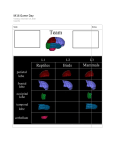
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
PHYSIOL0GICU REVIEWS Vol. 63, No. 1, January Printedin USA. 1983 Gas Exchange and Control of Breathing in Reptiles MOGENS L. GLASS AND STEPHEN C. WOOD I. Introduction ......................................................... A. Patterns of breathing ............................................. B. Gasexchange ..................................................... C. Ventilation-perfusion ratio ........................................ D. Pulmonary diffusing capacity ...................................... E. Control of breathing .............................................. F. Respiratory symbols .............................................. II. Ventilation .......................................................... ...................................................... A. Introduction B. Breathing patterns ................................................ C. Scaling of ventilation and air convection requirements .............. III. Gas Exchange ........................................................ A. Introduction ...................................................... B. Oxygen uptake during rest and activity ............................ C. Gas-exchange ratios ............................................... D. Diffusing capacity of the lungs ..................................... E. Ventilation-perfusion matching .................................... F. Nonpulmonary gas exchange ....................................... G. Gas exchange in periodic breathing ................................ IV. Effects of Temperature on Control of Breathing ........................ A. Concepts of constant relative alkalinity and alphastat regulation .... B. Consequences for ventilation and gas exchange ..................... C. Exceptions to alphastat regulation and constant relative alkalinity .. V. Control of Breathing at Constant Body Temperature ................... A. Ventilatory responses to carbon dioxide inhalation .................. B. Anoxia and hypoxia: tolerance and regulatory responses ............ C. Receptor systems ................................................. VI. Conclusions .......................................................... 232 233 236 237 238 238 239 241 242 243 245 246 246 247 248 I. INTRODUCTION This article reviews the respiratory physiology of reptiles, specifically ventilation, gas exchange, and control of breathing. We do not attempt to present a comprehensive or encyclopedic review but instead use data selected from the literature to illustrate major points. The diversity of modern reptiles and their many unique physiological and biochemical traits make them attractive as experimental animals. These 232 0031-9333/83/000099-06$01.50 Copyright 0 1983 the American Physiological Society Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Abteilung Phgsiologie, Max-Planck-Institut fiir experiwmatelle Medixin, G3tingen, West Gemnany; and Department of Phgsiologg, School of Medicine, University of N&w Mexico, Albuquerque, New Mexico January 1983 RESPIRATION IN 233 REPTILES features, however, also create problems in experimental design and interpretation of data. In fact the problems associated with interpreting reptilian physiology emerge as the central theme in this review. To pique the reader’s interest, perhaps it is worth summarizing these problems at the outset. A. Patterns of Breathing B. Gas Excharzge Most equations used by mammalian physiologists to analyze gas exchange are valid only under so-called steady-state conditions, i.e., when the tissue respiratory quotient (RQ) equals the respiratory exchange ratio (RE) = CO2 production (ko2)/02 uptake (VO,)]. This creates a problem in CRE analyzing reptilian data: steady state is difficult to determine and is an infrequent condition for reptiles. In reptiles, RE frequently deviates from RQ owing to phenomena such as cutaneous CO2 elimination, production of ammonium bicarbonate in urine, and breath holding. A further complication is the high anaerobic capacity of many reptiles. Alligators, for example, take up to 12 h to repay an O2 debt. Turtles can tolerate complete anoxia for days or months, accumulating lactic acid levels up to 200 mmol liter-l, and can survive a decrease in brain pH to 6.4. l C. Ventilation-Perfusion Ratio This concept presents a real problem when applied to reptiles because both ventilation (V) and pulmonary blood flow (QL) are often periodic. The presence of large and variable central vascular shunts provides reptiles with a flexible circulatory system. Although somewhat disadvantageous to O2 transport, the central shunts permit the pulmonary circulation to be both in series and in parallel with systemic circulation. For diving species this provides a redistribution of blood away from the lungs during submersion. Obviously, the concept of %VQL matching as the important determinant of gas exchange in mammalian lungs must be cautiously and carefully applied to reptiles. Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Show the respiratory trace of a normal turtle to a group of physicians, and they will probably identify it as a case of Biot’s breathing, a pathological pattern of breathing in humans in which breathing episodes and breath holds alternate. Such periodic breathing is found in most reptiles and creates a problem in characterizing minute ventilation as the product of tidal volume (VT) and breathing frequency (f). For example, a reptile may increase minute ventilation simply by reducing or eliminating the breath holds, without changing VT or f during the ventilatory periods (VPs). M. L. GLASS AND Vdume S. C. WOOD 68 E. Cimtrol of Breathing This area of reptilian respiratory physiology creates some interesting problems of data interpretation. For example, if CO2 is added to the inspired gas of reptiles, some species respond with the increase in VT expected by mammalian physiologists but show a paradoxical decrease in f and a net decrease in minute ventilation leading to aggravated CO2 retention. An interesting reptilian “trick” is the difference in control of breathing between conditions of constant and increasing body temperature. A given species may show increased ventilation when given exogenous CO2 at constant body temperature; however, if CO2 increases because of increased metabolism as temperature goes up, the same species shows no ventilatory response. F. Reqkatmq Symbols The use of equations is necessary to explain Common symbols are listed below. A a &I3 BTPS C DL Dko -2 Eoz f I M NVP basic respiratory concepts. alveolar arterial solubility of a gas in blood or water air convection requirement body temperature and pressure, saturated with water vapor concentration of a gas diffusing capacity of the lung carbon monoxide (CO) diffusing capacity of the lung O2 diffusing capacity of the lung pulmonary O2 extraction breathing frequency inspired body mass nonventilatory period, i.e., breath-hold period Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 The relatively low O2 consumption of reptiles compared with that of mammals of similar size (with both groups at 37°C) prompted some-authors to look at pulmonary diffusing capacity (DL) as a probable limiting factor. The presence of central vascular shunts creates considerable problems in assessing the importance of DL in reptiles. Even if DL were infinite, the admixture of venous blood would produce arterial desaturation. Thus the finding of PA% - Ph values [i.e., alveolar-arterial differences in O2 partial pressure (POT)]of 40-60 Torr in normal reptiles cannot be ascribed to inadequate lung architecture alone. January 1983 P Pa co2 PACO, Pa 02 PA 02 02 - pa02 PC02 PA PK' QL R RE RQ STPD T V. V WQL . VA vco . 2 VD . VE . vo 2 VT VP II. IN REPTILES 235 gas pressure (total or partial) partial pressure of CO2 in arterial blood partial pressure of CO2 in alveoli partial pressure of O2 in arterial blood partial pressure of O2 in alveoli alveolar-arterial difference in partial pressure of O2 partial pressure of CO2 apparent first dissociation constant of the bicarbonate buffer system partial pressure of O2 pulmonary blood flow gas constant gas-exchange ratio for a gas exchanger, i.e., ratio of CO2 output to O2 uptake respiratory quotient standard temperature and pressure (OOC, 760 Torr) and dry absolute temperature gas volume gas volume per unit time or ventilation ventilation-perfusion ratio alveolar ventilation, i.e., volume expired from the alveolar compartment per unit time co2 output dead-space ventilation total ventilation, i.e., total expired volume per unit time O2 uptake tidal volume, usually calculated as a mean value for expired volume ventilatory period VENTILATION A. Introduction Like birds and mammals, reptiles ventilate the lung by a suction-pump mechanism. Thus inspiratory flow is caused by the development of subatmospheric pressures within the lung (cf. 84). Pressure gradients result from rib cage movements alone because the diaphragm is absent in reptiles (121). The basic respiratory flow pattern in reptiles consists of an exhalation followed by inspiration. If the glottis closes at end expiration, a breath-hold period follows (30,36,53). Some studies report a second expiration preceding breath holds (91); however, these triphasic flow patterns apparently result Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 PO . 2 RESPIRATION 236 M. L. GLASS AND S. C. WOOD ih?ume 63 from excitation of the animal or are artifacts of the methods used to measure ventilation (36). Like mammals, reptiles have dead-space ventilation (VD), as well as alveolar ventilation (VA), i.e., ventilation of the gas-exchange surfaces (cf. 78). The value of VA determines intrapulmonary gas partial pressures and is consequently more important than total ventilation (VE) in relation to control of breathing and gas exchange. Unfortunately most of the data available are for TE. Patterns Reptilian breathing patterns may surprise those who are familiar with the continuous breathing activity typical of nondiving mammals and birds. In reptiles, breathing invariably alternates with periods of breath holding. Breath holds may be interrupted by a series of breaths (2, 24, 64, 88, 109, 117). This pattern is typical of some aquatic reptiles and persists when the animal is out of the water (cf. 60,84). An alternative breathing pattern that consists of single breaths interrupted by relatively short breath holds has been reported in some chelonians (57,87), lizards (118,154), and snakes (67). These two alternative breathing patterns are shown in Figure 1. Interruption of breathing activity by breath holds appears to be a general characteristic of pulmonary ventilation in ectothermic vertebrates. In amphibians pulmonary ventilation may alternate with periods during which only the buccal cavity is ventilated (96). Ventilation is also intermittent in lungfish (105). Discontinuous breathing patterns may even occur in nondiving mammals under some extreme conditions; for example, when undisturbed hibernating hedgehogs are in deep hypothermia, series of breaths are interrupted by protracted periods of breath holding (100). Reptilian breathing patterns are difficult to characterize in terms of f and VT. In mammalian studies, VE is conventionally reported as minute volume, i.e., the volume expired during a l-min period (VE = f VT). This concept is not satisfactory for description off in reptiles because some species may breath hold for hours (cf. 8). An alternative approach in describing f in reptiles is to quantitate the breath-hold period [nonventilatory period (NVP)], i.e., f,,,, = fvr [VP/(VP + NVP)], where fmean is mean breathing frequency, fvr is breathing frequency during ventilation, VP is duration of ventilatory periods, and NVP is duration of nonventilatory periods. Breathing activity may occupy a small fraction