Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
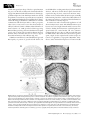
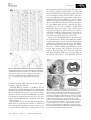
Review Blackwell Publishing Ltd. Tansley review Models of shoot apical meristem function Author for correspondence: Fiona Tooke Email: [email protected] Received: 16 January 2003 Accepted: 3 April 2003 Fiona Tooke† and Nick Battey †Department of Plant Sciences, Cambridge University, Cambridge, CB2 3EA, UK; School of Plant Sciences, The University of Reading, Whiteknights, Reading, RG6 6AS, UK doi: 10.1046/j.1469-8137.2003.00803.x Contents Summary 37 V. Developmental genetics 44 I. 37 VI. Conclusions 49 II. How things began 38 Acknowledgements 50 III. Cytology 39 References 50 IV. Morphology 41 Introduction Summary Key words: shoot apical meristem, models, flower initiation, leaf initiation, growth, developmental genetics. In this review we describe how concepts of shoot apical meristem function have developed over time. The role of the scientist is emphasized, as proposer, receiver and evaluator of ideas about the shoot apical meristem. Models have become increasingly popular over the last 250 years, and we consider their role. They provide valuable grounding for the development of hypotheses, but in addition they have a strong human element and their uptake relies on various degrees of persuasion. The most influential models are probably those that most data support, consolidating them as an insight into reality; but they also work by altering how we see meristems, redirecting us to influence the data we collect and the questions we consider meaningful. © New Phytologist (2003) 159: 37–52 I. Introduction During embryogenesis certain cells gain the potential to produce the roots and shoots of a plant. These zones of cells have become known as the root and shoot apical meristems. The latter, often after a period of dormancy in the seed, functions to produce the entire above-ground structure of the plant. What we know about this process, and how we now perceive the shoot © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com apical meristem is a consequence of a long history of research, through which has emerged a number of key models, influential to many but traceable back to a dominant few. The many facets of the shoot apical meristem provide tremendous scope for different interpretations. Below, in Fig. 1, are four views of the shoot apical meristem. Notice what aspects each view emphasizes: in the transverse section, the clear phyllotactic arrangement and the organ initiation function 37 38 Review Tansley review Fig. 1 Views of the shoot apical meristem. (a) Transverse section of the terminal bud of Ligustrum vulgare (1, shoot apex; 2, youngest leaf primordia; 3, older leaves). (b) Longitudinal section of the shoot apical meristem of Zea mays (1, shoot apex; 2, new leaf; 3, previous leaf; 4, primary thickening meristem; 5, procambial strand). (c) Transmission electron microscope image of Linum usitatissimum (1, stoma; 2, bud). (d) Top view of the shoot tip of Aeonium. Reprinted from Bowes (1996), with permission from T. Norman Tait. are of apparent; in the longitudinal section, the cellular organization and outgrowth of primordia at the periphery can be seen; by scanning electron microscopy the shoot architecture is visible; and in the living plant, the lateral outgrowths from the meristem and the integration of growth and development over time is shown. How would you describe how this system works? What or who influences you in your description? In nearly 250 years of shoot apical meristem research, many would probably agree with the experience of being ‘thrilled by the translucent, glistening beauty of the apical meristem and surrounding leaf primordia’ described by Sussex (1998). From this perspective, driven by an enthusiasm to find out how the shoot apical meristem works, it is perhaps hard to conceive that very different perceptions of it could exist. Indeed, it might be assumed that progress to today’s understanding of the genetic principles underlying shoot apical meristem structure and function would have been quicker, but for technological or resource constraints. Yet this is only partly correct. The recognition that the paths we are now on were not always taken at the first opportunity shows the importance of individual judgements, and that arrival at today’s models of genetic determinism was perhaps not inevitable. Goebel (1926) commented that, ‘it is not the facts thereby attained but the conclusions drawn from them that determine the progress of science. These conclusions are influenced not only by each particular investigator’s individuality but by the general posture of science in his day.’ The purpose of this review is to chart the prominent people, from Wolff in the 1700s to Weigel now, who have influenced the way we view meristems, and to describe their approaches and contributions to current understanding of the shoot apical meristem. We have had to be somewhat selective in our treatment of the topic; for comprehensive reviews of various aspects of the shoot apical meristem readers are referred to Adler et al. (1997), Jean (1994) and Lyndon (1994, 1998). II. How things began In 1759, in his mid 20s, at a time when the microscope was still relatively underdeveloped and plant anatomy was failing to command great attention, Caspar Wolff published Fig. 2 The punctum vegetationis. Diagram by Caspar Wolff ( Wolff, 1896), which he described as follows, ‘The tip was peeled and all leaves were removed from the front view, so that it is possible to see the vegetation point. Leaves were not removed from the back to demonstrate their attachment to the vegetation surface. (v) The convex, juicy and translucent vegetation surface. (p) The first leaf to appear, with its concave inner surface adjacent to the vegetation surface. The consistency of this leaf is barely more substantial than a viscous fluid. (a) A different leaf, which is larger and more substantial than the previous. (c) A leaf which has already developed a surrounding edge. (e) Half a leaf. (d) Complete leaf.’ ‘Theoria Generationis’ (Wolff, 1896). The work had little influence then and for some time afterwards; its later revival, according to Sachs, resulted from Wolff ’s thoughtfulness rather than his accuracy (Sachs, 1890). The observations led Goebel to declare Wolff the ‘true founder of the history of development’ (Goebel, 1926). For it was Wolff who discovered the growing point of the plant, the ‘punctum vegetationis’ as he named it (Fig. 2). Here, he claimed, was evidence that growth of the plant proceeded by a process of epigenesis, that is, construction de novo, and not by ‘preformation’, whereby the mature plant resulted simply from the unfolding of preformed structures. He recognized too that leaf and flower primordia were derived by similar processes leading to the suggestion, many years before it was made by Goethe, that floral organs were modified leaves (Singer, 1931). www.newphytologist.com © New Phytologist (2003) 159: 37 – 52 Tansley review That Wolff made this proposal, we can surmise, was the result of a combination of factors: he had a question he wanted answered (epigenesis vs preformation), patience and observational skill, and suitable equipment (the microscope). In this way the most essential of plant structures was revealed, Review together with its mode of action (epigenesis) and, by implication, the ‘open’ form of development of plants in contrast to the ‘closed’ development of animals. The punctum vegetationis did not, however, become a clear focus of research at once. Thus in 1842, Schleiden could suggest, presumably with a sense of originality, that the study of ‘developmental history’ as opposed to complete structures, might be the key to morphology (Goebel, 1926). The approach to phyllotaxis further illustrates this reluctance to view plants as dynamic entities. Despite dating back to the time of Pliny, morphological explanations of phyllotaxis that involved the apex and primordia were not put forward until the 1860s at the earliest (Adler et al., 1997). Before this time studies were made of the mature, complete leaf arrangement. Transverse sections through the shoot apical meristem to reveal the organization of the primordia were an innovation of the early 20th century (see Church, below). III. Cytology The term ‘meristem’ appears not to have entered botanical vocabulary until 1858. It is attributed to Nageli who in his work ‘Beitrage zur wissenschaftichen Botanik’ classified tissues as ‘generating’ or ‘permanent’, according to their morphology. Parenchymatous generating tissue was termed ‘primary meristem’ (Sachs, 1890). The application of this term implies a recognition of function because ‘meristem’ is from the Greek word ‘merizein’, meaning ‘to divide’. The early published views of the shoot apical meristem were largely observational, aiming to give accurate descriptions of meristems under natural, nonexperimental conditions (Sifton, 1944). Nageli’s work centred on the apical cell, which he found in algae and moss, and its power to divide continuously. At the time it was assumed that this large, dividing cell was a feature of all plants, and Nageli believed that the sequence of cell divisions was of great importance in determining the form of the plant (Reed, 1942). Whilst the apical cell concept remains important today in the study of bryophytes and pteridophytes, in 1868 Hanstein redirected attention to cell layers, Fig. 3 Cytological