Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
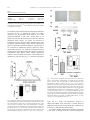
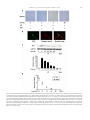
Cardiovascular Research 74 (2007) 471 – 479 www.elsevier.com/locate/cardiores Adiponectin accumulates in myocardial tissue that has been damaged by ischemia-reperfusion injury via leakage from the vascular compartment Rei Shibata a , Kaori Sato a , Masahiro Kumada b , Yasuhiro Izumiya a , Mina Sonoda b , Shinji Kihara b , Noriyuki Ouchi a , Kenneth Walsh a,⁎ a Molecular Cardiology/Whitaker Cardiovascular Institute, Boston University School of Medicine, 715 Albany Street, W611, Boston, MA 02118, USA b Department of Metabolic Medicine, Graduate School of Medicine, Osaka University, 2-2, Yamada-oka, Suita, Osaka, 565-0871, Japan Received 11 October 2006; received in revised form 25 January 2007; accepted 2 February 2007 Available online 12 February 2007 Time for primary review 31 days Abstract Objectives: Adiponectin, a circulating adipocyte-derived hormone, exerts beneficial actions on hearts subjected to ischemia-reperfusion injury. Adiponectin exists in plasma as three different oligomeric forms: trimer, hexamer and high molecular weight. This study investigated the expression and myocardial accumulation of adiponectin in a murine model of ischemia-reperfusion injury. Methods: Wild-type and adiponectin deficient mice were subjected to left anterior descending artery ligation followed by reperfusion. Plasma adiponectin levels were analyzed by ELISA and adiponectin in heart was determined by immunohistochemical, Western blot and real-time PCR analyses. Results: Plasma adiponectin levels declined after myocardial ischemia-reperfusion injury due to reductions in high molecular weight and, to a lesser extent, trimer and hexamer isoforms. Adiponectin protein was detected in injured but not sham-operated heart, and this was accompanied by a negligible increase in adiponectin transcript in the myocardium. Systemic delivery of adiponectin to adiponectin knockout (APN-KO) mice led to the accumulation of adiponectin in ischemia-reperfusion-injured, but not-uninjured hearts at levels comparable to wild-type suggesting that cardiac expression of adiponectin does not appreciably contribute to its accumulation in the infarcted heart. The serum half-life of adiponectin was 7.4 ± 0.3 h in ischemia-reperfusion and 9.7 ± 0.5 h in sham-operated mice (P N 0.05), whereas the half-life of adiponectin in the damaged heart was 26.9 ± 2.2 h (P b 0.05). Conclusions: These data show that adiponectin accumulates in the heart following ischemic damage primarily through leakage from the vascular compartment, and that adiponectin has a longer half-life in damaged heart tissue than in plasma. © 2007 European Society of Cardiology. Published by Elsevier B.V. All rights reserved. Keywords: Adiponectin; Ischemia-reperfusion; Myocardium; Infarction 1. Introduction Adiponectin, also referred to as adipocyte complementrelated protein of 30 kDa (ACRP30) [1,2], is an adipocyte- Abbreviations: AAR, area at risk; APN, adiponectin; CAD, coronary artery disease; HMW, high molecular weight; IA, infarct area; I-R, ischemiareperfusion injury; KO, knockout; LAD, left anterior descending artery; LV, left ventricular; WAT, white adipose tissue. ⁎ Corresponding author. Tel.: +1 617 414 2390; fax: +1 617 414 2391. E-mail address: [email protected] (K. Walsh). derived bioactive factor that is abundantly present in plasma. In clinical studies, circulating adiponectin levels are downregulated in association with obesity-linked disorders including hypertension, low grade inflammation and type 2 diabetes [3,4]. These obesity-linked disorders are implicated in the severity and outcome of ischemic heart disease including myocardial infarction [5,6]. Conversely, high plasma adiponectin levels are associated with a lower risk of CAD in some [7–10] but not all [11,12] studies. It is also recognized that high plasma adiponectin levels are associated with a lower risk of myocardial infarction [13] and that 0008-6363/$ - see front matter © 2007 European Society of Cardiology. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.cardiores.2007.02.010 472 R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 adiponectin levels rapidly decline in men following acute myocardial infarction [14]. Adiponectin exists in plasma as a trimer, hexamer or high molecular weight (HMW) forms in humans and mice [15,16]. The HMW form of adiponectin is lower in CAD patients, whereas the HMW form increases during weight reduction in obese subjects [16]. Several reports have shown that the HMW form of adiponectin determines insulin sensitivity and glucose tolerance [17–19]. Consistent with clinical observations, experimental studies showed that adiponectin-knockout (APN-KO) mice exhibit diet-induced insulin resistance, impaired endothelium-dependent vasodilation on atherogenic diets and impaired angiogenic response to ischemia [20,21]. Aortic constriction in APN-KO mice results in enhanced concentric cardiac hypertrophy and increased mortality [22,23], whereas adenovirus-mediated adiponectin expression attenuates cardiac hypertrophy in APN-KO, wild-type (WT) and diabetic db/db mice [22]. Adiponectin inhibits agonist-stimulated hypertrophy in