Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Plant breeding wikipedia , lookup
Plant evolutionary developmental biology wikipedia , lookup
Plant ecology wikipedia , lookup
Plant secondary metabolism wikipedia , lookup
Plant reproduction wikipedia , lookup
Plant physiology wikipedia , lookup
Perovskia atriplicifolia wikipedia , lookup
Plant morphology wikipedia , lookup
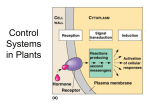
Chapter 35 COMMUNICATION: CHEMICAL SIGNALS The tissues that sense environmental change are not necessarily those that respond to the change. Plant hormones are chemical messengers. Produced in one part of the plant. Transported to another part of the plant. Causes a physiological response: regulate growth and development. Each hormone type causes several responses. The responses of different hormones overlap. There are five classes of plant hormones. PHOTOTROPISM The Darwins published their hypothesis about chemical signals and phototropism in 1881. Peter Boysen-Jensen conclude in 1913, after conducting experiments, that the signal was indeed a chemical and that it could diffuse from one part of the plant to another. In 1925, Frits Went conducted his classical experiment with oat coleoptiles. Went was able to collect the phototropic chemical in blocks of agar. Went was able to produce a phototropic-like response without the stimulus of light. Cholodny and Went proposed independently that the response is caused by an asymmetrical distribution of the hormone. Went named the hormone auxin from the Greek auxein, to increase. The Cholodny-Went Hypothesis. Auxin is produced in the tips of the coleoptiles. The auxin is then transported from one side to another of the coleoptile in response to light. Cells on the side with the greater concentration of auxin will elongate more causing the entire stem to bend towards the light. Other scientists have proposed that the auxin is destroyed in the side where the light strikes causing a difference in auxin concentration along the stem. Kögl and Thimann independently isolated the auxin hormone. It turned out to be indole acetic acid or IAA. The concentration of IAA is about 50 nanograms for every 50 grams of fresh tissue. 1 ng = 1 billionth of a gram or 0.000 000 000 1 g When IAA arrives at a target cell, then its message must be received and transduced to produce the appropriate response. Researchers found that IAA binds to a receptor protein called ABP1. The Acid-Growth Hypothesis It proposes that... 1. IAA produces or activates additional proton pumps. 2. The pumping of protons into the extracellular matrix causes K+ and other positive ions to enter the cell. 3. This increase in solutes brings an influx of water into the cell. 4. There is then an increase in turgor pressure that makes cell expansion possible. Hager and colleagues found that cells treated with addition IAA increased the number of proton pumps by 80% relative to untreated control cells. They also found that the acidity of the of the cell wall changed from a pH of 5.5 to one of 4.5. The cell wall is rigid. So how does the cell wall expands? Cosgrove found two classes of cell wall proteins that actively increase cell length when the pH in the cell wall drops below 4.5. These proteins are called expansins. Expansins have been found in many species and tissues but how they work is not known yet. One hypothesis proposes that these protein break the bonds between cellulose fibers and pectin fibers or other wall components, allowing for stretching and expansion of the wall. APICAL DOMINANCE In apical dominance, the majority of the stem growth takes place in the apical meristem of the shoot, and inhibits the growth of other meristems (e.g. lateral buds) located down the stem of the plant. Apical dominance occurs because auxin flows from the apex of the shoot down to the tissues below. Auxin transport is polar, unidirectional. Radioactively labeled auxin molecules have shown that the hormones travel all the way down to the central portion of the root, and when it reaches the root tip, the hormone moves out to the epidermal cells and up for a short distance - the fountain model of flow. The Cholodny-Went Hypothesis maintains that asymmetrical distribution of auxin causes gravitropism in roots. In a horizontal root tip, gravity-sensing cells redistribute the auxin; more auxin goes to the lower side of the root tip. Asymmetrical auxin distribution causes the cell in the upper side to elongate causing the root tip to bend downward - gravitropism. The Chemiosmotic Model. This model attempts to explain how polar transport takes place. The auxin in an acidic cell wall (pH 5.5) accepts a proton, H+, and becomes neutral. There are influx carrier proteins located only on the upper side of the cell membrane. Auxin is taken into the cell via this influx protein carrier that takes the auxin in with the attached proton. Inside the cell the pH is neutral (pH = 7) and the auxin loses the proton and becomes negative, anionic. There are carrier proteins specific for negative auxin called efflux carrier proteins, located only on the cell membrane at the base of the cell. Auxin leaves the cell through these carrier protein following an electrochemical gradient. These events repeat and the auxin is transported from top of the cell to the bottom of the cell and out to be pick up again by influx carrier of the cell below. Gälweiler and colleagues identified a protein coded by the gene PIN1 that is located only at the base of stem cells. It is hypothesized that these are the efflux carriers. An overview of auxin action. It is produce in the apical meristem of shoots, in young leaves and in seeds. It is transported downward in parenchyma cells. It causes cell elongation, promotes xylem and phloem differentiation, inhibits lateral bud development, stimulates fruit development but delays ripening, and inhibits leaf abscission. Auxin is the root-growth hormone sold in nurseries. It promotes root growth on cut-off shoots. Because of its many effects on plants, some biologists have proposed