Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
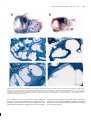
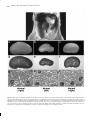
1999 Oxford University Press Human Molecular Genetics, 1999, Vol. 8, No. 4 645–653 Inner ear and kidney anomalies caused by IAP insertion in an intron of the Eya1 gene in a mouse model of BOR syndrome Kenneth R. Johnson*, Susan A. Cook, Lawrence C. Erway1, Angela N. Matthews3, L. Phillip Sanford2, Nancy E. Paradies2 and Rick A. Friedman3 The Jackson Laboratory, 600 Main Street, Bar Harbor, ME 04609, USA, 1University of Cincinnati, Cincinnati, OH 45221, USA, 2Children’s Hospital Medical Center, Cincinnati, OH 45267, USA and 3House Ear Institute, Los Angeles, CA 90057, USA Received November 18, 1998; Revised and Accepted January 17, 1999 A spontaneous mutation causing deafness and circling behavior was discovered in a C3H/HeJ colony of mice at the Jackson Laboratory. Pathological analysis of mutant mice revealed gross morphological abnormalities of the inner ear, and also dysmorphic or missing kidneys. The deafness and abnormal behavior were shown to be inherited as an autosomal recessive trait and mapped to mouse chromosome 1 near the position of the Eya1 gene. The human homolog of this gene, EYA1, has been shown to underly branchio-otorenal (BOR) syndrome, an autosomal dominant disorder characterized by hearing loss with associated branchial and renal anomalies. Molecular analysis of the Eya1 gene in mutant mice revealed the insertion of an intracisternal A particle (IAP) element in intron 7. The presence of the IAP insertion was associated with reduced expression of the normal Eya1 message and formation of additional aberrant transcripts. The hypomorphic nature of the mutation may explain its recessive inheritance, if protein levels in homozygotes, but not heterozygotes, are below a critical threshold needed for normal developmental function. The new mouse mutation is designated Eya1 bor to denote its similarity to human BOR syndrome, and will provide a valuable model for studying mutant gene expression and etiology. INTRODUCTION Hearing loss is the most common sensory deficit in humans and affects ∼1 in 2000 live births (1). Tremendous progress has been made during the last 5 years in mapping and cloning of genes responsible for both syndromic and non-syndromic hereditary hearing loss (2,3). The mouse is an excellent animal model for the DDBJ/EMBL/GenBank accession nos AF097544–AF097546 study of these human conditions because the anatomy, function and hereditary abnormalities of the inner ear have been shown to be similar in both humans and mice (4,5). Genetic and molecular analyses of mouse deafness mutations have aided the discovery of genes that underly several human hearing loss syndromes. For example, the mouse shaker-1 mutation (sh1) was shown to be a mutation of Myo7a (6), and the homologous gene in humans subsequently was shown to be responsible for both dominant (DFNA11) and recessive (DFNB2) forms of non-syndromic deafness (7,8), as well as for Usher syndrome type 1B (9). Recently, the mouse shaker-2 mutation (sh2) was shown to be a mutation of Myo15 (10), and the homologous gene in humans to be responsible for DFNB3 (11). In mice, mutations affecting the vestibular system of the inner ear often result in a characteristic circling or head-bobbing phenotype; many of these mutations also affect the cochlea and cause deafness (4). As part of our research program at the Jackson Laboratory to identify genes causing deafness, we have been selecting and studying mutant mice that exhibit behavior characteristic of vestibular dysfunction. Pathological analysis of one such circling mutant revealed gross morphological abnormalities of the inner ear and also dysmorphic or missing kidneys. Here, we describe the inner ear and kidney pathology of these mutant mice and our genetic mapping results that place the new mutation on proximal chromosome 1, near the position of the Eya1 gene (12). The human homolog of this gene, EYA1, has been shown to underly branchio-oto-renal (BOR) syndrome, an autosomal dominant disorder characterized by hearing loss with associated branchial and renal anomalies (13). Because of the coincident map position and the mutation’s phenotypic similarity to human BOR syndrome, we considered Eya1 to be a likely candidate gene for the new mouse mutation and undertook a molecular analysis of this gene in mutant mice. We show that Eya1 mRNA expression is altered in mice homozygous for the new mutation as compared with their normal, co-isogenic littermates, and