated in several types of cancer, it remains to be determined how the loss of an r-protein would contribute to neoplasm. Given the specificity in the pathological features that are observed in Diamond–Blackfan anaemia patients, it has yet to be seen whether S19 can regulate translation in a qualitative manner, as proposed for the S6 r-protein, which facilitates translation of 5′ TOP genes. Therefore, downregulation in translation of specific mRNA, such as those encoding tumour suppressors, could be a possible mechanism for the loss of a r-protein. Creation of animal models 188 | MARCH 2003 | VOLUME 3 for Diamond–Blackfan anaemia and primary cells that lack S19 will be useful in determining the mechanism by which ribosome biogenesis promotes cancer. Future directions Several pathways that lead to increased ribosome biogenesis have been associated with the transformation process (BOX 2). Alterations can occur either at the level of rRNA transcription, modification or assembly of the ribosome. Evidence has clearly shown that several tumour suppressors and oncogenes regulate ribosome function, but determining the precise effects of these interactions will require further investigation. We have loosely used the term ‘increased total translation’ throughout the review as an obvious potential outcome for increased ribosome production. Although this might occur in tumours that show increases in rRNA synthesis, how can increased translation predispose cells to neoplasm? At the heart of this question is the debate about whether increased cellular growth or proliferation brought about by elevated proteins synthesis is a means to cancer progression, or whether it actually serves as a bona fide ‘hit’ which itself is necessary and sufficient for cancer initiation. Several possibilities exist, each of which remain highly speculative (BOX 2). One consequence of an increase in translation of cellular mRNAs would be the increased production of certain proteins that normally show low or moderate levels of expression. Expression of most oncogenes is tightly regulated to prevent deleterious effects on cell growth. One mechanism by which cells regulate oncogene expression involves the long, highly structured 5′ UTR that is present in these mRNAs129. Initiation factors are believed to unwind the secondary structure of 5′ UTRs and recruit ribosomes to these sites. This process limits the expression rate of these proteins. It has been proposed that initiation factors such as eIF4E could be rate limiting in the cell, and that their upregulation might result in increased cancer susceptibility through enhanced translation of mRNAs with a highly structured 5′ UTR114,115. An upregulation in ribosome production could facilitate the translation of these mRNAs by increasing the rate of formation of the initiation complex. It is worth noting that the overexpression of proto-oncogenes such as MYC results not only in an upregulation in ribosomal proteins but also in initiation factors for translation (FIG. 3). Therefore, an increase in translation, mediated by a rise in ribosome production and initiation factors, could actually promote translation of proteins involved in cellular growth control that are the least abundantly expressed in the cell and often encode proto-oncogenes. Specificity in translation of certain mRNAs could also be achieved by an upregulation of certain r-proteins. As discussed above, phosphorylation of S6 results in an increase of specific target mRNAs that contain a terminal oligopyrimidine tract in their 5′ UTR. S6 is constitutively phosphorylated in many types of cancer cell, and the activity of tumour suppressors that is important for the phosphorylation of this ribosomal protein is also deregulated. However, www.nature.com/reviews/cancer REVIEWS Box 2 | Different steps in the pathway to ribosome production that can lead to cancer, when deregulated The regulation of ribosome biogenesis is a highly coordinated process that leads to accurate initiation and regulation of protein synthesis. There are three steps in this process, which, when deregulated, can contribute to increased tumorigenesis. The first step in ribosome production requires the synthesis of the 45S rRNA precursor. The transcription of this rRNA gene is negatively regulated by tumour suppressors such as p53 and retinoblastoma (RB) and augmented on mitogenic stimuli by several kinases that phosphorylate components of the transcription complex that are responsible for 45S synthesis. The accurate regulation of rRNA synthesis can be lost in cancer cells through inactivating mutations in tumour suppressors or upregulation of these kinases. Another step in ribosome biogenesis