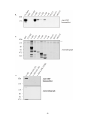
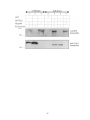
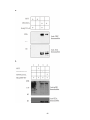
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
G protein–coupled receptor wikipedia , lookup
Cell nucleus wikipedia , lookup
Protein phosphorylation wikipedia , lookup
Cellular differentiation wikipedia , lookup
Magnesium transporter wikipedia , lookup
Protein moonlighting wikipedia , lookup
Protein domain wikipedia , lookup
Signal transduction wikipedia , lookup
Intrinsically disordered proteins wikipedia , lookup
FUNCTIONAL CONSEQUENCES OF THE DIRECT PHYSICAL INTERACTION BETWEEN E2A TRANSCRIPTION FACTORS AND CBP/p300 by Brandy Dawn Hyndman A thesis submitted to the Department of Pathology and Molecular Medicine In conformity with the requirements for the degree of Doctor of Philosophy Queen’s University Kingston, Ontario, Canada September, 2007 Copyright © Brandy Dawn Hyndman, 2007 Abstract The E2A locus is involved in chromosomal translocations associated with acute lymphoblastic leukemia. The most common of these involves a translocation between chromosomes 1 and 19 (t1;19), resulting in expression of the chimeric oncoprotein E2APBX1. A direct interaction between transcriptional activation domain 1 (AD1) of E2A and KIX domain of the histone acetyltransferase (HAT) /co-activator CBP is required for E2A-PBX1-mediated leukemia induction in mice. This thesis examines the functional consequences of the direct, physical interaction between E2A and CBP, for both proteins. We demonstrate that the interaction between E2A and CBP/p300, as well as another HAT/co-activator, p/CAF, results in acetylation of E2A. Mutagenesis-based mapping studies identify several lysine residues as substrates for acetylation. Of particular interest, a conserved lysine (K34) located within AD1 is acetylated in vitro and in vivo. Substitution of this residue to arginine impairs transcriptional activation of a luciferase reporter while substitution to glutamine, mimicking the acetylation, restores E2Amediated transcriptional activation. Recent studies have shown that several transcription factors can modulate the intrinsic HAT activity of CBP/p300. We were surprised to find that E2A proteins enhance acetylation of histones by CBP, in vitro and in vivo, in a KIX domain-independent manner. Acetylation of E2A is also not required for stimulation of CBP/p300 histone acetylation. It appears that E2A interacts with the other CBP domains to mediate this effect, presumably through allosteric effects. ii In summary, we demonstrate that acetylation of E2A plays a role in mediating the transcriptional activation activity of E2A. Furthermore, acetylation of E2A enhances its interaction with CBP/p300, at least in the presence of additional nuclear factors. We show evidence that p/CAF may mediate this effect. Enhancement of CBP/p300 HAT activity by oncogenic E2A-PBX1 proteins in vivo, suggests that some of its leukemiapromoting effects may be due to E2A-induced gain of function effects on CBP/p300. The enhanced interaction between acetylated E2A and CBP/p300, as well as the E2Amediated stimulation of histone acetyltransferase activity might play a role in the DNAbinding-independent induction of proliferation. iii Co-Authorship Mr. Richard Bayly generated the results shown in Figure 12. Mr. Bayly was also responsible for preparing some reagents. I performed all other experiments iv Acknowledgements I have met many great people during my time a Queen’s University. First and foremost, I would like to thank my thesis supervisor, Dr. David LeBrun, for allowing me to be a part of his lab. He is very enthusiastic about research and I have learned a great deal from him. David, I greatly appreciate your mentorship during the course of my PhD. Since there is no ‘I’ in research, I would also like to acknowledge our excellent research assistant, Mr. Richard Bayly. Dick always has a smile on his face and time to teach those around him. I couldn’t have done it without your guidance. I’ll miss hearing about the horses and the annual pond hockey games. Thanks also go out to all the past and present members of the LeBrun lab, including Mr. Richard Casselman and Cheryl Foster. I am also grateful to Ms. Barbara Saunders, the Graduate Secretary for the Department of Pathology and Molecular Medicine, for letting me bend her ear over the years. (Yes Barb, I checked the Graduate Handbook….) Finally, and most importantly, I would like to thank my family for their unconditional love and support. To Mom and Dad, I couldn’t have done it without your support and encouragement. I love you both. Especially, thanks for the airmiles and plane tickets home for the holidays over the past 8 years. Did it really take that long?! Kingston is great city but Alberta will always be home. My brothers, Cody and Kyle, started their PhD’s a year after me but gained the title of ‘Dr. Hyndman’ two years before me. I guess v that’s the difference between Health Science research and Mathematics/Finance and Economics! Thank you for your encouragement to get finished and for keeping life interesting. Cody, maybe I’ll have time now to learn how to knit that sweater you’ve been bugging me for over the past 10 years! vi Table of Contents Abstract ............................................................................................................................... ii Co-Authorship ................................................................................................................... iv Acknowledgements ............................................................................................................. v Table of Contents .............................................................................................................. vii List of Figures ..................................................................................................................... x Chapter 1 Introduction .................................................................................................... 1 Chapter 2 Literature Review ........................................................................................... 4 2.1 Structure and Function of the E2A Gene Products .................................................... 4 2.1.1 E2A Gene Products are Basic Helix-Loop-Helix Proteins ................................. 