demonstrate that this altered expression is caused by the spontaneous insertion of an intracisternal A particle (IAP) element into intron 7 of the Eya1 gene. *To whom correspondence should be addressed. Tel: +1 207 288 6228; Fax: +1 207 288 6149; Email: [email protected] 646 Human Molecular Genetics, 1999, Vol. 8, No. 4 RESULTS Mice with circling and head-bobbing behavior were discovered in a C3H/HeJ colony at the Jackson Laboratory. The abnormal behavior was shown to be inherited as an autosomal recessive trait. Affected male homozygotes would sometimes breed; females did not. Auditory-evoked brainstem response (ABR) threshold measurements demonstrated that homozygous mutant mice are deaf (no evoked response to sound pressure levels >95 dB) at the earliest testable age (3–4 weeks), but heterozygotes hear normally. Mice homozygous for the mutation were also characterized by absent or dysmorphic kidneys. The adult inner ears of the mutant mice displayed a number of morphogenetic abnormalities (Fig. 1). Whole-mount preparations revealed subtle abnormalities of the pars superior, or vestibular portion of the inner ear. Specifically, the lateral semicircular canal, the last to appear developmentally, is foreshortened, with a much narrower diameter than that of the wild-type (not shown). Several of the postnatal inner ears studied also revealed an incomplete common crus, the region of the joined non-ampullated ends of the superior and posterior semicircular canals (not shown). The abnormalities of the pars inferior, or cochlear portion of the inner ear, were the most severe and constant. All but the most basal one-quarter of the cochlea was absent in the adult mutant inner ear (Fig. 1a and b). Histological analysis demonstrated the rudimentary basal portion of the mutant cochlea with a spiral ligament and no overlying stria vascularis. Additionally, there was complete absence of the organ of Corti, or sensory epithelium, in the mutant (Fig. 1c and d). No abnormalities of the pharyngeal pouches were noted. The kidney phenotype seen in post-wean mutants showed variable expressivity, ranging from bilaterally normal kidneys to unilateral absence (Fig. 2). Most often, bilateral deficiencies were observed, with the left kidney being more severely affected than the right. Even in cases of severe hypoplasia, the cellular morphology of mutant kidneys was very similar to that of heterozygotes and wild-type controls. However, functional stress in mutant kidneys was indicated by plasma urea nitrogen values which corresponded to the degree of kidney deficiency. The reduced function of mutant kidneys, therefore, is most likely caused by an insufficiency in the number of nephrons rather than to structural defects at the cellular level. Plasma urea nitrogen values ranged from 26 to 56 mg/dl (mean ± SEM = 36.0 ± 8.1, n = 13) in mutant mice compared with 18–28 mg/dl for sibling controls (mean ± SEM = 23.4 ± 3.0, n = 14). Student’s one-tailed t-test indicated that the mutant urea nitrogen levels were significantly elevated (P = 0.01). Matings between mutationcarrying mice produced small litter sizes and fewer than expected numbers of affected mice, suggesting that homozygosity for the mutation may sometimes cause perinatal lethality, perhaps because of occasional bilateral kidney agenesis or severe bilateral hypoplasia. To map the new mutation genetically, an intercross was made between F1 hybrids produced from matings between homozygous mutant mice and mice from the wild-derived inbred strain CAST/Ei. Intercross progeny with obvious vestibular dysfunction were typed for polymorphic markers dispersed throughout the genome. Linkage was found with markers on chromosome 1. The haplotypes of 58 affected intercross progeny (116 tested meioses) were examined to establish gene order and interlocus recombination distances. No recombination was observed be- tween the new mutation and the markers D1Mit68, D1Mit4 or D1Mit52, and ∼7% recombination (8/116) was observed between this cluster and D1Mit71. These results placed the mutation ∼10 cM from the chromosome 1 centromere, near the recently mapped Eya1 gene (12). DNA sequence comparisons of RT–PCR products covering the protein-coding region of the Eya1 cDNA (nucleotides 153–1926; 12) failed to reveal any differences between mutant and control mice; however, a gross alteration of the Eya1 gene in affected mice compared with wild-type controls was detected by Southern blot analysis (Fig. 3). Genomic DNA from affected homozygous mice, heterozygotes and +/+ control mice hybridized with a 1.6 kb Eya1 cDNA