cultured cardiac myocytes [22,24]. Recently, it was demonstrated that APN-KO mice develop larger infarcts following ischemia-reperfusion injury [25,26]. Conversely, supplementation of adiponectin diminishes infarct size in APN-KO and WT mice. Furthermore, adiponectin inhibits the development of severe myocarditis in leptin-deficient obese ob/ob mice [27]. These findings have led to the proposal that the short-term administration of this factor may have clinical utility in the treatment of patients suffering from acute myocardial infarction [28]. Recently a number of studies have shown that adiponectin transcript is upregulated in damaged tissues. For example, adiponectin mRNA is detected in the liver of mice after carbon tetrachloride-induced hepatic injury [29], and oxidative stress upregulates adiponectin mRNA in skeletal muscle cells [30]. With regard to the heart, adiponectin protein is detected at periphery of damaged myocytes in patients with myocardial infarction and dilated cardiomyopathy [31,32], and this protein is also detected in heart during the acute phase of virus-induced myocarditis in mice [33]. Adiponectin transcript has been detected in cultured cardiac myocytes [34] and in mouse models of myocardial injury [33,35]. Collectively, these data suggest that the endogenous production of this protein in injured tissues may contribute to its protective actions, but this has never been directly demonstrated. Therefore, a more thorough understanding of the expression and pharmokinetic properties of adiponectin is required to evaluate the therapeutic potential of this protein for the treatment of heart disease. In the present study, we examined the effect of myocardial ischemia-reperfusion injury in mice on circulating levels of adiponectin isoforms and assessed adiponectin expression and accumulation in the heart. Our observations indicate that despite a transient drop in plasma adiponectin following ischemia-reperfusion, adiponectin accumulates in the heart following injury, largely from leakage from the vascular compartment, and that it has a longer half-life in damaged heart tissue than in plasma. 2. Materials and methods 2.1. Materials Bacterially-produced mouse adiponectin was prepared as described previously [36]. The adenoviral vectors expressing β-galactosidase (Ad-βgal) or murine adiponectin (Ad-APN) have been described previously [21]. Adiponectin antibody that recognizes globular domain of mouse adiponectin, was purchased from R&D systems. Collagen type III antibody was purchased from Rockland Immunochemical. GAPDH antibody was purchased from Biogenesis Inc. 2.2. Mouse model of myocardial ischemia-reperfusion WT and APN-KO mice in a C57/BL6 background were used for this study [25]. Study protocols conform to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (Publication No. 8523, revised 1996). Mice at the ages of 10 to 12 weeks were anesthetized with sodium pentobarbital (50 mg/kg intraperitoneally). A left thoracotomy was performed, and the left anterior descending artery (LAD) was visualized under a microscope and ligated with 8–0 silk suture using snare occluder. Area at risk and infarct size were determined as described previously [25]. Mice were subjected to 30 min of LAD ligation, followed by 1, 3, 6, 12, 24, 36, 48, 96 and 168 h of reperfusion. In some experiments, recombinant adiponectin protein or PBS vehicle was injected into the jugular vein of mice at 30 min before LAD ligation. In some experiments, 2 × 108 plaque-forming units (pfu) of Ad-APN or Ad-β-gal were injected into the jugular vein of APN-KO mice 3 d prior to the ischemia-reperfusion injury. For Western blot and real-time PCR analyses, Evans blue dye was injected through the jugular vein for delineation of the area at risk (AAR), and non-necrotic areas within the AAR regions were collected under a dissecting microscope. 2.3. Adiponectin level and oligomeric state in plasma Total mouse adiponectin levels were determined with adiponectin ELISA kits (Otsuka Pharmaceutical Co Ltd.). For this measurement blood (20 μl) was collected from tail vein at the time of ischemia-reperfusion surgery and at 0, 1, 3, 6, 12, 24, 36, 48, 96 and 168 h after reperfusion. To determine the oligomeric form of adiponectin, blood samples were collected by heart puncture from mice on postoperative 24 h. The oligomeric state of adiponectin was analyzed by gel filtration chromatography and ELISA on fractions of eluted proteins as described previously [16]. 2.4. Immunohistochemical analysis Mice were sacrificed and Left Ventricular (LV) tissue was obtained at 0, 3, 12, 24, 48, 96 and 168 h after R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 ischemia-reperfusion injury. Tissue samples were embedded in OCT compound (Sakura Finetech USA Inc) and frozen in liquid nitrogen. Tissue sections (7 μm in thickness) were incubated with goat polyclonal antiadiponectin antibody or control IgG antibody (Santa Cruz Biotechnology). In some experiments, double fluorescence staining was performed on frozen heart sections. Sections were stained with rabbit polyclonal anti-collagen type III antibody followed by the treatment with rhodamineconjugated secondary antibody to detect collagen type III, and subsequently with goat polyclonal anti-adiponectin antibody followed by the treatment with FITC-conjugated secondary antibody to detect adiponectin. The signals were detected and analyzed using a fluorescence microscope (Nikon Diaphot). 