4 2.1.2 E2A Contains Two Transcriptional Activation Domains................................... 5 2.2 E2A Proteins Are Required For B-Cell Commitment And Differentiation ............ 10 2.2.1 E2A is Specifically Required for the Earliest Stages of B-Cell Differentiation ................................................................................................................................... 10 2.2.2 E2A is Involved in Later Stages of B-Cell Development ................................ 11 2.3 Regulation of E2A Activity..................................................................................... 11 2.3.1 Regulation of E2A by Dimerization with Other bHLH Proteins .................... 11 2.3.2 E2A Proteins are Substrates for Phosphorylation............................................. 12 2.3.3 E2A Stability is Regulated by Ubiquitination .................................................. 13 2.4 E2A Involvement In Translocations And Leukemia ............................................... 15 2.4.1 The E2A Gene is Located on Chromosome 19................................................. 15 2.4.2 PBX1 Structure and Function ........................................................................... 16 2.4.3 E2A-PBX1 Structure and Function .................................................................. 16 2.5 Chromatin and Transcriptional Regulation ............................................................. 18 2.5.1 Chromatin Structure ......................................................................................... 18 2.5.2 Enzymes That Covalently Modify Histones ..................................................... 19 2.5.3 Chromatin Remodeling Enzymes ..................................................................... 22 2.6 CBP and p300 are Transcriptional Co-Activator Proteins ...................................... 24 2.6.1 CBP and p300 Have Intrinsic Acetyltransferase Activity ................................ 27 2.7 Acetylation and Transcription Factor Regulation ................................................... 28 2.8 Recognition Motifs for Acetyltransferase Proteins ................................................. 29 vii 2.9 CBP Interacts Directly with E2A Proteins .............................................................. 30 2.9.1 Identification of E2A-CBP Interactive Domains.............................................. 30 2.9.2 Functional Significance of the E2A-CBP Interaction for E2A-PBX1-mediated Leukemogenesis ............................................................................................... 32 Chapter 3 Hypothesis and Research Objectives .......................................................... 34 Chapter 4 Materials and Methods ................................................................................ 36 Chapter 5 Acetylation of Lysine 34 in E2A Proteins Contributes to Transcriptional Activation ....................................................................................................... 47 5.1 Contributions ........................................................................................................... 47 5.2 Rationale.................................................................................................................. 47 5.3 Results ..................................................................................................................... 48 5.3.1 The Portion of E2A Included in E2A-PBX1 Interacts with Acetyl-Transferase Proteins Capable of Acetylating Both Histones and E2A ................................ 48 5.3.2 Acetylation of E2A Correlates with Recruitment of CBP ................................ 52 5.3.3 In Vitro Acetylation of E2A ............................................................................. 57 5.3.4 Both p300 and p/CAF Interact Directly with E2A ........................................... 61 5.3.5 Acetylation of E2A Promotes its Interaction with CBP ................................... 64 5.3.6 Mapping of Acetylated Lysines ........................................................................ 67 5.3.7 Transcriptional activation by AD1 Involves Acetylation of K34..................... 78 5.3.8 A Role for Acetylation of Lyisne 34 in the Transcriptional Fynction of Activation Domain ........................................................................................... 82 5.4 Discussion ............................................................................................................... 87 Chapter 6 E2A Enhances the Acetyl-transferase Activity of CBP ............................. 94 6.1 Contributions ........................................................................................................... 94 6.2 Rationale.................................................................................................................. 94 6.3 Results ..................................................................................................................... 95 6.3.1 E2A Proteins Enhance in vitro Acetylation of Histones by CBP/p300 ............ 95 6.3.2 Mapping of the E2A Domains Required to Increase HAT Activity of CBP . 103 6.3.3 The KIX and Bromo Domains are not Required for E2A-induced Increase of CBP HAT Activity ......................................................................................... 107 6.3.4 E2A Does Not Enhance CBP Acetylation of Nucleosomes in vitro .............. 113 viii 6.3.5 E2A Enhances CBP Catalytic Activity in vivo............................................... 116 6.4 Discussion ............................................................................................................. 121 Chapter 7 General Discussion and Future Directions ............................................... 124 References ..................................................................................................................... 130 ix List of Figures Figure 1: Domain structures of E2A, PBX1 and E2A-PBX1 fusion proteins and location of regulatory elements within E2A ..................................................................... 6 Figure 2: Histone modifications and Lysine acetylation ................................................ 20 Figure 3: Domain structure of the co-activator CREB-binding protein .......................... 25 Figure 4: E2A is acetylated by acetyltransferase activity recruitment from nuclear extracts ............................................................................................................ 