Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
/ . Embryol. exp. Morph., Vol. 11, Part 4, pp. 697-714, December 1963
Printed in Great Britain
The Development of Somites in the Chick Embryo
by RUTH BELLAIRS 1
From the Department of Anatomy, University College London
WITH ONE PLATE
INTRODUCTION
T H I S paper is concerned with the influences which cause undifferentiated mesoderm to become converted into somites in the chick embryo. The experiments
that are reported were designed to test several theories which already exist in
the literature. Each of these theories ascribes an essential, inductive role to
one of the following regions:
1. The 'somite centres*. Spratt (1955) put forward the idea that two 'somite
centres' exist in the chick blastoderm and that these actively induce the
formation of somites. His evidence is based mainly on a series of transection experiments.
2. HenserCs node. This theory is based on the results of experiments in which
the node is damaged or extirpated (e.g., Peebles, 1898; Wetzel, 1929;
Fraser, 1954). The literature already contains evidence against this
theory.
3. The neural tissue. This theory, which has been put forward by various
authors (e.g. Griinwald, 1936; Fraser, 1960), is based on the fact that
neural tissue and somites are usually closely associated.
The test that I have applied to each theory involves the following argument.
If a particular tissue (or region) plays an essential role in somite formation,
then somites cannot develop in the absence of that tissue. In the present paper
it will be shown that somites can form in the absence of the 'somite centres',
of the node, and of neural tissue. In no case, however, have they been found to
develop in the absence of regression movements.
MATERIAL AND METHODS
One hundred and nineteen chick blastoderms were removed from the egg
and explanted in vitro. The blastoderms were taken from eggs which had
been incubated for between 18 and 24 hr. and were usually at the full-length
1
Author's address: The Department of Anatomy, University College, Gower Street,
London W.C.I., U.K.
698
R U T H BELLAIRS
primitive streak stage, the head process stage or the early head fold stage (i.e. the
L-, h.p.- and h.f.-stages respectively of Waddington, 1932, and of Abercrombie,
1950). Most of the experiments consisted of extirpating parts of the area pellucida (see Text-figs. 2, 5, 8), though some transplantation experiments were also
performed (see Text-fig. 9).
The culture technique of New (1955) was used, which consists of growing the
blastoderm on a piece of vitelline membrane stretched out tautly over a
glass ring. Normally, if no operation is performed on the blastoderm it expands
to cover the available area of membrane, this expansion being due largely to
the activity of the peripheral cells of the area opaca (New, 1959; Bellairs &
New, 1962; Bellairs, 1963). If there is a small wound in the area pellucida,
however, it tends to become pulled into a large hole by the force of the expansion. In extreme cases the area pellucida is reduced to a narrow ring of cells
surrounding a large hole. In most of the present experiments therefore, a
narrow band, about 1 mm. wide, was trimmed from the periphery of the
blastoderm and this reduced the expansion.
After the operation had been performed, each specimen was incubated in vitro
for about 24 hr. and then examined. If development had taken place, the
specimen was fixed in Bouin's fluid, serially section at 10/t and stained with
eosin and Ehrlich's haematoxylin. The criterion taken for somite differentiation
was that segmentation should have occurred. The actual size of individual
somites or their number was considered to be relatively unimportant since it is
related to both speed of development during the period of incubation, and to
the amount of mesoderm available. A minimum of four distinct somites or
pairs of somites was, however, considered necessary to demonstrate segmentation.
The presumptive somite areas and the 'somite centres'
Maps of the presumptive areas at the full length primitive streak stage and
later have been provided by a number of authors (e.g. Wetzel, 1929; Graper,
1929; Pasteels, 1937; Waddington, 1952; Spratt, 1955; Vakaet, 1962). The
main differences in interpretation are concerned with the question of how much
mesoderm has already invaginated by this stage.
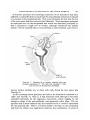
Text-fig. 1A shows the arrangement of the presumptive notochord and somites
at the early head process stage, according to the fate map of Pasteels (1937).