of total time. At 25°C the ratio VP/ (VP + NVP) is 0.38 in Caiman sclerops (ll?), 0.18 in Crocodylus niloticus (60), and 0.10 in the snake Acrochordus javanicus (59). Breath-hold duration decreases with increased body temperature (1,59,107), but even at 20°C breath holds may exceed 1 h. For example, dive duration in Pseudemys concinna may reach 141 min (8). An accurate description of ventilation in reptiles may therefore require measurements over hours or even days. Ventilation l l Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 B. Breathing Jammy 1983 RESPIRATION IN 237 REPTILES Crocodylus Test udo 1 min FIG. 1. Typical breathing patterns of reptiles. U’@pey trace, ventilation crocodile (Crocodylus niloticus); lower trace, breathing pattern of a 3-kg d&s). Both species inspire before breath holding. Measurements were tachography on resting unrestrained animals. I in a young, 5-kg Nile tortoise (Testudo parobtained by pneumo- should be characterized by VP and NVP and not only by VT and an overall f value. The need to measure over prolonged periods is emphasized by reports of large diurnal variations in ventilation (123, 154). Problems is assessing resting values for f and VT in reptiles may be caused by the methods used to measure ventilation. Reptiles are often sensitive to disturbances, which may prevent measurement of true resting ventilation (1, 81, 117, 130). Even when noise and disturbances are kept at a minimum, resting values may not be achieved, because measurement of ventilation may involve restraints and unnatural conditions that may influence the reptile’s behavior. A quiet animal is not necessarily resting if it is restrained. For example, Cragg (36) measured ventilation in the small lizard Lace&x viridis (-30 g). Minimum values for ventilation of sleeping animals were less than half as large as values measured during the daytime on restrained but quiet lizards. Some studies report unpredictable breathing patterns (117, 122). Such patterns would conform to the idea that reptiles are animals without precise control of breathing. Alternatively we suggest that some erratic breathing patterns result from inadequate methods of measuring ventilation. The finding that breathing patterns may be very regular if measurements are taken with the least possible disturbance of the animal (59) supports our theory. C. Scaling of Ventilation and Air Convection Requirements It may be difficult to establish true resting values for ventilation in reptiles, but all studies are consistent in reporting resting values for ventilation that are markedly lower than those reported, for similar-sized mam- Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 I 238 M. L. GLASS AND S. C. WOOD Votaww 63 l l l III. GAS EXCHANGE A. Introduction The gas-exchange surfaces of reptilian lungs are covered by faveoli, which are honeycomblike extensions of the pulmonary wall (46, 121). Rep- Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 mals or birds. Most data are available for lizards, for which Bennett (14) derived an interspecies relation (y) expressing the dependence of ventilation on body mass. Such relations take the general form y = aMb, where M = body mass (in kilograms) and a and b are empirically derived constants. For ventilation of lizards at a body temperature of 3537”C, Bennett calculated the relation VE = 4,614 Mo*76 (where VE is measured in ml BTPWh). In reptiles this relation predicts ventilation volumes that are about 20% of the values for mammals (143). Some later studies have confirmed predictions for ventilation (56, 154). In contrast, Bennett (14) also attempted to derive interspecies relations for the subparameters of ventilation, i.e., VT and f. These predictions contrast considerably with later studies (cf. X9), perhaps because the relationships between VT and f can be highly flexible in lizards (63). It is presently impossible to derive reliable relations for scaling of ventilation in nonsaurian reptiles because data are incomplete. The relationship between v and 00~ of a gas exchanger is expressed as . = v CI~, Eo,, where CI~, is the inspired O2 concentration and Eo, is vo pulmonary O2 extraction. This equation may be rearranged to (~/TO& CI~, Eo, = 1 (cf. 43). The ratio of ventilation to O2 uptake (~/VO,, measured in ml BTPS/ml STPD) is termed the air convection requirement (ACR) because it represents the ventilation volume required to extract a unit amount of 02. The equation shows that ACR and Eo, are inversely related. Obviously the degree of ex .traction of O2 from the inspired gas is an important variable in determi ning PACT, and recent studies have emphasized the role of ACR in respiratory control (cf. 83). Bennett (14) compiled data for ACR in lizards and concluded that ACRs did not vary significantly with M. Lizards resemble birds and mammals in this regard (43,102,138,143). Reptilian ventilation and the ACRs may change with temperature (see sect. IvB). Bennett (14) compared lizards at about 37OC to mammals and found almost identical values for ACR. Thus the interspecies relation to lizards predicts that ACR = 37.7 ml BTPWml STPD against 35.5 ml BTPWml STPD for mammals (43). The ACR of chelonians is generally lower than that for lizards. At 35OC ACR is 10.8 ml BTPWml STPD in Pseudemys scripta (81) and about 25 ml BTPWml STPD in Pseudemysjbridana (98). This is consistent with predictions by Howell and Rahn (78). They derived equations to show *that ACR is inversely related to the blood bicarbonate ion concentration ([HCOJ). Since plasma [HCO,] is generally greater in chelonians than in lizards (44), a difference in ACR is predicted. Data for v and TO, of snakes and crocodilians are too scarce to allow general statements. January 1983 RESPIRATION IN REPTILES 239 B. Oxygen Uptake During Rest and Activity Bennett and Dawson (16) reviewed data for vo2 in reptiles and calculated interspecies relations for scaling of resting VO, at several body temperatures. They recognized that the data were of limited usefulness because few studies report vo2 on undisturbed reptiles resting in the dark. Despite this finding, some preliminary conclusions can be derived from these data. Dependence of resting-state VO, on M is expressed by the relation vo2 = aM! The ex- Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 tilian lungs range in complexity from simple saclike structures, as in the tegu lizard Tupinambis teguixin (99), to multicameral lungs, as in varanid lizards (121) or in the turtle Pseudemys sctipta (120). Thus the gas-exchange units in reptilian lungs are not alveoli as in mammals. We emphasize this point because discussions of gas exchange in reptiles may include topics such as alveolar ventilation or alveolar gas partial pressure. It might seem appropriate to substitute the termfaveoZar for alveolar in discussing reptilian ventilation. Introduction of a new term is unnecessary, however, because the alveolar and faveolar compartments are functi .onally identical. Some difficulties in describing venti lation in reptiles also apply to gas exchange. A fundamental problem is the acquisition of true resting values for voz or CO2 elimination (ho2). Note that resting values are not always obtained even when a reptile has been quiet for hours. Food intake in reptiles may cause pronounced metabolic changes that persist during the digestive process, lasting days (12, 23, 34, 35, 63). For example, 60, in snakes may increase several-fold during periods of digestion (12). Other problems arise from discontinuous breathing patterns. These problems are aggravated at low temperatures, because breathing activity may be absent for hours (1, 58). Clearly a l-h measurement could lead to highly unreliable conclusions, e.g., that the animal is not consuming OZ. Diurnal changes in 60, may also occur (23, 68, 154, 158). A useful variable in description of mammalian gas exchange is the pulmonary ventilation-perfusion ratio (~/QL), which is the ratio of i7A (measured in ml BTPS/min) to pulmonary perfusion (measured in ml BTPS/min). In healthy mammals this ratio is close to unity (128, 148). In reptiles the concept of ~/QL is problematic because the lungs are ventilated periodically, and perfusion is also quite variable. Another useful concept in mammalian physiology is that of DL (cf. 51). This variable was originally defined as a diffusive conductance for gas transfer through the pulmonary membrane, i.e., the tissue barrier separating alveolar gas and pulmonary capillary blood (20). All the above concepts have been applied in discussions of reptilian gas exchange (83,120,154,159). In this review we discuss the validity of applying these traditional concepts to reptiles. In addition, we discuss some aspects of reptilian gas excha nge that seem exotic but that may be interesting and en lightening to those primaril .y fam iliar w ith mam malian physiology. 240 M. L. GLASS AND S. C. WOOD Volume 63 Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 ponent b has attracted the most interest because it is close to 0.75 in overall relations based on large groups of organisms (72). Bennett and Dawson calculated the following interspecies relation for scaling of resting state 90, at 20°C as (TO2 = ml OZ/h) = 25.6 MO**‘. Hemmingsen (72) had previously derived a general relation for scaling of the TO, of ectothermic animals at 20°C. Comparing the two relations, Bennett and Dawson (16) concluded that reptiles resemble other ectothermic vertebrates in general metabolic level. This conclusion is not too surprising, because Hemmingsen included data for reptiles in his calculations for the more general relation. The value of 00, in reptiles increases predictably with body temperature. The Qlo values for the increase of resting VO, are frequently 2-3, but exceptions occur. In the range of temperatures at which a reptile is normally active, Qlo may decrease and approach 1. Examples have been reported for lizards (cf. 125), chelonians (54), a crocodilian (23), and snakes (42, 90). This effect is species specific. For example, the To2 value of Pseudemys scripta is a simple exponential function of temperature, whereas the vo2 value of the box turtle Terrapene ornata changes little between 20 and 30°C but changes markedly with temperatures outside that range (54). At body temperatures of 37-40°C the weight-specific VO, of a reptile is much less than that of an endothermic vertebrate. Bennett and Dawson (16) calculated that a l-kg lizard at 37OC consumes 122 ml OJh, which is 18% of the vo2 for a l-kg mammal (143). The difference in 60, values between reptiles and endotherms correlates well with variations in values for fundamental variables in pulmonary function. The ventilation volume of a reptile is about 20% of the value for a mammal of similar size. Similar differences exist for pulmonary perfusion and DL (see sect. III@. The different demands for O2 of reptiles and mammals are also reflected at the tissue level. Bennett (13) reported that the number of mitochondria per unit weight of tissue is greater in mammals than in reptiles but did not provide quantitative evidence. A recent study by Else and Hulbert (48) compared mitochondrial volume density in tissues of the lizard Amphibolurus nuchalis to values for a mammal of similar size (Mus); they found the greatest mitochondrial volume density in the mammal, which also had the greatest mitochondrial membrane surface area. Another important difference at the tissue level is the amount of capillary surface area available per unit volume of tissue. Pough (124) estimates that the capillary surface area per unit muscle weight of a reptile is about 20% of the value for a mammal of similar size. Some reptiles can expand voz values considerably relative to resting values. The turtle Pseudemys scripta, for example, may increase vo2 ZO-fold (54). Maximum 00, of reptiles increases with body temperature (cf. 125), which implies that the capacity for sustained exercise is greatest at high temperatures. Interestingly, Moberly (116) found that the maximum sustainable walking speed of Iguana iguana nearly tripled with an increase of body temperature from 20 to 40°C. The range at which the greatest increase of aerobic metabolism is possible may correspond to the active temperature January RESPIRATION 1983 IN REPTILES 241 C. Gas-Exchange Ratios Values for the pulmonary RE in mammals reflect (at steady state) the stoichiometry of reactions in food catabolism. Therefore values of RE outside the range of 0.7-1.0 are not expected. In reptiles the RE value may be 0.71.0 (cf. 16), but values lower than 0.7 have been reported. The pulmonary RE value of CrocodyZus ~orosus may be as low as 0.32 (69). In crocodilians a decrease in pulmonary RE values relative to the tissue RQs can be due to incorporation of COZ into the urine to form ammonium bicarbonate (34, 69). This mechanism has only been studied in crocodilians, but it probably is not important in all reptiles because nitrogen excretion in the form of ammonia is negligible in many terrestrial species (cf. 39). In some reptiles, gas exchange may occur through the skin (see sect. 1113’). This also decreases the overall pulmonary exchange ratio (cf. 43). Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 range (15, 125). More studies will be needed to assess whether the temperature dependence of maximum 00, is limiting for the activity of reptiles in their normal habitat. Despite the ability of reptiles to increase vo2 by several times, aerobic metabolism may account for less than half of the energy expenditure measured at the highest metabolic rate (54, 68, 135). When, for example, the water snake Natrix rhombifera was stimulated to maximal activity for 10 min, anaerobic metabolism accounted for as much as 86% of total energy consumption (68). Coulson and Herbert (33) have pointed out that most animals are provided with similar amounts of usable muscle glycogen to provide fuel for a burst of intensive muscular activity. Therefore the anaerobic power available for a short, intensive activity burst is not very different for a 70kg crocodile or a 70-kg human. If activity is prolonged, however, the lower aerobic capacity of the reptile will lead to development of a considerable O2 debt, which will be repaid very slowly. For example, elevated VO, indicated repayment of an O2 debt in an alligator for no less than 12 h after cessation of exercise (35). Recovery time depends on the weight-specific VO, of the animals and will be most time-consuming in large reptiles because weightspecific 00, decreases with increased body weight (35). The capacity for increasing VO, relative to resting values may depend on body weight. Bennett and Dawson (16) compiled data for VO, in reptiles with M values ranging from 4 to 795 g and found that the ratio of maximum vo2 to resting vo2 was independent of M, i.e., the exponents for scaling of resting vo2 and maximum To2 values are the same. This conclusion may be premature because the resting and maximum 00, values were not derived for the same combination of species and because the analyzed weight range was limited. In addition the conclusion conflicts with data for scaling of vo2 in large chelonians such as the Aldabra giant tortoise [Testudo gigantea (79)] and the green sea turtle [Chelonia mydas (126)]. In these chelonians the capacity to expand metabolism increases with body weight. More intraspecies studies are needed to establish if scaling of vo2 in these chelonians is unique. 242 M. L. GLASS AND S. C. WOOD Vohme 63 D. Di$ikag Capacity of the Lungs In 1909, Bohr (20) derived equations to describe the diffusive conductance of a pulmonary membrane. For To2 an equation states TO, = DL~~ APo2, where DL~~ is the O2 diffusing capacity of the pulmonary membrane and APO, is the mean difference in PO, between alveolar gas and pulmonary capillary blood. Thus a low DL o2 relative to vo2 could lead to large PO, differences between alveolar gas and arterial blood. The DL value increases with surface area and is inversely related to diffusion distance (cf. 43). Morphometric measurements have provided estimates of DL values and show a difference of an order of magnitude in the DL values of reptiles and mammals (120). It is possible to measure DL directly by a method introduced by Krogh and Krogh (lOl), but in that technique DL is measured for carbon monoxide (CO) and not for OZ. Crawford et al. (37) measured DL~~ in small chelonians and reported values ranging from 0.033 to 0.114 ml CO kg-l mine1 Torr-l, corresponding to 535% of values for mammals of similar size (145). This order of magnitude is very likely correct, but it is problematic that Crawford et al. only measured DL co at one temp,erature (ZO-23°C) because reptiles are functional over a wide range of body temperatures. A recent study reports that DL~~ in lizards increases markedly with temperature (62); this emphasizes that all variables in pulmonary function of reptiles must be reported at specified body temperatures. At 35-37OC the DL~~ value of lizards is about 20% of values for similar-sized mammals (62). This value closely matches a fivefold difference in resting vo2. Thus the ratio of DL~O to 60, is much the same in lizards and mammals. Clearly it would be more useful to know the DL values for respiratory gases (02 and C02) than to know the values for CO, but direct measurements are complicated (cf. 4). Recently obtained direct measurements of pulmonary variables in the tegu lizard Tupinambis teguixin show that DL~~ increases with temperature and is of the same-order of magnitude as DL~~(61). l l l l Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Some non-steady-state conditions influence the pulmonary RE. 1) Breath holding causes a gradual decrease in RE. Conversely, RE increases during breathing activity (see sect. IIIG). 2) Changes in activity level affect RE because COZ is released during exercise and replenished during recovery (89, i26). This situation may in part result from a decrease in NVP during exercise, but it could also be caused by metabolic changes. 3) Transient effects of temperature changes influence RE (cf. 16). 4) Altered feeding status may decrease values for RE as a consequence of changes in acid-base status; such changes are caused by the large amounts of acid needed for digestion in reptiles that swallow whole animals or large chunks of meat (34). Thus several explanations should be considered if low RE values are measured in reptiles. In addition such measurements should be performed over prolonged periods, preferably days, to establish if steady-state conditions exist or if low RE values are transient. January 1983 RESPIRATION IN REPTILES 243 E. Ventilation-Perfusion Matching Pulmonary gas exchange is ineffective if perfusion of the lung is not matched quantitatively with ventilation (cf. 148). Therefore it is significant that pulmonary perfusion is also periodic in reptiles. Perfusion is greatest during and immediately after breathing, and it decreases during breath holds (26,28,93,141,149,151,152). These changes in perfusion of the lungs correlate well with adjustments of heart rate and stroke volume. However, perfusion of the lungs also depends on selective redistribution of cardiac output between systemic and pulmonary circulation (cf. 151). The anatomical basis is incomplete separation of systemic and pulmonary circulations. Thus the ventricle is incompletely divided in noncrocodilian reptiles (cf. 151). During breath holds, blood is progressively shunted away from the pulmonary circulation (right-to-left shunt), whereas during breathing activity the net blood shunt may be reversed. The decrease in pulmonary perfusion during breath holds is partly related to a progressive increase of pulmonary resistance that is cholinergically mediated in chelonians (25). Matching of ventilation and perfusion during breathing activity may involve stretch receptors in the lungs rather than chemoreceptors. In Pseudemgs sctipta artificial ventilation of the lungs causes increased perfusion. In contrast, sudden changes in alveolar gas composition have no effect on perfusion (92). Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Once values for DL~~ are established for various reptiles, one must still ask whether DL~$ is a useful variable in describing reptilian gas exchange. This question remains because the PACT- Paoz value is determined not only by the diffusive conductance of the pulmonary gas exchanger but also by the central cardiovascular admixture of systemic venous blood into the systemic arteries (see sect. IIIE). The PACT- PaoZ value may be as large as 45-50 Torr in chelonians (28) and may exceed 60 Torr in sea snakes (140). Mitchell et al. (114) have recently reported PACT - Pao, values of lo-20 Torr in resting lizards. These values could be explained on basis of poor DL values, as Mitchell’s group suggested. But because present data on DL in reptiles indicate values that are not particularly low relative to To2 (62), it seems more likely that large PACT- PaoZ values develop as a consequence of central cardiovascular mixing of blood. In addition the shunting of blood within the pulmonary circulation could prevent equilibration between a fraction of pulmonary arterial blood and alveolar gas. In a recent study of the sea snake Laticauda culobina, Seymour (139) concluded that large PACT- Paoz values primarily result from shunting of blood within the pulmonary circulation and from central vascular mixing of blood, whereas the diffusive resistance of the pulmonary membrane is relatively unimportant in creating PO, differences. Some caution is required in generalizing these results because the reptiles are a diverse group and because further information on temperature effects is desirable for evaluation of the causes for nonequilibration between alveolar gas and systemic arterial blood. 244 M.L.GLASSAND S.C.WOOD Vohme 63 F. Nonpulmonary Gas Exchange In 1960, Mertens (110) stated that the skin of reptiles is “almost impervious to water” and “of scant importance for respiration.” These statements compared reptiles with amphibians, which lose water from the skin at rates of the same magnitude as those for free water surfaces (138) and which may also have high rates of cutaneous respiration (cf. 43). This schematic distinction between amphibians and reptiles is not valid in light of studies published since 1960. Romer (134) discussed fossil evidence that Carboniferous amphibians were completely scaled. He suggested that the highly water-permeable skin of modern amphibians represents a specialized secondary condition. Moreover reptilian integuments are not impermeable to water. This is especially true for species that occupy aquatic habitats. No less than 70-90% of total evaporative water loss may be cutaneous in caimans and in Pseudemys scripta (cf. 17). Respiratory gases may also permeate the skin. Nonpulmonary CO2 elimination is substantial in many aquatic reptiles. For example, the nonpulmonary CO2 elimination is 65% of total COZ output in the aquatic turtle Trionyx mucita (85), and up to 94% of COB output is cutaneous in the sea snake PeZamis pZaturus (65). Nonpulmonary O2 uptake is usually a small fraction of total uptake. This observation agrees with a general principle: in bimodal breathers the nonpulmonary CO2 output is greater than nonpulmonary O2 uptake (cf. 43). In the aquatic snake Acrochordus javanicus cutaneous respiration accounts for up to 33% of CO2 elimination but 8% of O2 uptake (144). In the freshwater turtle Sternothaerus minor (at 22°C) and in the sea snake PeZamis pZaturus (30°C) the extrapul- Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 The concept of ~/QL is a complicated idea to apply to reptiles because these ratios can be based on mean long-term values for