50 Figure 5: Acetylation of E2A correlates with recruitment of CBP.................................. 54 Figure 6: Acetylation of E2A by purified CBP/p300 or p/CAF ...................................... 58 Figure 7: p/CAF interacts directly with E2A. .................................................................. 62 Figure 8: Acetylation of E2A promotes its interaction with CBP/p300. ......................... 65 Figure 9: Schematic indicating the location and phylogenetic conservation of lysine residues in the E2A-PBX1a oncogenic protein ............................................... 68 Figure 10: The catalytic domain of recombinant p/CAF acetylates lysines 34, 101 and 144 of E2A. .................................................................................................... 71 Figure 11: Recombinant p300 and the co-activator complex recruited from nuclear extracts acetylate identical lysine residues within E2A amino acids 161-483. ........................................................................................................ 75 Figure 12: Post-translational modification of lysine 34 is involved in transcriptional activation by E2A. ......................................................................................... 80 Figure 13: The oncogenic portion of E2A is not a substrate for ubiquitination or sumoylation. ................................................................................................... 84 Figure 14: E2A enhances the histone acetyltransferase activity of p300. ....................... 98 Figure 15: Time course experiment showing that E2A enhances the catalytic activity of CBP approximately two-fold ....................................................................... 101 Figure 16: The α-helical domain within AD1 and AD2 of E2A are dispensable for enhancement of CBP histone acetylation. ................................................... 105 Figure 17: E2A interacts with the HAT domain of CBP to enhance intrinsic HAT activity of CBP. The KIX and BROMO domains of CBP are dispensable for enhancement of HAT activity by E2A......................................................... 110 x Figure 18: E2A is unable to enhance CBP-mediated acetylation of nucleosomes in vitro ......................................................................................................................................... 114 Figure 19: The E2A portion of E2A-PBX1 mediates enhanced acetyltransferasedependent transcriptional activation by CBP in vivo. ................................... 118 xi Chapter 1 Introduction Leukemia remains the most common childhood cancer, accounting for approximately twenty-five percent of all pediatric cancer cases in Canada (1). Acute lymphoblastic leukemia (ALL), characterized by rapid accumulation of abnormal, often immature B- or T-lineage cells in the bone marrow and blood, accounts for seventy-five percent of childhood leukemias. While the current five year survival rate for pediatric ALL is approximately eighty-percent, due to improved treatment protocols developed over the last thirty years, ALL still represents the leading cause of cancer fatalities in persons under the age of nineteen years (1). Clues to the molecular mechanisms of leukemogenesis have been provided by the discovery of non-random chromosomal abnormalities in the leukemic cells. More than half of all leukemias have detectable chromosomal abnormalities by karyotypic analysis (2). Some of these abnormalities are consistently associated with various sub-classes of ALL; such non-random chromosomal translocations are seen in approximately forty-five percent of ALL cases (2). Many of these translocations lead to juxtaposition of the coding sequence of two genes, normally located on separate chromosomes, leading to expression of chimeric oncoproteins. Translocations may also alter the regulation of a gene by placing it under the control of new promoter elements. One chromosomal translocation involving chromosomes 1 and 19, t(1:19), is found in the leukemic cells of approximately five percent of ALL cases and is the second most 1 common translocation in ALL. First reported in 1984, t(1;19) was determined to create a fusion gene involving two genes, PBX1 on chromosome 1 (3,4) and E2A on chromosome 19 (5). This translocation results in expression of a chimeric protein called E2A-PBX1 and is found in approximately 30% of patients diagnosed with precursor Bcell ALL (pre-B ALL). This translocation was initially associated with a poorer prognosis. However, more intensive treatment regimens have since erased this difference (6). Interestingly, the E2A locus is also involved in at least two other leukemia-associated translocations. A translocation between chromosomes 17 and 19 creates the fusion gene E2A-HLF (7,8) while an internal rearrangement of chromosome 19 fuses the same region of the E2A gene to the FB1 gene (9), resulting in expression of truncated forms of E2A. E2A gene products normally function to regulate lymphocyte development by regulating the transcription of B-lineage specific genes. In addition, E2A proteins can interact with tissue specific factors to regulate tissue differentiation in other non-lymphoid cell types. In recent years abundant evidence has suggested a functional interaction between E2A and several transcriptional co-activator proteins in mediating E2A-target gene expression (10-12). Work in Dr. LeBrun’s lab has uncovered a direct interaction between E2A and the transcriptional co-activator CREB binding protein (CBP) (13). This interaction is required for the induction of tumours in mice by E2A-PBX1 and the outgrowth of immortal cells from primary bone marrow cultures (13,14). Transcriptional co-activators, such as CBP and the highly homologous protein p300, are involved in transcriptional regulation by two general mechanisms. First, by affecting 2 chromatin conformation by acetylation of lysine residues within nucleosomal histone tails and, second, by mediating recruitment of the initiation complex to promoters through interaction with DNA binding transcription factors. Over the past decade several investigators have determined that CBP/p300 can also mediate acetylation of non-histone proteins such as transcription factors (15). This acetylation has been shown to affect several aspects of transcription factor function, including DNA binding affinity, proteinprotein interactions, sub-cellular localization and protein stability. Reciprocally, interaction of several viral and cellular proteins with histone acetyltransferase (HAT) coactivators, including CBP/p300, alters the ability of the co-activator to acetylate histone proteins. The work described in this thesis investigates the functional consequences of the direct physical interaction between E2A and CBP/p300. The overall, longer-term objective is the elucidation of potential novel mechanisms of leukemia induction by E2A-proteins and, consequently, the identification of new targets for therapeutic intervention. 