This stage is immediately followed by the regression movements (Text-fig. IB)
during which the embryonic axis forms anteriorly whilst the primitive streak
shortens posteriorly. These movements include the backward migration of the
node (relative to fixed points on, say, the area opaca) and to a lesser extent of
the regions of the area pellucida on either side of the node. (See also schemes
by Spratt, 1947; Vakaet, 1962.)
The regions which Spratt (1955) has called 'somite centres' are shown in
Text-fig. 1C. They lie 'one on each side of the posterior half of the node.
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
699
Each extends slightly (0-05-0-02 mm.) under the node and laterally for about
0-2 to 0-25 mm.' (Spratt, 1955).
B
1. A: The presumptive areas of notochord, somite and lateral
plate mesoderm at the early head process stage (after Pasteels, 1937).
B: The regression movements during the development of the head process and head fold (after Pasteels, 1937). C: The location of the' somite
centres' at the head fold stage (after Spratt, 1955).
TEXT-FIG.
RESULTS
Experiment 1
This experiment was designed to test the ability of presumptive somite mesoderm to develop when the 'somite centres' had been removed, but the regression
RUTH BELLAIRS
700
movements could still take place. The anterior part of the area pellucida was
extirpated except for a narrow strip of primitive streak (see Text-fig. 2, A & B).
Control
Series
Experiment I
D
Discarded
Control Series b
2. Diagram illustrating the position of the cuts made in experiment 1 and the accompanying control experiments at the L-stage (A & C)
and the h.p.-stage (B & D). A & B: Main experiment (stipple) and control series a (white). C & D: Control series b (fine stipple).
TEXT-FIG.
This strip was retained in order to promote regression movements (see discussion). The length of the strip was between one third and one half that of the
primitive streak (i.e. the strip was usually between about 0-6 and 1-0 mm.
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
701
long at the L-stage, the anterior cut passing through the anterior border of the
node). The strip was, however, made as narrow as possible (i.e. about0-1 mm.
wide) in order to eliminate completely the 'somite centres'; i.e. the longitudinal
cuts passed through the primitive folds rather than lateral to them. (A similar
experiment is described by Butros, 1962, except that in his specimens the longitudinal cuts were made some distance lateral to the primitive streak and did
not therefore eliminate the 'somite centres'; indeed, they were not designed to
do this.)
Thirty-three specimens were operated upon, nineteen at the L-stage, fourteen
at the head process or head fold stage. In addition, two series of control experiments were performed. Control series a consisted of culturing the remaining
anterior part of the area pellucida (Text-fig. 2, A & B), a region which contained some presumptive somite material (see Text-fig. 1); thus if somites failed
to form in both anterior (control) and posterior (experimental) isolates it was
considered that the embryo was either defective or so retarded as to be unreliable.
Control series b is described below (see also Text-fig. 2, C & D). The results of
the experiment are given in Table 1.
TABLE 1
Summary of the results of experiment 1 and of the associated control series a
and of control series b.
Presence of somites is indicated by + , absence of somites by —
Number of
specimens
Experiment 1 and
control series a
Total:
Control series b
f 8
\ 3
Stages of
specimens
6L:2h.p.
1 L:2h.p.
5L:3h.p.
5L:3h.p., 2 h.f.
2L:2h.p.
Anterior piece Posterior piece
{Control series a) {Experiment X)
—
+
+
Damaged or lost
—
—
+
+
+
33
8
5L:3h.p.
+
-
Eight specimens failed to form somites and well defined axes in both anterior
and posterior portions; six of these specimens were oedematous. Three other
specimens had formed somites in the anterior piece but not in the posterior
piece; in two of these the main horizontal cuts were made very far posteriorly,
the length of the strip being 1 -6 mm.; it is possible therefore that in these embryos little, if any, presumptive somite material remained in the posterior piece.