probe revealed differences in banding patterns with five restriction enzymes. The mutant fragment sizes were larger than control fragments in EcoRI and PvuII digests and smaller than controls in PstI, MspI and BglII digests, suggesting that the mutation was a sizeable (3–5 kb) genomic insertion or deletion within the Eya1 gene. The overall size and exon–intron structure of the mouse Eya1 gene appeared similar to that described for the human EYA1 gene (14). Because of its large size (the human EYA1 gene consists of 16 coding exons and extends over 156 kb), the region of the Eya1 gene containing the mutation was narrowed by sequential hybridization of Southern blots with probes from increasingly restricted portions of the Eya1 cDNA (Fig. 3). The site of the mutation was thus refined to a 3 kb genomic region containing exon 8. To compare DNA sequences in the mutated region of the Eya1 gene, genomic clones were identified by screening phage libraries from both mutant and wild-type mice with a 106 bp exon 8 probe. Restriction site mapping was then used to localize the mutation to intron 7 (Fig. 4), and ∼3500 nucleotides of DNA from that region were sequenced (GenBank accession no. AF097544). PCR amplification with various combinations of primers designed from this sequence further narrowed the region of mutation to a 300 bp interval; primers spanning this interval failed to amplify a product with mutant genomic DNA. Sequence analysis of the mutant DNA clone in this region revealed an insertion, ∼1480 bp upstream from the 3′ splice acceptor sequence of exon 8. A BLAST search of GenBank DNA sequences identified the insertion as an IAP transposon; 32 matches had alignment scores >200, including those IAP elements causing pale ear (ep) and agouti (A) mutations. The IAP insertion was in the same transcriptional orientation as the Eya1 gene. The single mutant genomic clone did not contain the 5′ end of the IAP element; therefore, the DNA sequence of the cloned 3′ long terminal repeat (LTR) together with the Eya1 intron sequence 5′ to the IAP insertion site were used to amplify the uncloned 5′ LTR junction fragment from mutant genomic DNA. Sequence analysis of this PCR product revealed that the 5′ and 3′ LTR sequences were identical and that the IAP element was flanked by a 6 bp direct repeat (GGTAGG), a characteristic of retrotransposition. The nucleotide sequences of the IAP junctions with intron 7 of the Eya1 gene have been deposited in GenBank under accession nos AF097545 (5′LTR) and AF097546 (3′ LTR). Northern blot analysis of total RNA from adult skeletal muscle hybridized with a 1.6 kb Eya1 cDNA probe showed an ∼50% reduction in the level of wild-type message in homozygous mutant mice compared with +/+ controls (Fig. 5a). Furthermore, in RNA from mutant mice, two transcripts of ∼7 and 10 kb were 647 Human Genetics, 1999, 8, No. NucleicMolecular Acids Research, 1994, Vol. Vol. 22, No. 1 4 647 Figure 1. Effect of the Eya1 mutation on adult inner ear morphology. Cleared whole-mount preparations (a and b) compared at equal magnification, and histological sections (c and d, 80× magnification, and e and f, 200× magnification) of inner ears from C3H/Hej +/+ control (a, c and e) and C3H/HeJ-Eya1 bor/Eya1 bor mutant (b, d and f) mice. Asterisks indicate the distinct interscalar septi of a normal cochlea (a); arrowheads designate a normal cochlea in the +/+ control (a) and its absence in the mutant inner ear (b). The stria vascularis (small arrow) and organ of Corti (large arrow) shown in the +/+ control (e) are absent in the mutant inner ear (f). sl designates the spiral ligament (e and f). seen in addition to the 4 kb wild-type message. Additional transcripts were also seen in RNA from eye and brain tissue of mutant mice (data not shown); however, Eya1 expression is much reduced and more difficult to quantify in these tissues compared with skeletal muscle expression. RT–PCR analysis with primers flanking exon 8 also produced additional products from mutant RNA, larger in size than the product expected from the wild-type Eya1 transcript (Fig. 5b). 