3 Chapter 2 Literature Review 2.1 Structure and Function of the E2A Gene Products 2.1.1 E2A Gene Products are Basic Helix-Loop-Helix Proteins The E2A gene products, E12 and E47, referred to herein as E2A proteins, represent the founding members of the basic helix-loop-helix (bHLH) family of transcription factors. The carboxy-terminus of E2A proteins contains two regions of conserved hydrophobic residues separated by a linker sequence (16). Examination of these residues using a helical representation revealed two amphipathic helices separated by the linker sequence containing amino acids characteristic of a loop structure. This motif was subsequently designated as helix-loop-helix (HLH) (16). Additional structural analysis revealed a region of basic amino acids immediately amino-terminal to the HLH motif (17) responsible for mediating interaction with the major groove of DNA, while the HLH structure mediates homo- or hetero- dimerization of two HLH containing proteins. Dimerization of bHLH proteins is independent of DNA binding, however dimerization is essential for DNA binding at the consensus DNA sequence CANNTG. This sequence, named the E-Box, was originally identified in the enhancers of the immunoglobulin heavy chain (IgH) and kappa light chain genes (18-20) but has since been found in a variety of other cell-type specific gene promoters and enhancers. 4 Several other bHLH containing proteins have been identified since the identification of the bHLH domain in E2A proteins. These have been characterized based on structural similarities, tissue distribution and interaction with other HLH containing proteins. E12 and E47 belong to Class I bHLH proteins. This class of proteins is widely expressed in many tissues and includes the related proteins HEB, E2-2 and Drosophilia protein daughterless (da) (16). Class I proteins heterodimerize with Class II bHLH proteins. The latter show restricted expression in specific tissues or developmental stages. MyoD (21,22), MRF4 , Myf5 and myogenin (23), for example, are muscle specific bHLH proteins involved in regulating myogenic differentiation in combination with E2A proteins; specifically, MyoD forms heterodimers with E2A proteins (24). 2.1.2 E2A Contains Two Transcriptional Activation Domains E2A proteins are proposed to function as transcriptional activators mediating expression of target genes containing E-Box elements. Henthorn et al (25) noted in yeast and mammalian cells that the amino terminal two-thirds, i.e. the portion not encoding the bHLH domain, of the E2A splice variant E2-5 and a related bHLH protein E2-2 were required for transactivation of reporter genes, suggesting the presence of transcriptional activation domain(s) in this region (25). Mapping studies using transient co-transfection of portions of E2A protein fused to the DNA binding domain of the yeast transcription factor GAL4 identified two independent transcriptional activation domains, AD1 and AD2, capable of inducing expression of a reporter gene. AD1 is located at the extreme amino terminus (amino acids 1-99) (10,26) while AD2 is centrally located (amino acids 394-406) (26,27) (Figure 1a). Originally considered to function independently, more 5 Figure 1: a) Domain structures of E2A, PBX1 and E2A-PBX1 fusion proteins. The numbers indicate the first and last amino acid positions in a domain or protein. The vertical line represents the fusion point between E2A and PBX1 proteins. AD, activation domain; bHLH, basic helix-loop-helix domain; HD, homeodomain; vertical red lines indicate amino acid differences in E47 compared to E12. b) The genomic E2A locus and location of protein domains or amino acids involved in the regulation of E2A function. Black lines represent exons. Exon 1 contains sequence for a 5’ untranslated region (UTR) as well as the translation start site. Exons 2-16 are coding exons. The blue and red boxes represent the alternatively spliced E12 and E47 exons (17a and 17b) encoding the helix-loop-helix domain of the proteins. A small part of exon 18 is also encodes the C-terminus of the protein as well as a large 3’UTR. * indicates a nuclear localization signal (NLS); # indicates a inhibitory domain involved in inhibition of DNA binding by E12 homodimers; S, serine residues known to be phosphorylated. 6 7 8 recent evidence from the LeBrun Lab suggests AD1 and AD2 act co-operatively to promote maximal E2A transcriptional activation (13). Activation domain 1 contains a stretch of highly conserved amino acids that form a potential α–helical domain located from residues 11-28 of E2A proteins. Mutagenesis of residues within the region that disrupt the α–helical conformation dramatically reduces the transcriptional activation capability of AD1 in both yeast and mammalian cells, indicating that the helix located within AD1 is required for transcriptional activation and interactions mediated by this domain are conserved between mammals and yeast (10,13). Located within the α–helical region of AD1 is Leucine-Aspartic Acid-PhenyalanineSerine (LDFS) motif that is highly conserved in class I HLH proteins, including E2A (10). Mutation of the LDFS motif (amino acids 20-23) significantly impairs transcriptional activation of reporter genes in yeast and mammalian cells (10). In addition, AD1 interacts with the SAGA co-activator complex in yeast in a manner that requires an intact LDFS motif (12). This interaction is required for transcriptional activation by AD1 in yeast, suggesting a role for co-activator complexes in E2Amediated stimulation of transcription. A highly conserved LXXLL (where X represents any amino acid, amino acids 16-20) is also present in this region of AD1. The LXXLL motif was first identified in nuclear hormone receptors as a binding site for co-activator proteins (28,29) further supporting a role for transcriptional co-activator complexes in E2A function. 