Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
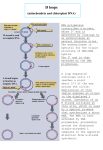
FEMS Microbiology Letters 111 (1993) 109-114 © 1993 Federation of European Microbiological Societies 0378-1097/93/$06.00 Published by Elsevier 109 FEMSLE 05523 Unique genetic arrangement in the dnaA region of the Borrelia burgdorferi linear chromosome: Nucleotide sequence of the dnaA gene Iain G. Old, Danielle Margarita and Isabelle Saint Girons Unit~ de Bact~riologie Mol~culaire et M~dicale, lnstitut Pasteur, Paris, France (Received 8 April 1993; revision received 28 April 1993; accepted 29 April 1993) Abstract." The complete nucleotide sequence of the Borrelia burgdorferi dnaA gene (encoding the initiator protein of chromosome replication) and its flanking regions was determined. The putative DnaA polypeptide exhibited 29-42% identity with those of other eubacteria. The gene order in the dnaA region at the centre of the B. burgdorferi linear chromosome is rnpA-rpmH-dnaN-dnaAgyrB-gyrA in contrast to the consensus eubacterial order of rnpA-rprnH-dnaA-dnaN-recF-gyrB, suggesting a rearrangement during the evolution of the Borrelia chromosome. We did not detect the multiple 9-nucleotide repeats known as DnaA boxes, which characterise origin of replications, in the dnaA-gyrB and dnaA-dnaN intergenic regions. In addition B. burgdorferi DnaA protein differs considerably from those of other eubacteria in a normally highly conserved region at the C-terminus of the polypeptide which may be involved in DNA binding. Key words."Borrelia burgdorferi; Linear chromosome; dnaA; dnaN; gyrB Introduction T h e dnaA g e n e p r o d u c t (the i n i t i a t o r p r o t e i n for c h r o m o s o m e r e p l i c a t i o n ) is o n e o f several p r o t e i n s which a r e essential for t h e r e p l i c a t i o n of t h e b a c t e r i a l c h r o m o s o m e . It is also t h e only p r o t e i n which is r e q u i r e d for initiation f r o m b a c t e r i a l origins a n d is n o t r e q u i r e d for D N A replic a t i o n events after initiation. D n a A b i n d s specifi- Correspondence to: I.G. Old, Unit6 de Bact6riologie Mol6culaire et M6dicale, D6partement de Bact6riologie et Mycologie, Institut Pasteur, 28 rue du Dr Roux, 75724 Paris Cedex 15, France. cally to 9 b p n u c l e o t i d e r e p e a t s k n o w n as D n a A boxes. O r i g i n r e g i o n s a r e c h a r a c t e r i s e d by nont r a n s l a t a b l e r e g i o n s c o n t a i n i n g at least 4 D n a A boxes in a c h a r a c t e r i s t i c a r r a n g e m e n t [1,2]. T h e c o n s e n s u s s e q u e n c e for a D n a A box is 5'T T A T C C A C A - 3 ' a n d a s e q u e n c e differing by up to 2 b a s e s while r e t a i n i n g t h e fourth, s e v e n t h a n d e i g h t h n u c l e o t i d e s m a y be d e f i n e d as such [3]. In G r a m - p o s i t i v e b a c t e r i a a n d p s e u d o m o n a d s , origins a r e f o u n d next to t h e g e n e s dnaA a n d gidA (Fig. 1) while in Escherichia coli t h e origin is f o u n d next to gidA, a n d a single D n a A box, essential for dnaA a u t o r e g u l a t i o n , is f o u n d in t h e dnaA p r o m o t e r r e g i o n [1,2]. T h e g e n e t i c o r g a n i s a t i o n of t h e dnaA r e g i o n of t h e c h r o m o s o m e has b e e n s t u d i e d in several