ventilation and perfusion, or, alternatively, on values measured only during breathing activity. The value for ~/QL ratio in mammals is close to unity. A temperatureindependent value of 1.2 during breathing has been reported for Varanus exanthematicus, which has the reputation of being an active mammallike lizard (154, 156). Larger deviations from a 1:l ratio are found in turtles (26, 98). Note that ~/QL may change with temperature. Several studies report that pulmonary perfusion increases in proportion to 60, when the body temperature of a reptile increases (62, 98, 156; R. N. Gatz, M. L. Glass, and S. C. Wood, manuscript in preparation). In most reptiles ventilation increases less than 00, with increases in body temperature, i.e., the ACRs are inversely related to temperature (cf. 83). Kinney’ and colleagues (98) studied the consequences of these relationships in Pseudemysjbridana, in which an overall ~/QL value decreased from 1.16 at 10°C to 0.62 at 35OC. These changes in ~/QL indicate that when body temperature changes, ventilation and pulmonary perfusion are independently regulated relative to vo2. January RESPIRATION 1983 IN REPTILES 245 G. Gas Exchange in Periodic Breathing Weight-specific lung volumes of reptiles are often large relative to those of mammals (37,62). Consequently the lungs may serve as a significant store during breath holds. In the alligator the pulmonary O2 store is 85% of the total (6). A calculation further illustrates the significance of the pulmonary O2 store. Crawford et al. (37) found a lung volume of 160 ml/kg in the semiaquatic turtle Pseudemys scripta and an O2 transfer from alveolar gas to blood of 0.9 ml kg-’ min-’ at ZO-23OC. With an alveolar PACT of about 120 Torr during ventilation preceding its dives (27), the turtle can dive with an initial O2 store of about 24 ml/kg, which, if fully depleted, would suffice for O2 delivery to the blood for 27 min. The pulmonary O2 store usually is not fully used, and voluntary dives in Pseudemys end when PACTfalls to about 22 Torr (1). The O2 demands may be smaller than those Crawford et al. (37) measured,“especially in turtles resting at low temperatures, where the pulmonary O2 store may meet aerobic demands for hours (I). l l Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 monary O2 uptake is sufficient to prolong underwater survival time (9, 65), but the increase is only 1 h or less with the water Po2 at 150 Torr. In vertebrates that rely on a combination of pulmonary and cutaneous gas exchange, the latter becomes relatively more important at low body temperatures (cf. 43) because the values for cutaneous gas exchange are relatively independent of temperature, whereas pulmonary vo2 and ho2 increase with body temperature (cf. 83). Some turtles can remain submerged for months if the water temperature is close to the freezing point (29). A recent study (147) shows that the low metabolic needs of turtles at temperatures close to freezing may be covered by anaerobic stores alone, but surnonvival is significantly promoted if water PO, is high, thus permitting pulmonary O2 uptake. The integuments of land-dwelling reptiles are much less permeable to water than those of aquatic species. This condition limits cutaneous evaporation (17) but also reduces cutaneous gas exchange. At 20°C only about 3% of CO2 output is cutaneous in the tortoise Testudo dendricuzata (85). In the chuckawalla (SaurmaZus obesus, a desert lizard) about 4% of CO2 output and less than 2% of O2 uptake take place through the skin at 25°C (38). Even in land-dwelling reptiles, however, cutaneous CO2 elimination may become important at low temperatures. In the box turtle Terrapene ornata a large fraction of CO2 output is cutaneous when the turtles are hibernating at 5OC (58). Nonpulmonary gas exchange in reptiles is not always cutaneous. Pharyngeal respiration occurs during submergence in the soft-shelled turtle Trionyx spinifer and can be seen as rapid movements of the hyoid apparatus (47). Such specialized sites for nonpulmonary gas exchange may have been overlooked in studies that assumed nonpulmonary gas exchange occurs only through the skin. 246 M. L. GLASS AND S. C. WOOD Volume 63 IV. EFFECTS OF TEMPERATURE ON CONTROL A. Concepts of Constant Relative Alphastat Regulation OF BREATHING AlkCclinity and In many ectothermic vertebrates the arterial pH (pH,) decreases with an increase in body temperature. Some researchers interpret this effect as a preservation of constant relative alkalinity, i.e., OH-/H+ (77, 78,129,130). Because the ionization constant of water (K. = [H+][OH-1) increases with temperature, constant water OH-/H+ requires increases in [H+] and [OH-] with temperature. Since [H+] = [OH-], the neutral pH of water (PA&) at any temperature is defined as pN, = 0.5 pK,. A constant relative alkalinity requires that arterial pH at any temperature be equal to the neutral pH of Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Numerous studies describe pulmonary gas exchange during breath holds in reptiles (1, 6, 18, 28, 45, 60, 67, 106, 140). All agree in reporting a gradual decrease in the pulmonary RE value with breath holding, i.e., the amount of O2 transferred from alveolar gas to blood is replaced by decreasing amounts of C02. This effect of breath holding on RE is consistent with data for mammals (cf. 115). During breath holding in mammals the CO2 gradient between mixed venous blood and alveolar gas gradually decreases because of the cessation of CO2 removal from the lung. This in turn reduces COZ entry into the alveolar gas from the blood. Oxygen continues to be removed from the alveolar gas via the pulmonary circulation to meet the requirements of aerobic metabolism. As a result the lungs shrink, which concentrates CO2 in the alveoli and increases PA coZ. This further diminishes the partial pressure of CO2 (Pco~) difference between mixed venous blood and alveolar gas and reduces CO2 entry into the alveolar compartment. Thus a self-perpetuating cycle is in operation (cf. 115). This explanation of gas exchange during breath holds applies to reptiles, although the existence of cutaneous respiration may complicate the interpretation. In Pseudemys at 20°C only about 10% of CO2 output is nonpulmonary (85). This amount is too small to explain decreases in RE during breath holds (dives) to values as low as 0.2, and it strongly indicates CO2 loading in tissues (2). This interpretation receives support from measurements of gas exchange during postdive ventilation (1,28). Initially RE is low, but it increases during breathing, indicating a gradual release of tissue CO2 stores. This observation again emphasizes the difficulty of applying the concept of steady state to reptiles and the caution that overall values for gas exchange should be based on measurements over hours or days. The decrease of RE during dives in the turtle CheZys fimbriata has been attributed to nonpulmonary COZ elimination (106). To find out if CO2 is stored in tissues or eliminated in the water, however, it is necessary to measure RE over a prolonged period. January 1983 RESPIRATION IN REPTILES 247 B. Consequences fbr Ventilution From und Gus Exchunge equation pH = pK’ + log ([HCO;]/ of COZ in blood, both pK’ and CVCO~ decrease with increased temperature (132). A regulated decrease of pH with increase of body temperature, however, can only result from alterations of the [HCO,]/Pco, ratio. Blood [HCOJ could be regulated by ion exchange in the kidneys or by an exchange between the intra- and extracellular compartments (71). Several studies on reptiles report that plasma [HCO,] does not change with temperature, however, in which case the decrease in pH requires that the arterial PCO~ (Pace,) increase with temperature (78). The alveolar Pco2 (PA& and Pa coZvalues depend on the ratio of VA to pulmonary ko2 (cf. 83). This is expressed in the relationship iTA/bC02 = RT/PA~~~, where R is the gas constant and T is absolute temperature. The ~~~~~ value rises if iTA/h02 decreases. As we have said, the VE value is usually measured instead of VA. Furthermore To2 is more frequently measured than VCO, and the effects of temperature on the ACRs are reported. Measurements of acid-base status in freshwater turtles (Pseudemys sp.) are thought to support the concepts of constant relative alkalinity and alphastat regulation (86, 88, 98). Plasma [HCOJ is not influenced by temperature changes, whereas Paco2increases with temperature owing to the decrease in the ACRs. For example, when the body temperature of Pseudemys scripta increased from 10 to 3O”C, vo2 increased (QIO = 1.8), whereas v did not change significantly. As a result, Pa coZ increased with 14.1 Torr to 31.9 Torr, while pH, decreased from 7.76 to 7.56 (88). The ACRs also decrease with temperature in the lizards Lace&a sp. (118) and Iguana iguana (56, 160), in the snake Thamnophis elegans (74), and in Alligator mississippienCVCO~ l the Henderson-Hasselbalch Pco~), where CYCO~ is the solubility sis (40). At high body temperatures, an elevated ~~~~~ may lead to the disad- Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 water plus a constant (C), or pH, = pNW + C. Differentiating with respect to temperature gives dpH,/dT = dpH,/dT N -0.018. Reeves (131) suggested that constant arterial OH-/H+ reflects regulation of net protein charge. This regulation requires pH changes with temperature that are similar but not identical to those for constant OH-/H+ (132). The theory is based on a closed-system model in which CO, content is assumed to be constant, and it predicts that constant net protein charge is preserved in all body compartments (108,131). Reeves (131) emphasized that constant net protein charge optimizes enzyme activity when body temperature changes. This theoretical prediction has recently received some experimental support (70, 161). Constant net protein charge implies constant fractional dissociation of imidazole groups of proteins (constant cw-imidazole). Reeves therefore used the term a@hns~at regulation for his model of vertebrate acidbase control. 248 M. L. GLASS AND S. C. WOOD Volume 63 vantage that PA o2 is low. This is clear from the relationship PACE,N PIED, - PA~,/RE, where PI,, is the partial pressure of inspired O2 (cf. 148). In the turtle Pseudemys jbridana the pulmonary venous O2 content is decreased at high body temperatures because low PACTvalues prevent saturation of the pulmonary capillary blood (98). C. Exceptions to Alphastat Relative Alkalinity Regulation and Constant Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 The time courses of regulatory responses of acid-base regulation were largely neglected until 1976, when Jackson and Kagen (86) found that adjustments of ACRs and pH to temperature changes in Pseudemys scripta were immediate and similar to those reported earlier. However, the longterm or seasonal effects of temperature on gas exchange, ventilation, and acid-base balance have been studied only recently and appear to be different. During winter the body temperature of the box turtle Terrapene ornata is frequently 5OC or less (103). At 5OC the breathing frequency is 3 breaths/ h as a mean value for a 24-h period, but overnight breath holds do occur (58). The ACRs of Terrapene increase with temperatures below 15”C, which is the minimum temperature for activation of hibernating box turtles (103). At temperatures greater than 15OC the ACRs decrease with increased body temperature. Figure 2 shows the effects of temperature on ACRs for several turtle species. The comparison emphasizes that the reported temperature effects are highly divergent at lower temperatures, whereas the results are consistent at ZO-35°C. It is presently not clear whether the diversities of responses at low temperatures are species-specific or are due to differences in exposure time to cold. Long-term measurements are frequently required for a full evaluation