9 2.2 E2A Proteins Are Required For B-Cell Commitment And Differentiation 2.2.1 E2A is Specifically Required for the Earliest Stages of B-Cell Differentiation A role in lymphocyte development was suggested by the original identification of the E2A proteins as immunoglobulin enhancer-binding proteins (16). Furthermore, ectopic expression of E2A in a macrophage cell line induces a more B-cell like phenotype (30) and over expression of E2A in a pre-T-cell line stimulates low but detectable levels of transcription and rearrangement of the immunoglobulin heavy chain gene (31). E2A knockout mice show severe reduction of B-lineage cells. Rearrangement of the immunoglobulin heavy chain segments is also absent, indicating that B-cell differentiation is blocked before the pre-B-cell stage (32,33). Expression of several genes that are regulated by E2A, and required for B-cell differentiation, is also significantly decreased in the knockout mice (34). In addition, mutant E2A protein lacking transcriptional activation domains but containing the bHLH domain is unable to induce expression of immunoglobulin genes or of several genes normally regulated by E2A, supporting the notion that the biological function of E2A requires the activation domains (30). Together, these results demonstrate a crucial role for the E2A proteins in promoting early stages of B-lymphoid development through regulating genetic recombination at the immunoglobulin loci and transcription of key target genes (34). 10 2.2.2 E2A is Involved in Later Stages of B-Cell Development In addition to being highly expressed (35) and active in early B-cell lineage commitment and differentiation (36), E2A shows increase mRNA levels in germinal centers (37) and is detectable in all secondary lymphoid tissues (38,39). Naïve mature B-cell express low levels of E2A proteins while activation of these cells results in induction of E2A protein expression and DNA binding (40). In addition, active B cells lacking E2A activity (i.e. DNA binding) fail to undergo immunoglobulin class switching (40). Together, these results indicate that E2A is also required at later stages of B-cell development. 2.3 Regulation of E2A Activity E2A proteins are expressed broadly in tissues and play a role in mediating differentiation of B-lymphocytes as well as muscle and other cells types (37,41). In addition, E2A plays a role in growth control, as over expression inhibits cellular proliferation and mediates cell cycle arrest (42). E2A levels are elevated in G0 arrested cells and decline with cell cycle progression, suggesting that E2A protein levels and/or activity must be downregulated for cell cycle progression (43). 2.3.1 Regulation of E2A by Dimerization with Other HLH Proteins E12 and E47 are generated by differential splicing of a single exon encoding the bHLH domain (Figure 1b). The bHLH domains of E12 and E47 show 80% amino acid identity (16), and the exons encoding the more amino-terminal polypeptide elements are invariant 11 in the isoforms E12 and E47.The differences in the basic DNA binding and HLH dimerization domains raise the possibility that these differences affect DNA binding and oligomerization with other proteins. In B-lymphocytes, E47 homodimers are stabilized by an inter-molecular disulfide bond and bind to DNA (44,45) while E12 proteins are less able to homodimerize and bind E-Box elements (16,24). Consistent with these observations, an inhibitory domain was identified in E12 that prevents it from forming homodimers but has no affect on heterodimerization with other class I and class II HLH proteins (46). A class of HLH proteins that lack a basic region has been identified and named Id, for Inhibitor of DNA binding (47). These proteins dimerize with E12, E47 and other bHLH proteins through a HLH domain, resulting in dramatically decreased DNA binding and transcriptional activation by heterodimers. Id expression is highest in immature progenitor-B (pro-B) cells and decreases as the cells differentiate to pre-B and mature Bcells (48,49). This pattern of expression correlates with up-regulation of E2A levels and function. In addition, over expression of Id1 in B-lineage cells impairs B-cell development in a mouse model (47,48) reminiscent of the phenotype seen in E2A knockout mice (32,33). Therefore, Id proteins appear to function physiologically as dominant-inhibitory regulators of E12/E47 function (47). 2.3.2 E2A Proteins are Substrates for Phosphorylation Protein phosphorylation regulates transcription factor function in a variety of ways, including by modulating DNA binding, protein-protein interactions and by targeting 12 transcription factors for proteolytic degradation (50). E2A proteins are phosphorylated in B-lymphoid and non-lymphoid cell lines (51). Further investigation revealed that phosphorylation at two serine residues (serine 514 and 529) of E47, adjacent to the bHLH domain, inhibits DNA binding by E47 homodimers but not E47/MyoD heterodimers (52). In addition, these two serine residues are hypo-phosphorylated in B-lineage cells compared to non-B-cells thus allowing DNA binding by E47 homodimers (53). In muscle progenitors, phosphorylation of E2A by the p38 kinase results in inhibition of E2A/MyoD transcriptional activation and myogenesis without effecting DNA binding, heterodimerization or protein levels (54). E2A proteins are also phosphorylated by several G1 cyclin-dependent kinases at amino-terminal serine residues (serine 48 and serine 154) to relieve E2A mediated growth suppression and allow entry into the cell cycle through a mechanism that does not affect E2A transcriptional activity or DNA binding (55). These results suggest that E2A proteins are regulated by phosphorylation downstream of signaling pathways that respond to extracellular signals or to cell cyclerelated cues. 2.3.3 E2A Stability is Regulated by Ubiquitination Regulated protein degradation is accomplished in cells through the ubiquitin-proteasome system (56). Ubiquitination of proteins involves a three step process culminating in covalent conjugation of the conserved 76 residue polypeptide ubiquitin to the ε-NH3+ group of lysine residues. Ubiquitin activating (E1) enzymes activate ubiquitin to generate 13 an E1-ubiquitin intermediate. Ubiquitin conjugating (E2) enzymes then transfer the ubiquitin from E1 to generate an E2-ubiquitin substrate that is specifically bound by an ubiquitin ligase (E3) enzyme complexed with the final substrate protein. The E3 enzyme transfers the ubiquitin to the ε-NH3+ group of a target lysine. Poly-ubiquitinated chains on lysine residues target proteins to the 26S proteasome where they are proteolytically degraded (56). E2A proteins contain three proline glutamine/aspartate, serine, threonine (PEST) rich regions, located within the bHLH region, within AD1 (amino acids 47-67) and at amino acids 169-189 (Figure 1b). PEST regions are commonly found within rapidly degraded proteins (57). E2A proteins interact with UBC9, a SUMO conjugating enzyme (58-62), in a manner that requires a bipartite binding site located at amino acids 476-494 and 505513, amino-terminal to the bHLH domain (Figure 1b) (63,64). Treatment of cells expressing E2A with proteasome inhibitors stabilizes E2A, as does anti-sense RNA to UBC9, suggesting that E2A is degraded through the proteasome (64,65). SUMOconjugation is not normally associated with degradation and E2A-SUMO conjugates have not been described suggesting that the E2A degradation domain (i.e. the UBC9 interactive region) may also be a recognition site for conventional ubiquitin conjugating enzymes or the E2A-UBC9 interaction may be a necessary intermediate step in the degradation pathway (65). Deletion of the UBC9 interacting region stabilizes E2A proteins and results in accumulation of hyper-phosphorylated E2A, suggesting that phosphorylation may be a 14 prerequisite for degradation of E2A through the proteasome (65). In fact, Notch-1induced inhibition of E2A transcriptional activity is a result of the induction of E2A ubiquitination and proteosomal degradation in a manner that requires the activity of MAP kinases (66-68). In addition, E2A phosphorylation promotes an interaction with Skp2 and the SCFSkp2 E3 ubiquitin ligase complex (66,69). 2.4 E2A Involvement in Translocations and Leukemia 2.4.1 The E2A Gene is Located on Chromosome 19 Using in situ DNA hybridization of a radio-labeled E47 probe to human metaphase chromosomes, Mellentin et al localized the E2A gene to the short (p) arm of chromosome 19 at band 13.2-13.3 (5). This region of chromosome 19 had previously been implicated in one of the most frequently reported chromosomal translocations associated with ALL, t(1;19)(q23,p13.3), described by several groups in 1984 (70-72). The E2A gene was further mapped to the breakpoint on chromosome 19 in eight leukemia specimens and two cell lines positive for the 1;19 translocation (5). Analysis of the RNA from one cell line indicated a larger than expected E2A mRNA, suggesting the existence of an E2Afusion gene in t(1:19)-positive leukemia cells and a pathological role in leukemogenesis (5). Sequencing of cDNAs representing fusion transcripts identified the translocation partner as PBX1 (3,5). 15 2.4.2 PBX1 Structure and Function The PBX1 gene was discovered and cloned as a result of its involvement in the 1;19 translocation (3,4). The gene product, PBX1, is a member of the three amino acid loop extension (TALE) class of homeodomain (HD) proteins (73). Differential splicing at the extreme 3’ end of the PBX1 pre-mRNA results in expression of two protein products, PBX1a and the shorter PBX1b (Figure 1a) (3,4). PBX1 interacts with a conserved hexapeptide sequence of HOX proteins located adjacent to the HOX HD (74-76). Reciprocally, HOX proteins interact with the three amino acid insert located within the PBX1 HD. HOX-PBX1 heterodimers co-operatively bind to overlapping DNA binding sites on opposite faces of the DNA (77) to regulate expression of genes involved in determination of cell fate and embryonic development (78). Importantly, PBX1 is not normally expressed in B-cells (79). 2.4.3 E2A-PBX1 Structure and Function The chimeric oncoprotein E2A-PBX1 consists of the N-terminal two-thirds of E2A, including its two transcriptional activation domains, fused in frame to most of the PBX1 protein, including its DNA binding homeodomain (Figure 1a) (3). The E2A-PBX1 oncoprotein is a potent transactivator in several studies (80,81). In the most intuitive model of E2A-PBX1 mediated leukemogenesis, DNA binding of E2A-PBX1 occurs at PBX1-defined target sites and is mediated by the PBX1-derived homeodomain. The activation domains of E2A recruit co-activators and the general transcription machinery, 16 thereby activating expression of genes not normally expressed in B-lymphocyte progenitors. This abnormal gene expression pattern is thought to disrupt B-cell development, causing leukemia (80-82). However, it has been demonstrated that deletion of the DNA binding domain HD from E2A-PBX1 does not abrogate growth deregulation in fibroblast or induction of malignant lymphomas in transgenic mice (79,83). In fact, replacement of the entire PBX1 portion of E2A-PBX1 with DNA-binding and dimerization domains from the yeast transcription factors GAL4 or GCN4 also does not abrogate transformation of cultured fibroblasts. Forced homodimerization of E2A 1-483 in fibroblasts is capable of causing growth deregulation, suggesting that PBX may, at least in part, contribute to oncogenesis by mediating oligomerization of E2A domains (84). In contrast, deletion of the E2A-derived transcriptional activation domains from the chimeric protein abrogates the oncogenicity of the protein (13,79,84). These results suggest that interactions mediated by the E2A portion of E2A-PBX1 may be more important than DNA binding mediated by PBX1 at least for growth deregulation by the protein. In an alternative model proposed for E2A-PBX1 induced leukemogenesis, the E2A portion interacts with transcriptional co-activators and, perhaps in a dominantinhibitory manner, sequesters these key co-activators from wild-type E2A or other transcription factors with tumour suppressive functions (85). Disruption of transcription factor/co-activator interactions could result in altered gene expression at many loci and contribute to development of leukemia (12,25,84). These two proposed oncogenic mechanisms are not mutually exclusive and leukemogenesis by E2A-PBX1 likely involves aspects of both models. 