2.5. Western blot analysis Heart tissue samples (non-necrotic AAR and sham) obtained at 0, 3, 12, 24, 48, 96 and 168 h after surgery and subcutaneous white adipose tissue (WAT) from control mice were homogenized in lysis buffer. Proteins (30 μg) were separated with denaturing SDS-PAGE. Following transfer to membranes, immunoblot analysis was performed with the indicated antibodies. This was followed by incubation with secondary antibody conjugated with HRP. 2.6. Determination of adiponectin mRNA Adiponectin mRNA level in myocardium and WAT was quantified by real-time PCR. Total RNA from heart (nonnecrotic AAR and sham) and WAT were prepared with the use of a Qiagen kit. The cDNA was produced using oligo-dT primer (ThermoScript RT-PCR Systems, Invitrogen). PCR was performed on iCycler iQ Real-Time PCR Detection System (BIO-RAD) using SYBR Green 1 as a double standard DNA specific dye (Applied Biosystems) [3]. In some experiments, total RNA were isolated from heart and liver of Ad-APN-treated APN-KO mice at 24 h after ischemia-reperfusion or sham-operated APN-KO mice. PCR was performed using Taq DNA Polymerase (Takara). Primers were: 5′-AGGTTGGATGGCAGGC-3′ and 5′GTCTCACCCTTAGGACCAAGAA-3′ for mouse adiponectin and 5′-TCACCACCATGGAGAAGGC-3′ and 5′GCTAAGCAGTTGGTGGTGCA-3′ for mouse GAPDH. 473 3. Results 3.1. Plasma adiponectin levels decline after ischemiareperfusion injury We assessed the time-dependent changes of plasma adiponectin levels in mice following myocardial ischemiareperfusion injury. For these experiments, the LAD was ligated for 30 min followed by 24 h of reperfusion in C57/ BL6 mice (WT). Under the conditions of these assays, this injury led to a 30.5% ± 2.4% (P b 0.05) infarct area (IA) relative to AAR at 24 h in a parallel set of 5 WT mice (Table 1). Circulating adiponectin levels, determined from sequential tail vein bleeds (n = 5 mice), declined by 35% (P b 0.01) at 24 h after ischemia-reperfusion (Fig. 1A). Plasma adiponectin returned to pre-injury levels 7 days after surgery. To corroborate these findings, plasma adiponectin levels at 24 h after sham-operation or ischemia-reperfusion were determined in a third set of WT mice (n = 12 for each condition). At this time point, plasma adiponectin levels were 29% lower (P b 0.01) in mice after ischemia-reperfusion compared with those in sham-operated mice (Table 1). The distribution of adiponectin oligomers in plasma at 24 h after ischemia-reperfusion or sham-operation was analyzed by ELISA on column elution fractions following gel filtration chromatography (n = 4 mice per condition). Representative elution profiles of plasma adiponectin from individual WT mice after sham-operation or ischemiareperfusion are shown in Fig. 2a. All oligomeric forms of adiponectin were significantly decreased after ischemiareperfusion injury, although the HMW form showed the greatest decrease (Fig. 2b). The level of HMW declined by 33% (P b 0.01), whereas hexamer and trimer forms declined by 13% (P b 0.05) and 12% (P b 0.05), respectively. 3.2. Expression and accumulation of adiponectin in injured heart The presence of adiponectin in the heart was assessed by immunohistochemistry, immunoblot and real-time polymerase chain reaction assays. Representative photographs of 2.7. Statistical analysis Data are presented as mean ± SD. The mean value was compared between 2 groups using an unpaired t test. The comparison among more than 3 groups was performed by analysis of variance (ANOVA) with Scheffe's Ftest. Comparison of adiponectin half-life between different experimental groups was performed by a repeated measure ANOVA. A value of P b 0.05 was accepted as statistically significant. Fig. 1. Decreased plasma adiponectin levels following ischemia-reperfusion injury. Plots of time course of plasma adiponectin levels from the same series of individual mice following ischemia-reperfusion injury (n = 5). ⁎P b 0.05, ⁎⁎P b 0.01 vs. WT mice before surgery. 474 R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 Table 1 Infarct size after ischemia-reperfusion WT APN-KO APN-KO + APN % AAR/LV % IA/AAR % IA/LV 51.9 ± 6.2 50.3 ± 4.1 49.6 ± 1.7 30.5 ± 6.2 50.1 ± 4.8⁎⁎ 31.9 ± 3.9⁎⁎ 15.5 ± 1.5 28.1 ± 2.2⁎⁎ 16.8 ± 2.7⁎⁎ Infarct size was determined 24 h after ischemia-reperfusion. Recombinant adiponectin protein (APN) was injected into jugular vein of APN-KO mice at 30 min before LAD ligation. ⁎⁎P b 0.01 APN-KO + APN versus APN-KO and APN-KO versus WT. myocardium tissue stained with anti-adiponectin antibodies are shown in Fig. 3a. Adiponectin protein was readily detected in the myocardium of the AAR at 24 h after ischemia-reperfusion in WT mice, whereas little or no adiponectin could be detected in hearts of WT mice that had undergone sham surgery. No signal was detected in histological sections of hearts from APN-KO mice (data not shown). Western blotting analysis detected adiponectin protein in heart after ischemia-reperfusion, whereas little or no expression of adiponectin could be detected in shamoperated WT mice (Fig. 3b). The level of adiponectin protein in injured heart was