that maintains accurate cellular function involves the modification of rRNA. One type of modification, which is catalysed by an enzyme known as dyskerin, converts uridine into pseudouridine (Ψ). Mutations in the gene encoding dyskerin, DKC1, result in increased tumour susceptibility. Animal models that have lost DKC1 function show a marked increase in tumour incidence associated with a decrease in rRNA processing. Ribosome assembly, which involves the association of rRNA with more than 70 ribosomal proteins (made in the cytoplasm), is another important step. The rRNA and proteins are assembled into the large subunit (60S) and small subunit (40S) of the ribosome. An increase in ribosomal protein production and activity has been observed in many cancer types. Mutations in ribosomal proteins such as S19 have also been associated with a human syndrome that is characterized by increased tumour susceptibility. Each of the three highlighted steps (blue boxes) might have deleterious effects on the cell that could contribute to tumour initiation or cancer progression, or both, through aberrant regulation of protein synthesis. This can be manifested either by an increase in ribosome production, thereby leading to an upregulation in total translation, or by alterations in translation of specific mRNAs, which are involved in the regulation of cell proliferation. Furthermore, when key checkpoints in the cell that are important in coordinating ribosome production with accurate cell-cycle progression are lost,‘nucleolar stress’ can result, and subsequently unrestrained cellular proliferation occurs. The advent of proteomics will aid in identifying protein targets that are deregulated as a result of perturbations in these pathways. Increase in rRNA synthesis by inactivating mutations in tumour suppressors rRNA gene 45S rRNA precursor transcription Nucleus ψ 5′ Modification ψ ψ 3′ CH3 CH3 Nucleolus Processing and modifications by snoRNPs and RNase CH3 ψ ψ CH3 28S CH3 Ribosome assembly ψ 18S 5,8S 5S CH3 Ribosomal proteins made in cytoplasm Large subunit (60 S) Small subunit (40 S) Cytoplasm Mutations and overexpression of r-proteins it remains unclear whether the modification of S6 contributes at the level of translation of its target mRNAs towards tumorigenesis. Modification of other r-proteins following mitogen activation could similarly regulate the translation of specific mRNAs. For example, r-proteins were recently shown to be ubiquitylated, and this modification corresponds with their selective destruction during a specific window of the cell cycle 130. These and other types of modifications could provide the ribosome with specificity for certain messengers, or enable cell-cycle control of ribosome function to take place. The precise consequence of r-protein overexpression, and whether these mechanisms promote cellular transformation, remains to be determined. NATURE REVIEWS | C ANCER Alterations in the modification of rRNA (DKC1) • Increased total translation • Altered translation of specific mRNAs • Nucleolar stress and deregulated cell-cycle progression Although ribosome biogenesis is tightly linked to regulation of translation, and cells that overexpress MYC show an increase in total translation, other possibilities exist for how alterations in ribosome biogenesis could lead to cancer. This is highlighted by findings during the course of analysing the S6−/− mice62. Although loss of this r-protein results in decreased ribosome biogenesis, total cellular translation is relatively unaffected. However, following challenge, the S6-null cells show a defect in proliferation. This defect occurred concomitantly with a decrease in cyclin E at the mRNA level, which is important for the G1-to-S transition. How could alterations in ribosome biogenesis per se affect the cell cycle? Growth and proliferation are associated with changes in the rate of ribosome production, and ribosome VOLUME 3 | MARCH 2003 | 1 8 9 REVIEWS biogenesis can serve as a sensor for cells to bypass important checkpoints during the cell cycle. In transformed cells that show increased ribosome production, the cell cycle can be altered as a consequence of alterations in ribosome biogenesis. This tight feedback between the ribosome and the cell cycle could function to maintain cellular homeostasis. Therefore, an increase in ribosome amount, irrespective of any consequent affects on protein translation, could contribute to the transformation process. Unfortunately, the proteins that are responsible for mediating cross-talk between the ribosome and the cell cycle remain largely unknown. For example, the mechanism by which S6-null cells, which contain fewer ribosomes than