17 2.5 Chromatin and Transcriptional Regulation 2.5.1 Chromatin Structure Nucleosomes represent the smallest unit of DNA compaction. They consist of 147 base pairs of DNA wrapped 1.65 turns around an octamer of core histones. Each nucleosome contains two units of each of the core histones, H2A, H2B, H3 and H4 (86). Individual nucleosomes are joined by a short segment of linker DNA of approximately 10 to 80 base pairs in length. This linker DNA is bound and stabilized by histone H1 proteins and the nucleosomes are then further compacted into higher order chromatin structures. These exclude other proteins from interacting with DNA and must be modified in order for the DNA to become more accessible to transcription factors, the RNA polymerase II initiation complex and the DNA replication machinery (87,88). In eukaryotic organisms, gene transcription must be highly regulated so as to ensure that genes are expressed at the proper time and in the correct cell-types. Gene transcription is regulated by sequence specific DNA binding of transcription factors to promoter sites followed by recruitment of co-regulatory complexes and consequent repression or enhancement of gene transcription (89). Transcriptional initiation requires alteration of the chromatin structure and recruitment of the basal transcription machinery. Two general classes of co-regulators have been described: a) those that covalently modify histones; and, b) ATP-dependent chromatin remodelers (90). 18 2.5.2 Enzymes That Covalently Modify Histones While the structured globular domains of the histone octamer are located within the nucleosome particle, flexible amino-terminal histone tails extend freely from the core. These histone tails are positively charged, lysine rich and contain sites amenable to posttranslational modifications important for chromatin function. Several different modifications have been identified on histones, including: acetylation, methylation, sumoylation, ubiquitination and phosphorylation (Figure 2a). Perhaps the two best characterized histone modifications are acetylation and methylation (91). Actively transcribed genes are associated with increased histone acetylation while heterochromatic, untranscribed regions of the genome are typically associated with hypoacetylated histones (92,93). Several enzymes have been identified that are able to transfer an acetyl group from acetyl-co-enzyme A to the ε-NH3+ group of lysine residues, mainly in the histone tails (Figure 2b). These enzymes, called histone acetyltransferases or HAT proteins (94), can be recruited to promoters by interaction with sequence-specific DNA binding transcriptional activators (87,88). Subsequent acetylation of the surrounding nucleosomal histones and the non-histone, chromatin-associated high mobility group (HMG) proteins (95,96) neutralizes the positive charge of the lysine residues, affecting the contacts between histones, histone and DNA as well as binding of regulatory proteins to acetylated chromatin (94). Acetylation of lysine residues is a reversible process and another group of histone modifying enzymes, the histone deacetylases (HDACs) are able to remove the acetyl group allowing the DNA to return to the repressive state (97). 19 Figure 2: Histone Modifications and Lysine Acetylation. a) Nucelosome core particles showing 8 N-terminal tail domains. Post-translational modifications known to occur on each tail are indicated by coloured symbols. Modified from (98) b) Histone acetyltransferase (HAT) enzymes are able to catalyze the transfer an acetyl group from acetyl-co-enzyme A to the ε-NH3+ group of lysine residues resulting in the neutralization of the positive charge of the lysine residues. 20 21 Histone methylation is also implicated in transcriptional control. Methylation occurs at lysine and arginine residues by a class of enzymes called histone methytransferases (HMTs) and can be either repressive or activating, depending on the specific lysine or arginine modified in each histone. To make matters more complex, HMT enzymes can add one, two or three methyl groups to each substrate amino acid. As with acetylation, methylation of lysines and arginines is reversible by several demethylating enzymes (99). Both acetylation and methylation create binding sites for protein-protein interactions, resulting in recruitment of additional factors involved in transcription. Acetylated lysines, for example, are recognized by bromodomains of chromatin-interactive proteins while methylated residues are recognized by chromodomains. Both bromo- and chromodomains are found in many histone modifying enzymes as well as in components of the ATP-dependent remodeling complexes and the basal transcription machinery (99). 2.5.3 Chromatin Remodeling Enzymes Chromatin remodeling enzymes are found within large multi-subunit complexes and function to reposition DNA on nucleosomes. All chromatin remodeling complexes contain an ATPase subunit belonging to the SNF2 superfamily of DNA helicase/ATPases and use the energy from ATP-hydrolysis to alter protein-DNA interaction. The two best characterized sub-families of ATP-dependent remodeling complexes are: 1) SWI/SNF; and, 2) ISWI (100). 22 The SWI/SNF complex was first described in yeast as a regulator of mating type switching (101). Human homologues have subsequently been described and are involved in regulating gene expression. SWI/SNF-type complexes contain subunits capable of interacting with DNA and histones. The histone binding activity is mediated by a bromodomain found within the catalytic subunit, that recognizes acetylated lysines in histone tails. In addition to interacting with acetylated histones, SWI/SNF complexes interact with components of the basal transcription machinery and RNA pol II and are generally localized to actively expressed genes (102). SWI/SNF complexes are also known to interact with sequence-specific DNA-binding transcriptional activators (102). For example, acetylation of erythroid Kruppel like-factor (EKLF) by CBP enhances its interaction with the SWI/SNF complexes resulting in enhanced transcriptional activation (103). Unlike the SWI/SNF complexes, ISWI complexes have not been found to associate with RNA pol II and have been suggested to reposition nucleosomes to establish a repressive chromatin environment. ISWI complexes are primarily associated with non-transcribed regions of the chromatin (104) and contain an activity not present in SWI/SNF type complexes, the ability to generate regularly spaced nucleosomes. This suggests that ISWI type complexes play a role in maintaining chromatin structure (100). However, recently ISWI components containing plant homeodomain (PHD) motifs were shown to be able to recognize histone 3 (H3) tri-methylated at lysine 4 (H3K4me3) (105,106), a histone modification associated with transcription start sites of active genes (107). These results 23 suggest that activation or repression of gene transcription by ISWI complexes is promoter dependent (100). 