Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Biomineralisation of
Magnetosomes in Bacteria
Microbial Bionanotechnology
Chapter 5
Magnetotactic
• Magnetosome
– Crystalline particles of iron oxide or sulfide
– Magnetite Fe3O4
– Greigite Fe3S4
• All are either obligate microaerophiles or strict
anaerobes
• Motile, aquatic bacteria
• Direction of motility is affected by the Earth’s
geomagnetic field
• Strains are either north- or south-seeking depending
upon oxic conditions
– North-seekers predominate in the northern hemisphere
– South-seekers predominate the southern hemisphere
– Exist in equal numbers at the equator
• Current hypothesis states that these bacteria use
the geomagnetic field to locate lower O2 or
anaerobic habitats
2
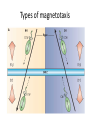
Types of magnetotaxis
Types of magneto-aerotaxis
There are Two Types of Magnetoaerotaxis...
Axial Magnetoaerotaxis
e.g., Magnetospirillum
magnetotacticum
Polar Magnetoaerotaxis
e.g., strain MC-1, a magnetotactic coccus
5
Magnetite Crystals
Produced by
Magnetotactic
Bacteria
Pc2
Sw3
Pm3
Pm2
Sw7
dnaA
st p2
Sw6
bf r1, bf r2,
nm1, st p1
magA,
st p5
Sw1
~4. 3 mb
st p3, st p4
dms,
f dx
Pm4
ni f l 1, rpl 6, Sw2
rpoA
f t sH,
ni f L2
Sw5
napA, rrn2,
sodB
Sw4
Pc1
Pm1
The MS-1 Genome
From http://www.soton.ac.uk/~serg/biotech/mtb-main.htm
Bertani, Weko, Phillips, Gray & Kirschvink,
Physical and genetic characterization of the
Genome of Magnetosprillum Magnetotacticum,
Strain MS-1, Gene 264: 257-263, 2001
Global
Swimming
Preferences of
Magnetotactic
The equator has
Bacteria
North-Seeking in the
Northern Hemisphere
(Blakemore, 1975)
N
Pelau
Equator
small numbers
of both N- and
S-seekers
These bugs use the
(Frankel et al.,
1981; Chang & geomagnetic field as
Kirschvink, 1989) an up/down indicator
at redox gradients.
S
South-Seeking in the
Southern Hemisphere
(Kirschvink, 1980)
Other bugs use tumblerun random walks.
The Kalmijn-Blakemore (‘78) Pulseremagnetization Experiment
Start with Northseeking bacteria
In a weak biasing
Field (~50 uT)
Hit with a sharp,
antiparallel pulse
exceeding Hc
North
South
100 mT magnetic pulse!
+
=
The remagnetized bacteria roll over and start swimming to the South …
North
South
Typical Bacterial Magnetosomes
(Courtesy of H. Vali)
3D Imaging of Bacterial Magnetosomes
Buseck et al., PNAS 98, 13490-13495, 2001
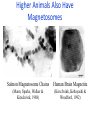
Higher Animals Also Have
Magnetosomes
Salmon Magnetosome Chains
Human Brain Magnetite
(Mann, Sparks, Walker &
Kirschvink, 1988)
(Kirschvink, Kobayashi &
Woodford, 1992)
MV-1 and Magnetofossils
Biologically Important Features of
Magnetosomes:
(Darwinian Selection for Magnetic Properties!!!)
•
•
•
•
•
•
•
Size & Shapes with the Single-Domain field
Elongation of the ultrafine crystals
Orienting [111] axis along length
Truncation of ends.
Exclusion of Trace Metal Impurities
Perfect Crystal Lattice
Alignment in chains of similarly-sized particles
Truncated
Xtl Ends
Particle
Elongation
[111]
Alignment
Seven Criteria
for Identifying
Magnetofossils*
*All 7 of these act to maximize the magnetic
moment at the cellular level; however, not all
magnetofossils will plot in the center
Magnetosomes show the effect of Natural Selection for
their magnetic properties. They are just the right size to be
perfect biological bar magnets. Any bacterium that makes
crystals outside the SD field will die!