eu- 110 bacteria: for reviews see [1,2]. The consensus is gid B-gidA - 5 OK-6O K-rnpA -rm p H-dnaA -dna NrecF-gyrB [4] (Fig. 1B,C) and between Pseudomonas putida and Bacillus subtilis a total of 12 genes in a 15 kb region are similar [4]. In E. coli there is less similarity as gyrA is elsewhere on the chromosome while gidB, gidA and the origin have been translocated by the inversion of a 40 kb region [4] (Fig. 1A). Due to the highly conserved nature of dnaA region of chromosome, it has been proposed that this was the replication origin of ancestral bacteria and that its organisation has been conserved in most eubacteria [1]. The spirochaete B. burgdorferi is an exception to the normal organisation in the dnaA region of the chromosome. While the rpmH, gyrB and gyrA genes of B. burgdorferi mapped to the centre of the 1 Mb linear chromosome, gidA was located between 293 and 395 kb from these genes [5-7]. In this study we present the sequence of the B. burgdorferi dnaA gene and demonstrate that the genetic organisation in this normally highly conserved region is unique among the eubacteria. Materials and Methods Bacterial strains and media E. coli strains XLl-blue and DH5a were grown as previously described [6]. Cloning of B. burgdorferi genes The dnaA clone, pB22, was isolated from a previously constructed library of EcoRI-partially digested B. burgdorferi 212 DNA in pUC18 [6]. Pulsed field gel electrophoresis (PFGE) and DNA hybridisations These were carried out as previously described [6]. DNA sequencing and computer analysis of sequences Deletions of pB22 were constructed with Exonuclease III using an Erase-a-base kit (Promega). DNA sequencing was carried out as previously described [6]. The GCG computer package [8] was used to carry out Fasta analyses and multiple sequence alignments. Results and discussion Sequencing of randomly chosen clones from our B. burgdorferi library allowed us to identify a clone, pB3, with a 3.5 kb insert, which carried gyr'BA' (DNA gyrase a and /3 subunits) [6]. We isolated a further clone, pB22, with a 4.8 kb insert which carried rpmH (ribosomal protein L34) [7]. Further sequence analysis revealed pB22 also carried dnaA, dnaN (DNA polymerase III /3 subunit), rnpA' (Ribonuclease P, protein component) and gyrB', the insert being contiguous with gyr'BA' carried by pB3 (Fig. 1D). The identification of these genes was inferred from Fasta analyses of their deduced amino acid sequences against the SWISS-PROT database [8]. The B. burgdorferi dnaA gene and its flanking regions were sequenced (Genbank accession number L14948). The 1458 nucleotide open reading frame encodes a 486 amino acid polypeptide of 56.8 kDa. The open reading frame is preceded by a possible ribosome binding site A G G A while a possible promoter, TFCACA-17N-CCTAAT is located 60 nucleotides upstream of the start codon. The GC % of B. burgdorferi is one o f the lowest in eubacteria (28-31%) and the codon usage is heavily biased to use A and T: 78.6% of codons in dnaA have A or T in the 3rd position. The deduced DnaA polypeptide of B. burgdorferi exhibits 29-42% identity with those from other eubacteria (Fig. 2). DnaA can be divided into three domains [9]: I, with moderate similarity; II (which may act as a hinge), with little similarity; and III, with high similarity. It