648 Human Molecular Genetics, 1999, Vol. 8, No. 4 Figure 2. Effect of the Eya1 mutation on adult kidney morphology. Top center: autopsy of a 4-month-old male mutant showing complete absence of the left kidney. The right kidney, indicated by an arrow, shows normal morphology. The intestinal tract was excised intentionally and the left ureter is partially obscured by the pancreas. Both testes were present. Bottom panels: the right kidney from a 7.5-month-old female +/? control (a, d and g) is compared with the left (b, e and h) and right (c, f and i) kidneys of a 7.5-month-old female sibling mutant: kidney whole-mounts 10× (a, b and c); longitudinal sections, 10×, H&E stained (d, e and f); and cortex sections, bar = 100 µm, PAS (g, h and i). Although hypoplasia is obvious in both mutant kidneys, their cellular morphology is similar to that of the control kidney. However, functional stress is indicated by elevated plasma urea nitrogen (32 mg/dl for the mutant versus 18 mg/dl for control). 649 Human Genetics, 1999, 8, No. NucleicMolecular Acids Research, 1994, Vol. Vol. 22, No. 1 4 649 Figure 3. Southern blot analysis of the Eya1 mutation. Genomic DNA from C3H/HeJ controls (+/+; lanes marked +), heterozygotes (+/Eya1 bor; lanes marked H) and homozygous mutant mice (Eya1 bor/Eya1 bor; lanes marked M) were digested with EcoRI, PstI, PvuII, MspI and BglII and hybridized with three Eya1 cDNA probes: (a) exons 1–13 (nucleotides 1–1600), (b) exons 5–10 (nucleotides 592–1277) and (c) exon 8 (nucleotides 992–1097). Exon numbering corresponds to Abdelhak et al. (13) and nucleotide numbering to Xu et al. (12). The same fragment polymorphisms were detected with all three probes. The mutant fragment sizes were larger in EcoRI and PvuII digests and smaller in PstI, MspI and BglII digests. DISCUSSION Both the new mouse mutation and human BOR syndrome are caused by alterations of homologs of the Drosophila eyes absent gene (mouse Eya1, human EYA1). These disorders are consistent with the role of this gene in early embryonic development. BOR syndrome is characterized by developmental anomalies of the branchial arches (pre-auricular pits, branchial cysts, outer and middle ear malformations), the inner ear (absent or undeveloped cochlea and semicircular canals) and the kidneys (kidney hypoplasia to bilateral agenesis) (13,15). The prominent features of the mouse Eya1 mutation, including cochlear and kidney hypoplasia, are strikingly similar to those of BOR syndrome. The reduced, but still present, expression of normal Eya1 transcripts in affected homozygotes is evidence that the mouse mutation is hypomorphic, which may explain its recessive nature compared with the dominant inheritance of human BOR syndrome. The ∼50% level of Eya1 expression observed in homozygous mutant mice (Fig. 5) might be below a critical physiological threshold necessary for normal function, but the expression level in heterozygous mice may be high enough to allow normal development. The EYA1 mutations reported for human BOR syndrome alter protein-coding sequences in exons or disrupt splice sites (13,14,16), and thus are likely to be more severe than the insertional disruption of an intron reported here for the mouse. Consequently, human heterozygotes may have gene expression levels similar to those of homozygous mice. The additional transcripts produced by the mouse mutation are unlikely to cause a gain of function or have dominant-negative effects because no phenotypic abnormalities were seen in heterozygotes even though aberrant transcripts were detected in these mice. The mouse genome contains ∼2000 copies of retroviral-like IAPs (17). IAP retrotranspositions have been shown to underly several inherited mutations in the mouse, including agouti, A (18); pale ear, ep (19); vibrator, Pitpn vb (20); Lamb3 (21); reeler, Reln rl (22); albino, Tyrc (23); fused, Axin Fu (24); and β-glucuronidase, Gus mps-2J (25). IAP element insertions into non-coding introns have been shown to cause reduced expression and/or formation of chimeric transcripts in the Pitpn vb (20), Axin Fu (24) and Gus mps–2J (25) mutations, similar to our findings for the Eya1 bor mutation. It is thought that IAP element insertions introduce cryptic splice and polyadenylation sites that disrupt normal transcript processing or stability and consequently reduce gene expression. In the Eya1 bor mutation, the junction of the 6 bp direct repeat with the 5′ LTR of the IAP created a consensus 5′ splice site (AGGTGTG; Fig. 4c), which, along with two LTR polyadenylation sites, may contribute to the aberrant RNA expression observed in northern blots and RT–PCR (Fig. 5). The DNA sequence of the Eya1 IAP LTR (Fig. 4) indicates that the U3 region was derived from the LS-type and the R-region from the T-type of IAP elements; this composite LTR structure is characteristic of other IAP insertions causing germline mutations, most of which have occurred in the C3H/He inbred strain (25). The Eya1 IAP LTR sequence is very similar to the LTRs of other IAP elements isolated from C3H/He mice. It is identical to the LTR of the IAP causing the pale ear mutation (GenBank AF003867), except that 30 bp are deleted at the position indicated in Figure 4, possibly the result of unequal crossing-over in this region of imperfect CT-rich repeats. 