cells that express S6, show a selective downregulation of cyclin E mRNA at the transcript level is unknown. Interestingly, a novel nucleolar protein involved in rRNA processing known as BOP1 (REF. 131) has been shown to cooperate with p53 in regulating the G1-to-S transition132. The ability of a dominant-negative form of Bop1 to induce cellcycle arrest was abrogated in p53-null cells, but rRNA processing was still impaired. These findings indicate that the p53 pathway could link ribosome biogenesis and the cell cycle. Alterations in ribosome biogenesis in 1. Gani, R. The nucleoli of cultured human lymphocytes. I. Nucleolar morphology in relation to transformation and the DNA cycle. Exp. Cell Res. 97, 249–258 (1976). 2. Heiss, N. S. et al. X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nature Genet. 19, 32–38 (1998). 3. Ruggero, D. et al. Dyskeratosis congenita and cancer in mice deficient in ribosomal RNA modification. Science 299, 259–262 (2003). In this study, animal models for dyskeratosis congenita were generated, which are hypomorphic mutants for the pseudouridine synthase, DKC1. These mice are faithful models of the disease and show marked cancer susceptibility associated with an impairment in ribosome biogenesis. 4. Draptchinskaia, N. et al. The gene encoding ribosomal protein S19 is mutated in Diamond-Blackfan anaemia. Nature Genet. 21, 169–175 (1999). Mutations in the ribosomal protein S19 were found to be associated with a disease characterized by an increased susceptibility to cancer. 5. Pardee, A. B. G1 events and regulation of cell proliferation. Science 246, 603–608 (1989). 6. Pyronnet, S. & Sonenberg, N. Cell-cycle-dependent translational control. Curr. Opin. Genet. Dev. 11, 13–18 (2001). 7. Grummt, I. Regulation of mammalian ribosomal gene transcription by RNA polymerase I. Prog. Nucleic Acid Res. Mol. Biol. 62, 109–154 (1999). 8. Grummt, I., Smith, V. A. & Grummt, F. Amino acid starvation affects the initiation frequency of nucleolar RNA polymerase. Cell 7, 439–445 (1976). 9. Kief, D. R. & Warner, J. R. Coordinate control of syntheses of ribosomal ribonucleic acid and ribosomal proteins during nutritional shift-up in Saccharomyces cerevisiae. Mol. Cell. Biol. 1, 1007–1015 (1981). 10. Klein, J. & Grummt, I. Cell cycle-dependent regulation of RNA polymerase I transcription: the nucleolar transcription factor UBF is inactive in mitosis and early G1. Proc. Natl Acad. Sci. USA 96, 6096–6101 (1999). An important paper showing that UBF activity is regulated in a cell-cycle-dependent manner and that fluctuations in rRNA synthesis during the cell cycle are controlled by UBF phosphorylation. 11. Cavanaugh, A. H. et al. Activity of RNA polymerase I transcription factor UBF blocked by Rb gene product. Nature 9, 177–180 (1995). 190 | MARCH 2003 | VOLUME 3 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. cancer cells might result in a general ‘nucleolar stress’, leading to alterations in cell-cycle progression. In this model, the rate or efficiency of ribosome production in the cell could serve as a signal by which cells could negatively or positively regulate cell-cycle progression independently from its effect on protein synthesis. A significant challenge in the future will be to understand the relative contribution of proteins directly involved in ribosome biogenesis and the control of translation, as these could be important to the molecular aetiology of specific tumour types, as well as their precise effects within the context of a multistep pathway that leads to cancer. This avenue of research will be greatly benefited by the advent of proteomics, which will enable the identification of proteins directly deregulated as a result of alterations in ribosome biogenesis. The finding that components of the translation machinery are overexpressed or deregulated in cancer cells could lead to the discovery of new therapeutic agents, designated to target factors that control translation. A compelling example in this respect is represented by rapamycin, which specifically inactivates mTOR and has been recently shown to act as a powerful tumour-suppressive drug in clinical trials133. A landmark paper which shows that the tumoursuppressor gene Rb directly represses transcription of rRNA through inhibition of UBF function. There is an accumulation of Rb in the nucleolus of differentiated cells, indicating that Rb might exert some of its growth-suppressive properties by regulating rRNA synthesis. Beckmann, H., Chen, J. L., O’Brien, T. & Tjian, R. Coactivator and promoter-selective properties of RNA polymerase I TAFs. Science 270, 1506–1509 (1995). Brandenburger, Y., Jenkins, A., Autelitano, D. J. & Hannan, R. D. Increased