has been noted that a remarkably conserved stretch found in the C-terminal region may be involved in DNA binding (unpublished data in [1]). The sequence G G R D H T T V is almost invariably present towards the C terminus of eubacterial DnaA polypeptides (SGRDHTTV is found in Buchnera aphidicola [10]), whereas in B. burgdorferi only 5 of the 8 amino acids are conserved (Fig. 2C). If this conserved stretch does represent a DNA-binding region of DnaA, this would suggest that B. 111 burgdorferi DnaA boxes, to which DnaA protein binds, may differ from those found in other eubacteria. Further sequencing of pB22 revealed an open reading frame 177 bp upstream of dnaA corresponding to the 5' end of gyrB (Fig. 1D). Like DnaA, GyrB is highly conserved and the corresponding region of the deduced polypeptide exhibited between 45 and 54% identity with those of other eubacteria. 242 bp downstream of dnaA was an open reading frame that would encode a polypeptide with 19-21% identity to the S-subunit of DNA polymerase III encoded by dnaN. DnaN is less well conserved than GyrB and DnaA and this low identity is in the order of that found between other eubacteria [11]. We also found an open reading frame following rpmH which, when translated, exhibited 35% identity with RnpA of Bacillus subtilis (results not shown). In Gramnegative bacteria the dnaA-dnaN intergenic region is a few nucleotides and the two genes form a single operon, while in Gram-positive bacteria the intergenic region is larger [1,2]. B. burgdorferi is more like Gram-positive bacteria in that the dnaA-dnaN intergenic region is 242 bp and the gyrBA genes are in tandem. The arrangement of the genes in the B. burgdorferi dnaA region is shown in Fig. 1D. We A E s c h e r i c h i a coli oriC --~¢ ~g/dA ~(40kb)~¢--.-.4~---- .~.~ g/d.B rnpA 60K rpmH 50K B P s e u d o m o n a s putida oriCll g/dB g/dA ~ dnaA .~ dnaN recF gyrB dnaN recF gyrB oriCl rnpA 60K rpmH 50K dnaA C B a c i l l u s subtilis orilll oril orUI i gidB gidA 50K mpA 60K rpmH dnaA dnaN recF gyrB gyrA D BorreUa burgdorferi (293-395kb) - . 4 ~ ~ r m gidA rnpA rpmH dnaN dnaA gyrA gyrB y pB4 pB22 pB3 Fig. 1. Genetic organisation in the dnaA regions of four different eubacteria, adapted from [1,2,4]. The open reading frames (boxed regions) and intergenic regions (gaps) are drawn approximately to scale. The direction of transcription is shown by single headed horizontal arrows while origin regions are marked with vertical arrows. The genes shown are: gidA, gidB (glucose inhibited division proteins); 50K, 60K (proteins of undetermined function); rnpA (ribonuclease P, protein component); rpmH (ribosomal protein L34); dnaA (chromosome replication initiation protein); dnaN (DNA polymerase III, /~-subunit); recF (inducer of SOS DNA repair); gyrB, gyrA (DNA gyrase, /3 and a subunits). (.4,) Escherichia coli: gyrA is located over 1500 kb distance on the chromosome; (B) Pseudomonas putida: The region corresponding to gyrA has not been sequenced; (C) Bacillus subtilis: A fourth origin is located between gyrA and rrnO; (D) Borrelia burgdorferi: The extent of the inserts of pB3 [6], pB4 [6] and pB22 (this work) are indicated and the sequence data described in this paper (Genbank accession number L14948) is marked by a dashed line with double headed arrows. The direction