5
Two- Domai n
I nt er act i on
st abi l i zed
1 Bact er i a
10
Pr ot i st s
I sol at ed
cr yst al s
Si ngl eDomai n
0. 1
4
10
Pi geons Bact er i a
3
MS- 1
Fi sh, Human
Super par amagnet i c
. 001
0. 0
0. 2
0. 4
0. 6
0. 8
10
1. 0
Wi dt h/ Lengt h
2
Lengt h ( A)
10
o
Lengt h ( um)
10
Magnetosome Elongation by the
Magnetotactic Bacteria
Octahedral to Centrosymmetric Hexagonal Prisms
(vibroid and coccoid
cells)
_
_
(111)
_
(111)
(111)
_
(111)
__
(111)
_
(111)
(111)
_
(111)
__
_
(111)
(011)
(111)
_
(111)
_
__
(111)
(111)
(111)
_
(111)
_
(011)
_
__
(111)
Cubo-Octahedral to Elongated Cubo-Octahedral
BIOGENIC MAGNETITE GEOMETRIES
Blue = {100}
Red = {110}
Green = {111}
{111} {111} @ 109°
{111} {100} @ 125°
{111} {110} @ 145°
{100} {110} @ 135°
Hexaoctahedron
KT-K
Truncated
Hexaoctahedron
Why should the bacteria truncate the
ends of their magnetosomes?
Because sharp edges produce magnetic ‘Flower Structures’, which
warp the internal magnetization directions. Putting those iron atoms
in the next crystal increases the magnetic moment of the cells!
See Fabian et al., GJI 124:89-104, 1996; Newell & Merrill JGR 105: 19377-19391)
A PDF file for this model is available on the
Magnetofossil home page At Caltech
www.gps.caltech.edu/users/jkirschvink/
magnetofossil.html
111
100
110
Martian Magnetofossil
(enlarged ~1,600,000 times)
Modeled after K. Thomas-Keprta et al. PNAS 98: 2165, 200
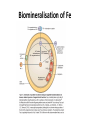
Iron Uptake & Purification in the
Magnetotactic Bacteria
Source
Rock
Ma g n e t o s o me Me mb r a n
es
3+
2+
I ron (Fe and Fe
)
Ot her Cat i ons
Trans-membrane
i ron channel (mag-A?)
Si derophores
Transf erri n
Ce l l
me mb r a n e
Biomineralisation of Fe
Magnetotactic Bacteria
•
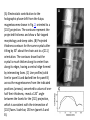
Electron cryotomography of Magnetospirillum magneticum sp. AMB-1 reveals
that magnetosomes are invaginations of the inner membrane. (A) General
features of AMB-1 cells highlighted in a 12-nm-thick section of an ECT
reconstruction. Outer membrane, OM; inner membrane, IM; peptidoglycan
layer, PG; ribosomes, R; outer membrane bleb, B; chemoreceptor bundle, CR;
poly-ß-hydroxybutyrate granule, PHB; gold fiduciary marker, G; magnetosome
chain, MG. Scale bar, 500 nm. (B to E) Representative magnetosomes
containing no magnetite (B), small (C), mediumsized (D), and fully-grown (E)
crystals are invaginations of the inner membrane. Scale bar, 50 nm.
•
Magnetosome chains are flanked by long cytoskeletal filaments. (A) Larger view of the
magnetosome chain in Fig. 1A. (B) Similar view of a magnetosome chain grown in the
absence of iron, which prevents the formation of magnetite crystals. Arrows point to the long
filaments. (C) Three-dimensional organization of magnetosomes (yellow) and their associated
filaments (green) shown in (B) with respect to the whole cell (blue). Scale bars, 100 nm.
Bakterielle Magnetosomer
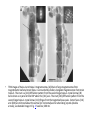
TEM image of two Itaipu-1 cocci. Each
bacterium has two chains of magnetosomes
(arrows) and two phosphorus-rich globules (P).