approximately 30% of the level found in subcutaneous white adipose tissues (WAT). Adiponectin mRNA in the myocardial non-necrotic AAR and WAT was quantified by real time PCR (Fig. 3c). Ischemia-reperfusion Fig. 3. The expression of adiponectin in the ischemic area of WT mouse heart. a) Representative immunostaining of adiponectin from WT heart sections at 24 h after sham-operated or ischemia-reperfusion injury (magnification, ×400; bar indicates 50 μm) (n = 5 in each group). b) Expression of adiponectin protein in heart tissues from WT mice at 24 h after sham-operation or ischemia-reperfusion injury and in white adipose tissue (WAT) by Western blot analysis. Relative adiponectin levels were quantified using the NIH image. Immunoblots were normalized to GAPDH (⁎⁎P b 0.01 vs. sham-operated WT, mice n = 3–5). c) Adiponectin mRNA levels in WT mouse heart at 24 h after sham-operation or ischemia-reperfusion. WAT was used as a positive control for adiponectin expression. Results are expressed adiponectin mRNA levels relative to GAPDH mRNA levels. ⁎P b 0.05, ⁎⁎P b 0.01 vs. sham-operated WT mice (n = 3–6). Fig. 2. Adiponectin isoforms are decreased after ischemia-reperfusion injury. a) Representative elution profiles of adiponectin in plasma from WT mice after sham-operation (closed circle) or ischemia-reperfusion injury (open circle). Plasma was collected at 24 h after sham-operation or ischemiareperfusion. Plasma was fractionated by gel filtration chromatography, and the adiponectin levels in each fraction were determined by ELISA. b) Percent changes of each form of adiponectin between sham-operation and ischemia-reperfusion surgery (⁎P b 0.05, ⁎⁎P b 0.01, n = 4 in each group). injury led to a small, but reproducible, increase in adiponectin mRNA levels in the heart (1.5-fold). However adiponectin mRNA levels in heart under these conditions are more than 33,000-fold lower than in WAT. To examine whether adiponectin accumulates in the heart as the result of leakage from the vascular compartment, R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 475 Fig. 4. Accumulation and kinetics of exogenous adiponectin in the heart after acute ischemic injury. APN-KO mice were subjected to ischemia-reperfusion injury at 30 min after injection of exogenous adiponectin. a) Immunostaining of adiponectin at 3 and 96 h in myocardial ischemic area of APN-KO mice after injection of exogenous bacterially-produced adiponectin (magnification, ×400; bar indicates 50 μm) (n = 3 in each group). b) Double fluorescence staining of adiponectin (green) and collagen type III (red) in myocardial ischemic region of APN-KO mice at 3 h after injection of exogenous adiponectin. Co-localization is indicated by yellow in the merged images (magnification, ×400; bar indicates 50 μm). c) Detection of exogenous adiponectin in ischemic heart tissues at 0, 3, 12, 24, 48, 96 and 168 h in APN-KO mice after injection of adiponectin by Western blot analysis. Relative adiponectin levels were quantified using the NIH image. Immunoblots were normalized to GAPDH. (n = 3 in each group). d) Plasma adiponectin levels at 0.5, 3, 12, 24, 36, 48 and 96 h in APN-KO mice in response to ischemia-reperfusion (I-R, open circle) or sham (closed circle) treatment after injection of exogenous adiponectin. Plasma adiponectin levels were determined by ELISA (n = 3 in each group). 476 R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 Fig. 5. Accumulation of adiponectin protein in damaged heart tissue. a) Adiponectin mRNA in heart and liver from sham-operated APN-KO mice or APN-KO mice treated with Ad-APN at 24 h after ischemia-reperfusion injury. Adiponectin mRNA expression was determined by PCR. b) Expression of adiponectin protein in heart tissues from WT mice and APN-KO mice treated with Ad-APN or Ad-βgal at 24 h after shamoperation or ischemia-reperfusion injury. Relative adiponectin levels were quantified from Western blot analyses using the NIH image (n = 3–5). Immunoblots were normalized to GAPDH. ⁎⁎P b 0.01 relative to shamoperated WT mice. APN-KO mice were subjected to ischemia-reperfusion injury 30 min after the intravenous injection of 1.0 μg/g recombinant adiponectin protein. Parallel experiments were performed on APN-KO mice that received vehicle (PBS). In the absence of exogenous adiponectin, IA/AAR × 100% was 50.1 ± 4.8% in APN-KO mice after 24 h of reperfusion (Table 1), whereas adiponectin protein administration led to a 37% decrease in infarct size at 24 h after injury, consistent with our previous findings [25]. At 3, 12, 24, 48, 96 and 168 h after injection of adiponectin protein, mice were killed and adiponectin accumulation in heart was assessed by immunohistochemistry (Fig. 4a). There was no detectable adiponectin prior to surgery in histological sections from hearts of APN-KO mice that had received adiponectin protein 3 h prior. In contrast, adiponectin protein was readily observed in the vasculature and myocardium of ischemic heart. This pattern of adiponectin distribution appeared similar to that seen in WT hearts subjected to ischemiareperfusion injury (Fig. 3a). The immunoreactivity appeared maximal at 3 h after injection of adiponectin and gradually declined at 12, 24, 48 and 96 h (Fig. 4a and data not shown). Adiponectin protein