expression of UBF is a critical determinant for rRNA synthesis and hypertrophic growth of cardiac myocytes. FASEB J. 15, 2051–2053 (2001). Bell, S. P., Learned, R. M., Jantzen, H. M. & Tjian, R. Functional cooperativity between transcription factors UBF1 and SL1 mediates human ribosomal RNA synthesis. Science 241, 1192–1197 (1988). antzen, H. M., Admon, A., Bell, S. P. & Tjian, R. Nucleolar transcription factor hUBF contains a DNA-binding motif with homology to HMG proteins. Nature 344, 830–836 (1990). Read, C. et al. High resolution studies of the Xenopus laevis ribosomal gene promoter in vivo and in vitro. J. Biol. Chem. 267, 10961–10967 (1992). Voit, R., Hoffmann, M. & Grummt, I. Phosphorylation by G1-specific cdk-cyclin complexes activates the nucleolar transcription factor UBF. EMBO J. 18, 1891–1899 (1999). Kihm, A. J., Hershey, J. C., Haystead, T. A., Madsen, C. S. & Owens, G. K. Phosphorylation of the rRNA transcription factor upstream binding factor promotes its association with TATA binding protein. Proc. Natl Acad. Sci. USA 95, 14816–14820 (1998). Tuan, J. C., Zhai, W. & Comai, L. Recruitment of TATAbinding protein-TAFI complex SL1 to the human ribosomal DNA promoter is mediated by the carboxy-terminal activation domain of upstream binding factor (UBF) and is regulated by UBF phosphorylation. Mol. Cell. Biol. 19, 2872–2879 (1999). Voit, R. & Grummt, I. Phosphorylation of UBF at serine 388 is required for interaction with RNA polymerase I and activation of rDNA transcription. Proc. Natl Acad. Sci. USA 98, 13631–13636 (2001). Stefanovsky, V. Y. et al. An immediate response of ribosomal transcription to growth factor stimulation in mammals is mediated by ERK phosphorylation of UBF. Mol. Cell 8, 1063–1073 (2001). The study shows that the activation of the ERK1/2 MAP kinases by growth factors leads to rRNA 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. transcription through a mechanism that is dependent on UBF phosphorylation. This study provides a link between growth-factor signalling and increased ribosome production. O’Mahony, D. J., Xie, W. Q., Smith, S. D., Singer, H. A. & Rothblum, L. I. Differential phosphorylation and localization of the transcription factor UBF in vivo in response to serum deprivation. In vitro dephosphorylation of UBF reduces its transactivation properties. J. Biol. Chem. 267, 35–38 (1992). Voit, R. et al.The nucleolar transcription factor mUBF is phosphorylated by casein kinase II in the C-terminal hyperacidic tail which is essential for transactivation. EMBO J. 11, 2211–2218 (1992). Voit, R., Kuhn, A., Sander, E. E. & Grummt, I. Activation of mammalian ribosomal gene transcription requires phosphorylation of the nucleolar transcription factor UBF. Nucleic Acids Res. 23, 2593–2599 (1995). Wang, T. C. et al. Mammary hyperplasia and carcinoma in MMTV-cyclin D1 transgenic mice. Nature 369, 669–671 (1994). Bortner, D. M. & Rosenberg, M. P. Induction of mammary gland hyperplasia and carcinomas in transgenic mice expressing human cyclin E. Mol. Cell. Biol. 17, 453–459 (1997). Navolanic, P. M., Steelman, L. S. & McCubrey, J. A. EGFR family signaling and its association with breast cancer development and resistance to chemotherapy. Int. J. Oncol. 22, 237–252 (2003). Cavanaugh, A. H. et al. Rrn3 phosphorylation is a regulatory checkpoint for ribosome biogenesis. J. Biol. Chem. 277, 27423–27432 (2002). Yuan, X., Zhao, J., Zentgraf, H., Hoffmann-Rohrer, U. & Grummt, I. Multiple interactions between RNA polymerase I, TIF-IA and TAFI subunits regulate preinitiation complex assembly at the ribosomal gene promoter. EMBO Rep. 3, 1082–1087 (2002). Classon, M. & Harlow, E. The retinoblastoma tumour suppressor in development and cancer. Nature Rev. Cancer 2, 910–917 (2002). Voit, R., Schafer, K. & Grummt, I. Mechanism of repression of RNA polymerase I transcription by the retinoblastoma protein. Mol. Cell. Biol. 17, 4230–4237 (1997). Hannan, K. M. et al. Rb and p130 regulate RNA polymerase I transcription: Rb disrupts the interaction between UBF and SL-1. Oncogene 19, 4988–4999 (2000). Ciarmatori, S. et al. Overlapping functions of the pRb family in the regulation of rRNA synthesis. Mol. Cell. Biol. 21, 5806–5814 (2001). www.nature.com/reviews/cancer REVIEWS 34. Hannan, K. M. et al. RNA polymerase I transcription in confluent cells: Rb downregulates rDNA transcription during confluence-induced cell cycle arrest. Oncogene 19, 3487–3497 (2000). 35. Zhai, W. & Comai, L. Repression of RNA polymerase I transcription by the tumor suppressor p53. Mol. Cell. Biol. 20, 5930–5938 (2000). Shows that the tumour suppressor p53 can directly regulate rRNA synthesis through its interaction with UBF. 36. Cordon-Cardo, C. et al. Cooperative effects of p53 and pRB alterations in primary superficial bladder tumors. Cancer Res. 57, 1217–1221 (1997). 