of transcription of gidA relative to dnaA is not known, while B. burgdorferi homologues of gidB and the 50K and 60K genes have not been isolated. B.subtilis E.coli M.luteus P.putida B.burgdorferi Domain I->l<-Domain II M _ _ _ E N i L D L W N Q A L A Q i E K K L S - K P S F E T W M K S T K A H S L Q G D T L T I T A P N E F A R D W L E S R Y L H L I A D T I YE L T G E E L S I K F V I P Q N Q D V E D F M P K . . . . . . . . . . MS ..... LSLWQQCLARLQDELPATE-F S M W i Rp L Q A E - L S D N T L A L Y A P N R F V L D W V R D K Y L N N INGLLT S-FCGRI APQLRFEVGTKPVTQ---TP QAAVTSNV M V A D Q A V L S S W R S V V G S L E D D A R V S A R L M G F V Y L A Q P Q G L I GN T L L L A V P N E T T R E T L Q G T - - - Q V A D A L T D A L T Q E F R E E I L L A I S I D A N LQP P R TP S S E A R R S S MS ..... VELWQQCVELLRDELPAQQ_FNTWI Rp L Q V E - A E G D E L R V Y A P N R F V L D W V N E K Y L G R L L E L L G E - N G S G IA P A L S L L I G S R R S S A P R A A P N A P V S A A V MEKSKN I___WSLILTEIKKELS-EEEFYVWFENLCFLE S I G D N IKI S T P N L F H K N Q I E K R F T K K I K E ILI K N G Y N N I V I V F . . . . . . . . T N Q P P K T H S N K . . . . . B.subtilis E.coli M. luteus P.putlda B.burgdorferi Domain II->L<-Domain III .......... PQV ........... KKAVKEDTSDFP ................................... QNMLNPK Y TFDTFVI GS GNRFAHAASLAVAEAP AK AAPAQVAQTQPQRAAP ................... STRSGWDNVPAPA ........... EPTYRSNVNVKHT ........ FDNFVEGK SNQLARAAARQVADNP GG LAGGP SGAAAP DVELPPAATAAT SRRAVAEELPGFR IEPPADVVPAANAAPNGNG .... KPTPAPp STSAETSRLNDRYHFETFVI GS SNRFAHAAANAVAEAPAK A A S _ _ L A Q T Q A H K T A P A A A V E P V A V A A A E P V L V E T S S R D S F D A M A E P A A A P P S G G G R A E Q R T V Q V E G A L K H T SY L N R T F T F D T F V E G K S N Q L A R A A A W Q V A D N P K H .................... QETKNPALNETFSKFDKLKEKTT S K E A I QN I Q D R I K M Y I K K E E E E P T N F K N - P F L K K R Y T F E N F I I G P N N K L A Y N A S L S I S K N P G K *. *. * * * *. .. * B.subtilis E.coli M. luteus P.putida B.burgdorferi I<-ATP Binding->1 A Y N P LF I Y G G V G L G K T H L M H A I G H Y V I D H N P S A K V V Y L S S E K F T N E F INS I R D N K A V D F R N R Y R N V D V L L A Y N P LF L Y G GT G L G K T H L L H A V G N G I M A R K P N A K V V Y M H S E R F V Q D M V K A L Q N N A I E E F K R Y Y R S V D A L L A Y N P LF I Y G E S G L G K T H L L H A I G H Y A R R L Y P G L R V R Y V N S E E F T N D F INS I R H D E G A S F K Q V Y R N V D ILL G Y N P LF L Y G G V G L G K T H L M H A V G N H L L K K N P N A K V V Y L H S E R F V A D M V K A L Q L N A I N E F K R F Y R S V D A L L K Y N P C L I Y G G V G L G K T H L L Q S I G N K T E E L H H N L K I L Y V T A E N F L N E F V E S IK T H E T K K F K K K Y R Y L D M L L B.subtilis E.coli M. luteus P.putlda B.burgdorferi Rpp KE i p T L E D R L R S R F E W G L I TD I T P P D L E T R I A RYPKE INGVEDRLKSRFGWGLTVA IEPPELETRVA LPPKQLSGFEDRLRSRFEWGL i TD I Q P P D L E T R I A RYPKE IEGLEERLKSRFGWGLTVAVEPPELETRVA RSP S E L T N F T D R L K S R F T R G L N V D ISKPNFE LRAA B.subtilis E. coli M. luteus P.putida B.burgdorferl S K P K V I T IKE I Q R V V G Q Q F N I K L E D F K A K K R T K S V A F P R Q I A M Y L S R E M T D S SLP Q E - K L V T i DN i Q K T V A E Y Y K I K V A D L L S K R R S R S V A R P R Q M A M A L A K E L T N H SLP E T A H E iT P E L I L H A