expression was not detected in hearts at 168 h after intravenous injection of protein. To examine whether adiponectin co-localizes with extracellular matrix in the injured myocardium, dual immunofluorescence staining was performed on adiponectin and collagen type III, a major component of the myocardial collagen matrix [37]. Hearts from APN-KO mice were harvested 3 h after the administration of adiponectin protein (2.5 h after the initiation of ischemia-reperfusion injury). Histological sections were co-stained for adiponectin (green) and collagen type III (red). Merged images revealed that these proteins co-localized (yellow) in the vasculature and myocardium of ischemic heart (Fig. 4b). The changes of adiponectin protein levels, assessed by semi-quantitative Western immunoblot analysis, paralleled the changes in immunoreactivity following ischemia-reperfusion injury (Fig. 4c). The highest levels of adiponectin protein were detected 3 h after the injection of adiponectin and levels declined at 12, 24, 48 and 96 h. No adiponectin could be detected at 168 h. No adiponectin was detected 3 h after adiponectin delivery in sham surgery hearts. Using these data, the calculated half-life of exogenous adiponectin in the myocardial AAR was 26.9 ± 2.2 h. We also examined the clearance of intravenously injected adiponectin in APN-KO mice. Recombinant adiponectin 1.0 μg/g was delivered intravenously through the jugular vein 30 min prior to ischemia or sham surgery (n = 3). At different time points blood was sequentially obtained from tail vein and serum adiponectin levels were determined by ELISA (Fig. 4d). The calculated half-life of adiponectin in plasma was similar under both these conditions (7.4 ± 0.3 h in ischemia-reperfusion-treated mice and 9.7 ± 0.5 h in shamoperated mice, P N 0.05). However, the half-life of adiponectin in the plasma was significantly lower than in the myocardial AAR (P b 0.05). Finally, we compared adiponectin levels in WT mice and in APN-KO mice that received 2 × 108 pfu of Ad-APN or Ad-βgal via the jugular vein 3 d prior to myocardial Table 2 Plasma adiponectin levels Mouse genotype I-R Adenovirus Adiponectin (μg/ml) WT WT APN-KO APN-KO APN-KO APN-KO − + − + − + − − Ad-βgal Ad-βgal Ad-APN Ad-APN 13.9 ± 1.3 10.1 ± 1.4 b0.05 b0.05 10.0 ± 1.7 8.4 ± 1.2 Measurements were made at 24 h after surgery. Adenoviral vectors were delivered intravenously via the jugular vein 3 days prior to surgery. Adiponectin levels are reported as mean ± standard error. R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 ischemia-reperfusion injury (30 min to 24 h, respectively) or sham-operation. This mode of delivery leads to the transduction of liver (Fig. 5a), and no adiponectin could be detected in hearts of sham-operated APN-KO mice that had received Ad-APN (Fig. 5b). In contrast, a low level of adiponectin could be detected in hearts from sham-operated WT mice, and this could represent the endogenous production of adiponectin by cardiac tissues. Following ischemia-reperfusion injury, similar levels of adiponectin protein were detected by Western immunoblot in the ischemic regions of the myocardium between WT mice and the APN-KO mice that had been treated with Ad-APN (Fig. 5b). Under these conditions, plasma levels of adiponectin were 10.1 ± 1.4 μg/ml in WT and 8.4 ± 1.2 μg/ml in APN-KO mice that received Ad-APN at 24 h after reperfusion (Table 2). These data suggest that leakage from the vascular compartment is the predominant source of adiponectin in the damaged myocardium. 4. Discussion Recently it has been shown that adiponectin exerts cardioprotective activities in models of pathological hypertrophy [22], viral myocarditis [27] and ischemia-reperfusion injury [25,26]. In the present study, we analyzed adiponectin levels in plasma and heart tissue before and after myocardial ischemia-reperfusion injury to elucidate aspects of the pharmacokinetics of adiponectin therapy for cardiac disorders. Total plasma adiponectin levels were found to decline transiently in mice following myocardial ischemia-reperfusion injury. At the 24 h timepoint decline was attributed to reductions in HMW and, to lesser extents, hexamer and trimer isoforms of adiponectin. Relatively abundant adiponectin protein could be detected in the ischemic area of hearts but the level of adiponectin protein in non-injured hearts was lower by a factor of 3. In experiments involving systemic delivery of adiponectin protein to APN-KO mice, adiponectin could be detected in hearts following ischemiareperfusion injury, but not in non-injured hearts. These data suggest that myocardial damage is required for the appreciable accumulation of adiponectin protein in heart. Previously, it has been shown that adiponectin transcript is almost exclusively expressed by adipose tissue [2]. Recently, however, it was reported that adiponectin transcript is synthesized by mouse cardiomyocytes [34], and is upregulated in virus-induced damaged myocardium [33,35]. Consistent with these latter reports, we find a 1.5fold elevation of adiponectin mRNA in heart following ischemia-reperfusion injury. However, quantitative RT-PCR analysis reveals that adiponectin transcript levels in injured heart are lower by a factor of 33,000, and by a factor of 49,000 in non-injured heart, compared to that in adipose tissue. In contrast, the level of adiponectin protein in injured heart is within a factor of 3 of that in adipose tissue, suggesting that endogenous production of adiponectin is unlikely to be a major source of this protein in injured heart. 