37. White, R. J., Trouche, D., Martin, K., Jackson, S. P. & Kouzarides, T. Repression of RNA polymerase III transcription by the retinoblastoma protein. Nature 382, 88–90 (1996). 38. Sutcliffe, J. E., Brown, T. R., Allison, S. J., Scott, P. H. & White, R. J. Retinoblastoma protein disrupts interactions required for RNA polymerase III transcription. Mol. Cell. Biol. 20, 9192–9202 (2000). 39. Cairns, C. A. & White, R. J. p53 is a general repressor of RNA polymerase III transcription. EMBO J. 17, 3112–3123 (1998). 40. Kaye, F. J., Kratzke, R. A., Gerster, J. L. & Horowitz, J. M. A single amino acid substitution results in a retinoblastoma protein defective in phosphorylation and oncoprotein binding. Proc. Natl Acad. Sci. USA 87, 6922–6926 (1990). 41. Larminie, C. G., Alzuherri, H. M., Cairns, C. A., McLees, A. & White, R. J. Transcription by RNA polymerases I and III: a potential link between cell growth, protein synthesis and the retinoblastoma protein. J. Mol. Med. 76, 94–103 (1998). 42. Johnson, L. F., Williams, J. G., Abelson, H. T., Green, H. & Penman, S. Changes in RNA in relation to growth of the fibroblast. III. Posttranscriptional regulation of mRNA formation in resting and growing cells. Cell 4, 69–75 (1975). 43. Baxter, G. C. & Stanners, C. P. The effect of protein degradation on cellular growth characteristics. J. Cell. Physiol. 96, 139–145 (1978). 44. Frank, D. J., Edgar, B. A. & Roth, M. B. The Drosophila melanogaster gene brain tumor negatively regulates cell growth and ribosomal RNA synthesis. Development 129, 399–407 (2002). 45. Frank, D. J. & Roth, M. B. ncl-1 is required for the regulation of cell size and ribosomal RNA synthesis in Caenorhabditis elegans. J. Cell Biol. 140, 1321–1329 (1998). 46. Fatica, A. & Tollervey, D. Making ribosomes. Curr. Opin. Cell. Biol. 14, 313–318 (2002). 47. Draper, D. E. & Reynaldo, L. P. RNA binding strategies of ribosomal proteins. Nucleic Acids Res. 27, 381–388 (1999). 48. Maguire, B. A. & Zimmermann, R. A. The ribosome in focus. Cell 104, 813–816 (2001). 49. Ramakrishnan, V. Ribosome structure and the mechanism of translation. Cell 108, 557–572 (2002). 50. Ferrari, S. et al. Noncoordinated expression of S6, S11, and S14 ribosomal protein genes in leukemic blast cells. Cancer Res. 50, 5825–5828 (1990). 51. Loging, W. T. & Reisman, D. Elevated expression of ribosomal protein genes L37, RPP-1, and S2 in the presence of mutant p53. Cancer Epidemiol. Biomarkers Prev. 8, 1011–1016 (1999). 52. Kondoh, N. et al. Enhanced expression of S8, L12, L23a, L27 and L30 ribosomal protein mRNAs in human hepatocellular carcinoma. Anticancer Res. 21, 2429–2433 (2001). 53. Zhang, L. et al. Gene expression profiles in normal and cancer cells. Science 276, 1268–1272 (1997). 54. Alon, U. et al. Broad patterns of gene expression revealed by clustering analysis of tumor and normal colon tissues probed by oligonucleotide arrays. Proc. Natl Acad. Sci. USA 96, 6745–6750 (1999). 55. Uechi, T., Tanaka, T. & Kenmochi, N. A complete map of the human ribosomal protein genes: assignment of 80 genes to the cytogenetic map and implications for human disorders. Genomics 72, 223–230 (2001). 56. Bassoe, C. F., Bruserud, O., Pryme, I. F. & Vedeler, A. Ribosomal proteins sustain morphology, function and phenotype in acute myeloid leukemia blasts. Leuk. Res. 22, 329–339 (1998). 57. Ferrari, S. et al. Abundance of the primary transcript and its processed product of growth-related genes in normal and leukemic cells during proliferation and differentiation. Cancer Res. 52, 11–16 (1992). 58. Warner, J. R. The economics of ribosome biosynthesis in yeast. Trends Biochem. Sci. 24, 437–440 (1999). 59. Rotenberg, M. O., Moritz, M. & Woolford, J. L. Jr. Depletion of Saccharomyces cerevisiae ribosomal protein L16 causes a decrease in 60S ribosomal subunits and formation of halfmer polyribosomes. Genes Dev. 2, 160–172 (1988). 60. Kongsuwan, K. et al. A Drosophila Minute gene encodes a ribosomal protein. Nature 317, 555–558 (1985). NATURE REVIEWS | C ANCER 61. Kay, M. A. & Jacobs-Lorena, M. Developmental genetics of ribosome synthesis in Drosophila. Trends Genet. 3, 347–351 (1987). 62. Volarevic, S. et al. Proliferation, but not growth, blocked by conditional deletion of 40S ribosomal protein S6. Science 288, 2045–2047 (2000). 63. Terada, N. et al. Rapamycin selectively inhibits translation of mRNAs encoding elongation factors and ribosomal proteins. Proc. Natl Acad. Sci. USA 91, 11477–11481 (1994). 