650 Human Molecular Genetics, 1999, Vol. 8, No. 4 Figure 4. Structure and sequence of the Eya1 gene in the region of IAP insertion. (a) Gene structure and site of IAP insertion, ∼1480 bp upstream of exon 8. Exons are indicated by black rectangles. Landmark restriction sites and PCR primers (shown as arrowheads indicating direction) are shown at their approximate locations. (b) DNA sequence of IAP LTRs flanked by the 6 bp direct repeat sequence GGTAGG, underlined. The start of each LTR is indicated above the sequence. A 30 bp deletion (compared with the pale ear IAP LTR, GenBank accession no. AF003867) in the CT-rich region of the LTRs is indicated by three asterisks. A consensus 5′ splice site, shown in bold, is created by the junction of the 5′ LTR with the 6 bp direct repeat. (c) PCR genotyping of mice. Two forward primers, one specific to the IAP insertion (IAPF1) and the other specific to the 5′-flanking region of intron 7 (gF5), were combined with one reverse primer specific to the 3′-flanking region of intron 7 (gR5). In combination, the three primers distinguish +/+ (lane 1), +/Eya1 bor (lane 2) and Eya1 bor/Eya1 bor(lane 3) genotypes. Primers IAPF1 and gR5 amplify mutant DNA (160 bp product) but not wild-type DNA, whereas primers gF5 and gR5 amplify wild-type (285 bp product) but not mutant DNA. In summary, we believe that retrotransposition of an IAP element into intron 7 of the Eya1 gene is responsible for the phenotype of the new mouse mutation for the following reasons. (i) The inner ear and kidney abnormalities of mutant mice are similar to the characteristic traits of human BOR syndrome, known to be caused by mutations of the human EYA1 gene. (ii) The new mouse mutation and the Eya1 gene co-segregated in 116 tested meioses from the linkage cross. (iii) Northern blot and RT–PCR analyses showed that Eya1 gene expression is altered in mutant mice compared with controls. (iv) Southern blot and DNA sequence analysis revealed that an IAP element was present in intron 7 of the Eya1 gene of mutant mice but not in co-isogenic controls. (v) PCR genotyping (Fig. 4c) has confirmed that all mutant mice are homozygous for this IAP element insertion. (vi) Other inherited mouse mutations have been shown to be caused by similar IAP element insertions into introns. This new mouse model will permit studies of Eya1 gene expression and etiology not possible in humans with BOR syndrome. For example, linkage backcross mice homozygous for the Eya1 mutation, with varying genetic backgrounds derived from parental C3H/HeJ and CAST/Ei genomes, displayed variable phenotypes for both inner ears and kidneys. Analysis of such genetically well-defined mice will permit identification of genes that modify the phenotypic manifestation of the Eya1 mutation and may provide insight into causes of the incomplete penetrance and variable expressivity characteristic of human BOR syndrome (15). The mouse mutation also provides a means to study mutant Eya1 gene expression during embryonic development. Other developmental control genes such as Pax2 are also known to contribute to inner ear and kidney patterning (26). Studies can now be undertaken to examine the role of Eya1 in molecular pathways leading to these common morphogenetic events. MATERIALS AND METHODS Genetic mapping PCR primer pairs (MapPairs) for microsatellite markers distributed throughout the mouse genome were purchased from Research Genetics (Huntsville, AL) and typing was performed as 651 Human Genetics, 1999, 8, No. NucleicMolecular Acids Research, 1994, Vol. Vol. 22, No. 1 4 651 gent Hearing Systems, Miami, FL) was used to obtain ABR thresholds for clicks and for pure-tone pips at 8, 16 and 32 kHz, as described previously (29). Genomic DNA and RNA, and cDNA preparation Figure 5. RNA expression analyses of mutant Eya1. (a) Northern blot analysis. Total cellular RNA extracted from skeletal muscle of wild-type C3H/HeJ (lane marked +) and homozygous mutant mice (lane marked M) was hybridized with a 1.6 kb Eya1 cDNA probe (exons 1–13). Relative optical density measurements were 1300 and 730 for the normal 4 kb Eya1 transcript from wild-type and mutant mice, respectively. The densities of the additional 7 and 10 kb transcripts in mutant mice were 420 and 120, respectively. Density measurements for the two Actb transcripts indicated that equal