of the regulatory responses of reptiles; this is especially true at low body temperatures. In some lizards the relationships between body temperature and pH, are different from those predicted for constant OH-/H+ and alphastat regulation. The deviations occur within temperature ranges that include preferred body temperatures (14, 153, 154, 157). Wood et al. (154) measured ventilation, gas exchange, and blood acid-base status in Varanus exanthematicus. Measurements were performed at night on resting undisturbed animals. In Varanus the pH, values and ACRs did not change between 25 and 35OC. Thus ventilation increases in proportion to vo2, and PaoZ is high (93.5 Torr) at 35OC. Wood et al. (153) argued that this is an advantage to an active predatory lizard with high preferred body temperature (37°C). Davies (40) suggested that conflicting results on acid-base balance are due to errors in the experiments on lizards. Wood and colleagues (157), however, have recently repeated measurements on Varanus, but these experiments were based on heating and cooling during the daytime. Over the range January 1983 RESPIRATION IN . Pseudemys floridana A Chrysemys picta bellii l Chelonia mydas 80 20 0 I 0 I 10 1 1 1 20 Temperature I I 30 1“c) FIG. 2. Relationships between air convection requirements (ACRs) and for various turtle species. Note that results are consistent at higher body highly divergent at lower temperatures. [Data from: n Jackson et al. (88); v A M. L. Glass, R. G. Boutilier, and N. Heisler, manuscript in preparation; l o Glass et al. (58).] body temperature temperatures but Kinney et al. (98); Jackson et al. (87); of 15-38OC the pH, value decreased with temperature but only by 0.005 units/ OC. This change is much too small to be consistent with constant relative alkalinity or alphastat regulation. Such results indicate that current concepts on temperature-dependent acid-base regulation need some revision. Moreover note that some studies may seem to support constant relative alkalinity but may also be interpreted differently. Thus in the turtle Pseudemys scripta the ApH/AT value for arterial blood was -0.010 units/“C between 10 and 30°C (88), whereas the predicted change for constant relative alkalinity is a ApH/AT value of -0.018 units/“C (78). Presently no explanation is available for these deviations from current theories on temperature-dependent acid-base regulation in reptiles. We feel that the development of consistent models will require further experiments involving exposure to temperatures consistent with the ecology of the animals. A recent study by Ackermann and White (2) emphasizes this point. In the marine iguana AmbZgrhynchus &status, pH, decreases by 0.015 units/ OC when body temperature increases from 24 to 35OC. At temperatures below 24”C, however, pH, changes little with temperature. Temperatures between 24 and 35OC appear to be consistent with the ecology of the species. Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 a 249 REPTILES 250 V. CONTROL M. L. GLASS OF BREATHING A. Ventilator9 I AT AND CONSTANT S. C. WOOD BODY TEMPERATURE Responses to Carbon Dioxide Inhalation Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Reptiles at a given body temperature regulate acid-base status by adjustments of VA relative to pulmonary ko2, i.e., ~A/~co~ = RT/PA,,, (cf. 43). The gain of this regulatory system is indicated by the ventilatory responses to imposed increases in Pacoz and ~~~~~ caused by CO2 inhalation. Effects of increased PCO~ on VA are difficult to measure, and VE or ACRs are normally studied. The ventilatory responses to COZ inhalation are diverse among reptiles. Chelonians and crocodilians increase VT and overall f values in response to elevated inspired Pu)~ (PI& (41,52,57,87,88). This may produce pronounced increases in VIX or ACR. In I?ww&w~~~~sscriJ)tcx,,for example, the ACRs increased U-fold in response to inhalation of 6% CO2 (88). Such positive ventilatory responses to COZ inhalation are also typical of birds (cf. 21) and mammals (cf. 97) and are easily explained from a functional point of viewi.e., increased ventilation promotes CO2 elimination and opposes the rise of ~~~~~ and Pace,. In aquatic reptiles, CO2 may also cause reductions in the duration of breath holds. In Pseudemys the ratio VP/(VP + NVP) increases when CO2 is inspired (52). Dive duration in the turtle Pebmedusa subrufa is reduced when ~~~~~ increases (57). Likewise the turtle Chrgsemys picta responds to COZ inhalation by increasing f through a shortening of breath holds (113). In Crocodylus niZoticus a shortening of breath holds is the only mechanism that produces a positive frequency response to hypercapnia (60). Most studies on the control of breathing in snakes and lizards report paradoxical responses to inspiration of C02. The value of VT increases, but f is depressed (59, 66, 118, 122, 146). In Lace&a viridis the depression of f leads to a decrease in ventilation relative to base-line values preceding COZ inhalation (118). This response seems meaningless because a decrease in ventilation diminishes the clearance of metabolically produced COB, thereby increasing the concentrations and PCO~ values in tissues. Therefore the responses of lizards and some snakes to CO2 inhalation possibly are different from those produced by a metabolic elevation in Pacoz and PACES. Recently Glass and co-workers (63) reported a positive frequency response to CO2 inhalation for the lizard Varanus exanthematicus. In Varanus both f and VT increased markedly during COZ inhalation, leading to a sevenfold increase in ventilation with an increase in PACT, of 15 Torr. The relationship between PA coZand ventilation is highly reproducible in Varanus in contrast to the relationships between the subparameters (VT and f) and PACES.It is difficult to predict whether the ventilatory response results from increased for from enlargement of VT. This is consistent with data on turtles, indicating that VT and f are independently regulated (cf. 84), a condition that resembles the situation in birds (cf. 21). January 1983 RESPIRATION IN 251 REPTILES The ventilatory responses of reptiles to COZ inhalation may be very fast (55, 57, 84). In the tegu lizard Tupinambis nigropunctatus a decrease in intrapulmonary PCO~causes breath holding within 0.8-1.4 s (55). This indicates a peripheral location of the receptor system, i.e., the receptors must be located close to the alveolar gas compartment (see sect. vC). B. Anoxia and Hypoxia: Tolerance and Regulatory Responses l Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Reptiles, (especially turtles) are very tolerant of anoxia. At their normal active body temperature, chelonians may survive for hours or days without O2 (7,18,80,94,133). Reptiles share this amazing tolerance with amphibians (104), whereas anoxia is deadly within minutes to endotherms at their normal body temperatures (7). The anoxia tolerance of ectotherms is due in part to low metabolic needs leading to a slow depletion of resources for anaerobic metabolism. Decreased metabolism in hypothermic mammals may also permit survival for hours without O2 (19). When reptiles are at body temperatures close to the freezing point, anaerobic glycolysis may provide sufficient energy for several months (147). The lack of O2 causes drastic changes in the animal’s acid-base status. Clark and Miller (31) measured a decrease in brain pH from 7.2 to 6.4 in Pseudemys scripta after the animal had been anoxic for 15 h at 22OC. Longterm survival of Chrysemys picta beZZii at 3OC was associated with the increase of plasma lactate to values exceeding 200 mmol liter-’ (147). As we have said, depletion of aerobic reserves (mainly intrapulmonary 02) is very slow during breath holds in reptiles. Therefore it is relevant to ask whether the tolerance to anoxia becomes important for a reptile in its normal habitat. Especially interesting in that context is the study by Ultsch and Jackson (147) on the freshwater turtle Chrysemys pi&a bellii, which is unusually tolerant of anoxia and low temperatures (142). Ultsch and Jackson correlate this tolerance with the northern distribution of this turtle, which inhabits areas where lakes are covered with ice for prolonged periods. Moberly (116) described another example of the survival value of anoxic tolerance. The common iguana (Ig&ana iguana) will attempt to escape into water when threatened by predators and may stay submerged for more than 4 h. The high blood lactate levels at the end of such dives indicate a strong dependence on anaerobic metabolism. In spite of this tolerance of anoxia, reptiles normally regulate ventilation to avoid a lack of oxygen. The values of PACT and Paoz depend on ventilation and VO, as expressed by a relationship that for RE = 1 becomes PACT = PI - RT(~o&A) (43, 128). Consequently ~~~~ rises when TE and VA 02 increase relative to TO,. Chelonians increase ventilation in response to large decreases in Pa 02, i.e., hypoxia (5, 10, 22, 26, 52, 57, 82). In aquatic species the ventilatory response to hypoxia may include a decrease in NVP (cf. 159) and 252 M. L. GLASS AND S. C. WOOD Vokurri~c? 63 an increase in the ratio VP/(VP + NVP), i.e., duration of ventilatory periods relative to total time (52). Hypoxia also causes cardiovascular changes. The responses involve an increase in heart rate and an increase in pulmonary perfusion (22, 26). The combination of ventilatory and cardiovascular responses is effective in preserving normal vo2. In Chelydra seripentina the 00~ value remained constant when the ambient O2 concentration decreased from 21to 2% (22). Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 The ventilatory responses to hypoxia have been studied in only a few nonchelonian reptiles. Nielsen (119) reported that ventilation is depressed by hypoxia in the lizard Lacerta. In contrast, ventilation increased with decreased Pro, in Varanus exanthematicus (63). Snakes increase ventilation in response to hypoxia (59, 66). For the aquatic snake Acrochordus, Glass and Johansen (59) reported a positive ventilatory response to hypoxia caused by a reduction in NVP. Data on the ventilatory responses of crocodilians to hypoxia are scarce and largely qualitative, but CrocodyZus niZoticus increased ventilation mainly by shortening breath holds (60). In reptiles the relationship between ventilation and PI,, usually resembles a hyperbolic function, i.e., the effect of hypoxia on ventilation is small unless inspired PIN, is considerably reduced. This resembles the mammalian ventilatory response to hypoxia, which has been studied extensively (cf. 97). In mammals a consequence of the shape of the response curve is that the role of a Paz-mediated drive to ventilation is relatively small unless PIN, is considerably reduced, as at high altitude (cf. 148). In reptiles a hypoxic stimulus may not be important unless Pa o2 is very low. For example, ventilation in Acrochordus changes little if ambient PO, is decreased from 155 to 74 Torr, whereas ventilation doubles with decreases to 37 Torr (59). A Po2-mediated ventilatory drive may still be important in normal respiratory control because PACT and Paoz values may fall to low values during breath holding, especially in diving reptiles (1, 59, 155). The conventional method for measuring ventilatory responses to hypoxia requires that a steady state be achieved, i.e., ventilation must reach a steady value at each level of hypoxia. This causes various si de effects that may mask a response (43). An alternative approach is to measure transient ventilatory responses after a single inhalation of pure OZ. Bench .etrit et al. used this technique to study ventilatory chemoreflexes in the tortoise Testudo horsJie+!di. Inhalation of pure O2 depresses or arrests ventilation within seconds. Consequently a tonic Po2-dependent stimulus to breathing is present during normoxic conditions. Techniques less refined than the single-breath method confirm that O2 breathing depresses ventilation in reptiles (52, 57, 59, 63). At low body temperatures the ventilatory responses of chelonians to a given concentration of inspired O2 are much reduced relative to responses at higher body temperatures (10, 82). This effect may depend on several factors: Demands for O2 are smallest at low temperature, and therefore a relatively smaller amount of O2 is extracted from the lung when tempera- January 1983 RESPIRATION IN 253 REPTILES 300 C‘c.3 - 80 -i E. 0, x 200 ci> L t- m 2 .