The remaining twenty-two specimens all formed somites in the posterior piece
of area pellucida.
Text-figure 3 is a drawing of a typical axis obtained from a posterior isolate.
The explant has become crescent shaped, somites and notochord lying along the
45
RUTH BELLAIRS
702
inner margin of the crescent. In six specimens a small clump of carbon particles was placed on the endoderm of the node and in four of these the mark
3. Drawing of a whole mount obtained after
performing experiment 1. A single row of somites has
developed.
TEXT-FIG.
carbon marks
4. Drawing of a whole mount obtained after
performing experiment 1. Note that a carbon mark was
placed on the anterior tip of the projecting strip and that
part of the mark has become displaced posteriorly. A
double row of somites is present and a V-shaped indentation lies to one side of it.
TEXT-FIG.
was subsequently found to have become displaced along the margin of one horn
of the crescent. Text-figure 4 is a drawing of one of the marked specimens; it
will be noted that a V-shaped indentation lies to one side of the axis.
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
703
Serial sections showed that typically these axes possess a notochord as well
as somites. Small pieces of neural plate are sometimes present, but in some
embryos neural tissue is absent along either the entire length of the somites
(three specimens), or along the greater part of the length (two specimens).
Control series b was carried out by transecting the blastoderm directly across
TEXT-FIG. 5. Diagram illustrating the position of the cuts made in
experiment 2 at the L-stage (A) and the h.p.-stage (B) respectively.
the primitive streak in eight specimens (see Text-fig. 2, C & D). The cut was
made so that the anterior third or half of the primitive streak was not included
in the posterior portion. These specimens showed no sign of somite differentiation in the posterior piece, whereas all of them had somites in the anterior
piece. Seven of the posterior pieces possessed a V-shaped indentation (see
Text-fig. 10). Serial sections showed that 24 hr. after operation the area pellucida of the posterior explant consisted of only ectoderm, endoderm and lateral
plate mesoderm. This control experiment has been performed previously by
other workers and the results confirm their findings (e.g. Waddington, 1935;
Spratt, 1955).
Experiment 2
This experiment was designed to remove the 'somite centre' from one side
of the area pellucida only, whilst permitting regression movements to take
place (see Text-fig. 5). Thirty-seven of these operations were performed. The
length of the piece removed varied from 0-35 to 1-6 mm. (average 0-6 mm.);
the width of the piece varied from 0-35 to 1-0 mm. (average 0-52 mm.). The
extirpation was made from the right side in twenty-eight specimens and from
RUTH BELLAIRS
704
the left in nine, but this fact did not appear to affect the type of result obtained.
The most crucial factor was apparently the fate of the hole in the tissues (see
Table 2).
TABLE 2
Summary of the results of experiment 2
Number of
specimens
12
14
2
7
2
Stages of
specimens
9 L:3 h.p.
6 L:5h.p.:3h. P2 L
4 L:2h.p.:l
1 L:l h.f.
h .f.
Fate of the
hole
Healed
Greatly enlarged
Split
Split
Split
Rows of
somites
2
1
0
2
1
Total: 37
forebrain
heart
edge of
area pellucida
6. Drawing of an embryo obtained after performing experiment 2. The hole has become so stretched
that the axis is confined to a narrow crescent of tissue on
the unoperated side and a single row of somites is present.
TEXT-FIG.
In twelve specimens the hole healed completely and a normal embryo with
notochord, neural tube and paired somites developed. Moreover, the area
pellucida had changed shape and elongated.
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
705
In fourteen specimens the centrifugal expansion of the blastoderm had been
sufficient to extend the hole so much that the area pellucida had become reduced
to a crescent at the unoperated side. This occurred despite the fact that the rim
of the area opaca had been removed (see Methods). These specimens possessed
an embryonic axis on the unoperated side which was abnormal principally in
that it possessed a single row of somites, although notochord was usually
present. On the operated side no area pellucida as such remained, apart from a
forebrain
heart
heart
split
edge of
area pellucida
TEXT-FIG. 7. Drawing of an embryo obtained after performing experiment 2. The hole has extended a little,
but two rows of somites have developed.
narrow border, perhaps two or three cells wide, lining the area opaca (see
Text-fig. 6).