477 To test directly whether leakage from the vascular compartment is a major source of this protein in damaged heart, an adenoviral vector expressing adiponectin was delivered to APN-KO mice prior to ischemia-reperfusion injury. Systemic delivery results in adiponectin expression predominantly from liver, and no expression could be detected in non-injured heart in our experiments. Following systemic delivery, adenoviral vectors are reported to interact with blood proteins that direct their binding to receptors on the hepatic cell surface [38]. The rescue of APN-KO mice with the systemic delivery of Ad-APN restored plasma adiponectin levels to 80% the level found in WT mice. Furthermore, following ischemia-reperfusion injury, myocardial adiponectin levels were within 87% of that in WT heart (Fig. 5, no statistically significant difference between WT and APN-KO treated with Ad-APN). Collectively, these data suggest that adiponectin protein accumulates in the heart following injury largely as a result of leakage from the vascular compartment. Consistent with the notion that adiponectin accumulates in damaged heart tissue due to breaches in the endothelial barrier, adiponectin protein can be detected in the subendothelial space at the early phase of catheter-injured vessels but not in the intact artery [39]. Adiponectin protein is also detected in the endothelium and subendothelial space of injured human vessels identified by thrombus attachment, but not in vessels with an intact endothelium [40]. Solid-phase binding assays have shown that adiponectin can bind to collagen type I, III and V, presumably through interactions with its collagen-like domain [39]. Collagens type I and III are major components of the cardiac extracellular matrix, and cardiac fibroblasts and endothelial cells produce collagens type I and III after myocardial infarction [37]. In this regard, adiponectin protein has been detected at the periphery of damaged myocytes in patients with myocardial infarction and dilated cardiomyopathy [31,32]. In the present study, it was shown that adiponectin protein was systemically delivered in APN-KO mice, and adiponectin co-localized with myocardial collagen type III following ischemia-reperfusion injury in APN-KO mice that were treated with recombinant adiponectin. In our study, adiponectin levels in WT mice significantly declined 35% after ischemia-reperfusion, which is comparable to the 29% decrease found in human after myocardial infarction [14]. The accumulation of adiponectin in damaged regions of the heart could contribute to the decline of circulating adiponectin levels following acute cardiac injury. In addition, tumor necrosis factor (TNF)-α and interleukin (IL)-6 are known to suppress adiponectin transcript expression in adipose tissue [4], and serum levels of TNF-α and IL6 rapidly increase after ischemia-reperfusion injury in the heart [41,42]. Thus, the upregulation inflammatory cytokines following acute myocardial injury may contribute to the decline in endogenous adiponectin levels in WT mice through a reduction in its rate of synthesis in adipose tissue. Adiponectin transcript is upregulated in the injured heart in response to ischemia-reperfusion or virus injury [33,35]. 478 R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 Furthermore, adiponectin mRNA levels in skeletal muscle and liver are increased under conditions of inflammation or oxidative stress [29,30,43]. In contrast, adiponectin is negatively regulated by pro-inflammatory cytokines or oxidative stress in adipose tissue [4,44]. Collectively, these data suggest that the upregulation of adiponectin by stress in non-adipose tissues may confer localized protection to injured tissue at a time when its expression by adipose tissues decline leading to a reduction in circulating levels of adiponectin. However, the mechanisms that contribute to the differential regulation of adiponectin between adipose and non-adipose tissues are unknown. Combs et al. have reported that the half-life of recombinant murine adiponectin is 6 h in FVB mice [45], which is compatible with our data using APN-KO mice after injection of bacterially-produced adiponectin protein (9.7 ± 0.5 h). More recently, Peake et al. reported that human adiponectin had a half-life of 14.3 h when injected into rabbits [46]. Furthermore, they reported that the HMW and trimer forms had half-lives of 13.0 and 17.5 h, respectively. Currently, the cardioprotective role of each oligomeric form of adiponectin is unknown, and future studies will be required to clarify the specific actions of each adiponectin oligomer on the injured heart. Of note, it is shown here that adiponectin has a longer half-life in damaged myocardial tissue, where it is bound to collagen matrix, than in the plasma. It is conceivable that the different oligomeric forms of adiponectin have different affinities for matrix and, thus, would exhibit different halflives in injured tissues. In summary, this work shows