-E 100 5 .-E 9 0 0 20 40 PO, 60 80 (To4 FIG. 3. Ventilatory responses to hypoxia in the freshwater turtle Chrysemys picta be&%. Pulmonary ventilation is shown as function of arterial PO,. Highest value corresponds to normoxie conditions, i.e., inspiration of room air, whereas lower values result from inspiration of hypoxic gases. Relationships are depicted for 3 body temperatures. Note pronounced temperature dependence of ventilatory responses. Increased temperature causes shift to the right of the hemoglobin 02-binding curve, thus impairing O2 transport at low arterial PO,. (From M. L. Glass, R. G. Boutilier, and N. Heisler, manuscript in preparation.) Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 tures are low (82). Moreover the O2 affinity of the blood increases with lowering of the body temperature, thereby facilitating O2 transport at low PACT and Pao, values (cf. 124). Consequently the effects of a given decrease in PI,, will be least severe at low temperatures. As Wood and Lenfant (159) have shown, however, a full evaluation of such relationships requires that the blood gas picture be known. Recently ventilatory responses to hypoxia were measured in the freshwater turtle Chrysemys picta be&i along with and pH,. Furthermore the VO, and 02-binding curves were depao,, pace,, termined (M. L. Glass, R. G. Boutilier, and N. Heisler, manuscript in preparation). The study emphasizes the temperature-dependence of the ventilatory responses to hypoxia. Thus at IOOC ventilation did not increase until Pao, fell to 5 Torr. In contrast, at 30°C ventilation approximately doubled as Pao, decreased from 60 to 30 Torr (Fig. 3). These responses correlate with the ability of the turtle to maintain normal To2 despite a decrease in Pao,. 254 M. L. GLASS AND S. C. WOOD Vohme 63 C. Receptor Sys t erms Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Investigators have studied several receptor systems in relation to the chemical control of breathing in reptiles, but only a few species have been examined. Therefore it is difficult to generalize concerning such receptor systems. Moreover some data suggest the systems may be different for various groups of reptiles. The most extensive data on PO, receptors are available for the tortoise Testudo horsfieZdi (10). In Testudo bilateral vagotomy abolishes ventilatory responses to hypoxia and hyperoxia, which suggests that Po2-sensitive receptor systems are innervated by the vagus nerves and provide a tonic drive to breathing. Some evidence indicates that the receptors are screening pulmonary arterial blood, but their exact location is not known (10). In mammals, Po2-sensitive receptors are located at the bifurcations of the external and internal carotids (32, 73), whereas the carotid bodies are situated at the common cdrotid in birds (150). A careful anatomical study (3) failed to demonstrate carotid bodies in reptiles. However, Frankel et al. (52) claim an ovoid structure is situated at the bifurcation of the carotid arteries in Pseudemys, but they have not provided evidence for the functional role of this structure. Although only a few species have been studied, the available information on CO2 and pH receptors suggests interesting perspectives. The presence of intrapulmonary CO2 receptors in lizards is well established (50), and Hitzig and Jackson (76) have discovered central nervous pH receptors in turtles. Moreover there is some evidence for peripheral COZ receptors in the tortoise Testudo horsfieldi (11). The presence of intrapulmonary COZ receptors in the lizard Tupinambis teguixin was first proposed by Gatz et al. (55), who measured an immediate decrease in ventilation with abrupt lowering of PACT,. An intrapulmonary location for the receptors for this response was strongly indicated because vascular isolation of the lung did not abolish the ventilatory response. Later studies confirmed that CO2 receptors cover large pulmonary areas in Tupinambis (136) and that output of these receptors increased with decreases in PACT, (50). The CO2 receptors are insensitive to stretch of the lung in contrast to pulmonary mechanoreceptors in Tupinambis. These mechanoreceptors may also be sensitive to changes in PACES,but they are not as sensitive as the CO2 receptors (50). Carbon dioxide-sensitive mechanoreceptors are also located in turtle lungs but seem to be of limited importance in ventilatory responses to CO2 inhalation (95, 111, 112). In Chqysemys picta the mechanoreceptors are involved in the regulation of VT, whereas the response to CO2 inhalation primarily consists of a shortening of the NVPs (95, 113). The intrapulmonary CO2 receptors of Tupinambis have not been reported for other reptiles. Intrapulmonary COZ receptors are well established in birds, however (21, 137). The avian and lacertilian intrapulmonary CO2 receptors may be functionally similar, whereas the mechanoreceptors in turtle January 1983 RESPIRATION IN REPTILES 255 VI. CONCLUSIONS Substantial progress has occurred over the past decade in understanding respiration and its control in reptiles. Information has accumulated concerning values for variables in respiratory function. A large difference in O2 requirements between reptiles and endothermic vertebrates is clearly reflected on all levels of aerobic metabolism. The lower O2 requirements of reptiles imply that values for pulmonary ventilation and perfusion are also low by mammalian standards and that continuous ventilation of the lungs is not required to supply sufficient Oz. Instead breathing activity is interrupted by breath-hold periods. Correspondingly pulmonary perfusion and gas exchange are periodic, with peak perfusion occurring during breathing activity. The cyclic character of ventilation and pulmonary perfusion implies Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 lungs belong to a type widespread among vertebrates with pulmonary ventilation (49). Central nervous receptors sensitive to the acid-base status of the cerebrospinal fluid (CSF) have been extensively studied in mammals (cf. 97) but were unknown in ectothermic vertebrates until recently, when such receptors were reported in Pseudemys scripta (75, 76). Subnormal pH values of the CSF in Pseudemys cause a pronounced increase in ACRs (76). Significantly the magnitude of this response was independent of body temperature (76). The information from this specifically pH-sensitive receptor system overrides the peripheral receptor’s input to the central nervous system (76). The ventilatory response to the decreased pH of the CSF occurs by an elevated f value. Values for VT, however, also increase during CO2 inhalation. This suggests that VT and f in Pseudemys are regulated by independent receptor systems (75). The studies on central receptors in Pseudemys provide important keys to the relative roles of receptors, but more studies are needed to quantify their relative importance and in particular to relate receptor function to the normal breathing patterns of the animals. Furthermore (as is often the case in reptilian physiology) too few species have been too little studied. An important question must still be answered: Do reptiles possess a receptor system that functions in relation to an increase of ventilation with temperature? Reeves (131) proposed that such receptors could be sensitive to the net protein charge of the blood, which should remain constant despite variations in body temperature. A recent study by Davies et al. (41) reports that the alligator defends net protein charge at all body temperatures. The increase in ACRs for a given change in net protein charge of arterial blood proved to be independent of body temperature. This may provide an important key, but not all reptiles regulate net protein charge when body temperature changes; furthermore the alphastat receptors have not yet been located. 256 M. L. GLASS AND S. C. WOOD Volume 63 We thank Professor Kjell Johansen for useful comments on several versions of the manuscript. We are also grateful for comments by Dr. D. C. Jackson and Dr. D. P. Toews. We also acknowledge assistance from Kirsten Horup in preparing the manuscript. M. L. Glass was the recipient of a fellowship from the Alexander von Humboldt-Stiftung during much of the time needed to prepare this review. REFERENCES 1. ACKERMANN, R. A., AND F. N. WHITE. Cyclic carbon dioxide exchange in the turtle Pseudemys scripta Physiol. Zool. 52: 378-389, 1979. 2. ACKERMANN, R. A., AND F. N. WHITE. The effects of temperature and acid-base balance and ventilation of the marine iguana. Respir. Physiol 39: 133-147, 1980. 3. ADAMS, W. E. The Comparative Morphology of the Carotid Body and Carotid Sinus. Springfield, IL: Thomas, 1958. 4. ADARO, F., P. SCHEID, J. TEICHMANN, AND J. PIIPER. A rebreathing method for estimating pulmonary Ds: theory and measurements in dog lungs. Respir. Physiol. 18: 43-63, 1973. 5. ALTLAND, P. D. AND M. PARKER. Effects of hypoxia upon the box turtle. Am J Physiol. 180: 421-427, 1955. 6. ANDERSEN, H. T. Physiological adjustments to prolonged diving in the American alligator, Alligator mississippiensis. Acta Physiol Scund 53: 23-45, 1961. 7. BELKIN, D. A. Anoxia: tolerance in reptiles. Science 139: 492-493, 1963. 8. BELKIN, D. A. Variations in heart rate during voluntary diving in the turtle Pseudemys concinna Copeia 1964: 321-336, 1964. 9. BELKIN, D. A. Aquatic respiration and underwater survival of two freshwater turtle species. Respir. Physid 4: 1-14, 1968. 10. BENCHETRIT, G., J. ARMAND, AND P. DEJOURS. Ventilatory chemoreflex drive in the tortoise Testudo horsfieldi. Respir. Physiol 31: 183-191, 1977. 11. BENCHETRIT, G., AND P. DEJOURS. Ventilatory COa drive in the tortoise Testudo hors&e,!.& J. Exp. Biol 87: 229-236, 1980. 12. BENEDICT, F. G. The Physiology of Large Reptiles. Washington, DC: Carnegie Inst., Publ. 425, 1932. 13. BENNETT, A. F. A comparison of activities of metabolic enzymes in lizards and rats. Comp. Biochxm. Physiol B 42: 637-647, 1972. 14. BENNETT, A. F. Ventilation in two species of lizards during rest and activity. Comp. Biochem Physiol. A 42: 653-671, 1973. 15. BENNETT, A. F., AND W. R. DAWSON. Aerobic and anaerobic metabolism during activity in the lizard, Dips+ saurus dorsalis. J. Camp. Physiol. 81: 289-299, 1972. 16. BENNETT, A. F., AND W. R. DAWSON. Metabolism. In: Bio&y of the Reptilia. Physiology A, edited by C. Gans Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 that measurements must be performed over prolonged periods. Prolonged measurements are also needed to assess the metabolic status of the animal. Moreover data should be obtained at several body temperatures, because variables such as pulmonary diffusing capacity, perfusion, and ventilation all increase along with O2 uptake when body temperature increases. Interspecies comparisons are thus impossible unless the temperature dependence of the compared variables is known. Reptiles are different from mammals or birds in being highly tolerant of a lack of 0,; in addition, reptiles tolerate variations in acid-base status that would be fatal to endotherms. In spite of this ability, reptiles possess regulatory systems to monitor acid-base status and blood oxygenation. Recently several receptor systems have been reported in relation to these regulatory responses, but the relative roles of these systems are incompletely known. These include intrapulmonary CO2 receptors, PO, receptors, and central nervous pH receptors. During the past decade several studies have addressed general theories concerning effects of changes in body temperature on acid-base regulation in ectotherms. Studies on acid-base regulation in alligators and turtles agree to some extent with predictions of these theories, whereas data on some lizards do not conform. Therefore it seems that further experimental work is needed to evaluate and modify current theories. January 17. 