Scale bar, 1 μm.
High-resolution TEM images of Itaipu-1 magnetosomes with indexed bars parallel to
lattice planes. Obvious symmetries between even the very small facets on opposite sides
of the crystal diagonals can be seen. Comparison with other crystals of the chain in Fig. 2
also indicates that this symmetry regularly alternates between crystals.
(A) Electrostatic contribution to the
holographic phase shift from the Itaipu
magnetosomes shown in Fig. 2, oriented to a
[110] projection. The contours represent the
projected thickness and show a flat-topped
morphology and steep sides. (B) Projected
thickness contours for the same crystals after
tilting by 30° about the chain axis to a [211]
orientation. The contours show that the
crystal is much thicker along its center than
along its edges, having a central ridge formed
by intersecting faces. (C) Line profiles (solid
line for panel A and dashed line for panel B)
across the magnetosomes from the indicated
positions (arrows), converted to values of onehalf their thickness, reveal a 120° angle
between the facets for the [211] projection,
which is consistent with the intersection of
[110] faces. Scale bar, 150 nm (panels A and
B).
•
TEM images of Itaipu-1 and Itaipu-3 magnetosomes. (A) Chain of large magnetosomes from
magnetotactic bacterial strain Itaipu-1 surrounded by smaller, elongated magnetosomes from strain
Itaipu-3. The inset is a [211] diffraction pattern from the second large Itaipu-1 crystal (arrow). (B)
Same chain as in panel A tilted 30o about the [111] axis. The inset [110] diffraction pattern from the
second large Itaipu-1 crystal shows (111) fringes from the magnetically easy axis. Corner faces {111}
and {200} are mirrored about the vertical (or horizontal) axis for alternating crystals (double
arrows); see detailed image in Fig. 3. Scale bar, 200 nm.
Tomographic reconstruction of a magnetite nanocrystal from an undescribed coccus
collected from Sweet Springs Nature Reserve, Morro Bay, CA, reconstructed from a
tilt series of STEM HAADF images obtained at 300 kV on a Philips CM300 FEG TEM
over a range of ± 56°. The tableau shows the three-dimensional morphology of the
crystal viewed from a range of directions.
Magnetosome Element Analysis
Cu-Fe Analysis
Magnetosomes
Magnetosome crystal morphology
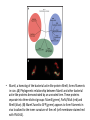
• MamK, a homolog of the bacterial actin-like protein MreB, forms filaments
in vivo. (A) Phylogenetic relationship between MamK and other bacterial
actin-like proteins demonstrated by an unrooted tree. These proteins
separate into three distinct groups: MamK (green), ParM/StbA (red) and
MreB (blue). (B) MamK fused to GFP (green) appears to form filaments in
vivo localized to the inner curvature of the cell (cell membrane stained red
with FM4-64).
• MamK is required for the proper organization of the magnetosome chain.
(A) Three-dimensional reconstruction of a wild-type AMB-1 cell. The cell
membrane (gray), magnetosome membrane (yellow), magnetite (orange),
and magnetosome-associated filaments (green) are rendered. (B) mamK
mutant, where magnetosomes appear disordered and no filaments are
found in their vicinity. (C) mamK cell expressing mamK-GFP on a plasmid
showing full reversal of the mutant phenotype.
Magnetosome membrane proteins
Comparative genome analysis
Magnetotaxis genes
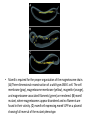
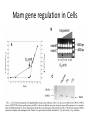
Mam gene regulation in Cells
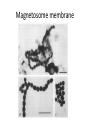
Magnetosome membrane
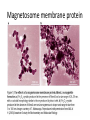
Magnetosome membrane protein
Biotechnological applications
•
•
•
•
•
•
Delivery systems
Separation systems
DNA arrays
RNA arrays
Thermo treatment
Sensor systems
Applications of Magnetosome particles