T G E Y F N L T L E E L T S K S R T R T L V T A R Q I A M Y L L R E L T E M S L P Q D - K L V S V D N i Q R T V A E Y Y K IK I S D L L S K R R S R S V A R P R Q V A M A L S K E L T N HSLP E P N N K I N IENI K K I L L R E L K I T H K D I E G H S K K P E I T K A R H I Y A Y L L R N F T E L S T V E , * , , , I D D I QF L A G K E Q T Q E E F F H T F N T L H E E S K Q IV I S SD I D DI Q F F A N K E R S Q E E F F H T F N A L L E G N Q Q I I L T SD I DDI Q F L A D K E A T V E E F F H T F N T L Y N N N K Q V V IT SD I D DI QF F A R K E R S Q E E F F H T F N A L L E G G Q Q V I LT SD ID D IHDLQKKEG IQEELF HTFNALYEDNKQLVFTCD I L R K K A K A E G L D I P N E V M L Y I A N Q I D S N I R E L E G A L I R V V A Y S S L I N K D I N A D L A A E A L K D I I . . . . . . PS I L M K K A D E N D IR L P G E V A F F I A K R L R S N V R E L E G A L N R V I A N A N F T G R A IT I D F V R E A L R D L L . . . . . . A L I L R K K A E A E G L V A P P E A L E Y I A S R I S T N I R E L E G A L I R V T A F A S L N R Q T V D IE L A E H V L K D LI . . . . . . TD ILMKKADQAKVELPHDAAFF I A Q R I R S N V R E L E G A L K R V I A H S H F M G R D IT IE LI R E S L K D LL . . . . . . A L IV E K K A E E D G I N V P K N IL N L V A Q K V T T N V R D L E A A V T K L K A Y I D L D N I E ID IE I V E K I I K E I I I Y E K E T T N DNA Binding? KIGEEFGGRDHTTVI HAHEKI SKLLADDEQLQQHVKE IKEQLK ...... E IGDAFGGRDHTTVLHACRK I E Q L R E E S H D I K E D F SN LI R T L . . . . . SS KIGQVLGGRDHTTVI HADRKIRELMAERRT IYNQVTELTNE IKRKQRGA E IGDMFGGRDHTTVLHACRK I N E L K E S D A D I R E D Y K N L L R T L . . . . . TT IGKI IGGKTHSTVLYS INKIDRDRNNDKE INNLI TELMNKIKKN .... ** ** , ** ** . . . . . B C B.subtilis B.aphidicola E.coll M. luteus M. caprlcolum P.mirabilis P.putida S.marcescens S. typhimurium S.coellcolor I<-ATP B i n d i n g - > l IYGGVGLGKTHLMHAI LYGGTGLGKTHLLHAI LYGGTGLGKTHLLHAV IYGESGLGKTHLLHAI IYGESGMGKTHLLKAA LYGGTGLGKTHLLHAV LYGGVGLGKTHLMHAV LYGGTGLGKTHLLHAV LYGGTGLGKTHLLHAV IYGESGLGKTHLLHAI .** *.*****. , DNA Binding? IGEEFGGRDHTTVIHAHEKI IGDAFSGRDHTTVLHACRKI IGDAFGGRDHTTVLHACRKI IGQVLGGRDHTTVIHADRKI IGEEFGGRDHTTVINAERKI IGDAFGGRDHTTVLHACRKI IGDMFGGRDHTTVLHACRKI IGDAFGGRDHTTVLHACRKI IGDAFGGRDHTTVLHACRKI IGALFGGRDHTTVMHADRKI B.burgdorferi IYGGVGLGKTHLLQSI .** ******. . IGKIIGGKTHSTVLYSINKI ** * *.**. . ** Fig. 2. Alignment of DnaA sequences. The amino acid sequences, presented in the one letter code, were aligned using Clustal V. Identical residues are marked with an asterisk ( * ) while similar sequences are denoted by a point (.) below the alignment. (A) Alignment of 5 different D n a A proteins from representative eubacteria: Bacillus subtilis, and Micrococcus luteus, respectively low and high GC% Gram-positive bacteria, Escherichia coli and Pseudomonas putida, Gram-negative bacteria, and Borrelia burgdorferi. The three different domains (1, 1I, III) are marked above the alignment as is the ATP binding site and a possible DNA binding site. (B) Conservation of the ATP binding site of DnaA. The relevant regions of 11 different DnaA proteins are aligned. The amino acid sequences are from Bacillus subtilis, Buchnera aphidicola, Escherichia coli, Micrococcus luteus, Mycoplasma capricolum, Proteus mirabilis, Pseudomonas putida, Salmonella typhimurium, Serratia marcescens, Streptomyces coelicolor [1,10,14-16] and