quantities of RNA were loaded in each lane. Migration positions for 28S and 18S rRNAs are shown on the left. (b) RT–PCR analysis. cDNA prepared from skeletal muscle RNA from one control (lane marked +) and three mutant (lanes marked M) mice was used as PCR template with primers amplifying the exon 5–8 region (nucleotides 592–1097) (12) of the Eya1 cDNA. previously described (27), except that PCR reactions were carried out for 30 cycles and products were separated on 3% agarose gels (Metaphor; FMC BioProducts, Rockland, ME) and visualized by ethidium bromide staining. Gene order, determined by minimizing the number of obligate cross-over events, and recombination frequency estimates were calculated with the aid of the Map Manager computer program (28). Histological and clinical analyses Adult inner ears were harvested after transcardial perfusion with 1% paraformaldehyde, 1% glutaraldehyde in phosphate buffer, pH 7.2. For light microscopy, inner ears were dissected and post-fixed in buffered 4% OsO4, dehydrated in ethanol and propylene oxide, and embedded in SPURR resin (Polysciences, Warrington, PA). Semithin sections (1–2 µm) were cut with a diamond knife and stained with 0.5% toluidine blue in 0.5% sodium borate. Kidneys were fixed in Bouin’s, halved longitudinally and embedded in paraffin. Sections (5 µm) were then stained with hematoxylin and eosin (H&E) or periodic acid–Schiff (PAS). Plasma was obtained by retro-orbital sinus bleed, and urea nitrogen values were assessed by Affiliated Laboratories (Bangor, ME). ABR threshold determinations Mice were tested at the University of Cincinnati for ABR thresholds. A computer-assisted evoked potential system (Intelli- Genomic DNA was prepared from mouse spleens by standard phenol–chloroform extraction and ethanol precipitation methods. Total RNA was purified from mouse brain, eye and skeletal muscle tissues with TRIzol reagent, according to the manufacturer’s protocol (Gibco BRL, Gaithersburg, MD). DNA and RNA concentrations were estimated by spectrophotometric measurements of absorbance. cDNA was prepared from total RNA from adult mouse skeletal muscle with the SuperScript Preamplification System for First Strand cDNA Synthesis (Gibco BRL). Southern and northern blots Blotting, probe labeling and hybridization procedures used for both Southern and northern blots were as previously described (30). A 1.6 kb fragment of the mouse Eya1 cDNA, containing most of the protein-coding sequence, was used as a probe for initial Southern and northern blot analyses. Additional probes for Southern analysis were produced by PCR amplification with the primers described below. For northern analysis, 15 µg of total RNA was loaded per lane on a 0.8% agarose gel with 2.2 M formaldehyde, electrophoresed in 1× MOPS buffer at 2 V/cm for 4 h, vacuum blotted in 10× SSC onto a positively charged nylon membrane, and UV cross-linked. For quantitative estimation of transcript abundance, densitometry measurements were made on developed X-ray film using the Computing Densitometer and ImageQuant software from Molecular Dynamics (Sunnyvale, CA). Genomic clones The wild-type Eya1 clone was isolated by standard filter hybridization screening methods from a commercially available lambda phage library prepared with 129/SvJ mouse genomic DNA (Lambda FIX II; Stratagene, La Jolla, CA). The mutant Eya1 clone was isolated from a custom-made phage library (Lambda FIX II) prepared with genomic DNA from homozygous mutant mice. PCR primers PCR primers used to amplify portions of the mouse Eya1 cDNA for sequence comparisons and as probes for Southern blot analysis are given in Table 1. Primers used for sequence comparisons of genomic DNA are given in Table 2; their approximate positions are shown in Figure 4a. Primers ex8F and ex8R were used to amplify the genomic region around exon 8 (nucleotides 992–1097). Primers gF5 and IAPR1 were used to amplify the junction of intron 7 with the IAP 5′ LTR. Primers gF5 and gR5 were used to amplify wild-type, but not mutant DNA. Primers IAPF1 and gR5 were used to amplify mutant, but not wild-type DNA. All three primers (gF5, IAPF1 and gR5) were used simultaneously to distinguish genotypes (Fig. 4c). 