that auxin overall function is to signal where cells are in space, where the cell is located along the axis of the plant. It helps to determine the overall shape of the plant due to changes in light availability, wind strength, etc. Auxin concentration signals how tissues should respond. GROWTH AND DORMANCY Abscisic acid (ABA) signals when the plant should stop growing. Gibberillic acids signal when a plant should start growing again. There are more than 100 gibberillic acids identified but only a few have been shown to act as hormones. Seed dormancy and germination Many plants produce seeds that must suffer a period of cold or hot temperatures, or drought before a favorable period for growth arrives. Abscisic acid is the signal that inhibits seed germination. Gibberillic acid signals the onset of embryonic development and germination. The protein α-amylase is a digestive enzyme that breaks the bonds between the sugar subunits in starch. α-Amylase is release from the aleurone layer of the seed during germination. Gibberillic acid (GA) activates the production of α-amylase. The embryo absorbs water and triggers the diffusion of GA. GA produced by the embryo reaches the membranes of cells in the aleurone layer. Receptors in the membrane receive this signal. The receptors activate the production of Myb, a transcription activator. Myb travels to the nucleus, binds to the α-amylase promoter, and triggers transcription and α-amylase production. The Myb protein that binds to the promoter has been named GAMyb. Abscisic acid decreases the production of α-amylase. Preliminary data suggests that ABA induces the production of Myb proteins that act as gene repressors. Myb proteins bind to the α-amylase promoter and shut down amylase production. ABA activates transcription repressors. Both activators and repressors compete for the same site in the promoter gene. If ABA is in higher concentration, repression dominates and dormancy occurs. If GA is in higher concentration, activators dominate and germination proceeds. Therefore... 1. A cell's response to a hormone often occurs because specific genes are turned on or off. 2. Hormones rarely act on DNA directly. 3. Different hormones interact at the molecular level because they induce different transcription activators and repressors. The relative amount of these regulatory factors determines which activators or repressors dominate the response. Closing guard cells Guard cells open in response to light in order to allow carbon dioxide to enter the mesophyll spaces for photosynthesis. In cases of drought, the roots cannot obtain enough water and the stomata close. Experiments have shown that plants under water stress have a higher concentration of ABA in their roots and leaves. Shoot elongation Japanese scientist isolated a substance in the 1930s that causes rice seedlings to elongate abnormally and fall over before harvest. These rice plants were infected with the fungus Gibberella fujikuroi. Treating seedling with extract of the fungus caused abnormally long plants. Analyzing stem-length mutants One of the traits studied by Gregor Mendel was dwarfism in peas. Mendel found that one allele coded for normal stem length, tall, and another for dwarf stems. Dwarf pea plants attain normal height when treated with GA1. This suggests that plants can react normally but cannot manufacture gibberellin. Researches found that a locus called Le (for length) was responsible for the synthesis of GA1. Diane Lester found a locus that encodes for the enzyme 3β-hydroxylase. 3β-hydroxylase catalyzes the addition of -OH to a gibberellin called GA20 and converts it to GA1. Lester found that in mutant DNA, the Le gene coded for the AA threonine instead of alanine. This produces an abnormal enzyme that cannot add -OH to GA20 to form the active gibberellin GA1. Role of gibberellins in plant growth It is produced in young leaves, roots, shoot apical meristem and in the seed embryo. Method of transport in the plant is unknown. It promotes seed germination, cell division and elongation, fruit development, flowering in some plants and breaks seed dormancy and winter dormancy. OTHER HORMONES 1. CYTOKININS They are produced in the roots. Travel upward in the xylem. Promote cell division and differentiation in which unspecialized cells become specialized, promotes chloroplast development, stimulates lateral bud development, inhibits abscission and delays senescence. There is evidence that cytokinins activate genes that keep the cell cycle going. Zeatin was the first isolated naturally occurring cytokinin. 2. ETHYLENE It is a gaseous hormone produced in stem nodes, aging tissues and ripening fruits. It probably diffuses out of the tissue that produces it. It promotes ripening of the fruits, senescence and abscission, inhibits cell elongation, stimulates germination of seeds and it is involved in responses to wounds and infections by microorganisms. OTHER CHEMICALS INVOLVED IN PLANT GROWTH AND DEVELOPMENT. Polyamines are organic molecules with two or more amine groups (- NH2). They may be involved in gene expression and increase the transcription of DNA and translation of mRNA. They exist in high concentration in plant tissues and are not transported extensively through the plant. Systemin, a plant polypeptide, stimulates plant defenses that produce chemicals that disrupt insect digestion. Present in very small quantities. Oligosaccharins are cell-wall fragments consisting of short, branched chains of sugar residues. Present in quantities lower than hormones. Bind to membrane receptors and affect gene expression and have many effects on plants. Some have antibiotic properties and kill fungi and other plant pathogens. Others promote vegetative growth and inhibit flowering. Salicylic acid help to defend the plant against insect attack and promotes wound healing. The function of brassinosteroid hormones in plants is not clear. One steroid hormone seems to be involved in light regulated developmental steps, others in growth. Plants without these hormones are dwarf.