Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
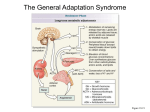
St Petersburg Scientific Forum ‘Science and Society: Physiology and Medicine of the 21st Century’ St Petersburg, Russia, September 19–23, 2011 From Hans Selye’s discovery of biological stress to the identification of corticotropin-releasing factor signaling pathways: implication in stress-related diseases Yvette Taché Center for Neurobiology of Stress and Digestive Diseases Research Center, Digestive Diseases Division, UCLA and VA Greater Los Angeles Health Care System, Los Angeles, CA, USA The term “stress” was first coined by the endocrinologist Hans Selye more than 70 years ago [1] to define the physiological adaptive responses of the organism to emotional or physical threats (“stressors”). While the stress response is adaptive in nature, when the severity and/or chronicity of the stressors are exceeding the limits, and the adaptive system becomes defective or excessive, the organism is no longer brought back to basal homeostasis leading to a state of allostatic load [2] also named “cacostasis” [3] which lies at the origin of a variety of stress-related diseases that develop in the context of a vulnerable background (genetic, epigenetic and/or constitutional). The pathogenesis of stress-induced disorders affects the whole body, including the viscera of which the gastrointestinal (GI) tract is a sensitive target [1]. In particular, dysregulation of the stress circuit affects motor function of the gut, contributes to visceral hypersensitivity as observed in patients with irritable bowel syndrome (IBS) and increases incidence to develop post infectious IBS following bacterial gastroenteritis. In recent decades, the biochemical coding of the stress response was unraveled through the identification of the 41 amino acid peptide, corticotropin releasing factor (CRF), and related peptides, urocortin 1 (ucn 1), ucn 2 and ucn 3, the characterization of CRF receptors CRF1 and CRF2 and their variants which display specific affinity for CRF and related agonists along with the development of selective CRF receptor subtype antagonists [4]. When a stressor is perceived, a convergence of stimulatory inputs from different brain regions (amygdala, prefrontal cortex, pons, medulla) activates the paraventricular nucleus of the hypothalamus (PVN) which releases CRF which binds to CRF1 receptors located on corticotrope cells of the pituitary gland to release the adrenocorticotropic hormone leading to glucocorticoids secretion from the adrenal glands. Far beyond an exclusive neuroendocrine role, CRF, which is widely distributed outside of the hypothalamus, also acts as a neurotransmitter/ neuromodulator to coordinate the behavioral, autonomic, im62 Yvette Taché 63 mune, and visceral efferent limbs of the stress response. For instance brain CRF activates the sympathetic nervous system inducing the systemic release of catecholamines involved in the ”fight or flight” response. The locus coeruleus is also activated by CRF and its noradrenergic projections to the forebrain contributes to the arousal and alertness. Convergent preclinical evidence has accumulated suggesting that stress-related alterations of colonic motor and hypersensitivity to colorectal distention (CRD) are primarily mediated by the activation of brain CRF/CRF1 signaling pathway, while CRF2 receptor activation may exert a counteracting influence [5,6]. In addition, recent experimental and clinical studies point to an equally important contribution of the peripheral CRF/CRF1 signaling locally expressed in the gut as a key component of the local arm of the colonic response to stress [7]. CRF gene is expressed in the colon and CRF immunoreactivity (IR) is located in the epithelia, lamina propria and crypts, and co-localized with tryptophan hydroxylase, a marker for enterochromaffin cells, and in enteric neurons. When CRF is injected peripherally, the peptide selectively activates myenteric plexus of the colon as shown by Fos immunoreactivity, a marker of neuronal activation, while no Fos was found in other gut segment myenteric neurons. Fos immmunoreactivity induced by CRF was located in CRF1 receptor-IR myenteric neurons and 40% of cholinergic neurons with Dogiel type I morphology, and in 20 and 80% of nitrergic neurons in proximal and distal colon respectively. In addition, the CRF1 receptor agonists, cortagine or stressin1 injected peripherally in rodents recaptures cardinal features of IBS-diarrhea predominant patients (stress-related hyperalgesia to CRD, increased colonic permeability, mast cell degranulation, activation of the colonic enteric nervous system, motility, transit, and defecation, induction of diarrhea and bacterial translocation). By contrast, the activation of CRF2 receptors attenuated the CRF1 mediated colonic and hyperalgesic responses to CRF1 agonists or stress [5,6]. These data suggest that an overactivity of CRF1 signaling and/or defective mounting of the CRF2 inhibitory mechanisms may play a role in IBS manifestations. Hans Selye, also stated that “an intense psychic shock may exert pronounced effects on the hair, e.g., graying and generalized loss of hair”. Subsequent experimental and clinical evidence indicated that chronic stress can inhibit hair growth and demonstrated the expression of CRF signaling system in normal skin and cycling hair follicles. We used CRF over-expressing (OE)-mice that display phenotype of chronic stress including alopecia. The non-selective CRF receptors antagonist, astressin-B (5 μg/mouse) injected peripherally once a day for 5 days in 4-9 months old CRF-OE alopecic mice induced pigmentation and hair re-growth that was largely retained for over 4 months. In young CRF-OE mice, astressin- 64 From Hans Selye’s discovery of biological stress. . . B prevented the development of alopecia that occurred in saline-treated mice. Histological examination indicated that alopecic CRF-OE mice had hair follicle atrophy and that astressin-B revived the hair follicle from the telogen to anagen phase. However, astressin-B did not show any effect on the elevated plasma corticosterone levels and the increased weights of adrenal glands and visceral fat in CRF-OE mice. The selective CRF2 receptor antagonist, astressin2 -B had moderate effect on pigmentation, but not on hair re-growth. The commercial drug for alopecia, minoxidil showed partial effect on hair re-growth. Therefore these data support the existence of a key molecular switching mechanism triggered by blocking peripheral CRF receptors to reset hair growth in a mouse model of alopecia associated with chronic stress [8]. References [1] [2] [3] [4] [5] [6] [7] [8] Selye H, Nature 1936; 138: 32. McEwen BS, Ann. N. Y. Acad. Sci. 1998; 840: 33–44. Chrousos GP, Nat. Rev. Endocrinol. 2009; 5: 374–381. Hauger RL et al. Pharmacol. Rev. 2003; 55: 21–26. Taché Y, Bonaz B, J. Clin. Invest 2007; 117: 33–40. Gourcerol G et al. Gastroenterology 2011; 140: 1586–1596. Larauche M, Kiank C, Taché Y, J. Physiol Pharmacol. 2009; 60 Suppl 7: 33–46. Wang L et al, PLoS. One. 2011; 6: e16377. Supported by NIH DK 57238 and 33061.