18. 19. 20. 22. 23. 24, 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. 36. RESPIRATION and W. R. Dawson. New York: Academic, 1976, vol. 5, p. 127-224. BENTLY, P. J. Osmoregulation. In: BioZogy of the Rep tibia. PhysioZogy A, edited by C. Gans and W. R. Dawson. New York: Academic, 1976, vol. 5, p. 365-413. BERKSON, H. Physiological adjustments to prolonged diving in the Pacific green turtle (Chelonia mydus ugas&ii). Comp. B&hem, PhysioL 18: 101-119, 1966. BIeRCK, G., B. JOHANSSON, AND H. SCHMID. Reactions of hedgehogs, hibernating and non-hibernating, to inhalation of oxygen, carbon dioxide and nitrogen. Actu PhysioL Scand 37: 71-83, 1956. BOHR, C. uber die spezifische Titigkeit der Lungen bei der respiratorischen Gasaufnahme und ihr Verhalten zu der durch die Alveolenwand stattfindenden Gasdiffusion. Skand Arch. Physiol 22: 221-280, 1909. BOUVEROT, P. Control of breathing in birds compared with mammals. Physiol Rev. 58: 604-655, 1978. BOYER, D. R. Hypoxia: effects on heart rate and respiration in the snapping turtle. Science 140: 813-814,1963. BROWN, C. R., AND J. P. LOVERIDGE. The effect of temperature on oxygen consumption and evaporative water loss in Crocodylus n&.&us. &nap. Biochem PhysioL A 69: 51-57, 1981. BURGGREN, W. W. A quantitative analysis of ventilation tachycardia and its control on two chelonians, Pseudemys scripta and Testudo graeca J Exp. BioL 63: 367-380,1975. BURGGREN, W. W. The pulmonary circulation of the chelonian reptile: morphology, haemodynamics and pharmacology. J. Comp. PhysioL 116: 303-323, 1977. BURGGREN, W. W., M. L. GLASS, AND K. JOHANSEN. Pulmonary ventilation: perfusion relationships in terrestrial and aquatic chelonian reptiles. Can, J. Zod 55: 2024-2034,1977. BURGGREN, W. W., M. L. GLASS, AND K. JOHANSEN. Intrapulmonary variation of gas partial pressures and ventilation inequalities in chelonian reptiles. J. Comp. Physiol 126: 203-209, 1978. BURGGREN, W. W., AND G. SHELTON. Gas exchange and transport during intermittent breathing in chelonian reptiles. J. Exp. BioL 82: 75-92, 1979. CARR, A. Hundbook of Turtles. Ithaca, NY: Cornell Univ. Press, 1952. CLARK, B. D., C. GANS, AND H. I. ROSENBERG. Airflow in snake ventilation. Respir. PhysioL 32: 207-212, 1978. CLARK, V. M., AND A. T. MILLER, JR. Studies on anaerobic metabolism in the fresh-water turtle (Pseudemys scripta elegans). Camp. B&hem, PhysioL A 44: 55-62, \ 1973. COMROE, J. H., JR. The peripheral chemoreceptors. In: Handbook of Physiology. Respiration, edited by W. 0. Fenn and H. Rahn. Washington, DC: Am. Physiol. Sot., 1964, sect. 3, vol. 1, chapt. 23, p. 557-583. COULSON, R. A., AND J. D. HERBERT. Relationship between metabolic rate and various physiological and biochemical parameters. A comparison of alligator, man and shrew. Comp. B&hem, PhysioL A 69: 1-13, 1981. COULSON, R. A., AND T. HERNANDEZ. Biochemistrl/ of the Alligator. A Study of Metabolism in Slow Motion. Baton Rouge, LA: Louisianna State Univ. Press, 1964. COULSON, R. A., AND T. HERNANDEZ. Oxygen debt in reptiles: relationship between the time required for repayment and metabolic rate. Comp. B&hem PhysioL A 65: 453-457, 1980. CRAGG, P. A. Ventilatory patterns and variables in rest IN 37. 38. 39. 40. 41 42 43 44 45 46 47 48. 49. 50. 51. 52. 53. 54. 55. REPTILES 257 and activity in the lizard Lacerta, Comp. B&hem. PhysioL A 60: 399-411, 1978. CRAWFORD, E. C., JR., R. N. GATZ, H. MAGNUSSEN, S. F. PERRY, AND J. PIIPER. Lung volumes, pulmonary blood flow and carbon monoxide diffusing capacity of turtles. J Comp. PhysioL 107: 169-178, 1976. CRAWFORD, E. C., JR., AND R. R. SCHULTETUS. CUtaneous gas exchange in the lizard Suuromulus obesus. Cope,iu 1970: 179-180,197O. DANTZLER, W. H. Renal function (with special emphasis on nitrogen excretion). In: BioZogy of the Reptilicz. Physiobgy A, edited by C. Gans and W. R. Dawson. New York: Academic, 1976, vol. 5, p. 447-504. DAVIES, D. G. Temperature-induced changes in acidbase status in the alligator, A&g&or mi.ssissippien&. J. AppL PhysioL: Reqwirat. Environ. Exercise PhysioL 45: 922-926, 1978. DAVIES, D. G., J. L. THOMAS, AND E. N. SMITH. Effect of body temperature on ventilatory control in the alligator. J. AppL PhysioL: Respirat. Environ Exercbe Physiol 52: 114-118, 1982. DAVIES, P. M. C., AND E. L. BENNETT. Non-acclimatory latitude-dependent metabolic adaptation to temperature in juvenile natricine snakes. J. Comp. PhysioL 142: 489494,198l. DEJOURS, P. Principle-s of Comparative Respiratory PhysioZogy. New York: Elsevier, 1975. DESSAUER, H. C. Blood chemistry of reptiles: physiological and evolutionary aspects. In: Biobgy of the Rep tiliu, edited by C. Gans and T. S. Parsons. New York: Academic, 1970, vol. 3, p. l-54. DILL, D. B., AND H. T. EDWARDS. Respiration and metabolism in a young crocodile (Crocodylus acutus Cuvier). Copeia 1931: l-3, 1931. DUNCKER, H.-R. General morphological principles of amniotic lungs. In: Respiratory Function in Birds, Adult and Embyonic, edited by J. Piiper. Berlin: Springer-Verlag, 1978, p. 2-15. DUNSON, W. A. Aquatic respiration in Trionyx spin+r aqwr. Herpetologica 16: 277-283, 1960. ELSE, P. L., AND A. J. HULBERT. Comparison of the “mammal machine” and the “reptile machine”: energy production. Am J PhysioL 240 (Regulatory Integrative Comp. PhysioL 9): R3-R9, 1981. FEDDE, M. R., AND W. D. KUHLMANN. Intrapulmonary carbon dioxide-sensitive receptors: amphibians to mammals: In: Respiratory Function in Birds, Adult and Emhonic, edited by J. Piiper. Berlin: Springer-Verlag, 1978, p. 33-50. FEDDE, M. R., W. D. KUHLMANN, AND P. SCHEID. Intrapulmonary receptors in the Tegu lizard: I. Sensitivity to COz. Respir. PhysioL 29: 35-48,1977. FORSTER, R. E., AND E. D. CRANDALL. Pulmonary gas exchange. Annu. Rev. PhysioL 38: 69-95, 1976. FRANKEL, H. N., A. SPITZER, J. BLAINE, AND E. P. SCHOENER. Respiratory responses of turtles (Pseudemys scripta) to changes in arterial gas composition. Comp. Biochem PhysioL 31: 535-546, 1969. GANS, C., AND B. D. CLARK. Air flow in reptilian ventilation. Camp. Biocti PhysioL A 60: 453-459,1978. GATTEN, R. E., JR. Effects of temperature and activity on aerobic and anaerobic metabolism and heart rate in the turtles Pseudemys scriptu and Terrapene orruzta Camp. Biochem PhysioL A 48: 619-648, 1974. GATZ, R. N., M. R. FEDDE, AND E. C. CRAWFORD, JR. Lizard lungs: COz-sensitive receptors in Tupinumbis nigropunctatus. Experientia 31: 455-456, 1975. Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 21. 1983 258 M. L. GLASS 71. HEISLER, N. Bicarbonate exchanges between body compartments after changes of temperature in the larger spotted dogfish (Scyliorhinus stellaris). Respir. PhysioL 33: 145-X0,1978. A. M. Energy metabolism as related to 72. HEMMINGSEN, body size and respiratory surfaces and its evolution. Rep. Steno. Mem Hosp. Nurd Insulinlub. 9: 7-110, 1960. HEYMANS, C., J.-J. BOUCKAERT, AND L. DAUTREBANDE. Sinus carotidien et reflexes respiratoires. II. Influences respiratoires reflexes de l’acidose, de l’alcalose, de l’anhydride carbonique, de l’ion hydrogene et de l’anoxemie. Sinus carotidien et echanges respiratoires dans les poumons et au dela des poumons. Arch. Ink Phar??Lac&yn Ther. 39: 400-450, 1930. HICKS, J. W., M. L. RIEDESEL, AND M. L. GLASS. Ventilation and gas exchange in Thumwphis elegans after S. C. WOOD 75. 76. 77. 78. 79. 80. 81. 82. 83. 84. 85. 86 87 88. 89. 90. 91. 92. Vot!um 63 acute and chronic exposure to various thermal environments (abstr.). J. Co& Wgo. Acud Ski 2: 99-100, 1979. HITZIG, B. M. Control of ventilation by central respiratory chemosensors in the unanesthetized turtle, Pseudemys scripta ekgans (PhD Thesis). Providence, RI: Brown Univ., 1977. HITZIG, B. M., AND D. C. JACKSON. Central chemical control of ventilation in the unanesthetized turtle. Am J. PhysioL 235 (Regulatory Integrative Comp. PhysioL 4): R257-R264, 1978. HOWELL, B. J., F. W. BAUMGARDNER, K. BONDI, AND H. RAHN. Acid-base balance in cold-blooded vertebrates as a function of body temperature. Am J. PhysioL 218: 600-606, 1970. HOWELL, B. J., AND H. RAHN. Regulation of acid-base balance in reptiles. In: Biology of the Reptilia PhysioZogy A, edited by C. Gans and W. R. Dawson. New York: Academic, 1976, vol. 5, p. 335-364. HUGHES, G. M., R. GAYMER, M. MOORE, AND A. J. WOAKES. Respiratory exchange and body size in the Aldabra giant tortoise. J. Exp. Bid 55: 651-665, 1971. JACKSON, D. C. Metabolic depression and oxygen depletion in the diving turtle. J. AppL PhysioL 24: 503-509, 1968. JACKSON, D. C. The effect of temperature on ventilation in the turtle Pseudemys scripta ekgans. Respir. PhysioL 12: 131-140,197l. JACKSON, D. C. Ventilatory response to hypoxia in turtles at various temperatures. Respir. PhysioL 18: 178-187, 1973. JACKSON, D. C. Respiratory control and CO2 eonductance: temperature effects in a turtle and a frog. Respir. PhysioL 33: 103-114, 1978. JACKSON, D. C. Respiratory control in air-breathing ectotherms. In: Research Topics in Physiology. Regulation of Ventilation and Gas Exchange, edited by D. G. Davies and C. B. Barnes. New York: Academic, 1978, vol. I, p. 93-130. JACKSON, D. C., J. ALLEN, AND P. K. STRUPP. The contribution of non-pulmonary surfaces to CO2 loss in 6 species of turtles at 20°C. Camp. Biochem PhysioL A 55: 243-246, 1976. JACKSON, D. C., AND R. D. KAGEN. Effects of temperature transients on gas exchange and acid-base status of turtles. Am J PhysioL 230: 1389-1393, 1976. JACKSON, D. C., D. R. KRAUS, AND H. D. PRANGE. Ventilatory response to inspired COz in the sea turtle: effects of body size and temperature. Respir. PhysioL 38: 71-81, 1979. JACKSON, D. C., S. E. PALMER, AND W. L. MEADOW. The effects of temperature and carbon dioxide breathing on ventilation and acid-base status of turtles. Respir. PhysioL 20: 131-146, 1974. JACKSON, D. C., AND H. D. PRANGE. Ventilation and gas exchange during rest and exercise in adult green sea turtles. J. Comp. PhysioL 134: 315-319, 1979. JACOBSON, E. R., AND W. G. WHITFORD. The effect of acclimation on physiological responses to temperature in the snakes, Thumnophis proximus and Natrix rhom&j&a Comp. Biochem, PhysioL 35: 439-449,197O. JAMMES, Y., AND C. GRIMAUD. Ventilation, pulmonary and cutaneous gas exchange in the awake lizard h& viridis. Camp. Biochem PhysioL A 55: 279-285, 1976. JOHANSEN, K., W. W. BURGGREN, AND M. L. GLASS. Pulmonary stretch receptors regulate heart rate and pulmonary blood flow in the turtle Pseudemys scripta Camp. B&&em PhysioL A 58: 185-191, 1977. Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 56. GIORDANO, R. V., AND D. C. JACKSON. The effect of temperature on ventilation in the green iguana. Comp. Biochem PhysioL 45: 235-238,1973. 57. GLASS, M. L., W. W. BURGGREN, AND K. JOHANSEN. Ventilation in an aquatic and in a terrestrial chelonian reptile. J. Exp. BioL 72: 165-179, 1978. 58. GLASS, M. L., J. W. HICKS, AND M. L. RIEDESEL. Respiratory responses to long-term temperature exposure in the box turtle, Terrapene ornuta J. Comp. PhysioL 131: 353-359, 1979. 59. GLASS, M. L., AND K. JOHANSEN. Control of breathing in Acrochordw javanicus, an aquatic snake. PhysioL ZooL 49: 328-340, 1976. 60. GLASS, M. L., AND K. JOHANSEN. Periodic breathing in the crocodile CrocodyZus niloticus. J. Exp. ZooL 208: 319-326,1979. Pulmonary oxygen 61. GLASS, M. L., AND K. JOHANSEN. diffusing capacity of the lizard Tupinumbis teguixin. J. Exp. ZooL 219: 385-388,1982. 