In the remaining eleven specimens the hole in the blastoderm remained as a
'split' (see Text-fig. 7). That is, it had remained small although it had been
extended backwards by the regression movements, which, judging by the
change in shape of the area pellucida, had apparently taken place. The unoperated side in these embryos may be considered to be a 'control' experiment
for the operated side. That is, if somites failed to develop on the unoperated
side then the embryo was regarded as defective (two specimens). In seven of
RUTH BELLAIRS
706
the remaining nine specimens somites formed on both the operated and unoperated sides (see Text-fig. 7). In each case a row of somites lay down either
side of the split, but the two rows were apparently not arranged in regular
pairs except in the region posterior to the split. Sections showed that on the
operated side the somites had no notochord associated with them; five of the
seven specimens also had no neural tissue on the operated side, the remaining
two possessing only tiny strips of neural plate (see Plate 1, fig. A).
In two further specimens the somites failed to form on the operated side, a
single row developing from the unoperated side. One of these had formed a
very distorted axis.
B
8. Diagram to illustrate the position of the cuts made in
experiment 3 at the L-stage (A) and the h.p.-stage (B).
TEXT-FIG.
Experiment 3
This experiment was designed to eliminate the greater part of the notochord
by extirpating the node (see Text-fig. 8). The piece of tissue removed was
between 0 • 2 and 0 • 5 mm. long (average 0 • 3 mm.) and between 0 • 2 and 0 • 3
mm. wide (average 0-28 mm.). This experiment has been performed previously
by several authors (see discussion). Only ten specimens were therefore treated
in this way in the present investigation. The results are summarized in Table 3.
In two specimens which failed to heal the hole was so enlarged that no axial
tissues formed at all. The remaining eight specimens were sectioned and each
showed regions in which well defined somites were present even though notochord was absent. In all eight specimens, elongation of the area pellucida had
occurred.
J. Embryol. exp. Morph.
Vol. 11, Part 4
B
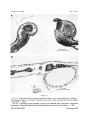
PLATE 1
FIG. A. Transverse section across a specimen similar to the one illustrated in Text-fig. 7
The operated side is on the left. Note that no neural tissue is associated with the somite
at this side, x 200.
FIG. B. Transverse section through a dwarf axis obtained after experiment 4 had been
performed. Note that somites have developed although neural tissue is absent, x 240.
RUTH BELLAIRS
(Facing page 706)
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
707
TABLE 3
Summary of the results of experiment 3
Number of
specimens
Stages of
specimens
Fate of the
hole
Rows of
somites
2
5
1
2
1 L:l h.p.
2L:2h.p. 1 h.f.
L
2L
Greatly enlarged
Split
Healed
Healed
0
2
1
2
Total: 10
As a 'control' experiment the extirpated graft was replaced in seven specimens
and in each of these a normal, single embryo formed complete with notochord
and paired somites.
Experiment 4
This experiment was carried out in collaboration with Professor M. Abercrombie as part of a different investigation, the results of which have not yet
been published in detail.
9. Diagram to illustrate the operation performed in
experiment 4 at the L-stage (A) and the h.p.-stage (B). The posterior rectangle was cut out and replaced by a graft from the
node region.
TEXT-FIG.
The operation consisted of cutting out a small rectangle from the posterior
end of the primitive streak and replacing it by a graft taken from the node
(Text-fig. 9). Both pieces of tissue included the whole thickness of the primitive
streak, ectoderm, mesoderm and endoderm. The size of the graft was about
708
R U T H BELLAIRS
0-1 to 0-2 mm. wide by 0-2 mm. long and the experiment was carried out at
either the L-stage or the h.p.-stage.