approaches to the shoot apical meristem. (a) Hanstein: histogenic layers. ep, epidermis; ec, subepidermis; cc, central core; d, dermatogen initial layer; pe, periblem initial layer; pl, plerome initial layer. (b) Schmidt: tunica-corpus. t, tunica; c, corpus. (c) Foster: cytohistological zones. zia, apical initial zone; cmc, central mother cells; zp, peripheral zone; mc, central meristem (rib zone). (d) Buvat: méristème d’attente. mm, méristème médullaire; ma, méristème d’attente; pmsp, proméristème sporogène; pmr, proméristème réceptaculaire. (e) Comparison of terminology. Composed from Majumdar (1942) and Buvat (1952). Diagrams reprinted from Annales des Sciences Naturelles (Botanique) 8: Buvat, Structure, évolution et fonctionnement du méristème apical de quelques dicotylédons. pp. 199 –300, copyright (1952), with permission from Masson, Paris. © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com 39 40 Review Tansley review with his histogenic layer theory of the shoot apical meristem (Fig. 3a,e). On the basis of cell layers he found and studied in angiosperms, Hanstein divided the meristem into dermatogen, periblem and plerome, from which layer initials were derived the epidermis, cortex and stele, respectively (Foster, 1939; Reed, 1942). In Hanstein’s thinking there is an implicit idea of ‘destiny’, certain cells being predetermined to differentiate in a manner commensurate with their layer. Echoes of these ‘prospective values’ (Foster, 1939) can be seen in later clonal analysis of cell layer behaviour. Satina’s colchicine-induced chimeras of Datura (Satina et al., 1940) provided evidence supporting Hanstein’s histogen concept, but some redefinition was required (see Fig. 4a,b,c). In his work on cranberry chimeras however, Dermen (1947) found that there was no reason to support these inherent functions of the cell layers (Fig. 4d,e). Schmidt in 1924 followed a somewhat different approach. Based purely on patterns of growth and cell division, his model defined the overlying tunica layer(s) by their anticlinal divisions, with more variable division planes found in the corpus below (Fig. 3b; Foster, 1939; Steeves & Sussex, 1989). Dermen (1947) considered that the tunica-corpus essentially lacked meaning but Sifton (1944) believed that adherents of the tunica-corpus theory were liberated from the limitations that consideration of destiny imposed. In 1938 Foster made a much broader application of cell division analysis (Foster, 1938). Like Schmidt’s tunica-corpus layers, Foster’s was a largely structural model, but it was more extensive, dividing the meristem into cytohistological zones (apical initial zone, central mother cell zone, transition, peripheral and rib zones) (Fig. 3c). This recognized the array of cellular features that Foster found. Foster used a gymnosperm, Gingko, for this original work, but the concept was found to be applicable to angiosperms (Majumdar, 1942). Clowes (1961) criticized the imprecision of zonation models Fig. 4 Chimeras. (a) (b) and (c) Colchicine-induced periclinal chimeras of Datura. Below each diagram the chromosome number of the cell layers is given. Each layer had an independent response to colchicine treatment. The authors recognized a 2-layered tunica, plus corpus. The 3 germ layers proposed by Hanstein were evident, but the constancy of chromosome number of layer 3 and the central core implied that ‘plerome’ in Datura is not an independent tissue but a derivative of L3. Reprinted from American Journal of Botany, 27: Satina et al. Demonstration of the three germ layers in the shoot apex of Datura by means of induced polyploidy in periclinal chimeras. pp. 895–905, copyright (1940), with permission from the Botanical Society of America. (d) Longitudinal section through the shoot apical meristem of a cranberry chimera. The first 3 layers, at least, are diploid and the inner layers, tetraploid. (e) Transverse section of the stem of the plant shown in D. The dark black line marks the boundary between the inner tetraploid area and outer diploid zone. Reprinted from American Journal of Botany, 34: Dermen, Periclinal cytochimeras and histogenesis in cranberry. pp. 32–34, copyright (1947), with permission from the Botanical Society of America. www.newphytologist.com © New Phytologist (2003) 159: 37 – 52 Tansley review saying, ‘… it depends so much on the pattern which catches the eye, and the pattern is made of many different visual impressions’. Yet the terms derived from Foster’s work – central zone (CZ), peripheral zone (PZ) and rib zone (RZ), are now in regular usage as locational reference points in the meristem. What is the basis of that revival today? Clark (1996) suggests that you could ‘superimpose’ these regions with those of the central undifferentiated cell pool and its surrounding cells heading for differentiation and organ formation. Hence, although Clark admits to a lack of evidence for this interpretation, a functional significance fortifies a histological model. Furthermore, it takes on a molecular significance with the suggestion that the genes WUSCHEL (WUS) CLAVATA (CLV), SHOOTMERISTEMLESS (STM) are involved in delimiting/maintaining the CZ and PZ regions (see Developmental genetics, below; Clark, 1996; Weigel & Clark, 1996). With the exception of Hanstein, concepts of the shoot apical meristem based on cell division patterns were until the 1940s generally structurally based. Few attempts were made to ascertain the functional significance of cell division rates or patterns. This makes the méristème d’attente theory of Buvat (1952) something of a turning point. To Buvat, the infrequency of cell divisions in the central region of the meristem was of great significance. It was this area he named the méristème d’attente, a zone of cells he envisaged waiting, inactively, until the initiation of floral activity (Fig. 3d,e; Buvat, 1952; Steeves & Sussex, 1989). Until flowering, the focus of vegetative development of the plant was the peripheral region, the ‘anneau initial’. In arriving at this model, Buvat defined the ‘French School’ (Fig. 5; Wardlaw, 1957; Cutter, 1959). The theories of Majumdar (1942) and Plantefol (1946) were acknowledged by the attention given to the peripheral regions of the meristem. Majumdar had seen the apex as a ‘self-perpetuating group of central initial cells, surrounded by a cylinder of more active flank meristem from which the primordia originate’ (Majumdar, 1942). This idea was reinforced in Plantefol’s phyllotaxis theory (see Morphology below) in which leaves Fig. 5 The French School. © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com Review arise from the ‘anneau initial’ incorporated into Buvat’s theory. There would appear also to be some influence of Grègoire’s ‘radical viewpoint’ (Foster, 1939) which strayed far from the classical interpretation of the flower, by suggesting that vegetative and floral meristems bore no relation to each other. To Grègoire, whilst tunica and corpus were recognizable in vegetative meristems, the floral meristem was defined as a ‘manchon meristematique’, a cloak or mantle of meristematic tissue overlying the ‘massif parenchymateux’, a region of highly vacuolated infrequently dividing cells (Foster, 1939; Philipson, 1949; Buvat, 1952). This perception of the floral meristem as independent, rather than a derivative of the vegetative meristem, is reflected in Buvat’s delimitation of vegetative and floral zones in the meristem. Beyond the French School, criticized by Wardlaw (1957) for observations ‘somewhat selective in character’, the méristème d’attente theory was a view not shared by many. Nevertheless, the theory did galvanise enquiry into cell behaviour in the meristem. This was in spite of differences of interpretation and crucially, the limited evidence for the inactivity required of the waiting cells in the meristem summit (Wardlaw, 1957; Cutter, 1959; Steeves & Sussex, 1989). IV. Morphology Morphological approaches to the shoot apical meristem are essentially concerned with how and where the meristem establishes growth. Featuring strongly in this aspect of shoot apical meristem research were the Snows who recognized that models of phyllotaxis until that time (1931) fell into two groups – those that proposed pattern to be determined by an unknown property of the stem itself, and those that advocated leaf arrangement to be dictated by contact with existing primordia. The Snows set about testing the validity of the models – assuming that if those in the second group were correct, manipulating primordia as they arose could influence the positions of those that formed later. Where others had observed and calculated (for review see Schwabe, 1984), the Snows became experimentally involved using a ‘cataract knife’ and a dissecting microscope to perform microsurgery on the shoot apical meristem. The Snows observed that, under certain circumstances isolating a primordium or its site caused the phyllotactic spiral to be reversed with a primordium and its successors forming at opposite sides to those expected, had normal phyllotaxy been retained (Fig. 6a,b). This led the Snows to propose a concept of ‘first available space.’ Existing primordia determined where a new primordium formed – and this was in the widest gap, furthest from the growing apex (Snow & Snow, 1931). The Snows also pioneered the study of how a leaf forms. They found that application of auxin to the shoot apical meristem of Lupinus albus and Epilobium hirsutum gave rise to enlarged, united primordia in an altered arrangement and interpreted these results to support their model of phyllotaxis 41 42 Review Tansley review Fig. 6 The Snows and Wardlaw – first available space/inhibitory fields. (a) and (b) Diagrams of transverse sections of the shoot meristem of Lupinus albus from Snow & Snow (1931). The straight line across (a), a normal bud, marks the position of the incision made to isolate the area of tissue (I1 region), about to initiate the next primordium. The phyllotactic alteration that this causes is shown in (b). Note the different positions of I4 and I5 after incision (b). P1, P2, P3, etc. = existing primordia, in order of increasing age. I1, I2, etc. = incipient primordia, invisible at the time of operation, in order of appearance. Reprinted from Philosophical Transactions of the Royal Society, London, Series B 221: Snow M, Snow R. Experiments on phyllotaxis. I. The effect of isolating a primordium. pp. 1 – 43, copyright (1931), with permission from The Royal Society. (c) and (d) Representations of the shoot apex of Dryopteris showing, in (c), the position of leaf primordia and incipient primordia, and in (d), the possible ‘fields’ associated with these ‘growth centres’. ac, apical cell; b, bud rudiments; m-m1, lower limits of apical meristem. The dashed line indicates the approximate base of the apical cone. I1, I2, etc., as above. Numbers = existing primordia (increasing age). Reprinted from Growth (supplement) 13: Wardlaw, Experiments on organogenesis in ferns. pp. 93–131, copyright (1949), with permission from Growth Publishing Co. Inc. (Snow & Snow, 1937). To an extent this work foreshadows contemporary morphological approaches (see below). The 1940s brought two influential models of phyllotaxis. The first was Plantefol’s ‘multiple foliar helices’ model which is rarely considered now but was a very significant feature of the French School (Fig. 7; Wardlaw, 1957; Cutter, 1959). In this model, leaves were arranged in helices of variable numbers depending on the plant. Helices terminated in leaf-generating centres in the anneau initial and these were under the control of an organizer (Plantefol, 1946; Cutter, 1959). This theory has not gained wide acceptance because it is not clear how to determine which foliar helices have most ‘biological reality.’ Neither has it been given rigorous mathematical testing. In the second of the post-Snow models, we return to their surgical approach, the knife this time wielded by Wardlaw. He favoured the large meristems and widely spaced primordia of the fern Dryopteris, and using a similar technique arrived at similar results to those of the Snows. He did not reject the Snows’ ‘first available space’ hypothesis but instead set about interpreting ‘space’ along the lines of Schoute in 1913, drawing inhibitory fields across the meristem (Fig. 6c,d). Wardlaw argued that space itself does not ‘do’ anything and is not a ‘causal factor’ in morphogenesis. A new primordium emerged at the position of weakest inhibition with the most recently formed primordium the inhibitor source (Wardlaw, 1949). The view that phyllotactic patterns can be explained by inhibitory fields is an enduring one. Richards (1951) sought a phyllotactic description that completely defined the arrangement of leaves and, most significantly, was independent of any theory of the origin of phyllotactic pattern. He achieved this using the divergence angle and the plastochron ratio (the radial distances of successively initiated primordia from the centre of the system). Richards clearly saw the importance (and limitations) of treatments by Schimper and Braun more than 100 years earlier, in which phyllotaxis was partly defined by the divergence between two successive leaves in a tangential direction; Church’s classification by pairs of orthogonally intersecting contact www.newphytologist.com © New Phytologist (2003) 159: 37 – 52 Tansley review Review shoot apical meristem. He wanted to explain the origin of pattern as well as its further propagation (Green et al., 1996), and he recognized that initiation and patterning mechanisms were not necessarily divorced from each other (Green, 1992). Perhaps most significantly, Green suggested that concepts of pattern formation need not be anticipatory (i.e. that no regional differences need precede and direct development), a contrasting view to that which predominates in developmental genetics today. Whilst Meyerowitz has advocated that ‘global patterning’ arises out of the coordination of local control mechanisms (Meyerowitz, 1996), Green considered this approach to be anticipatory and favoured a view of development which was reflective of global pattern; so for example, in some cases, ‘a tissue-level physical process appears to be upstream of organ-specific expression’ (Green, 1997). Green proposed a biophysical solution to shoot apical meristem pattern formation, similar to that which gives ‘wrinkles in wet skin’ (Hernandez & Green, 1993); this was the physical buckling theory. Patterning in this model arose from instability, in turn arising from the ‘need to expand’ of a uniform sheet tied to a uniform underlying layer which gave upward pressure. These attributes were provided by the tunica and corpus of the meristem (Fig. 8). To redress this unbalanced state, physical buckling would occur and generate bulges on the meristem surface (Green, 1992; Hernandez & Green, Fig. 7 Plantefol: multiple foliar helices. (a) Stem of Lilium candidum showing three foliar helices. (b) The classical view of leaf generation and the multiple foliar helices model for a stem with three foliar helices. Reprinted from Annales des Sciences Naturelles (Botanique) 7: Plantefol, Fondements d’une théorie phyllotaxique nouvelle. pp. 1–77, copyright (1946), with permission from Masson, Paris. parastichies (Church, 1904, 1920); and van Iterson’s studies of packing of spheres on cylinders. Although Richards’ treatment of phyllotaxis did not depend on any particular mechanism, it was consistent with the inhibitor theory adopted by Wardlaw. Later mathematical models simulating the growing shoot apex by, for example, Thornley (1975a,b), and Veen & Lindenmayer (1977) used the idea that primordia (and the bare apex) are sources of an inhibitor of primordium initiation (see Schwabe, 1984; Jean, 1994 for reviews). These models tend to be concerned with the mechanism by which the shoot apical meristem continues to function. A more contemporary morphologist, Green, was, however, not so concerned with maintenance of existing pattern at the © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com Fig. 8 Green: undulations. (a) Wild-type floral meristem of Antirrhinum. Stamens arising in whorl 3. (b) Floral meristem of a deficiens mutant of Antirrhinum, which makes sepals in the first two whorls of the flower, followed by carpels. (c) and (d) Mechanical simulations of undulations in a fixed circular region. It is proposed that in the meristem, undulations depend on the ratio of corpus (flexible) to tunica (rigid). A shift from out-of-plane undulations, giving stamens, to in-plane undulations will give rise to carpels, mimicking the effects of the deficiens mutant. s, sepals; p, petals; st, stamens; c, carpels. Reprinted from American Journal of Botany 86: Green, Expression of pattern in plants: combining molecular and calculusbased biophysical paradigms. pp.1059–1076, copyright (1999), with permission from the Botanical Society of America. 43 44 Review Tansley review 1993; Green et al., 1996). These bumps could then provide a new set of physical parameters for continued buckling to propagate pattern (Green, 1992). A nice example of the process of leaf initiation and the subsequent organization of a shoot apical meristem is provided by the work of Selker & Lyndon (1996) with axillary explants of watercress (Nasturtium officinale). Roughly triangular surfaces, showing none of the characteristic structural organization of a meristem, were able to initiate new buds. The phyllotactic pattern of these explant buds was sometimes different to that of the parent plant. The authors interpret their observations as showing that the pattern is related to the shape of the bud-forming space and mechanical stresses within it (Selker & Lyndon, 1996; Lyndon, 1998). Green perceived that in the shoot apical meristem pattern was generated from ‘nothing’ (Hernandez & Green, 1993) and he saw this model as preferable to the more commonly applied positional information and reaction-diffusion genre, both of which require an anticipatory prepattern for which he found very little evidence (Green, 1992; Hernandez & Green, 1993). Nor under the physical buckling mechanism was there a need for inhibitory fields to account for phyllotaxis. Green suggested that there was no reason to believe that organ initiation would be spontaneous unless inhibited (Green, 1992). In Green’s model ‘inhibition’ derives from the ‘intrinsic reluctance’ of the surface to undergo a sharp change in shape (Green et al., 1996). To a degree Green’s work was carried out on inanimate models, simulations which gave phenocopies of plant mutants and fused organs (Green, 1996). Applying the principles in vivo, growing sunflower heads in constraints resulted not only in predictable alterations in pattern but also in altered organ identity (Hernandez & Green, 1993). Green interpreted this as ‘abnormal buckling’, that is, a physical process having an influence on gene expression (Hernandez & Green, 1993; Green, 1994). Overall, Green’s