64. Jefferies, H. B., Reinhard, C., Kozma, S. C. & Thomas, G. Rapamycin selectively represses translation of the ‘polypyrimidine tract’ mRNA family. Proc. Natl Acad. Sci. USA 91, 4441–4445 (1994). 65. Jefferies, H. B. et al. Rapamycin suppresses 5′ TOP mRNA translation through inhibition of p70s6k. EMBO J. 16, 3693–3704 (1997). 66. Meyuhas, O. & Hornstein, E. in Translational Control of Gene Expression (eds Sonenberg, N., Hershey, J. W. B. & Mathews, M. B.) (Cold Spring Harbor Laboratory Press, New York, 2000). 67. Anand, N. et al. Protein elongation factor EEF1A2 is a putative oncogene in ovarian cancer. Nature Genet. 31, 301–305 (2002). 68. Thomas, G., Siegmann, M. & Gordon, J. Multiple phosphorylation of ribosomal protein S6 during transition of quiescent 3T3 cells into early G1, and cellular compartmentalization of the phosphate donor. Proc. Natl Acad. Sci. USA 76, 3952–3956 (1979). 69. Gressner, A. M. & Wool, I. G. The phosphorylation of liver ribosomal proteins in vivo. Evidence that only a single small subunit protein (S6) is phosphorylated. J. Biol. Chem. 249, 6917–6925 (1974). 70. Kozma, S. C. Thomas G. p70s6k/p85s6k: mechanism of activation and role in mitogenesis. Cancer Biol. 5, 255–260 (1994). 71. Montagne, J. et al. Drosophila S6 kinase: a regulator of cell size. Science 285, 2126–2129 (1999). 72. Shima, H. et al. Disruption of the p70(s6k)/p85(s6k) gene reveals a small mouse phenotype and a new functional S6 kinase. EMBO J. 17, 6649–6659 (1998). 73. Kawasome, H. et al. Targeted disruption of p70(s6k) defines its role in protein synthesis and rapamycin sensitivity. Proc. Natl Acad. Sci. USA 95, 5033–5038 (1998). 74. Stolovich, M. et al. Transduction of growth or mitogenic signals into translational activation of TOP mRNAs is fully reliant on the phosphatidylinositol 3-kinase-mediated pathway but requires neither S6K1 nor rpS6 phosphorylation. Mol. Cell. Biol. 22, 8101–8113 (2002). 75. Naora, H., Takai, I., Adachi, M. & Naora, H. Altered cellular responses by varying expression of a ribosomal protein gene: sequential coordination of enhancement and suppression of ribosomal protein S3a gene expression induces apoptosis. J. Cell. Biol. 141, 741–753 (1998). 76. Kim, J. et al. Implication of mammalian ribosomal protein S3 in the processing of DNA damage. J. Biol. Chem. 270, 13620–13629 (1995). 77. Boxer, L. M. & Dang, C. V. Translocations involving c-myc and c-myc function. Oncogene 20, 5595–5610 (2001). 78. Coller, H. A. et al. Expression analysis with oligonucleotide microarrays reveals that MYC regulates genes involved in growth, cell cycle, signaling, and adhesion. Proc. Natl Acad. Sci. USA 97, 3260–3265 (2000). 79. Boon, K. et al. N-myc enhances the expression of a large set of genes functioning in ribosome biogenesis and protein synthesis. EMBO J. 20, 1383–1393 (2001). SAGE analysis of MYCN target genes in human neuroblastoma cells reveals that most MYC target genes are involved in ribosome biogenesis and protein synthesis. 80. Menssen, A. & Hermeking, H. Characterization of the cMYC-regulated transcriptome by SAGE: identification and analysis of c-MYC target genes. Proc. Natl Acad. Sci. USA 99, 6274–6279 (2002). 81. Iritani, B. M. & Eisenman, R. N. c-Myc enhances protein synthesis and cell size during B lymphocyte development. Proc. Natl Acad. Sci. USA 96, 13180–13185 (1999). The direct overexpression of c-Myc during B-cell development results in increased cell growth that is independent of the cell cycle and correlates with an increase in total protein synthesis in the cell. 82. Kim, S., Li, Q., Dang, C. V. & Lee, L. A. Induction of ribosomal genes and hepatocyte hypertrophy by adenovirus-mediated expression of c-Myc in vivo. Proc. Natl Acad. Sci. USA 97, 11198–11202 (2000). 83. Iritani, B. M. Modulation of T-lymphocyte development, growth and cell size by the Myc antagonist and transcriptional repressor Mad1. EMBO J. 21, 4820–4830 (2002). 84. Piedra, M. E., Delgado, M. D., Ros, M. A. & Leon, J. c-Myc overexpression increases cell size and impairs cartilage 85. 86. 87. 88. 89. 90. 91. 92. 93. 94. 95. 96. 97. 98. 99. 100. 101. 102. 103. 104. 105. 106. 107. 108. 109. 110. differentiation during chick limb development. Cell Growth Differ. 13, 185–193 (2002). Schmidt, E. V. The role of c-myc in cellular growth control. Oncogene 18, 2988–2996 (1999). Kozma, S. C. & Thomas, G. Regulation of cell size in growth, development and human disease: PI3K, PKB and S6K. Bioessays 24, 65–71 (2002). Di Cristofano, A., Pesce, B., Cordon-Cardo, C. & Pandolfi, P. P. Pten is essential for embryonic development and tumour suppression. Nature Genet. 19, 348–355 (1998). Di Cristofano, A., De Acetis, M., Koff, A., Cordon-Cardo, C. & Pandolfi, P. P. Pten and p27KIP1 cooperate in prostate cancer tumor suppression in the mouse. Nature Genet. 