652 Human Molecular Genetics, 1999, Vol. 8, No. 4 Table 1. Primers used for cDNA sequence comparisons between mutant and control Forward primer Reverse primer Amplified nucleotides EYA1F1 EYA1R4 153–833 ATGGAAATGCAGGATCTAAC TGTGCGTACTGACCCTGGCC EYA1F3 EYA1R6 GGAAAGTGGATTGTCACAGT AAAAATAGATGTGTGTCTGC EYA1F5 EYA1R8 CGTCGAGGTTCAGATGGGAA CTATTGGAAACACAATTCCT 592–1277 1036–1720 EYA1F7 CON1R GATCTACAACACCTACAAAA CAGGTACTCTAATTCCAAGG 1478–1926 All primers are presented in the 5′→3′ orientation. An ‘F’ signifies a forward primer and an ‘R’ a reverse primer according to the Eya1 direction of transcription. Nucleotide numbering corresponds to the mouse Eya1 cDNA sequence reported by Xu et al. (1997), GenBank accession no. U61110. Table 2. Primers used for analysis of Eya1 genomic DNA Primer Sequence ex8F ACAGTCCTTCCACACCCATT ex8R GGAGGGGAGGGATTATTGTT gR2 CCAGGGAGCCTGATATTTGA gR4 TCTTCCTCACTGTGATCTTGTTG gR6 CAGAAAAATTGATGAAACTGGAGA gR7 GTCTCCACCATGGCTTCAAT gF1 TGCTGAGTCCTGGTGATCTG gF2 TAGTCCCCATGTGCTTCCAT gF3 TAGGGCTCTTCCCAAAGCTG gF5 TCTCCAGTTTCATCAATTTTTCTG gR5 CAGATCACCAGGACTCAGCA IAPR1 CAGACCAGAATCTTCTGCGA IAPF1 TCGCAGAAGATTCTGGTCTG Cycle Sequencing method. The same primers used for PCR amplification were also used for cycle sequencing. ACKNOWLEDGEMENTS We thank Dr Richard Maas (Harvard Medical School, Boston, MA) for his generous gift of the Eya1 cDNA clone and for sharing primer sequence information. We thank personnel of the the Jackson Laboratory (TJL) Microchemistry Service for rapid and high quality DNA sequencing (Amy Lambert and Doug McMinimy) and library screening and clone purification (Kevin Johnson). We thank Emma Lou Cardell (University of Cincinnati) and Rod Bronson (TJL) for their histological expertise. We also thank Patsy Nishina and Babette Gwynn (TJL) for their careful review of this manuscript. This study was supported by National Institutes of Health grants GM46697, RR01183, CA34196, DC00119, and contract DC62108. REFERENCES For explanation, see Table 1. Primer positions are indicated in Figure 4a. PCR reactions The following reaction conditions were used for PCR amplifications: 20–50 ng of template DNA, 50 mM KCl, 10 mM Tris–HCl, 0.01% Triton X-100, 2.25 mM MgCl2, 100 nM of each primer, 100 µM of each of four deoxyribonucleoside triphosphates and 0.5 U of Taq DNA polymerase. Amplification consisted of one cycle of denaturation at 94C for 3 min followed by 35 cycles, each consisting of 94C for 15 s denaturation, 60C for 2 min of annealing and 72C for 2 min of extension. After the 35 cycles, the final product was extended for 7 min at 72C. PCR reaction products were separated on 2.5% Metaphor agarose gels, stained with ethidium bromide and visualized with UV light. DNA sequencing PCR-amplified products from cDNA and genomic DNA templates were excised from gels and purified with QIAquick Gel Extraction kit (Qiagen, Valencia, CA). DNA was sequenced using an Applied Biosystems 373A DNA Sequencer (Perkin Elmer, Norwalk, CT) and an optimized DyeDeoxy Terminator 1. Morton, N.E. (1991) Genetic epidemiology of hearing loss. Ann. NY Acad. Sci., 630, 16–31. 2. Kalatzis, V. and Petit, C. (1998) The fundamental and medical impacts of recent progress in research on hereditary hearing loss. Hum. Mol. Genet., 7, 1589–1597. 3. Van Camp, G. and Smith, R.J. (1998) Hereditary Hearing Loss Homepage. http://dnalab-www.uia.ac.be/dnalab/hhh 4. Brown, S.D. and Steel, K.P. (1994) Genetic deafness—progress with mouse models. Hum. Mol. Genet., 3, 1453–1456. 5. Steel, K.P. (1995) Inherited hearing defects in mice. Annu. Rev. Genet., 29, 675–701. 6. Gibson, F., Walsh, J., Mburu, P., Varela, A., Brown, K.A., Antonio, M., Beisel, K.W., Steel, K.P. and Brown, S.D.M. (1995) A type VII myosin encoded by the mouse deafness gene shaker-1. Nature, 374, 62–64. 7. Liu, X.Z., Walsh, J., Tamagawa, Y., Kitamura, K., Nishizawa, M., Steel, K.P. and Brown, S.D. (1997) Autosomal dominant non-syndromic deafness caused by a mutation in the myosin VIIA gene. Nature Genet., 17, 268–269. 8. Liu, X.Z., Walsh, J., Mburu, P., Kendrick-Jones, J., Cope, M.J., Steel, K.P. and Brown, S.D. (1997) Mutations in the myosin VIIA gene cause non-syndromic recessive deafness. Nature Genet., 16, 188–190. 9. Weil, D., Blanchard, S., Kaplan, J., Guilford, P., Gibson, F., Walsh, J., Mburu, P., Varela, A., Levilliers, J., Weston, M.D. et al. (1995) Defective myosin VIIA gene responsible for Usher syndrome type 1B. Nature, 374, 60–61. 10. Probst, F.J., Fridell, R.A., Raphael, Y., Saunders, T.L., Wang, A., Liang, Y., Morell, R.J., Touchman, J.W., Lyons, R.H., Noben-Trauth, K., Friedman, T.B. and Camper, S.A. (1998) Correction of deafness in shaker-2 mice by an unconventional myosin in a BAC transgene. Science, 280, 1444–1447. 653 Human Genetics, 1999, 8, No. NucleicMolecular Acids Research, 1994, Vol. Vol. 22, No. 1 4 11. Wang, A., Liang, Y., Fridell, R.A., Probst, F.J., Wilcox, E.R., Touchman, J.W., Morton, C.C., Morell, R.J., Noben-Trauth, K., Camper, S.A. and Friedman, T.B. (1998) Association of unconventional myosin MYO15 mutations with human nonsyndromic deafness DFNB3. Science, 280, 1447–1451. 