Borrelia burgdorferi (this work). For further references see [1]. (C) Lack of conservation in a possible D N A binding site of ~o 113 found no O R F on pB22 with similarity to recF (data not shown). The order dnaN-dnaA-gyrBgyrA is in agreement with that presented by others at the Fifth International Conference on Lyme borreliosis [12]. This is in contrast to the usual eubacterial arrangement dnaA-dnaN-recF-gyrB (Fig. 1A-C). It would appear that, in addition to a relative translocation of gidA [6], the B. burgdorferi dnaA and dnaN genes have been inverted and recF has been either lost or transposed elsewhere during the evolution of the B. burgdorferi linear chromosome. The proximity of dnaA to gyrBA suggested the gene lay between 457 and 488 kb on the B. burgdorferi genetic map [6] and this was confirmed by Southern hybridisations (data not shown). As replication origins are found next to dnaA in Gram-positive bacteria and Pseudomonads (Fig. 1B,C), we searched the gyrB-dnaA and dnaA-dnaN intergenic regions for DnaA boxes. We were unable to detect clusters of DnaA boxes characteristic of an oriC region [2] in the gyrBdnaA intergenic region while there was only one DnaA box (AAATCCACA) in the dnaA-dnaN intergenic region. The absence of any DnaA box in the dnaA upstream region might suggest that B. burgdorferi dna,4 is not autoregulated. As the dnaA and dnaN genes have been inverted relative to rprnH and gyrB, another possible origin location is between dnaN and rpmH. Initial analysis of 500 bp of sequence upstream of rpmH have revealed only two DnaA boxes (data not shown). Although a central location for the origin of replication on a linear chromosome is appealing, we cannot preclude the possibility that an origin is next to gidA (Fig. 1D). Location of the origin of replication was also hampered in Buchnera aphidicola, a Gram-negative intracellular symbiont of aphids with 28-30% GC. As in B. burgdorferi there is no recF gene, no DnaA boxes were detected in the dnaA region [10], and the D N A binding region may be atypical (Fig. 2C). It is not known whether the origin is located elsewhere, as in E. coli and Proteus mirabilis, or whether the DnaA boxes are atypical in this organism [10]. A feature of the oriC region of E. coli is the presence of some 20 G A T C sites. The state of methylation of G A T C sites in the origin region is an important factor in the regulation of chromosome replication in enteric bacteria such as E. coli and P. mirabilis, which possess an adenine methylation system, while organisms lacking the Dam system do not appear to be regulated in this manner [2]. Strain 212 (our unpublished results), like the majority of Borrelia burgdorferi (sensu lato) strains, does not possess an adenine methylation system [13]. It is therefore unlikely that B. burgdorferi uses the adenine methylation system for control of chromosome replication. Indeed, we found only 5 G A T C sites in 4.7 kb of sequence in the B. burgdorferi dnaA region. We conclude that B. burgdorferi is unique amongst the eubacteria: it has a linear chromosome, the arrangement of genes in the dnaA region is atypical, as is a possible D N A binding motif in the C-terminus of the DnaA protein and no arrangements of DnaA boxes characteristic of a eubacterial ori region were found in the dnaAdnaN