approach to meristems was strikingly original; this is apparent not least in the vocabulary he used to describe them. He placed emphasis on shape and form, writing of ‘tissue undulations’, ‘an initial bump pattern’ which is ‘roughly sinusoidal’ and primordia ‘delimited by parallel creases of equivalent clarity and depth’ (Hernandez & Green, 1993). Green’s work spanned a time when morphology was, to some extent, giving way to the emerging field of developmental genetics. Perhaps partly because of this, his work on pattern initiation was founded on almost uniquely held principles. Modern-day morphologists have much in common with the Snows in attempting to discover the workings of the shoot apical meristem through its manipulation. Reinhardt applied a similar technique to the Snows – the application of synthetic indole-3-acetic acid (IAA), but in combination with plant material in which auxin transport had been inhibited, chemically (with N-1-naphthylphthalamic acid (NPA) in tomato shoot apical meristems) or genetically (the pin-formed 1-1 (pin1-1) mutant of Arabidopsis) (Reinhardt et al., 2000). Tomato shoot apical meristems on NPA media failed to produce leaves, but leaf initiation could be restored by auxin. Similarly auxin application recovered flower formation in pin 1-1 mutants. The finding that leaves always appeared at the precise point of auxin application in the radial axis of the shoot apical meristem but at a constant distance from the meristem tip led Reinhardt to propose a model of shoot apical meristem function in which the apical-basal axis, Foster’s CZ/ PZ, is maintained by genes such as CLV, WUS, STM (see Developmental Genetics, below) but that radial patterning is auxin dependent. Organogenesis of leaves and flowers requires auxin, and, in line with the Snows, correct positioning of leaves (but not their initiation) requires existing primordia (Reinhardt et al., 2000; Kuhlemeier & Reinhardt, 2001). Recently a more comprehensive model, linking transcription factors, hormones and primordium outgrowth has been elaborated. A new leaf differentiates where there is gibberellin and a high concentration of auxin. These conditions arise through an auxin gradient in the meristem with existing primordia acting as auxin sinks. Gibberellin levels are affected when homeodomain transcription factors such as KNAT1 and STM, which normally inhibit gibberellin biosynthesis are down-regulated. In this way transcription factors can regulate meristematic cells through growth hormones (Vogler & Kuhlemeier, 2003). At a biophysical level, expansins are proposed to operate in cell wall expansion during cell extension. Reinhardt suggested that genes such as the tomato expansin LeEXP18 could be activated to induce bulging of the meristem in the final element of a phyllotactic mechanism (Reinhardt et al., 1998). This role of expansin in leaf development is supported by the work of Fleming et al. (1997) and Pien et al. (2001). In both cases expansin in localized areas induced leaf primordia and reversed phyllotaxis. Pien et al. suggested that this was ‘evidence of cell-division independent mechanisms controlling morphogenesis’ (Pien et al., 2001). V. Developmental genetics Although there is little to suggest that cytological and morphological considerations of the shoot apical meristem are now implausible, the developmental genetics era of the last 15 years is largely independent and disconnected from these earlier phases – phases which may now sometimes appear irrelevant. Models are a significant component of this research phase, being typically the end result and the starting point for further work. In the current climate of genetic determinism there is a sense that, in molecular biology, we have the answer; that this is the time when we will define the root causes of meristem functions. The molecular models do, however, elaborate on earlier structural models, being particularly concerned with the balance between self-perpetuating and differentiating cells that underlies meristem maintenance. Attention is also given to www.newphytologist.com © New Phytologist (2003) 159: 37 – 52 Tansley review how meristems form, initiate the organs characteristic of vegetative or floral phases, and terminate. Barton has considered the origin of leaves and of the shoot apical meristem itself. These are processes that require tissue differentiation, to distinguish the shoot apical meristem from other embryonic tissues, and, later to define leaf founder cells from amongst the undifferentiated cells of the shoot apical meristem. Barton’s research addresses patterning arising in the embryo and ‘genetic circuitry that distinguishes meristem from leaf ’ (Barton, 2001). She has studied these differentiations using the homeodomain transcription factor STM in Arabidopsis. stm mutants fail to make a functional shoot apical meristem. The gene is expressed in the late globular stage of the embryo and then in the shoot apical meristem throughout the life of the plant, suggesting a role in the formation and maintenance of the meristem. STM is down-regulated in cells destined to become leaves (Long et al., 1996), although this is not the first sign of leaf identity as the expression of PINHEAD/ZWILLE (PNH) precedes the down-regulation of STM (Lynn et al., 1999). Barton revived an earlier term, ‘promeristem’ to encompass the initials and their recent undifferentiated derivatives in the shoot apical meristem (Barton, 1998). STM and PNH are effectively markers for the outcome of decisions facing promeristem cells – to continue in a meristematic role or to become leaf. Barton proposes a model of cyclic action. In this model the shoot apical meristem initiates leaves, then the adaxial side of these leaves induces shoot apical meristem formation in the leaf axil – so shoot apical meristems make leaves of which make meristems (McConnell & Barton, 1998). In arriving at this model, Barton considered the correlation that exists between adaxial leaf tissue and the formation of meristems. This relationship is seen in phabulosa mutants of Arabidopsis, in which leaves are adaxialized, in overexpression studies with the maize homeodomain KNOX transcription factors and also in earlier work by the Snows, in which surgery resulting in leaf abaxialization was accompanied by a loss of axillary meristems. A significant concern of this model is whether shoot meristem tissue arises only once, as the original embryonic assignment, or many times over in the life of the plant. Does the new axillary meristem arising in association with the adaxial side of the leaf derive from ‘detached meristem’, that is, a few meristematic cells that remain undifferentiated? Or is it possible for cells to differentiate but then to respond to a localized signal that re-establishes their meristematic properties? (McConnell & Barton, 1998). These considerations of the distinction between the leaf and the meristem are reminiscent of the thoughts of Arber (1950). Arber observed the self-similarity of the system: ‘each branch shoot echoes the character of the parent shoot’ and consolidated the earlier botanical interpretations on this theme in her view of the leaf as a ‘partial-shoot’, adding that ‘the partial-shoot has an urge towards the development of whole shoot characters’ (Arber, 1950). © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com Review Laux and Clark have focused on how the meristem sustains its two main functions, providing cells to an undifferentiated pool, and differentiating cells into organs. Laux primarily considers the Arabidopsis homeodomain transcription factor, WUS (Mayer et al., 1998) and Clark investigates the role of the signal transduction pathway encoded by the CLAVATA genes. CLAVATA1 encodes a receptor kinase, CLAVATA2 a receptor-like protein and CLAVATA3 a peptide ligand (Clark et al., 1997; Jeong et al., 1999; Trotochaud et al., 2000). The involvement of these genes in meristem maintenance can be inferred from their mutant phenotypes. The meristems of wus mutants terminate prematurely (Laux et al., 1996), whilst those of clavata mutants proliferate (Clark et al., 1993, reviewed in Clark, 1996). The model of meristem function which emerges from mutant, expression and overexpression analyses uses the CZ/PZ terminology of Foster to provide grid references, within which gene expression patterns and cell functions can be defined (Fig. 9). Laux’s group picture the meristem as including an organizing centre defined by WUS expression. This organizing centre lets overlying cells know that they are stem cells by inducing CLV3 expression, which maintains the cells in an undifferentiated state. To become organs, cells need to be beyond the range of WUS, thus losing their stem cell identity and becoming available to be assigned organ fate (Schoof et al., 2000). Although WUS induces CLV3 expression, CLV3 is secreted from the outer layers to the lower layers of the meristem where the CLV1 and 2 genes are responsible for curtailing the expression of WUS. It is this WUS –CLV negative feedback interaction which is proposed to maintain the balance of cells in the meristem (Schoof et al., 2000). Revealingly, Laux’s group have expressed WUS under the control of the promoter of the AINTEGUMENTA (ANT ) gene, thereby directing its expression to organ primordia. The resulting misexpression of WUS extends CLV3 induction and undifferentiated cell fate to a tissue at an early stage Fig. 9 The molecular basis of shoot apical meristem maintenance. WUS