12. Xu, P.-X., Woo, I., Her, H., Beier, D.R. and Maas, R.L. (1997) Mouse Eya homologues of the Drosophila eyes absent gene require Pax6 for expression in lens and nasal placode. Development, 124, 219–231. 13. Abdelhak, S., Kalatzis, V., Heilig, R., Compain, S., Samson, D., Vincent, C., Weil, D., Cruaud, C., Sahly, I., Leibovici, M., Bitner-Glindzicz, M., Francis, M., Lacombe, D., Vigneron, J., Charachon, R., Boven, K., Bedbeder, P., Van Regemorter, N., Weissenbach, J. and Petit, C. (1997) A human homologue of the Drosophila eyes absent gene underlied branchio-oto-renal (BOR) syndrome and identifies a novel gene family. Nature Genet., 15, 157–164. 14. Abdelhak, S., Kalatzis, V., Heilig, R., Compain, S., Samson, D., Vincent, C., Levi-Acobas, F., Cruaud, C., Le Merrer, M., Mathieu, M., Konig, R., Vigneron, J., Weissenbach, J., Petit, C. and Weil, D. (1997) Clustering of mutations responsible for branchio-oto-renal (BOR) syndrome in the eyes absent homologous region (eyaHR) of EYA1. Hum. Mol. Genet., 6, 2247–2255. 15. OMIM (1998) Online Mendelian Inheritance in Man (OMIM) (Baltimore, MD: Center for Medical Genetics, Johns Hopkins University URL: http://www3.ncbi. nlm. nih. gov/omim/ ). 16. Kumar, S., Kimberling, W.J., Weston, M.D., Schaefer, B.G., Berg, M.A., Marres, H.A. and Cremers, C.W. (1998) Identification of three novel mutations in human EYA1 protein associated with branchio-oto-renal syndrome. Hum. Mutat., 11, 443–449. 17. Kuff, E.L. and Lueders, K.K. (1988) The intracisternal A-particle gene family: structure and functional aspects. Adv. Cancer Res., 51, 183–276. 18. Duhl, D.M., Vrieling, H., Miller, K.A., Wolff, G.L. and Barsh, G.S. (1994) Neomorphic agouti mutations in obese yellow mice. Nature Genet., 8, 59–64. 19. Gardner, J.M., Wildenberg, S.C., Keiper, N.M., Novak, E.K., Rusiniak, M.E., Swank, R.T., Puri, N., Figner, J.N., Hagiwara, N., Lehman, A.L., Gales, T.L., Bayer, M.E., King, R.A. and Brilliant, M.H. (1997) The mouse pale ear (ep) mutation is the homologue of human Hermansky–Pudlak syndrome. Proc. Natl Acad. Sci. USA, 94, 9238–9243. 653 20. Hamilton, B.A., Smith, D.J., Mueller, K.L., Kerrebrock, A.W., Bronson, R.T., van Berkel, V., Daly, M.J., Kruglyak, L., Reeve, M.P., Nemhauser, J.L., Hawkins, T.L., Rubin, E.M. and Lander, E.S. (1997) The vibrator mutation causes neurodegeneration via reduced expression of PITPa: positional complementation cloning and extragenic suppression. Neuron, 18, 711–722. 21. Kuster, J.E., Guarnieri, M.H., Ault, J.G., Flaherty, L. and Swiatek, P.J. (1997) IAP insertion in the murine LamB3 gene results in junctional epidermolysis bullosa. Mamm. Genome, 8, 673–681. 22. Royaux, I., Bernier, B., Montgomery, J.C., Flaherty, L. and Goffinet, A.M. (1997) Relnrl-Alb2, an allele of reeler isolated from a chlorambucil screen, is due to an IAP insertion with exon skipping. Genomics, 42, 479–482. 23. Wu, M., Rinchik, E.M., Wilkinson, E. and Johnson, D.K. (1997) Inherited somatic mosaicism caused by an intracisternal A particle insertion in the mouse tyrosinase gene. Proc. Natl Acad. Sci. USA, 94, 890–894. 24. Vasicek, T.J., Zeng, L., Guan, X.-J., Zhang, T., Costantini, F. and Tilghman, S.M. (1997) Two dominant mutations in the mouse Fused gene are the result of transposon insertions. Genetics, 147, 777–786. 25. Gwynn, B., Lueders, K., Sands, M.S. and Birkenmeier, E.H. (1998) Intracisternal A-particle element transposition into the murine beta-glucuronidase gene correlates with loss of enzyme activity: a new model for beta-glucuronidase deficiency in the C3H mouse. Mol. Cell. Biol., 18, 6474–6481. 26. Torres, N., Gomezpardo, E. and Gruss, P. (1996) Pax2 contributes to inner ear patterning and optic nerve trajectory. Development, 122, 3381–3391. 27. Dietrich, W., Katz, H., Lincoln, S.E., Shin, H.-S., Friedman, J., Dracopoli, N.C. and Lander, E.S. (1992) A genetic map of the mouse suitable for typing intraspecific crosses. Genetics, 131, 423–447. 28. Manly, K.F. (1993) A Macintosh program for storage and analysis of experimental genetic mapping data. Mamm. Genome, 4, 303–313. 29. Erway, L.C., Willott, J.F., Archer, J.R. and Harrison, D.E. (1993) Genetics of age-related hearing loss in mice: I. Inbred and F1 hybrid strains. Hear. Res., 65, 125–132. 30. Johnson, K.R., Cook, S.A. and Davisson, M.T. (1992) Chromosomal localization of the murine gene and two related sequences encoding high-mobility-group I and Y proteins. Genomics, 12, 503–509.