Twenty-four experiments of this type were performed and, in thirteen, regulation apparently took place for no trace of the graft could subsequently be
found. In eleven specimens, however, supernumerary axial tissues were formed
although they were never present as a complete, well-arranged axis. On sectioning, it was found that four of them had no neural tissue, although they possessed
a small notochord and miniature but well-segmented somites (Plate 1, fig. B).
DISCUSSION
The 'somite centres'1 .
Spratt (1955) regards the 'somite centres' as two regions, one on either side
of the node, which regress along with the node and induce somites from presumptive somite mesoderm. He states (p. 160): 'Removal of the somite centers
(including the overlying part of the neural plate and the hypoblast below) results in the cessation of further somite formation posterior . . . to the centers.'
His evidence is based on various types of experiment. In the main experiment
he transected the area pellucida, and found that if the cut was about 0 • 3 mm. posterior to the primitive pit, somites failed to form in the posterior part. The same
experiment was performed in the present investigation (control series b of
experiment 1) and the same result was obtained. Spratt concluded that somites
failed to form because the posterior presumptive somite material was not in
contact with the somite centres.
The present investigation (experiment 1) shows, however, that the posterior
isolate can form somites, in the absence of the somite centres, provided only
that a narrow strip of primitive streak remains attached. It seems probable
that in Spratt's experiments the regression movements were inhibited, whereas
in the ones reported in this paper they were not (see below). Spratt himself
regards the regression movements as important, though his experiments have
not been specifically designed to distinguish between the importance of somite
centres and regression.
Spratt's second line of evidence is derived from experiments in which he
extirpated square or rectangular pieces of the node and surrounding tissue.
After cutting out almost the entire somite centre on one side (see his Fig. 5,
operation 6), he failed to obtain somites posterior to the hole in four out of
nine specimens. It is difficult to accept this as evidence of the essential nature of
the somite centre however, for in the remaining five specimens Spratt obtained
somites posterior to the hole. Indeed, he sometimes obtained extra somites.
Similarly it has been shown in experiment 2 in the present investigation that,
if the ' somite centre' is extirpated, somites frequently form from mesoderm
posterior to the wound. Moreover, Spratt himself has shown that somite
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
709
formations can occur in the anterior part of the blastoderm following extirpation of the somite centre.
It is concluded from the results of experiments 1 and 2 of this investigation
that somites can differentiate from presumptive somite mesoderm in the absence
of the presumed somite centres. Hence the somite centres are not essential for
somite formation.
Henserfs node and no to chord
Hensen's node contains most, if not all of the presumptive notochord material
(Pasteels, 1937; Spratt, 1947). Removal of the node, therefore, frequently leads
to the production of embryos which are deficient in notochord. Many authors
have reported the presence of somites in such embryos (e.g. Waddington, 1932;
Waterman, 1936; Wolff, 1936; Abercrombie & Bellairs, 1954; Spratt, 1955;
19576; Grabowski, 1956; Fraser, 1960); and some of them have noted that if
the wound heals and notochord is absent then somites tend to fuse into a
single row in the mid-line. Similar results were obtained in the present investigation, and they serve to support the now widely held opinion that neither
the node nor notochord are necessary for somite formation. It is of interest in
this connexion that attempts by Fraser (1960) to induce somites from dissociated
mesenchyme cells by the insertion of notochord grafts produced at best a nonspecific clumping of cells.
The neural tissue
The possibility that the neural tissue plays a role in somite induction has
been suggested by several authors (e.g. Griinwald, 1936; Waddington, 1935;
Spratt, 1957a; Fraser, 1960). The evidence is based mainly on the fact that the
two tissues are usually closely associated, but attempts by Fraser to induce
somites with neural tissue produced no clear-cut result. In the present investigation, however, seven specimens have been obtained which are totally
deficient in neural tissue but have nevertheless developed somites. In five other
specimens a row of somites has developed which is separated from any neural
tissue by a hole in the blastoderm. It is considered that these specimens demonstrate that somites are not dependent on neural tissue for formative influences.