expression in a subset of cells in the central region of the meristem establishes an ‘organizing centre’ and activates CLV3 expression in the overlying outer cell layers of the meristem. CLV3 negatively regulates the expression of WUS via its interaction with CLV1. These interactions between WUS and the CLV genes, regulate the proliferation of stem cells in the meristem. STM is expressed throughout the meristem where it represses cell differentiation. It is down-regulated in organ primordia (see text for details). CZ, central zone; PZ, peripheral zone. 45 46 Review Tansley review of differentiation (Schoof et al., 2000). Thus, WUS can apparently respecify relatively undifferentiated tissue, such as organ primordia, and initiate stem cell identity there. More differentiated cells, however, require the action of both STM and WUS to reverse their fate (Lenhard et al., 2002). These genes appear to function independently, regulating different downstream targets. Their common task is to prevent differentiation, either through its suppression (STM) or by the specification of stem cells (WUS). Lenhard et al. (2002) found that coexpression of STM and WUS led to an extension of small CLV3expressing cells from the apex onto the lamina of cotyledons. Gallois et al. (2002) induced WUS expressing sectors in seedlings and independently activated STM, with the result that outgrowths occurred from both abaxial and adaxial sides of the cotyledons. These outgrowths expressed CLV1 transiently but then developed as leaf-like organs, suggesting that they had a meristem-like phase but ultimately were not selfmaintaining structures. Thus, the details of WUS / CLV/STM action still need to be worked out, particularly when the genes are expressed in pre-existing tissues in different stages of differentiation. In the era of developmental genetics the key question of how leaf primordia are positioned has received relatively little attention. Hake has described two mutants of maize which provide possible support for Wardlaw’s inhibitory field model. In the terminal ear 1 (te1) mutant alternate phyllotaxis is disrupted to an irregularly opposite or spiral arrangement (Veit et al., 1998); in the abphyl1 mutant it becomes decussate (Jackson & Hake, 1999). The expression of TE1, a putative RNA-binding protein, was found in ‘horseshoe’ formations alternating from side to side around the stem. New leaf primordia arose at the discontinuity of expression, leading Veit et al. to suggest that TE1 inhibited cells from ‘acting as organizers of leaf development’ and Scanlon to wonder whether TE1 might ‘divulge the first molecular evidence for the inhibitory field theory of plant phyllotaxy’ (Scanlon, 1998). Crucial to the interpretation of abphyl1 is the observation of the enlargement of the embryonic shoot apical meristem, and that the novel phyllotactic pattern is established in the embryo. This implies that, depending on the precise timing of leaf position determination, phyllotactic patterning may follow the change in meristem size and not vice-versa ( Jackson & Hake, 1999), a finding consistent with the model proposed by Richards (1951). Jackson and Hake envisage that the enlarged meristem affects fields – all inhibitory – but either chemical or biophysical. Up to this point we have considered how differentiated and undifferentiated tissues of the meristem are specified and maintained. This is a constant underlying feature of the meristem, even when alterations to meristem function occur in accordance with the growth phase of the plant. Mechanisms that elaborate on this basic pattern during the floral phase have been the subject of many models, which will now be considered. Coen’s view of the shoot apical meristem is unitary (i.e. the meristem is divisible into a set of simple, self-contained regions and functions), reductionist and highly conceptualized. He suggests that meristems have an ‘identity’ – vegetative, inflorescence or floral. This identity is controlled by ‘meristem identity genes’ and can be described and classified in terms of four main features, phyllotaxy, organ identity, determinacy and internode length (Coen, 1991; Coen & Carpenter, 1993). Consequently, in interpreting the floricaula ( flo) mutant of Antirrhinum majus, Coen proposes that the loss of FLO gene expression results in a failure of the transition of the meristem from inflorescence to floral identity (Fig. 10a; Coen et al., 1990). Hence FLO is a meristem identity gene orchestrating a switch in the crucial meristem properties that compose ‘identity’. The conceptual nature of this picture is illustrated by the work of Huala & Sussex (1993). Drawing largely on predevelopmental genetics work, they described cases in which meristems may be considered determined but their derivatives may not. For example, a maize meristem may be reproductively determined yet still initiate primordia which develop as vegetative shoots (Huala & Sussex, 1993). In such cases assessment of meristem identity must rely on characteristics of phyllotaxy, determinacy and internode length (Huala & Sussex, 1993). Fig. 10 Coen: FLORICAULA and partitioning. (a) Longitudinal section of an inflorescence of Antirrhinum majus, viewed in dark field. The section shows FLO expression in the meristem detected using in situ hybridization with digoxigenin-labelled probes (1990). Reprinted from Cell 63, Coen et al. FLORICAULA: a homeotic gene required for flower development in Antirrhinum majus. pp. 1311–1322, copyright (1990), with permission from Elsevier. (b) Top view of a wild type meristem of Antirrhinum majus, showing a regular initiation pattern of discrete primordia. The outline of a double helix is marked. (c) flo mutant of Antirrhinum majus lacking discrete primordia. Reprinted from Plant Cell 7, Carpenter et al. Control of flower development and phyllotaxy by meristem identity genes in Antirrhinum. pp. 2001–11, copyright (1995), with permission from the American Society of Plant Biologists. www.newphytologist.com © New Phytologist (2003) 159: 37 – 52 Tansley review For Coen a unifying factor underlying a number of these meristem properties is the idea of partitioning. Cells become partitioned off from the shoot apical meristem to become organ primordia or secondary meristems, and which of these two types of partitioning is favoured dictates determinacy. Conforming to a phyllotactic arrangement requires a precise pattern of partitioning (Coen & Carpenter, 1993). This concept was elaborated further in one of the first molecular approaches to phyllotaxis: the analysis of phyllotaxis of the meristem identity gene mutants of FLORICAULA (FLO) and SQUAMOSA (SQUA) (Carpenter et al., 1995). In both mutants phyllotaxis is altered from that of the wild type, implicating FLO and SQUA in promoting the transition from spiral to whorled phyllotaxis. This change is considered to be controlled by these genes in a two-stage process. Areas of the meristem are set aside for ‘potential primordium initiation’ in stage 1, and then partitioning of these areas in stage 2 results in primordia. Rarely, in flo mutants, this second stage fails and plants form double spirals, which led Carpenter et al. to question whether the primary initiation state is of discrete primordia (which unite in flo mutants) or of spirals which require partitioning. This second proposal is recognized as identifying with the ideas of Plantefol (Fig. 10b,c). Coen proposes that the nature of the organs which a meristem of floral identity initiates is controlled by ‘organ identity genes’ (Coen & Carpenter, 1993). The model of organ identity gene action, the ABC model, devised by Coen and Meyerowitz (1991, see below) is a current, highly influential concept of flower development. Coen’s interest in this area arose from a PhD on Drosophila, during which he became interested in studying development and evolution from genetic and molecular perspectives (Coen, 1996). By analyzing Antirrhinum flower structure mutants, Coen began to construct a genetic model, with first two (independent), then three gene functions to explain flower structure (Carpenter & Coen, 1990; Coen, 1991). The resulting ABC model proposes that organ identity genes operating in three overlapping domains, A, B and C control the identity of organs in the flower. a function expressed alone in whorl 1 gives rise to sepals; a and b in whorl 2 to petals; b and c in whorl 3 to stamens; c in whorl 4 to carpels. a and c are antagonistic and cannot be expressed in the same domain (Fig. 11a,b; Coen & Meyerowitz, 1991; Coen, 1991; Coen & Carpenter, 1993). For Huala & Sussex (1993), floral homeotic mutants demonstrate that determination of the floral meristem and its primordia are separable events (see above). It is also evident that, whilst mutation of organ identity genes alters identity, other floral properties (e.g. phyllotaxy, lack of internodes) persist. This narrow role for organ identity genes was apparent to Lord, who proposed the importance of relative timing in meristem functions, and emphasized the heterochronic as well as homeotic nature of floral mutants. One of the conclusions of her work was that whilst organ identity was altered, the characteristic cell division patterns that gave rise to the primordia © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com Review Fig. 11 Coen and Meyerowitz: the ABC model. (a) The ABC model of floral organ identity. In this diagrammatic representation the whorls of the flower are shown as concentric rings. Superimposed