27, 222–224 (2001). Vogt, P. K. PI 3-kinase, mTOR, protein synthesis and cancer. Trends Mol. Med. 7, 482–484 (2001). Backman, S., Stambolic, V. & Mak, T. PTEN function in mammalian cell size regulation. Curr. Opin. Neurobiol. 12, 516–522 (2002). Maehama, T. & Dixon, J. E. The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate. J. Biol. Chem. 273, 13375–13378 (1998). Myers, M. P. et al. The lipid phosphatase activity of PTEN is critical for its tumor supressor function. Proc. Natl Acad. Sci. USA. 95, 13513–13518 (1998). Brown, E. J. et al. Control of p70 s6 kinase by kinase activity of FRAP in vivo. Nature 377, 441–446 (1995). Burnett, P. E., Barrow, R. K., Cohen, N. A., Snyder, S. H. & Sabatini, D. M. RAFT1 phosphorylation of the translational regulators p70 S6 kinase and 4E-BP1. Proc. Natl Acad. Sci. USA 95, 1432–1437 (1998). Isotani, S. et al. Immunopurified mammalian target of rapamycin phosphorylates and activates p70 S6 kinase α in vitro. J. Biol. Chem. 274, 34493–34498 (1999). Sabatini, D. M. Interaction of RAFT1 with gephyrin required for rapamycin-sensitive signaling. Science 284, 1161–1164 (1999). Neshat, M. S. et al. Enhanced sensitivity of PTEN-deficient tumors to inhibition of FRAP/mTOR. Proc. Natl Acad. Sci. USA 98, 10314–10319 (2001). Podsypanina, K. et al. An inhibitor of mTOR reduces neoplasia and normalizes p70/S6 kinase activity in Pten+/− mice. Proc. Natl Acad. Sci. USA 98, 10320–10325 (2001). References 97 and 98 report that Pten-deficient tumours show an increase in mTOR activation and an upregulation in p70S6k activity. These tumours can be effectively treated with the macrolide rapamycin, the cellular target of which is mTOR. Gingras, A. C. et al. 4E-BP1, a repressor of mRNA translation, is phosphorylated and inactivated by the Akt(PKB) signaling pathway. Genes Dev. 12, 502–513 (1998). Sekulic, A. A direct linkage between the phosphoinositide 3-kinase-AKT signaling pathway and the mammalian target of rapamycin in mitogen-stimulated and transformed cells. Cancer Res. 60, 3504–3513 (2000). Pullen, N. et al. Phosphorylation and activation of p70s6k by PDK1. Science 279, 707–710 (1998). Aoki, M., Blazek, E. & Vogt, P. K. A role of the kinase mTOR in cellular transformation induced by the oncoproteins P3k and Akt. Proc. Natl Acad. Sci. USA 98, 136–141 (2001). Podsypanina, K. et al. Mutation of Pten/Mmac1 in mice causes neoplasia in multiple organ systems. Proc. Natl Acad. Sci. USA 96, 1563–1568 (1999). Suzuki, A. High cancer susceptibility and embryonic lethality associated with mutation of the PTEN tumor suppressor gene in mice. Curr. Biol. 8, 1169–1178 (1998). Gomez, M., Sampson, J. & Whittemore, V. The Tuberous Sclerosis Complex (Oxford Univ. Press, 1999). Kwiatkowski, D. J. A mouse model of TSC1 reveals sexdependent lethality from liver hemangiomas, and upregulation of p70S6 kinase activity in Tsc1 null cells. Hum. Mol. Genet. 11, 525–534 (2002). Inoki, K., Li, Y., Zhu, T., Wu, J. & Guan, K. L. TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nature Cell Biol. 4, 648–657 (2002). Gao, X. Tsc tumour suppressor proteins antagonize amino-acid-TOR signalling. Nature Cell Biol. 4, 699–704 (2002). Tee, A. R. Tuberous sclerosis complex-1 and -2 gene products function together to inhibit mammalian target of rapamycin (mTOR)-mediated downstream signaling. Proc. Natl Acad. Sci. USA 99, 13571–13576 (2002). Radimerski, T., Montagne, J., Hemmings-Mieszczak, M. & Thomas, G. Lethality of Drosophila lacking TSC tumor suppressor function rescued by reducing dS6K signaling. Genes Dev. 16, 2627–2632 (2002). VOLUME 3 | MARCH 2003 | 1 9 1 REVIEWS 111. Goncharova, E. A. et al. Tuberin regulates p70 S6 kinase activation and ribosomal protein S6 phosphorylation. A role for the TSC2 tumor suppressor gene in pulmonary lymphangioleiomyomatosis (LAM). J. Biol. Chem. 277, 30958–30967 (2002). 112. Jaeschke, A. Tuberous sclerosis complex tumor suppressor-mediated S6 kinase inhibition by phosphatidylinositide-3-OH kinase is mTOR independent. J. Cell Biol. 159, 217–224 (2002). 113. Lazaris-Karatzas, A., Montine, K. S. & Sonenberg, N. Malignant transformation by a eukaryotic initiation factor subunit that binds to mRNA 5′ cap. Nature 345, 544–547 (1990). The first demonstration that a translation initiation factor shows oncogenic properties when constitutively overexpressed. 114. De Benedetti, A. & Harris, A. L. eIF4E expression in tumors: its possible role in progression of malignancies. Int. J. Biochem. Cell Biol. 31, 59–72 (1999). 115. Gingras, A. C., Raught, B. & Sonenberg, N. eIF4 initiation factors: effectors of mRNA recruitment to ribosomes and regulators of translation. Annu. Rev. Biochem. 