that myocardial ischemiareperfusion injury leads to a reduction in serum levels of adiponectin in mice. Low levels of adiponectin protein are detected in uninjured heart, but relatively high levels of adiponectin are detected in the heart following acute injury. While ischemia-reperfusion injury results in a small upregulation of adiponectin transcripts in the heart, myocardial levels of this protein are similar between WT mice and Ad-APN-treated APN-KO mice that synthesize adiponectin from non-myocardial tissues. Collectively, these data indicate that adiponectin accumulates in damaged heart primarily as the result of leakage from the vascular compartment. These findings are consistent with the notion that adiponectin enters damaged myocardial tissue, and participates in the wound healing process by binding to matrix. Acknowledgments This work was supported by the National Institutes of Health grants HL66957, HL77774, AR40197 and AG15052 to K. Walsh and Grant-in-Aid for Scientific Research on Priority Areas to S. Kihara. R. Shibata was supported by a grant from the American Heart Association Postdoctoral Fellowship Award, Northeast Affiliate. N. Ouchi was supported by an American Heart Association Scientist Development Grant, Northeast Affiliate. References [1] Scherer PE, Williams S, Fogliano M, Baldini G, Lodish HF. A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem 1995;270:26746–9. [2] Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose most abundant gene transcript 1). Biochem Biophys Res Commun 1996;221:286–9. [3] Ouchi N, Kihara S, Funahashi T, Nakamura T, Nishida M, Kumada M, et al. Reciprocal association of C-reactive protein with adiponectin in blood stream and adipose tissue. Circulation 2003;107:671–4. [4] Ouchi N, Kihara S, Funahashi T, Matsuzawa Y, Walsh K. Obesity, adiponectin and vascular inflammatory disease. Curr Opin Lipidol 2003;14:561–6. [5] Wolk R, Berger P, Lennon RJ, Brilakis ES, Somers VK. Body mass index: a risk factor for unstable angina and myocardial infarction in patients with angiographically confirmed coronary artery disease. Circulation 2003;108:2206–11. [6] Orlander PR, Goff DC, Morrissey M, Ramsey DJ, Wear ML, Labarthe DR, et al. The relation of diabetes to the severity of acute myocardial infarction and post-myocardial infarction survival in Mexican– Americans and non-Hispanic whites. The Corpus Christi Heart Project. Diabetes 1994;43:897–902. [7] Schulze MB, Shai I, Rimm EB, Li T, Rifai N, Hu FB. Adiponectin and future coronary heart disease events among men with type 2 diabetes. Diabetes 2005;54:534–9. [8] Kumada M, Kihara S, Sumitsuji S, Kawamoto T, Matsumoto S, Ouchi N, et al. Association of hypoadiponectinemia with coronary artery disease in men. Arterioscler Thromb Vasc Biol 2003;23:85–9. [9] Ouchi N, Kihara S, Arita Y, Maeda K, Kuriyama H, Okamoto Y, et al. Novel modulator for endothelial adhesion molecules: adipocytederived plasma protein adiponectin. Circulation 1999;100:2473–6. [10] Zoccali C, Mallamaci F, Tripepi G, Benedetto FA, Cutrupi S, Parlongo S, et al. Adiponectin, metabolic risk factors, and cardiovascular events among patients with end-stage renal disease. J Am Soc Nephrol 2002;13:134–41. [11] Lindsay RS, Resnick HE, Zhu J, Tun ML, Howard BV, Zhang Y, et al. Adiponectin and coronary heart disease: the Strong Heart Study. Arterioscler Thromb Vasc Biol 2005;25:e15–6. [12] Lawlor DA, Davey Smith G, Ebrahim S, Thompson C, Sattar N. Plasma adiponectin levels are associated with insulin resistance, but do not predict future risk of coronary heart disease in women. J Clin Endocrinol Metab 2005;90:5677–83. [13] Pischon T, Girman CJ, Hotamisligil GS, Rifai N, Hu FB, Rimm EB. Plasma adiponectin levels and risk of myocardial infarction in men. JAMA 2004;291:1730–7. [14] Kojima S, Funahashi T, Sakamoto T, Miyamoto S, Soejima H, Hokamaki J, et al. The variation of plasma concentrations of a novel, adipocyte-derived protein, adiponectin, in patients with acute myocardial infarction. Heart 2003;89:667. [15] Pajvani UB, Du X, Combs TP, Berg AH, Rajala MW, Schulthess T, et al. Structure-function studies of the adipocyte-secreted hormone Acrp30/ adiponectin. Implications fpr metabolic regulation and bioactivity. J Biol Chem 2003;278:9073–85. [16] Kobayashi H, Ouchi N, Kihara S, Walsh K, Kumada M, Abe Y, et al. Selective suppression of endothelial cell apoptosis by the high molecular weight form of adiponectin. Circ Res 2004;94:e27–31. [17] Fisher FF, Trujillo ME, Hanif W, Barnett AH, McTernan PG, Scherer PE, et al. Serum high molecular weight complex of adiponectin correlates better with glucose tolerance than total serum adiponectin in Indo-Asian males. Diabetologia 2005;48:1084–7. [18] Pajvani UB, Hawkins M, Combs TP, Rajala MW, Doebber T, Berger JP, et al. Complex distribution, not absolute amount of adiponectin, correlates with thiazolidinedione-mediated improvement in insulin sensitivity. J Biol Chem 2004;279:12152–62. R. Shibata et al. / Cardiovascular Research 74 (2007) 471–479 [19] Lara-Castro C, Luo N, Wallace P, Klein RL, Garvey WT. Adiponectin multimeric complexes and the metabolic syndrome trait cluster. Diabetes 2006;55:249–59. [20] Ouchi N, Ohishi M, Kihara S, Funahashi T, Nakamura T, Nagaretani H, et al. Association of