Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
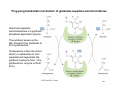
Both PS 7 and PS 8 are due next Thursday 1 Comments about PS6 Chymotrypsin Elastase Moderately Deep and Wide Accepts Phenyl group of Phe Shallow, Accepts smaller apolar residues Gly and Ala Trypsin Active Site Stabilizing Residues Cavity Deep, narrow channel Accepts Lysine Asp 189 acts as a stabilizing residue 2 Michaelis-Menten Equation v= Vmax [S] KM + [S] Vmax the reaction rate when the enzyme is fully saturated with substrate KM, the Michaelis constant, is the substrate concentration at which the reaction rate is half maximal. . 3 The turnover number • • • • A measure of catalytic activity kcat, the turnover number, is the number of substrate molecules converted to product per enzyme molecule per unit of time, when E is saturated with substrate. According to M-M , k2 = kcat = Vmax/Et Values of kcat usually range from less than 1/sec to 104/sec Catalase is 4 x 107 /sec 4 The catalytic efficiency kcat/Km • kcat/Km is an apparent second-order rate constant • It measures how the enzyme performs when S is low • The upper limit for kcat/Km is the diffusion limit the rate at which E and S diffuse together (~109/M for small substrates (glycerol) and 108/M for larger substrates (nucleotides)) 5 Example of MM Eqn Question triose phosphate isomerase Glyceraldehyde-3-P ÅÆdihydroxyacetone-P KM = 1.8 x 10-5 M When [glyceraldehyde-3-P] = 30 µM, v = 82.5 µmol mL-1 sec-1 What is Vmax for the triose phosphate isomerase? Assume [ET] = 3 nmol/mL, what is kcat for triose phosphate isomerase? What can we say about the catalytic efficiency of triose phosphate isomerase? 6 Linear Plots of the MichaelisMenten Equation Linear plots allow Km and Vmax to be estimated by extrapolation of lines rather than asymptotes • Lineweaver-Burk (double reciprocal plot) • Hanes-Woolf – Preferred because there isn’t an overemphasis of the data obtained at low [S] Deviation from the linear plot implies allostery (regulation) 7 Lineweaver-Burk y = m x + b The Lineweaver-Burk double-reciprocal plot, depicting extrapolations that allow the determination of the x- and y-intercepts and slope. 8 Hanes-Woolf A Hanes-Woolf plot of [S]/v versus [S], another straight-line rearrangement of the MichalelisMenten equation. 9 What Can Be Learned from the Inhibition of Enzyme Activity? • Enzymes may be inhibited reversibly or irreversibly • Reversible inhibitors may bind at the active site or at some other site • Enzymes may also be inhibited in an irreversible manner 10 Different Types of Inhibition 12 Vmax 10 Relative velocity No Inhibitor 8 Km Competitive Inhibitor 6 Uncompetitive Inhibitor Inhibitor binds to same site as substrate reversibly Inhibitor only binds to Enzyme/substrate complex 4 Pure Noncompetitive Inhibitor Inhibitor binds to different site as substrate and does not affect the binding of substrate Mixed NI – affects S binding 2 0 0 10 20 30 40 50 60 70 [S] 11 Effect of Inhibitors on the Michaelis-Menten Rate Equation Inhibition Type Rate Equation Apparent KM Apparent Vmax None v = Vmax[S]/(KM+[S]) KM Vmax Competitive v = Vmax[S]/ ([S]+ KM(1 + [I]/KI)) KM(1 + [I]/KI) Vmax Noncompetitive v = (Vmax[S]/(1 + [I]/KI))/(KM+[S]) KM Vmax/(1 + [I]/KI) Mixed v = Vmax[S]/((KM(1 + [I]/KI) +([S](1 + [I]/KI’))) KM(1+[I]/KI)/ (1+[I]/KI’) Vmax/(1 + [I]/KI’) Uncompetitive v = Vmax[S]/(KM+([S](1 + [I]/KI’)) KM/(1+[I]/KI’) Vmax/(1 + [I]/KI’) 12 Competitive Inhibition Inhibitor binds to same site as substrate reversibly As the concentration of a competitive inhibitor increases, higher concentrations of substrate are required to attain a particular reaction velocity. The reaction pathway suggests how sufficiently high concentrations of substrate can completely relieve competitive inhibition. KI = k-3/k3 13 Competitive Inhibition KI = [E][I]/[EI] = k-3/k3 Lineweaver-Burk plot of competitive inhibition, showing lines for no I, [I], and 2[I]. Note that when [S] is infinitely large (1/[S] = 0), Vmax is the same, whether I is -1 present of not. In the presence of I, the x-intercept = [I] ⎞ ⎛ Km ⎜1 + ⎟ KI ⎠ ⎝ Derivation on page 423 14 An example of a competitive inhibitor Structures of succinate, the substrate of succinate dehydrogenase (SDH), and malonate, the competitive inhibitor. Fumarate (the product of SDH action on succinate) is also shown. 