68, 913–963 (1999). 116. Ni, J., Tien, A. L. & Fournier, M. J. Small nucleolar RNAs direct site-specific synthesis of pseudouridine in ribosomal RNA. Cell 89, 565–573 (1997). 117. Lafontaine, D. L., Bousquet-Antonelli, C., Henry, Y., Caizergues-Ferrer, M. & Tollervey, D. The box H + ACA snoRNAs carry Cbf5p, the putative rRNA pseudouridine synthase. Genes Dev. 12, 527–537 (1998). 118. Dokal, I. Dyskeratosis congenita in all its forms. Br. J. Haematol. 110, 768–779 (2000). 119. Giordano, E., Peluso, I., Senger, S. & Furia, M. minifly, a Drosophila gene required for ribosome biogenesis, J. Cell Biol. 144, 1123–1133 (1999). 120. Zebarjadian, Y., King, T., Fournier, M. J., Clarke, L. & Carbon, J. Point mutations in yeast CBF5 can abolish in vivo pseudouridylation of rRNA. Mol. Cell. Biol. 19, 7461–7472 (1999). 121. Mitchell, J. R., Wood, E. & Collins, K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature 402, 551–555 (1999). 192 | MARCH 2003 | VOLUME 3 122. Rudolph, K. L. et al. Longevity, stress response, and cancer in aging telomerase-deficient mice. Cell 96, 701–712 (1999). 123. Decatur, W. A. & Fournier, M. J. rRNA modifications and ribosome function. Trends Biochem. Sci. 27, 344–351 (2002). 124. Kullmann, M., Gopfert, U., Siewe, B. & Hengst, L. ELAV/Hu proteins inhibit p27 translation via an IRES element in the p27 5’UTR. Genes Dev. 16, 3087–3099 (2002). 125. Cornelis, S. et al. Identification and characterization of a novel cell cycle-regulated internal ribosome entry site. Mol. Cell 5, 597–605 (2000). 126. Hoang, C. & Ferre-D’Amare, A. R. Cocrystal structure of a tRNA Psi55 pseudouridine synthase: nucleotide flipping by an RNA-modifying enzyme. Cell 107, 929–939 (2001). 127. Vagner, S., Galy, B. & Pyronnet, S. Irresistible IRES. Attracting the translation machinery to internal ribosome entry sites. EMBO Rep. 2, 893–898 (2001). 128. Da Costa, L., Willig, T. N., Fixler, J., Mohandas, N. & Tchernia, G. Diamond-Blackfan anemia. Curr. Opin. Pediatr. 13, 10–15 (2001). 129. Willis, A. E. Translational control of growth factor and protooncogene expression. Int. J. Biochem. Cell. Biol. 31, 73–86 (1999). 130. Spence, J. et al. Cell cycle-regulated modification of the ribosome by a variant multiubiquitin chain. Cell 102, 67–76 (2000). 131. Strezoska, Z., Pestov, D. G. & Lau, L. F. Bop1 is a mouse WD40 repeat nucleolar protein involved in 28S and 5.8S rRNA processing and 60S ribosome biogenesis. Mol. Cell. Biol. 20, 5516–5528 (2000). 132. Pestov, D. G., Strezoska, Z. & Lau, L. F. Evidence of p53dependent cross-talk between ribosome biogenesis and the cell cycle: effects of nucleolar protein Bop1 on G(1)/S transition. Mol. Cell. Biol. 21, 4246–4255 (2001). A defect in ribosome rRNA processing can directly affect cell-cycle progression through a p53dependent process. This is a hallmark study proposing that a nucleolar stress affects cellular proliferation through crosstalk between the ribosome and the cell cycle. 133. Hidalgo, M. & Rowinsky, E. K. The rapamycin-sensitive signal transduction pathway as a target for cancer therapy. Oncogene 19, 6680–6686 (2000). 134. Decatur, W. A. & Fournier, M. J. RNA-guided nucleotide modification of ribosomal and other RNAs. J. Biol. Chem. 278, 695–698 (2003). 135. Manning, B. D., Tee, A. R., Logsdon, M. N., Blenis, J. & Cantley, L. C. Identification of the tuberous sclerosis complex-2 tumor suppressor gene product tuberin as a target of the phosphoinositide 3-kinase/akt pathway. Mol. Cell 10, 151–162 (2002). 136. Potter, C. J., Pedraza, L. G. & Xu, T. Akt regulates growth by directly phosphorylating Tsc2. Nature Cell Biol. 4, 658–665 (2002). References 135 and 136 show that AKT phosphorylates TSC2 and thereby inhibits its function as negative regulator of p70S6k. Acknowledgments We are indebted to M. Barna for input, discussion and critical reading of the review. We apologize to the many scientists whose work we did not cite due to space constraints. This work is supported by the National Cancer Institute, the Mouse Model of Human Cancer Consortium (MMHCC) and the Leukemia Lymphoma Society (LLS). Online links DATABASES The following terms in this article are linked online to: CancerNet: http://www.cancer.gov.search/ leukaemia | small-cell lung carcinoma FlyBase: http://flybase.bio.indiana.edu/ brat LocusLink: http://www.ncbi.nlm.nih.gov/LocusLink CDK2 | CDK4 | cyclin D1 | DKC1 | EEF1A1 | EEF2 | EGF | MYC | NPM | nucleolin | p53 | PDK1 | PTEN | RB | TIF-IA | TIF-IB | TSC1 | TSC2 OMIM: http://www.ncbi.nlm.nih.gov/Omim Diamond–Blackfan anaemia | dyskeratosis congenita | lymphangioleiomyomatosis | tuberous sclerosis Access to this interactive links box is free online. www.nature.com/reviews/cancer