The endoderm and lateral plate
The endoderm and lateral plate mesoderm lie respectively ventral and lateral
to the developing somites. It is conceivable, therefore, that either one or both
of these tissues may play a role in somite formation, though I have not found
any suggestion to this effect in the literature. However, Waddington (1932)
obtained somites from endoderm-free epiblasts when they were grown together
in pairs. Similarly, one embryo was obtained in the present investigation in
which somites had developed which were devoid of contact with lateral plate.
710
RUTH BELLA1RS
On the basis of this scanty evidence it seems unlikely, therefore, that either
endoderm or lateral plate will prove to be essential for somite formation.
The Regression Movements
It is suggested as a result of the present investigation that regression movements play an important role in somite formation. The importance of regression
movements has been mentioned by previous authors, though merely as a corollary to one or other of the theories outlined above (e.g. Spratt, 1955; Fraser,
1960). Details of regression movements have been worked out by Pasteels
(1937) (see Text-fig. 1). The movements start near the anterior end of the area
pellucida and spread posteriorly, being maximal at the node and along the
primitive streak. The regression of the node results in the laying down of the
notochord. The area pellucida as a whole also changes shape and becomes
elongated backwards. Thus development of notochord and/or posterior
elongation of the area pellucida may be taken as evidence that regression movements have occurred.
The anterior end of the primitive streak has been shown to undergo regression
movements if it is either isolated (e.g. Spratt, 1957a) or grafted (Abercrombie
& Waddington, 1937). Nevertheless this regression is not entirely under its
own control, for Abercrombie (1950) found that the direction of the regression
movements in the streak can be reversed by the area pellucida as a whole.
The results of the present experiments will now be considered in relation to this
theory.
Experiment 1
There seems little doubt that regression occurred in the operated specimens
which developed somites, for these generally possessed notochord, a structure
which forms only by regression of the node. In order to decide whether the
regression is important as a factor in somite formation, it is necessary to know
if it occurred also in the control series b (see Text-fig. 2, C and D). The latter
experiment was originally performed by Waddington (1932, 1935) and has
since been repeated by Spratt (1955) and by Butros (1960, 1962). All authors
agree that somites fail to form unless the cut is made immediately behind the
node, and that a V-shaped opening develops in the anterior (cut) edge of the
isolate. Waddington (1935) and Butros (1962) regard this opening as evidence
that regression movements have occurred, but Jacobson (1938) has put
forward the idea that the V-shape is formed by a stretching or retraction of the
cut edge. The latter view is supported by the present results which show that a
V-shaped formation may occur independently of the primitive streak and hence
of the region of maximum regression movements (Text-fig. 4). Despite the
removal of the outer rim of the area opaca (see Methods) some centrifugal
expansion apparently takes place in these experiments; since only half the
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
711
blastoderm is present, however, the tension exerted is likely to be unequally
distributed so that the area pellucida may become distorted and the V-shape
form to one side of the primitive streak. An interpretation of the effect of this
type of expansion on posterior isolates (experiment 1 and accompanying controls) is shown in Text-fig. 10. It is concluded, therefore, that if the anterior
part of the primitive streak remains, then regression occurs and somites develop ; if the anterior part of the primitive streak is cut off, no regression takes
place and no somites form.
TEXT-FIG. 10. Diagram illustrating the probable effect of expansion and regression in
experiment 1 (C & D) and its accompanying control series b (A & B). It is suggested that
somites develop only if regression occurs along the primitive streak. In both experimental
and control series, however, centrifugal expansion movements may lead to the formation
of a V-shaped indentation.
Experiment 2
In experiment 2, also, the expansion movements may stretch the wound so
much that the area pellucida on the operated side becomes reduced to a narrow
band of cells. It is unlikely that normal regression movements could take place
on this side under such circumstances, and it is regarded as significant therefore
that no somites developed in the operated side of these specimens.