hypoadiponectinemia with impaired vasoreactivity. Hypertension 2003;42:231–4. [21] Shibata R, Ouchi N, Kihara S, Sato K, Funahashi T, Walsh K. Adiponectin stimulates angiogenesis in response to tissue ischemia through stimulation of amp-activated protein kinase signaling. J Biol Chem 2004;279:28670–4. [22] Shibata R, Ouchi N, Ito M, Kihara S, Shiojima I, Pimentel DR, et al. Adiponectin-mediated modulation of hypertrophic signals in the heart. Nat Med 2004;10:1384–9. [23] Liao Y, Takashima S, Maeda N, Ouchi N, Komamura K, Shimomura I, et al. Exacerbation of heart failure in adiponectin-deficient mice due to impaired regulation of AMPK and glucose metabolism. Cardiovasc Res 2005;67:705–13. [24] Fujioka D, Kawabata K, Saito Y, Kobayashi T, Nakamura T, Kodama Y, et al. Role of adiponectin receptors in endothelin-induced cellular hypertrophy in cultured cardiomyocytes and their expression in infarcted heart. Am J Physiol Heart Circ Physiol 2006;290:H2409–16. [25] Shibata R, Sato K, Pimentel DR, Takemura Y, Kihara S, Ohashi K, et al. Adiponectin protects against myocardial ischemia-reperfusion injury through AMPK-and COX-2-dependent mechanisms. Nat Med 2005;11:1096–103. [26] Tao L, Jiao X, Gao E, Lau WB, Yuan Y, Lopez B, et al. Nitrative inactivation of thioredoxin-1 and its role in postischemic myocardial apoptosis. Circulation 2006;114:1395–402. [27] Takahashi T, Saegusa S, Sumino H, Nakahashi T, Iwai K, Morimoto S, et al. Adiponectin replacement therapy attenuates myocardial damage in leptin-deficient mice with viral myocarditis. J Int Med Res 2005;33:207–14. [28] Ouchi N, Shibata R, Walsh K. Cardioprotection by adiponectin. Trends Cardiovasc Med 2006;16:141–6. [29] Yoda-Murakami M, Taniguchi M, Takahashi K, Kawamata S, Saito K, Choi-Miura NH, et al. Change in expression of GBP28/adiponectin in carbon tetrachloride-administrated mouse liver. Biochem Biophys Res Commun 2001;285:372–7. [30] Delaigle AM, Senou M, Guiot Y, Many MC, Brichard SM. Induction of adiponectin in skeletal muscle of type 2 diabetic mice: in vivo and in vitro studies. Diabetologia 2006;49:1311–23. [31] Takahashi T, Saegusa S, Sumino H, Nakahashi T, Iwai K, Morimoto S, et al. Adiponectin, T-cadherin and tumour necrosis factor-alpha in damaged cardiomyocytes from autopsy specimens. J Int Med Res 2005;33:236–44. [32] Ishikawa Y, Akasaka Y, Ishii T, Yoda-Murakami M, Choi-Miura NH, Tomita M, et al. Changes in the distribution pattern of gelatin-binding protein of 28 kDa (adiponectin) in myocardial remodelling after ischaemic injury. Histopathology 2003;42:43–52. 479 [33] Takahashi T, Zhu SJ, Sumino H, Saegusa S, Nakahashi T, Iwai K, et al. Inhibition of cyclooxygenase-2 enhances myocardial damage in a mouse model of viral myocarditis. Life Sci 2005;78:195–204. [34] Pineiro R, Iglesias MJ, Gallego R, Raghay K, Eiras S, Rubio J, et al. Adiponectin is synthesized and secreted by human and murine cardiomyocytes. FEBS Lett 2005;579:5163–9. [35] Takahashi T, Yu F, Saegusa S, Sumino H, Nakahashi T, Iwai K, et al. Impaired expression of cardiac adiponectin in leptin-deficient mice with viral myocarditis. Int Heart J 2006;47:107–23. [36] Ouchi N, Kobayashi H, Kihara S, Kumada M, Sato K, Inoue T, et al. Adiponectin stimulates angiogenesis by promoting cross-talk between AMP-activated protein kinase and Akt signaling in endothelial cells. J Biol Chem 2004;279:1304–9. [37] Cleutjens JP, Verluyten MJ, Smiths JF, Daemen MJ. Collagen remodeling after myocardial infarction in the rat heart. Am J Pathol 1995;147:325–38. [38] Shayakhmetov DM, Gaggar A, Ni S, Li ZY, Lieber A. Adenovirus binding to blood factors results in liver cell infection and hepatotoxicity. J Virol 2005;79:7478–91. [39] Okamoto Y, Arita Y, Nishida M, Muraguchi M, Ouchi N, Takahashi M, et al. An adipocyte-derived plasma protein, adiponectin, adheres to injured vascular walls. Horm Metab Res 2000;32:47–50. [40] Ouchi N, Kihara S, Arita Y, Nishida M, Matsuyama A, Okamoto Y, et al. Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocytederived macrophages. Circulation 2001;103:1057–63. [41] Cain BS, Harken AH, Meldrum DR. Therapeutic strategies to reduce TNF-alpha mediated cardiac contractile depression following ischemia and reperfusion. J Mol Cell Cardiol 1999;31:931–47. [42] Zingarelli B, Hake PW, Yang Z, O'Connor M, Denenberg A, Wong HR. Absence of inducible nitric oxide synthase modulates early reperfusion-induced NF-kappaB and AP-1 activation and enhances myocardial damage. FASEB J 2002;16:327–42. [43] Delaigle AM, Jonas JC, Bauche IB, Cornu O, Brichard SM. Induction of adiponectin in skeletal muscle by inflammatory cytokines: in vivo and in vitro studies. Endocrinology 2004;145:5589–97. [44] Iwashima Y, Katsuya T, Ishikawa K, Kida I, Ohishi M, Horio T, et al. Association of hypoadiponectinemia with smoking habit in men. Hypertension 2005;45:1094–100. [45] Combs TP, Berg AH, Rajala MW, Klebanov S, Iyengar P, JimenezChillaron JC, et al. Sexual differentiation, pregnancy, calorie restriction, and aging affect the adipocyte-specific secretory protein adiponectin. Diabetes 2003;52:268–76. [46] Peake PW, Kriketos AD, Campbell LV, Shen Y, Charlesworth JA. The metabolism of isoforms of human adiponectin: studies in human subjects and in experimental animals. Eur J Endocrinol 2005;153:409–17.