62. GLASS, M. L., K. JOHANSEN, AND A. S. ABE. Pulmonary diffusing capacity in reptiles (relations to temperature and Oz-uptake). J. Camp. PhysioL 142: 509-514,198l. 63. GLASS, M. L., S. C. WOOD, R. W. HOYT, AND K. JOHANSEN. Chemical control of breathing in the lizard Varanus exunthemuticua. Comp. B&hem PhysioL A 62: 999-1003,1979. The 64. GLASS, M. L., S. C. WOOD, AND K. JOHANSEN. application of pneumotachography on small unrestrained animals. Camp. Biock PhysioL A 59: 425-427, 1978. in the sea snake 65. GRAHAM, J. B. Aquatic respiration Pelumis pluturus. Respir. PhysioL 21: l-7, 1974. 66. GRATZ, R. K. Ventilatory response of the diamondback water snake, Natrix rhomln@ra, to hypoxia, hypercapnia and increased oxygen demand. J. Comp. PhysioL 129: 105110,1979. 67. GRATZ, R. K., A. AR, AND J. GEISER. Gas tension profile of the lung of the viper, Vipera xunthinu pakstinue. Respir. PhysioL 44: 165-176, 1981. Energetics for ac68. GRATZ, R. K.! AND H. HUTCHISON. tivity in the diamondback water snake, Natrix rhombifma PhysioL Zool. 50: 99-114, 1977. 69 GRIGG, G. C. Metabolic rate, Q10 and respiratory quotient (RQ) in Crocod&s porosus, and some generalizations about low RQ in reptiles. PhysioL ZooL 51: 354-361, 1978. 70 HAZEL, J. R., W. S. GARLICK, AND P. A. SELLNER. The effects of assay temperature upon the pH optima of enzymes from poikilotherms: a test of the imidazole alphastat hypothesis. J. Comp. PhysioL 123: 97-104, 1978. AND Janumy RESPIRJ~TION 1983 110. MERTENS, R. The World of Amphibians and Reptilea New York: McGraw-Hill, 1960. 111. MILSOM, W. K., AND D. R. JONES. Are reptilian pulmonary receptors mechano- or chemosensitive? Nature London 261: 327~323,1976. 112. MILSOM, W. K., AND D. R. JONES. Pulmonary receptor chemosensitivity and the ventilatory response to inhaled CO, in the turtle. Respir. Physid 37: 101-107, 1979. 113. MILSOM, W. K., AND D. R. JONES. The role of vagal afferent information and hypercapnia in the control of the breathing pattern in chelonia. J. Exp Bid 87: 52-63, 1980. 114. MITCHELL, G. S., T. T. GLEESON, AND A. F. BEN- REPTILES 259 NETT. Pulmonary oxygen transport during activity in lizards. Respir. Physid 43: 365-375, 1981. 115. MITHOEFER, J. C. Breath holding. In: Handbud 4f Phyeidc;;rgy. Respiratian, edited by W. 0. Fenn and H. Rahn. Washington, DC: Am. Physiol. Sot., 1964, sec. 3, vol. 2, chapt. 33, p. 1011-1026. 116. MOBERLY, W. R. The metabolic responses of the common iguana, Iguana *no, to walking and diving. Conzp B&n&em Physiol27: 21-37,1968. 117. NAIFEH, K. H., S. E. HUGGINS, H. E. HOFF, T. W. HUGG, AND R. E. NORTON. Respiratory patterns in crocodilian reptiles. Resp+. Phyeid 9: 31-42, 1970. 118. NIELSEN, B. On the regulation of respiration in reptiles. 1. The effect of temperature and CO, on the respiration of lizards (Lacertu). J. Em Bid 38: 301-304, 1961. 119. NIELSEN, B. On the regulation of respiration in reptiles. 2. The effect of hypoxia with and without moderate hypercapnia on the metabolism of lizards. J. Exp Bid 39: 107-117, 1962. 120. PERRY, S. F. Quantitative anatomy of the lungs of the red-eared turtle, P8faulemps scripta eltgana Respir. Physid 35: !&G-262,1978. 121. PERRY, S. F., AND H.-R. DUNCKER. Interrelationship of static mechanical factors and anatomical structure in lung evolution. J. tip Ph&d 138: 321~334.1980. 122. POUGH, F. H. Physiological aspects of burrowing of sand lizards (&nu, &uunw) and other lizards. hp Bit&em Physiol 31: 869-M&1969. 123. POUGH, F. H. Heart rate, breathing and voluntary diving of the elephant trunk snake, AcrocMw javanicw Camp Biachem. Phyti A 44: l&189,1973. 124. POUGH, F. H. Blood oxygen transport and delivery in reptiles. Am Zool 20: 173-185, 1980. 125. POUGH, F. H. The advantages of ectothermy in tetrapods. Am. Nat 115: 92-1X&1980. 126. PRANGE, H. D., AND D. C. JACKSON. Ventilation, gas exchange and metabolic scaling of a sea turtle. &q++. Phyti 27: 369-3’7’7, 1976. 127. RAHN, H. Gas transport from the external environment to the cell. In: Develqpmen of the Lung, edited by U. S. deReuck and R. Porter. Boston: Little, Brown, 1967, p. 223. 128. RAHN, H., AND W. 0. FENN. A Graphical Analysis crf the Respiratory Gas Exchange. Washington, DC: Am. Physiol. Sot., 1955. 129. RAHN, H., AND B. J. HOWELL. The OH-/H+ concept of acid-base balance: historical development. wr. Physiol 33: 91-97,1978. 130. RANDALL, W. C., D. E. STULLKEN, AND W. A. HIESTAND. Respiration of reptiles as influenced by the composition of the inspired air. copeia 1944: 136-144, 1944. 131 REEVES, R. B. An imidazole alphstat hypothesis for vertebrate acid-base regulation: tissue carbon dioxide content and body temperature in bullfrogs. Respir Phw 14: 219-236, 1972. 132. REEVES, R. B. Temperature-induced changes in blood acid-base status: pH and Pa in a binary buffer. J. Appl Phyti 40: 752-761, 1976. 133. ROBIN, E. D., J. W. VESTER, H. V. MURDAUGH, JR., AND J. E. MILLEN. Prolonged anaerobiosis in a vertebrate: anaerobic metabolism in the fresh water turtle. J. C&U Cmnp PhvsioL 63: 287-297.1964. 134. ROMER, A. S. Skin breathing-primary or secondary? Respir. Physid 14: 183-192, 1972. 135. RUBEN, J. Aerobic and anaerobic metabolism during activity in snakes. J. tip Physid 109: 147-157, 1976. 136. SCHEID. P.. W. D. KUHLMANN. AND M. R. FEDDE. Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 93. JOHANSEN, K., C. LENFANT, MD D. HANSON. Phylogenetic development of pulmonary circulation. Fe&r atian proc, Bid 29: 1124-1129,197O. 94. JOHLIN, J. M., AND F. B. MORELAND. Studies of the blood picture of the turtle after complete anoxia. J. Bid them. 103: 107-114,1933. 95. JONES, D. R., AND W. K. MILSOM. Functional characteristics of slowly adapting pulmonary stretch receptors in the turtle. J. Physid Landon 291: 37-49, 1979. 96. JONGH, H. J. DE, AND C. GANS. On the mechanism of respiration in the bullfrog Runu cartesbsiana: a reassessment. J. Ado+& 127: 259~290,1969. 97. KELLOGG, R. H. Central chemical regulation of respiration. In: Handbook uf Physidogy. Respiration, edited by W. 0. Fenn and H. Rahn. Washington, DC: Am. Physiol. Sot., 1964, sect. 3, vol. 1, chapt. 20, p. 507-534. 93. KINNEY, J. L., D. T. MATSUURA, AND F. N. WHITE. Cardiorespiratory effects of temperature in the turtle, Pwudem~jbridana Reqir. Physid 31: 309~325,1977. 99. KLEMM, R. D., R. N. GATE, J. A. WESTFALL, AND R. M. FEDDE. Microanatomy of the lung parenchyma of a tegu lizard %pinati nigropunctat~ J. Mnphol 161: 25’7-280, 1979. loo. KRISTOFFERSON, R., AND A. SOIVIO. Hibernation in the hedgehog (Erim europaeus L.). Anrr. Acud Sci Fenn Ser. A4 82: l-16,1964. 101. KROGH, A., AND 116.KROGH. Rate of diffusion of CO into lungs of man. Skand Arch Ph@uL 23: 236-24?,1999. R. C., AND W. A. CALDER, JR. A prelim102. LASIEWSKI, inary allometric analysis of respiratory variables in resting birds. R.tspir. Physid 11: 152-166,197l. 103. LEGLER, J. M. Natural history of the ornate box turtle, Tewxqwne ornata Agassiz. Univ. Kana Mwr Nat Hiat. Mist Pzdd 11: 527-665,196O. H. The effect of prolonged submersion on 104. LEIVESTAD, the metabolic and heart rate in the toad (Bzcfo 6&o). Acta Univ. Bergensis 5: 1-15, 1960. C., AND K. JOHANSEN. Respiration in the 105. LENFANT, aethiopicwx I. Respiratory African lung&h Pdopterw properties of blood and normal patterns of breathing and gas exchange. J. Exp Bid 49: 437-452,1968. C., K. JOHANSEN, J. A. PEDERSEN, AND 106. LENFANT, K. SCHMIDT-NIELSEN. Respiration in the fresh water turtle Ch&sfinrbriata, Respir. PhysiaL 8: 261-275, 1970. 107. LUCEY, E. C., AND E. W. HOUSE. Effect of temperature on the pattern of lung ventilation and on the ventilationcirculation relationship in the turtle Pseudemys scriptu Cbmp Biochem Physid A 57: 239-243,1977. 108. MALAN, A., T. L. WILSON, AND R. B. REEVES. Intracellular pH in cold-blooded vertebrates as a function of body temperature. Respr. Physid 28: 29-47, 1976. 109. McCUTCHEON, F. H. The respiratory mechanism in turtles. Physd ZooL 16: 255-269, 1943. IN 260 M. L. GLASS AND Vdume 68 ciples and Aduv, edited by M. S. Gordon. New York: Macmillan, 1972, p. 111-159. 151. WHITE, F. N. Circulation. In: Biology @ the Repcilia, Phm A, edited by C. Gans and W. R. Dawson. New York: Academic, 1976, vol. 5, p. 2’75-335. 152 WHITE, F. N., AND G. ROSS. Circulatory changes during experimental diving in the turtle. AM. J. Phpsid 211: 15 18,1966. 153. WITHERS, P. C. Acid-base regulation as a function of body temperature in ectothermic toads, a heliothermic lizard, and a heterothermic mammal. J. M Bid 3: 163-171,1978. 154. WOOD, S. C., M. L. GLASS, AND K. JOHANSEN. Effects of temperature on respiration and acid-base balance in a monitor lixard. J. &uzp Ph@d 116: 28’7~296.1977. 155. WOOD, S. C., AND K. JOHANSEN. Respiratory adaptations to diving in the Nile monitor lixard, Vumnus niloticua J. Cimp Physid 89: 145-X8,1974. 156. WOOD, S. C., K. JOHANSEN, AND R. N. GATE. Pulmonary blood flow, ventilation/perfusion ratio, and oxygen transport in a varanid lizard. Am J. Physid (Rep ulutuq In@pmtive http Physid 2): 233: RS-lW3, WM. 157. WOOD, S. C., K. JOHANSEN, M. L. GLASS, AND R. W. HOYT. Acid-base regulation during heating and cooling . in the lizard, Vamnw exa&hem&cw J Appl Ph&ol: Eizercke Physid Re8pirat. Envim 50: T19-783,198l. 158. WOOD, S. C., K. JOHANSEN, M. L. GLASS, AND G. M. 0. MALOIY. Aerobic metabolism of the lizard Vur. atczc8exan&m&cu~ effects of activity, temperature and size. J. Ph&d 127: 331336.1978. 159. WOOD, S. C., AND C. LENFANT. Respiration: mechanics, control and gas exchange. In: Bw d the Rep&&a Physidogy A, edited by C. Gans and W. R. Dawson. New York: Academic, 1976, vol. 5, p. 225274. 166. WOOD, S. C., AND W. R. MOBERLY. The influence of temperature on the respiratory properties of iguana blood. Req.+. Physid lo: 2O-29,197O. 161. YANCEY, P. H., AND G. N. SOMERO. Temperature dependence of intracellular pH: its role in conservation of pyruvate apparent Km values of vertebrate lactate dehydrogenases. J. Ph@ol125: 129-134,1978. C%nnp Cbmp Downloaded from http://physrev.physiology.org/ by 10.220.33.1 on September 12, 2016 Intrapulmonary receptors in the tegu lizard. II. Functional characteristics and localization. Beep+. Physid 29: 49-63.1977. 13’7. SCHEID, P., H. SLAMA, R. N. GATZ,AND M. R. FEDDE. Intrapulmonary Q receptors in the duck. III. Functional localization. Be@-. Phw8id 22: 123-136, 1974. 138. SCHMIDT-NIELSEN, K. Anid Physidogy. Adaptatian ud E&-me&. London: Cambridge Univ. Press, 1975. 139. SEYMOUR, R. S. Gas tensions and blood distribution in sea snakes at surface pressure and at simulated depth. Phjjsiol ZboL 51: 380-4071978. 146. SEYMOUR, R. S., AND M. E. D. WEBSTER. Gas transport and acid-base balance in diving sea snakes. J. E’ ad 191: 169-l& 1975. 141. SHELTON, G., AND W. BURGGREN. Cardiovascular dynamics of the chelonia during apnoea and lung ventilation. J. Ezp Bti 64~ 323-343,1976. 142. SMITH, H. M., AND D. C. NICKON. Preliminary experiments on the role of the cloaca1 bursae in hibernating turtles. Not Hi& Mist 178: l-8,1961. 143. STAHL, W. R. Scaling of respiratory variables in mammals. J. A& Phpiol22: a-460,1967. 144. STANDAERT, T., AND K. JOHANSEN. Cutaneous gas exchange in snakes. J. tip Ph@oi 89: 313~329,1974. 145. TAKEZAWA, J., F. J. MILLER, AND J. J. O’NEIL. Singlebreath diffusing capacity and lung volumes in small laboratory mammals. J. Appl Phvsiuk Respimt Envim Ezerciee Ph+L 48: 1652-1059,1986. 146. TEMPLETON, J. R., AND W. R. DAWSON. Respiration in the lixard mp/@s adlaria Ph@d ZooL 36: 104126,1963. 147. ULTSCH, G. R., AND D. C. JACKSON. Long-term submergence at 3°C of the turtle, Chtysemys p&a be&i, in normoxic and severely hypoxic water. I. Survival, gas exchange and acid-base status. J. Ezp Bid 96: 11-28, 1982. 148. WEST, J. B. Respiratory Phy8idogy-the Etwntiak Baltimore, MD: Waverly, 1974. 149. WHITE, F. N. Redistribution of cardiac output in the diving alligator. copeia 1963: 567-570,1969. 159. WHITE, F. N. Respiration. In: Animal Phy-: Prin- S. C. WO.OD