or dnaA-gyrB intergenic regions. We are unable to conclude whether this is because the B. burgdorferi origin is also atypical or because it is located elsewhere, perhaps beside gidA, as in E. coli (Fig. 1A). We are currently examining these possibilities. Acknowledgements We are grateful to Barrie Davidson for helpful discussions and to Guy Baranton for his continued interest in this work. This work was supported by grants from the Gould Foundation to the Institut Pasteur. References 1 Yoshikawa, H. and Ogasawara, N. (1991) Structure and function of DnaA and the DnaA-box in eubacteria: evolutionary relationships of bacterial replication origins. Mol. Microbiol. 5, 2589-2597. 2 Smith, D.W., Yee, T.W., Baird, C. and Krishnapillai, V. (1991) Pseudomonad replication origins: a paradigm for bacterial origins? Mol. Microbiol. 5, 2581-2587. 3 Holz, A., Schaefer, C., Gille, H., Jueterbock, W.R. and Messer, W. (1992) Mutations in the DnaA binding sites of 14 4 5 6 7 8 9 the replication origin of Escherichia coli. Molec. Gen. Genet. 233, 81-88. Ogasawara, N. and Yoshikawa, H. (1992) Genes and their organisation in the replication origin region of the bacterial chromosome. Mol. Microbiol. 6, 629-634. Davidson, B.E., MacDougall, J. and Saint Girons, I. (1992) Physical map of the linear chromosome of the bacterium Borrelia burgdorferi 212, a causitive agent of Lyme disease, and localization of rDNA genes. J. Bacteriol. 174, 37663774. Old, I.G., MacDougall, J., Saint Girons, I. and Davidson, B.E. (1992) Mapping of genes on the linear chromosome of the bacterium Borrelia burgdorferi: possible locations for its origin of replication. FEMS Microbiol. Lett. 99, 245-250. Old, I.G., Margarita, D. and Saint Girons, I. (1992) Nucleotide sequence of the Borrelia burgdorferi rpmH gene encoding ribosomal protein L34. Nucleic Acids Res. 20, 6097. Devereux, J. (1991) The GCG sequence analysis software package, version 7, Genetics Computer Group Inc., Madison, Wisconsin, Fujita, M.Q, Yoshikawa, H. and Ogasawara, N. (1990) Structure of the dnaA region of Micrococcus luteus: conservation and variations among eubacteria. Gene 93, 7378. 10 Lai, C.-Y. and Baumann, P. (1992) Genetic analysis of an aphid endosymbiont DNA fragment homologous to the rnpA-rpmH-dnaA-dnaN-gyrB region of eubacteria. Gene 113, 175-181. 11 Fujita, M.Q., Yoshikawa, H. and Ogasawara, N. (1992) Structure of the dnaA and DnaA-box region in the Mycoplasma capricolum chromosome: conservation and variations in the course of evolution. Gene 110, 17-23. 12 Casjens, S. and Huang, W.M. (1992) Type II DNA topoisomerase genes in Borrelia. V International Conference on Lyme borreliosis. Arlington, USA, p. A27. 13 Hughes, C.A.N. and Johnson, R.C. (1990) Methylated DNA in Borrelia species. J. Bacteriol. 172, 6602-6604. 14 Skovogaard, O. (1990) Nucleotide sequence of a Proteus mirabilis DNA fragment homologous to the 60K-rnpArpmH-dnaA-dnaN-recF-gyrB region of Escherichia coli. Gene 93, 27-34. 15 Skovogaard, O. and Hansen, F.G. (1987) Comparison of dnaA nucleotide sequences of Escherichia coli, Salmonella typhimurium and Serratia marcescens. J. Bacteriol. 169, 3976-3981. 16 Calcutt, M.J. and Schmidt, F.J. (1992) Conserved gene arrangement in the origin region of the Streptomyces coelicolor chromosome. J. Bacteriol. 174, 3220-3226.