When gross distortion of this type did not happen, however, even when the
wound failed to heal, the operated side changed shape and became elongated
in the same way as the unoperated (control) side. It may be inferred, therefore,
that regression had occurred on both sides. Somites are also present on both
sides, which supports the thesis that the development of somites is correlated
with regression movements.
712
RUTH BELLAIRS
Experiment 3
Regression movements may again be deduced to have occurred since there
was a characteristic change in shape and elongation of the area pellucida. In
these embryos, therefore, somite formation appeared to be correlated with
regression movements.
Experiment 4
The presence of the notochord in the axes derived from the grafts is evidence
that regression has occurred. Thus, these embryos also show a correlation
between somite formation and regression movements.
The present results suggest that regression movements are necessary for and
therefore influence somite development. An influence of this sort must be
distinguished from an embryonic induction proper, which, typically, is brought
about by the direct action of one tissue on another. There is no evidence, however, as to the way in which the regression movements could exert an influence
on somite formation. One possible mechanism is that the changes in cell shape
which occur during the movements of the cells lead to important alterations in
the surface properties of the cells. These, in turn, may affect the internal
structure and metabolism of the cells.
SUMMARY
1. In two groups of experiments the so-called ' somite centres' were removed
from chick blastoderms. Nevertheless, well-developed somites formed. It is
concluded, therefore, that the 'somite centres' are not essential for somite
formation.
2. The same experiments, and others which are described, also provided
evidence that somites can form in the absence of neural tissue. It is concluded,
therefore, that neural tissue is not essential for somite formation.
3. Experiments involving extirpation of the node confirmed the now generally accepted findings of various authors that neither the node nor the notochord are essential for the differentiation of somites.
4. It is suggested that the regression movements may play a decisive role in
influencing somite differentiation.
RESUME
Le developpement des somites chez Vembryon de poulet
1. Dans deux groupes d'experiences, les pretendus 'centres somitiques' de
blastodermes de poulet ont ete excises. Neanmoins, des somites bien developpes
se sont formes. On conclut done que les 'centres somitiques' ne sont pas indispensables a la formation des somites.
DEVELOPMENT OF SOMITES IN THE CHICK EMBRYO
713
2. Les memes experiences, et d'autres que Ton decrit, ont aussi apporte la
preuve que des somites peuvent se former en l'absence de tissu nerveux. On
conclut done que le tissu nerveux n'est pas indispensable a la formation des
somites.
3. Des experiences comportant l'extirpation du noeud de Hensen ont confirme les observations de divers auteurs, maintenant generalement acceptees,
selon lesquelles ni le noeud de Hensen ni la notochorde ne sont indispensables
a la differentiation des somites.
4. On suggere l'idee que les mouvements de regression peuvent jouer un
role decisif en influencant la differenciation des somites.
ACKNOWLEDGEMENTS
I am most grateful to Mrs J. Astafiev for drawing the text-figures, and to Miss J. Spillman
for preparing the plate.
REFERENCES
M. (1950). The effects of antero-posterior reversals of lengths of the primitive streak in the chick. Phil. Trans. B. 234, 317-38.
ABERCROMBIE, M. & BELLAIRS, RUTH (1954). The effects in chick blastoderms of replacing
the node by a graft of posterior primitive streak. /. Embryol. exp. Morph. 2, 55-72.
ABERCROMBIE, M. & WADDINGTON, C. H. (1937). The behaviour of grafts of primitive streak
beneath the primitive streak of the chick. / . exp. Biol. 14, 319-34.
BELLAIRS, RUTH (1963). Differentiation of the yolk sac of the chick studied by electron
microscopy. / . Embryol. exp. Morph. 11, 195-219.
BELLAIRS, RUTH & NEW, D. A. T. (1962). Phagocytosis in the chick blastoderm. Exp.
Cell Res. 26, 275-9.
BUTROS, J. (1960). Induction by desoxyribonucleic acids of axial structures in post-nodal
fragments of chick blastoderms. / . exp. Zool. 143, 259-82.