on this are the domains A, B and C. See text for details. Redrawn from Coen & Meyerowitz (1991). How this is proposed to translate to the meristem is shown in (b), a diagram of the meristem in longitudinal section, showing patterns of ABC expression in present or presumptive whorls 1–4. Reprinted from Plant Cell 5, Coen and Carpenter, The metamorphosis of flowers. pp. 1175–1181, copyright (1993), with permission from the American Society of Plant Biologists. in each whorl remained as wild type (Lord et al., 1994; Hill & Lord, 1989; Crone & Lord, 1994). Weigel has also considered the mechanisms which underlie the correct positions of ABC expression patterns, concluding that meristems have patterning systems common to vegetative and floral meristems, which involve the genes UNUSUAL FLORAL ORGANS and WUSCHEL (Parcy et al., 1998; Lohmann et al., 2001). Coen introduced a second dimension to complete the ‘polar coordinate model’ for the control of primordium fate (Coen, 1991). As well as the ‘r’ or radial axis, along which organ identity varies in the flower, there is the ‘y’ axis. Along this second axis, the expression of the gene CYCLOIDEA is proposed to vary – and control the asymmetry (where relevant) of the flower. It achieves this, it is proposed, by slowing the growth rate and reducing the organ number in the area of its expression (Luo et al., 1995). According to this model, the fate of a primordium is the reflection of its coordinates on the identity (r) and symmetry (y) axes (Coen & Meyerowitz, 1991; Coen, 1991). What is striking about these models of meristem function is that they rely on the ideas of identity and symmetry as specific end points to which genetic cascades are directed. As Meyerowitz, coproposer of the ABC model, points out, there are few, if any, reasons why a model of flower development along these lines could not have been arrived at 100 years earlier. Detailed descriptions of monstrous and normal flowers have been made for centuries, but, with some 47 48 Review Tansley review exceptions, it was not until the late 1980s that the study of flower development capitalized on the availability of flower abnormalities (Meyerowitz et al., 1989; Meyerowitz, 1995). The aim of these approaches was to ‘understand the flow of information from the genome that led to the 3-dimensional structure of the flower’ (Meyerowitz, 1995). For Meyerowitz this began with the description of recessive homeotic mutants of Arabidopsis thaliana. He established that the mutations in question were often flower-specific, the organs developed on a time course linked to their whorl rather than their identity, that replacement organs in the mutants were often mosaic in nature, and there were few constraints on the identity of organs in any whorl. This last discovery was evidence against the few earlier mechanistic models there had been, which proposed sequential means of organ specification, and were at odds with the data on abnormalities (Bowman et al., 1989; Meyerowitz et al., 1989). Consideration of the mutant phenotypes led Meyerowitz to the proposal that the flower primordium could be ‘divided into fields or compartments, each consisting of adjacent whorls’ with the genes acting in the ‘delineation of concentric-ring shaped compartments, each with a different fate’ (Bowman et al., 1989). The preliminary model proposed three classes of homeotic gene capable of producing a flower, with the leaf as a ‘ground state organ’ (Coen & Meyerowitz, 1991; Meyerowitz et al., 1991). Later tests of the model revealed the necessity for a further function, provided by the MADS-box SEPALLATA genes, to convert leaves to floral organs (Honma & Goto, 2001). Theissen (2001) considered this grounds to re-evaluate the ‘outmoded ABC model’, by then amounting to the ABCDE model, questioning whether ‘floral homeotic functions are still a useful concept at all’ since the concept ‘no longer provides a useful simplification’ (Theissen, 2001). In Coen’s opinion, we need now to think of the basis of primordium fate in genetic terms and consider, ‘how are the domains of whorl identity genes established?’ (Coen, 1991). Seen in relation to the cytological and morphological approaches outlined in earlier sections, this indicates a huge shift in perspective. The view of the meristem here is abstract, a recurrent feature of developmental genetics-based meristem models. What are the relative merits of this direction in meristem research and how have these new ideas come about? The ABC model is probably the most influential model of meristem function that there has been. It is simple and neat – two desirable features in any model. It is possible that the ABC model is so successful because it presents the closest explanation to the biological reality of how a flower forms. Certainly it is reassuring that, almost independently, and working on separate systems, Coen and Meyerowitz arrived at similar models. But it could be argued that this has more to do with their common perspective than with reality. The massive support found for this model shows that many choose to interpret their data in the light of it, and that they share the same perspective as Coen and Meyerowitz. A molecular model is necessarily abstract because, as Coen has pointed out, what has recently been revealed is ‘how organisms contain a whole series of regional differences that were previously hidden from view’ (Coen, 1996). But, whilst we can perceive the organization of floral organs, and in situ hybridization allows us to visualize gene expression, in these models we need to envisage partitioning, domains, ‘molecular territories’ (Coen, 1996) and boundaries (Vincent et al., 1995). This can be challenging, although similar demands have been made in the past. In Wardlaw’s proposal of phyllotactic determinants we had to visualize inhibitory fields radiating from growth centres. Interestingly, both models provide strong visual images, and it seems legitimate to ask how far they colour our perceptions. Ask yourself: ‘would I have thought like this from experience? How much is this a product of my imagination?’ The answer is that the model determines what you see, the questions you can ask; it defines a way of thought. How did we start to see plants in such unitary terms? Out of the beginnings of mutant analyses arose ‘animal-plant’ thought. Initially this was merely a recognition that the initiative to use mutants for the study of pattern formation followed a precedent set in animal developmental studies. Drawing from earlier work on Drosophila, Meyerowitz concluded that finding the genes underlying flower development was the next step (Meyerowitz et al., 1989). When the first of these genes was uncovered, the DEFICIENS A (DEF A) gene of Antirrhinum majus (Sommer et al., 1990), and the functional significance of its homology to transcription factors was considered, it was possible to envisage a regulatory network determining morphogenesis at the meristem (Sommer et al., 1990). It was, however, the report of the second transcription factor to be cloned, AGAMOUS (AG) from Arabidopsis (Yanofsky et al., 1990), that seemed to signal a turning point in perceptions of how the meristem might function. Discovery that this second gene had extensive homology to the DEF A gene prompted many thoughts in an animal-plant direction. The possibility that a family of transcription factors was at the heart of pattern formation of flowers as well as flies grew ever more likely. Meyerowitz proposed that we might consider some sort of ‘floreo’ box, similar to the homeobox of Drosophila homeotic selector genes (Meyerowitz et al., 1991) and the AG report concluded that, analogous to animal development, floral organ development might be regulated by a set of closely related genes (Yanofsky et al., 1990). Furthermore, with DEF and AG MADS box transcription factors (as they became known, Schwarz-Sommer et al., 1990) from different plant species, there was the possibility of an evolutionarily conserved mechanism. Thus the domains and compartments of meristems in today’s developmental biology are traceable to concepts of animal development. From a similarity of approach (analysis of developmental mutants) to a discovery of gene families, we have reached a similarity of perception and mechanism. Sablowski www.newphytologist.com © New Phytologist (2003) 159: 37 – 52 Tansley review Fig. 12 Meinhardt: segmentation. Schematic longitudinal section of a plant. M indicates an ‘elementary module’ composed of node (N) and internode (I), and producing a leaf (L) and bud (B). Each module is made of at least 3 subunits M1, M2, M3, which ensure polarity. A, shoot apical meristem; P, primordia. Stem shown, ‘unrolled’ in 2-dimensions. Cells produced by the shoot apical meristem aid stem elongation and differentiate into subunits. Note leaf initiation on M1, M2, border. Reprinted from International Journal of Developmental Biology 40, Meinhardt, Models of biological pattern formation: common mechanism in plant and animal development. pp. 123–134, copyright (1996), with permission from the University of the Basque Country Press. & Meyerowitz (1998) commented that ‘homeotic mutations have provided evidence for modularity in the development of both animals and plants; the identity of discrete body parts can be transformed by loss of function or ectopic expression of homeotic genes’. This concept of modularity extends beyond the meristem consumed with flowering, and also back to a pre-developmental genetics