15 Pure Noncompetitive Inhibition Inhibitor binds to different site as substrate and does not affect the binding of substrate KI = KI’ KI’ The reaction pathway shows that the inhibitor binds both to free enzyme and to enzyme complex. Consequently, Vmax cannot be attained, even at high substrate concentrations. 16 Pure Noncompetitive Inhibition KI = KI’ Lineweaver-Burk plot of pure noncompetitive inhibition. Note that I does not alter Km but that it decreases Vmax. In the presence of I, the y-intercept is equal to (1/Vmax)(1 + I/KI). 17 Mixed Noncompetitive Inhibition Inhibitor binds to different site as substrate and affects the binding of substrate KI ≠ KI’ KI’ Lineweaver-Burk plot of mixed noncompetitive inhibition. Note that both intercepts and the slope change in the presence of I. (a) When KI is less than KI'; (b) when KI is greater than 18 KI'. Pure Uncompetitive Inhibition Inhibitor only binds to enzyme/substrate complex Lineweaver-Burk plot of pure uncompetitive inhibition. Note that both intercepts 19 change, but the slope (KM/Vmax) remains constant in the presence of I. Which of the following graphs shows the results of reaction rate vs substrate concentration for an non-allosteric enzyme in the absence and presence of a noncompetitive inhibitor (non-competitive inhibitors bind to an enzyme at a site different than the active site)? 20 What Is the Kinetic Behavior of Enzymes Catalyzing Bimolecular Reactions? • Enzymes often use two (or more) substrates • Reactions may be sequential or singledisplacement reactions • And they can be random or ordered enzyme A + B ÅÆP + Q E + A + B ÆAEBÆPEQÆE + P + Q 21 Creatine kinase acts by a random, single-displacement mechanism The structures of creatine and creatine phosphate, guanidinium compounds that are important in muscle energy metabolism. 22 Single-displacement bisubstrate mechanism Single-displacement bisubstrate mechanism. Double-reciprocal plots of the rates observed with different fixed concentrations of one substrate (B here) are graphed versus a series of 23 concentrations of A. Note that, in these Lineweaver-Burk plots for single-displacement bisubstrate mechanisms, the lines intersect to the left of the 1/v axis. Random, single-displacement bisubstrate mechanism Random, single-displacement bisubstrate mechanisms where A does not affect B binding, and vice versa. Note that the lines intersect at the 1/[A] axis. (If [B] were varied in an experiment with several fixed concentrations of A, the lines would intersect at the 1/[B] axis in a 1/v versus 24 1/[B] plot.) Ping-pong bisubstrate mechanism of glutamate aspartate aminotransferase Glutamate:aspartate aminotransferase is a pyridoxal phosphate-dependent enzyme. The pyridoxal serves as the NH2 acceptor from glutamate to form pyridoxamine. Pyridoxamine is then the amino donor to oxaloacetate to form aspartate and regenerate the pyridoxal coenzyme form. (The pyridoxamine: enzyme is the E' form.) 25 Ping-pong bisubstrate mechanism Double-displacement (ping-pong) bisubstrate mechanisms are characterized by LineweaverBurk plots of parallel lines when double-reciprocal plots of the rates observed with different fixed concentrations of the second substrate, B, are graphed versus a series of 26 concentrations of A.