BUTROS, J. (1962). Studies on the inductive action of the early chick axis on isolated postnodal fragments. J. exp. Zool. 149, 1-20.
FRASER, R. C. (1954). Studies on the hypoblast of the young chick embryo. / . exp. Zool.
126, 349^00.
FRASER, R. C. (1960). Somite genesis in the chick. III. The role of induction. / . exp. Zool.
145, 151-67.
GRABOWSKI, C. T. (1956). The effects of excision of Hensen's node on the early development
of the chick embryo. / . exp. Zool. 133, 301-44.
GRAPER, L. (1929). Die Primitiventwicklung des Hiihnchens nach stereokinemato-graphischen
Untersuchungen, kontrolliert durch vitale Farbmarkierung und verglichen mit der
Entwicklung anderer Wirbeltiere. Arch. EntwMech. Org. 116, 382-429.
GRUNWALD, P. (1936). Experimentelle Untersuchungen iiber die Beziehungen der Medullaranlage zur Entwicklung der Urwirbel beim Huhn. Arch. EntwMech. Org. 135, 389—
407.
JACOBSON, W. (1938). The early development of the avian embryo. II. Mesoderm formation
and the distribution of presumptive embryonic material. /. Morph. 62, 445-501.
NEW, D. A. T. (1955). A new technique for the cultivation of the chick embryo in vitro.
J. Embryol. exp. Morph. 3, 326-31.
NEW, D. A. T. (1959). The adhesive properties and expansion of the chick blastoderm.
/. Embryol. exp. Morph. 7, 146-64.
PASTEELS, J. (1937). £tudes sur la gastrulation des Vertebres meroblastiques. III. Oiseaux.
Arch. Biol., Paris et Liege, 48, 381-463.
ABERCROMBIE,
714
RUTH BELLAIRS
PEEBLES, F. (1898). Some experiments on the primitive streak of the chick. Arch. EntwMech.
Org. 7, 405-29.
SHOGER, R. L. (1960). The regulative capacity of the node region. J. exp. Zool. 143, 221-38.
SPRATT, N. T. (1947). Regression and shortening of the primitive streak in the explanted
chick blastoderm. / . exp. Zool. 104, 69-100.
SPRATT, N. T. (1955). Analysis of the organizer center in the early chick embryo. I. Localization of prospective notochord and somite cells. J. exp. Zool. 128, 121-64.
SPRATT, N. T. (1957a). Analysis of the organizer center in the early chick embryo. II.
Studies on the mechanics of notochord elongation and somite formation. / . exp. Zool.
134, 577-612.
SPRATT, N. T. (1951b). Analysis of the organizer center in the early chick embryo. III. Regulative properties of the chorda and somite centers. / . exp. Zool. 135, 319-54.
VAKAET, L. (1962). Pregastrulatie en gastrulatie der Vogelkiem. Bruxelles: Presses Academiques Europeennes.
WADDINGTON, C. H. (1932). Experiments on the development of chick and duck embryos,
cultivated in vitro. Phil. Trans. B, 221, 179-230.
WADDINGTON, C. H. (1935). The development of isolated parts of the chick blastoderm.
J.exp. Zool. 71, 273-91.
WADDINGTON, C. H. (1952). The Epigenetics of Birds. Cambridge University Press.
WATERMAN, A. J. (1936). Experiments on young embryos cultured in vitro. Proc. nat. Acad.
Set, Wash. 22, 1-3.
WETZEL, R. (1929). Untersuchungen am Huhnchen. Die Entwicklung des Keims wahrend
der ersten beiden Bruttage. Arch. EntwMech. Org. 119, 188-321.
WOLFF, E. (1936). Les bases de la teratogenese experimentale des Vertebres amniotes, d'apres
les resultats de methodes directes. Arch. Anat., Strasbourg 22, 1-375.
(Manuscript received 28th May 1963)