era when vegetative modules became known as phytomers. More recently Meinhardt (1996) has proposed a segmentation model of plant development (Fig. 12). Disappointed in the inability of morphological spacing models to explain how leaves develop distinct abaxial and adaxial leaf surfaces with correct orientation to the shoot apical meristem, Meinhardt proposed that segments M1, M2 and M3 form a repeating sequence to construct a vegetative plant. An activator-inhibitor mechanism is involved in determining the generation of leaves but, crucially, leaves only form at the junction of segments. The junction position ensures that leaves are composed of two tissue types, M1 and M2 (abaxial-adaxial surface) and are correctly orientated. Thus the polar character of the boundary ensures the polar structure of the leaf. Meinhardt sets this model amongst others of biological pattern formation (tropical sea shells, leg, wing initiation … ) and proposes a ‘common mechanism’ in plant and animal development. Further endorsements of such analogies come from the comparisons of leaf and wing development ( Waites & Hudson, 1995; Scanlon et al., 1996). The end of the shoot apical meristem is most frequently considered as accompanying flower formation. This determinacy © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com Review dimension to flowering has been prominent in the thinking of developmental geneticists and is associated with the C function gene AG (Yanofsky et al., 1990). ag mutants fail to make the reproductive organs of the flower and produce an indeterminate proliferation of sepals and petals (Bowman et al., 1989), whilst ectopic AG expression can convert an indeterminate inflorescence meristem to a determinate floral meristem (Mizukami & Ma, 1997). The termination of growth associated with flowering results from an interaction between AG and WUS (Lenhard et al., 2001; Lohmann et al., 2001). As described above, WUS is involved in assigning stem cell fate via CLV3. In conjunction with the meristem identify gene LEAFY it also up-regulates AG expression but, later in development, AG acts to repress WUS, thus halting the supply of undifferentiated cells and preventing further growth. Although the C function may lie at the heart of meristem determinacy, development continues after the carpels it specifies. The D function genes FBP7 and FBP11 evoke ovule and placental column development from the central meristem of Petunia (Angenent & Colombo, 1996). In some cases determinacy is only incompletely maintained (Fig. 13). When and where growth termination occurs dictates the architecture of the plant (Battey, 2003). There are times when flowering does not stop a meristem (floral reversion), and other times when meristems cease growth without flowering. One such example is found in Arabidopsis itself: the induced apical meristem is an inflorescence meristem, and although growth at this apical point is indeterminate, it isn’t infinite. Another example is found in inflorescence meristems of Bougainvillea, which under long day conditions fail to develop and become lignified thorns (Hackett & Sachs, 1968). In calabrese, apical abortion or ‘blindness’, an environmentally induced disorder, results in cells losing their meristematic ability and differentiating as parenchyma. The tunica-corpus distinctions disappear and the meristem stops without death or flowering, a case of ‘terminal differentiation’ (Forsyth et al., 1999). VI. Conclusions Describing the study of botany in the early 1800s, Sachs observed that, ‘a curious misconception crept in among the phytotomists at this time; they believed that more correct and trustworthy figures would be obtained if the observer and writer did not himself make them, but employed other eyes and other hands for that purpose; they imagined that in this way every kind of prejudice, of preconceived opinion would be eliminated from the drawings’ (Sachs, 1890). Today we might feel that in an age of photography, sequencing, imaging, we have machines of objectivity that place us beyond that problem, yet we still find only what we look for; what we see depends on how we interpret the information available. Through recent history the shoot apical meristem has been a constant but its scientific representation has been ever-changing. A model of shoot meristem function summarizes 49 50 Review Tansley review Fig. 13 An indeterminate flower of Impatiens balsamina. Proliferations of petal-like organs arise on the placental column and burst out from within the seed pod. Photograph courtesy of Jason Pole. available knowledge, directs future research, and promotes a point of view. Acknowledgements FT would like to thank Lucy Cavendish College, Cambridge and the Stanley Smith (UK) Horticultural Trust. NHB acknowledges research funding from BBSRC, DEFRA and The University of Reading Research Endowment Trust Fund. The legend to Fig. 2 was kindly translated from the German by Alex Angenendt (The University of Reading). References Adler I, Barabe D, Jean RV. 1997. A history of the study of phyllotaxis. Annals of Botany 80: 231–244. Angenent GC, Colombo L. 1996. Molecular control of ovule development. Trends in Plant Science 1: 228–232. Arber A. 1950. The natural philosophy of plant form. Cambridge, UK: Cambridge University Press. Barton MK. 1998. Cell type specification and self renewal in the vegetative shoot apical meristem. Current Opinion in Plant Biology 1: 37–42. Barton MK. 2001. Leaving the meristem behind: regulation of KNOX genes. Genome Biology 2: 1–3. Battey NH. 2003. Plant Culture: thirteen seasonal pieces. February – constructing a corymb. Journal of Experimental Botany 54: 605–608. Bowes BG. 1996. A colour atlas of plant structure. London, UK: Manson Publishing. Bowman JL, Smyth DR, Meyerowitz EM. 1989. Genes directing flower development in Arabidopsis. Plant Cell 1: 37–52. Buvat R. 1952. Structure, évolution et fonctionnement du méristème apical de quelques dicotylédones. Annales des Sciences Naturelles (11th Series, Botany) 8: 199–300. Carpenter R, Coen ES. 1990. Floral homeotic mutations produced by transposon-mutagenesis in Antirrhinum majus. Genes and Development 4: 1483–1493. Carpenter R, Copsey L, Vincent C, Doyle S, Magrath R, Coen E. 1995. Control of flower development and phyllotaxy by meristem identity genes in Antirrhinum. Plant Cell 7: 2001–2011. Church AH. 1904. The principles of phyllotaxis. Annals of Botany 18: 227– 243. Church AH. 1920. On the interpretation of phenomena of phyllotaxis. New York, USA: Hafner. Clark SE. 1996. The shoot meristem as a site of continuous organogenesis. Seminars in Cell and Developmental Biology 7: 873–880. Clark SE, Running MP, Meyerowitz EM. 1993. CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development 119: 397– 418. Clark SE, Williams RW, Meyerowitz EM. 1997. The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell 89: 575–585. Clowes FAL. 1961. Apical meristems. Oxford, UK: Blackwell Scientific Publications. Coen ES. 1991. The role of homeotic genes in flower development and evolution. Annual Review of Plant Physiology and Plant Molecular Biology 42: 241–279. Coen ES. 1996. Floral symmetry. EMBO Journal 15: 6777–6788. Coen ES, Carpenter R. 1993. The metamorphosis of flowers. Plant Cell 5: 1175–1181. Coen ES, Meyerowitz EM. 1991. The war of the whorls: genetic interactions controlling flower development. Nature 353: 31–37. Coen ES, Romero JM, Doyle S, Elliott R, Murphy G, Carpenter R. 1990. FLORICAULA: a homeotic gene required for flower development in Antirrhinum majus. Cell 63: 1311–1322. Crone W, Lord EM. 1994. Floral organ initiation and development in wild-type Arabidopsis thaliana (Brassicaceae) and in the organ identity mutants apetala 2–1 and agamous-1. Canadian Journal of Botany 72: 384–401. Cutter EG. 1959. On a theory of phyllotaxis and histogenesis. Biological Reviews 34: 243–263. Dermen H. 1947. Periclinal cytochimeras and histogenesis in cranberry. American Journal of Botany 34: 32–43. Fleming AJ, McQueen-Mason S, Mandel T, Kuhlemeier C. 1997. Induction of leaf primordia by the cell wall protein expansin. Science 276: 1415–1418. www.newphytologist.com © New Phytologist (2003) 159: 37 – 52 Tansley review Forsyth JL, Pearson S, Hadley P, Barnett JR. 1999. Apical abortion in calabrese is induced by periods of low temperature and results in premature differentiation of apical meristem cells. Journal of Experimental Botany 50: 861–868. Foster AS. 1938. Structure and growth of the shoot apex in Gingko biloba. Bulletin of the Torrey Botanical Club 65: 531–556. Foster AS. 1939. Problems of structure, growth and evolution in the shoot apex of seed plants. Botanical Review 5: 454–470. Gallois J-L, Woodward C, Reddy GV, Sablowski R. 2002. Combined SHOOTMERISTEMLESS and WUSCHEL trigger ectopic organogenesis in Arabidopsis. Development 129: 3207–3217. Goebel K. 1926. Wilhelm Hofmeister. The life and work of a nineteenth century botanist. London, UK: The Ray Society. Green PB. 1992. Pattern formation in shoots: a likely role for minimal energy configurations of the tunica. International Journal of Plant Sciences 153: 59–75. Green PB. 1994. Connecting gene and hormone action to form, pattern and organogenesis: biophysical transductions. Journal of Experimental Botany 45: 1775–1788. Green PB. 1996. Expression of form and pattern in plants – a role for biophysical fields. Seminars in Cell and Developmental Biology 7: 903– 911. Green PB. 1997. Expansin and morphology: a role for biophysics. Trends in Plant Science 2: 365–366. Green PB. 1999. Expression of pattern in plants: combining molecular and calculus-based biophysical paradigms. American Journal of Botany 86: 1059–1076. Green PB, Steele CS, Rennich SC. 1996. Phyllotactic patterns: a biophysical mechanism for their origin. Annals of Botany 77: 515–527. Hackett WP, Sachs RM. 1968. Experimental separation of inflorescence development from initiation in Bougainvillea. Proceedings of the American Society for Horticultural Science 92: 615 –621. Hernandez LF, Green PB. 1993. Transductions for the expression of structural pattern: analysis in sunflower. Plant Cell 5: 1725–1738. Hill JP, Lord EM. 1989. Floral development in Arabidopsis thaliana : a comparison of the wild type and the homeotic pistillata mutant. Canadian Journal of Botany 67: 2922–2936. Honma T, Goto K. 2001. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 409: 525–529. Huala E, Sussex IM. 1993. Determination and cell interactions in reproductive meristems. Plant Cell 5: 1157–1165. Jackson D, Hake S. 1999. Control of phyllotaxy in maize by the abphyl1 gene. Development 126: 315–323. Jean RV. 1994. Phyllotaxis: a systematic study in plant morphogenesis. Cambridge, UK: Cambridge University Press. Jeong S, Trotochaud AE, Clark SE. 1999. The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 11: 1925–1933. Kuhlemeier C, Reinhardt D. 2001. Auxin and phyllotaxis. Trends in Plant Science 6: 187–189. Laux T, Mayer KFX, Berger J, Jürgens G. 1996. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 122: 87–96. Lenhard M, Bohnert A, Jürgens G, Laux T. 2001. Termination of stem cell maintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS. Cell 105: 805 – 814. Lenhard M, Jürgens G, Laux T. 2002. The WUSCHEL and SHOOTMERISTEMLESS genes fulfil complementary roles in Arabidopsis shoot meristem regulation. Development 129: 3195 – 3206. Lohmann JU, Hong RL, Hobe M, Busch MA, Parcy F, Simon R, Weigel D. 2001. A molecular link between stem cell regulation and floral patterning in Arabidopsis. Cell 105: 793 – 803. Long JA, Moan EI, Medford JI, Barton MK. 1996. A member of the KNOTTED class of homeodomain proteins encoded by the SHOOTMERISTEMLESS gene of Arabidopsis. Nature 379: 66–69. © New Phytologist (2003) 159: 37 – 52 www.newphytologist.com Review Lord EM, Crone W, Hill JP. 1994. Timing of events during flower organogenesis: Arabidopsis as a model system. Current Topics in Developmental Biology 29: 325–356. Luo D, Carpenter R, Vincent C, Copsey L, Coen E. 1995. Origin of floral asymmetry in Antirrhinum. Nature 383: 794–799. Lyndon RF. 1994. Control of organogenesis at the shoot apex. New Phytologist 128: 1–18. Lyndon RF. 1998. The shoot apical meristem: its growth and development. Cambridge, UK: Cambridge University Press. Lynn K, Fernandez A, Aida M, Sedbrook J, Tasaka M, Masson P, Barton MK. 1999. The PINHEAD/ZWILLE gene acts pleiotropically in Arabidopsis development and has overlapping functions with the ARGONAUTE1 gene. Development 126: 469–481. Majumdar GP. 1942. The organization of the shoot in Heracleum in the light of development. Annals of Botany 6: 49–82. Mayer KFX, Schoof H, Haecker A, Lenhard M, Jürgens G, Laux T. 1998. Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 95: 805–815. McConnell JR, Barton MK. 1998. Leaf polarity and meristem formation in Arabidopsis. Development 125: 2935–2942. Meinhardt H. 1996. Models of biological pattern formation: common mechanism in plant and animal development. International Journal of Developmental Biology 40: 123–134. Meyerowitz EM. 1995. The molecular genetics of pattern formation in flower development: a perspective after ten years of the flowering newsletter. Flowering Newsletter 20: 4–12. Meyerowitz EM. 1996. Plant development: local control, global patterning. Current Opinion in Genetics and Development 6: 475–479. Meyerowitz EM, Bowman JL, Brockman LL, Drews GN, Jack T, Sieburth LE, Weigel D. 1991. A genetic and molecular model for flower development in Arabidopsis thaliana. Development 112: 157–168. Meyerowitz EM, Smyth DR, Bowman JL. 1989. Abnormal flowers and pattern formation in floral development. Development 106: 209–217. Mizukami Y, Ma H. 1997. Determination of Arabidopsis floral meristem identity by AGAMOUS. Plant Cell 9: 393–408. Parcy F, Nilsson O, Busch MA, Lee I, Weigel D. 1998. A genetic framework for floral patterning. Nature 395: 561–566. Philipson WR. 1949. The ontogeny of the shoot apex in dicotyledons. Biological Reviews 24: 21–50. Pien S, Wyrzykowska J, McQueen-Mason S, Smart C, Fleming A. 2001. Local expression of expansin induces the entire process of leaf development and modifies leaf shape. Proceedings of the National Academy of Sciences, USA 98: 11812–11817. Plantefol L. 1946. Fondements d’une théorie phyllotaxique nouvelle. Annales des Sciences Naturelles (11th series, botany) 7: 1–77. Reed HS. 1942. A short history of the plant sciences. Waltham, MA, USA: Chronica Botanica Company Publication. Reinhardt D, Mandel T, Kuhlemeier C. 2000. Auxin regulates the initiation and radial position of plant lateral organs. Plant Cell 12: 507–518. Reinhardt D, Wittwer F, Mandel T, Kuhlemeier C. 1998. Localized upregulation of a new expansin gene predicts the site of leaf formation in the tomato meristem. Plant Cell 10: 1427–1437. Richards FJ. 1951. Phyllotaxis: its quantitative expression and relation to growth in the apex. Philosophical Transactions of the Royal Society of London B 235: 509–564. Sablowski RWM, Meyerowitz EM. 1998. A homolog of NO APICAL MERISTEM is an immediate target of the floral homeotic genes APETALA3/PISTILLATA. Cell 92: 93–103. Sachs J. 1890. History of botany. Oxford, UK: Clarendon Press. Satina S, Blakeslee AF, Avery AG. 1940. Demonstration of the three germ layers in the shoot apex of Datura by means of induced polyploidy in periclinal chimeras. American Journal of Botany 27: 895–905. Scanlon MJ. 1998. Force fields and phyllotaxy: an old model comes of age. Trends in Plant Science 3: 413–414. 51 52 Review Tansley review Scanlon MJ, Schneeberger RG, Freeling M. 1996. The maize mutant narrow sheath fails to establish leaf margin identity in a meristematic domain. Development 122: 1683 –1691. Schoof H, Lenhard M, Haecker A, Mayer KFX, Jürgens G, Laux T. 2000. The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 100: 635–644. Schwabe WW. 1984. Phyllotaxis. In: Barlow PW, Carr DJ, eds. Positional controls in plant development. Cambridge, UK: Cambridge University Press, 403–440. Schwarz-Sommer Z, Huijser P, Nacken W, Saedler H, Sommer H. 1990. Genetic control of flower development by homeotic genes in Antirrhinum majus. Science 250: 931–936. Selker JML, Lyndon RF. 1996. Leaf initiation and de novo pattern formation in the absence of an apical meristem and pre-existing patterned leaves in watercress (Nasturtium officinale) axillary explants. Canadian Journal of Botany 74: 625 – 641. Sifton HB. 1944. Developmental morphology of vascular plants. New Phytologist 43: 87–129. Singer C. 1931. A short history of biology. Oxford, UK: Clarendon Press. Snow M, Snow R. 1931. Experiments on phyllotaxis. I. The effect of isolating a primordium. Philosophical Transactions of the Royal Society, London, Series B 221: 1 – 43. Snow M, Snow R. 1937. Auxin and leaf formation. New Phytologist 36: 1 – 18. Sommer H, Beltrán J-P, Huijser P, Pape H, Lönnig W-E, Saedler H, Schwarz-Sommer Z. 1990. DEFICIENS, a homeotic gene involved in the control of flower morphogenesis in Antirrhinum majus: the protein shows homology to transcription factors. EMBO Journal 9: 605 – 613. Steeves TA, Sussex IM. 1989. Patterns in plant development, 2nd edn. Cambridge, UK: Cambridge University Press. Sussex I. 1998. Themes in plant development. Annual Review of Plant Physiology and Plant Molecular Biology 49: 8 – 12. Theissen G. 2001. Genetics of identity. Nature 414: 491. Thornley JHM. 1975a. Phyllotaxis. I. A mechanistic model. Annals of Botany 39: 491–507. Thornley JHM. 1975b. Phyllotaxis. II. A description in terms of intersecting logarithmic spirals. Annals of Botany 39: 511–526. Trotochaud AE, Jeong S, Clark SE. 2000. CLAVATA3, a multimeric ligand for the CLAVATA1 receptor-kinase. Science 289: 613–617. Veen AH, Lindenmayer A. 1977. Diffusion mechanism for phyllotaxis: theoretical physico-chemical and computer study. Plant Physiology 60: 127–139. Veit B, Briggs SP, Schmidt RJ, Yanofsky MF, Hake S. 1998. Regulation of leaf initiation by the terminal ear 1 gene of maize. Nature 393: 166– 168. Vincent CA, Carpenter R, Coen ES. 1995. Cell lineage patterns and homeotic gene activity during Antirrhinum flower development. Current Biology 5: 1449–1458. Vogler H, Kuhlemeier C. 2003. Simple hormones but complex signalling. Current Opinion in Plant Biology 6: 51–56. Waites R, Hudson A. 1995. PHANTASTICA: a gene required for dorsoventrality of leaves in Antirrhinum majus. Development 121: 2143–2154. Wardlaw CW. 1949. Experiments on organogenesis in ferns. Growth Supplement 13: 93–131. Wardlaw CW. 1957. The reactivity of the apical meristem as ascertained by cytological and other techniques. New Phytologist 56: 221–229. Weigel D, Clark SE. 1996. Sizing up the floral meristem. Plant Physiology 112: 5–10. Wolff CF. 1896. Theoria generationis. Leipzig, Germany: Verlag von Wilhelm Engelmann. Yanofsky MF, Ma H, Bowman JL, Drews GN, Feldmann KA, Meyerowitz EM. 1990. The protein encoded by the Arabidopsis homeotic gene AGAMOUS resembles transcription factors. Nature 346: 35–39. www.newphytologist.com © New Phytologist (2003) 159: 37 – 52