2.6 CBP and p300 are Transcriptional Co-Activators Two of the most widely studied transcriptional co-activators are CREB binding protein (CBP) and p300 (Figure 3). CBP is a large (265 kDa) protein that was originally identified in a screen for proteins that interact with the phosphorylated form of the transcription factor CREB (108). The phospho-CREB-CBP interaction stimulates CREBmediated expression of a target gene over that induced by unphosphorylated CREB (109,110). p300 was identified as an adenoviral E1A interacting protein (111,112). Analysis of the p300 and CBP amino acid sequences reveals that the two proteins are highly similar, especially over a centrally located 800 amino acid span (85% identical, 95% homologous) (113). This region includes a bromodomain as well as the C/H3 region identified by Eckner et al as responsible for p300 binding to E1A (114). Since CBP and p300 are highly identical and can substitute for one another in many, although not all, functional assays, they are often referred to collectively as CBP/p300 (115,116). Several studies have also determined that CBP and p300 are capable of interacting with the basal transcription factors TFIIB (109,117) and TATA binding protein (117,118) and form a complex with RNA polymerase II (119-122). Along with the evidence that CBP and p300 stimulate transcriptional activation by CREB (109,110), these results support the idea that CBP/p300 are transcriptional co-activator molecules. 24 Figure 3: CREB-binding protein (CBP) organization and interactions. C/H, cysteine/histidine rich regions 1-3; The KIX domain mediates interactions with a large number of transcription factors including CREB, c-Myb and E2A. The bromodomain (BROMO) mediates protein-protein interactions by association with acetylated lysines. HAT, the region of CBP containing histone acetyltransferase catalytic activity. TAZ, a zinc-finger domain involved in interactions with multiple proteins. Some known CBPinteracting proteins, discussed in this thesis, are indicated below the functional domains. The numbers represent the first and last amino acids of domains or the human CBP protein. 25 26 2.6.1 CBP and p300 Have Intrinsic Acetyltransferase Activity One mechanism of CBP/p300 mediated stimulation of transcription was postulated to be through recruitment of the p300/CBP Associated Factor (p/CAF) to promoters. p/CAF was cloned based on homology to the yeast GCN5, a transcriptional co-activator with intrinsic AT activity (123,124). Like GCN5, p/CAF also efficiently acetylates the free core histones H3 and H4 as well as nucleosome particles (123). p/CAF was subsequently shown to interact with p300 and CBP at a region adjacent to the E1A binding site (123). In fact, binding of p/CAF to CBP/p300 inhibits binding by E1A in a competition assay and E1A is able to disrupt the p300-p/CAF interaction in vivo (123). Based on this discovery, E1A was used as a ligand to recruit interactive proteins from nuclear extracts. E1A recruits significant amounts of HAT activity while an E1A mutant unable to interact with CBP/p300 cannot recruit HAT activity (125). Recombinant p300 and CBP were subsequently tested for their ability to acetylate histones. A region located between the bromodomain and the E1A binding site that is 91% similar between CBP and p300 was identified as the HAT catalytic domain (126). Unlike p/CAF, p300 preferentially acetylates H4, suggesting that the two HAT proteins may co-operate to acetylate histones at promoters. The HAT domain of CBP/p300 does not show any sequence similarity to the HAT domains of GCN5 and p/CAF, consistent with CBP/p300 and GCN5/p/CAF belonging to separate families of HAT proteins (125). 27 2.7 Acetylation and Transcription Factor Regulation Acetylation of histones has been convincingly identified as a key modulator of transcriptional activity. Hyper-acetylated histones are consistently associated with actively transcribed genes while hypo-acetylation and recruitment of HDAC enzymes is associated with a repressive chromatin state (99). Beyond histones, a growing list of nonhistone proteins have been identified as substrates for acetylation by co-activators, including CBP/p300 and p/CAF (15,127). Acetylation of transcription factors has been demonstrated to have a variety of functional effects and has been postulated to be as important a regulator of protein function as phosphorylation (128). Acetylation of lysines can have positive or negative effects on protein interactions with DNA. For example, acetylation of lysines adjacent to the DNA binding domain of p53 results in enhanced DNA binding (129) while acetylation within the DNA binding domain of High Mobility Group 1 protein (HMG1) antagonizes DNA binding, likely by neutralizing a positive charge that contributes to interaction with negatively charged DNA (130). Acetylation also plays a role in regulating protein-protein interactions by transcription factors. Acetylation of MyoD, for example, results in a higher affinity interaction with the bromodomain of CBP/p300 (131). Acetylation is also able to destabilize protein-protein interactions. The interaction between the nuclear hormone receptor co-activator ACTR 28 and promoter-bound estrogen receptor is disrupted by acetylation by CBP, resulting in down regulation of transactivation (132). Poly-ubiquitination of lysines targets proteins for degradation by the proteasome. Since a particular lysine residue cannot have two post-translational lysine modifications simultaneously, prior acetylation can antagonize ubiquitination, thereby affecting protein stability. For example, several lysine residues in p53 are substrates for both ubiquitination and acetylation. Acetylation of p53 prevents ubiquitination and stabilizes p53 (133,134). In conclusion, transcription factors are direct substrates for acetylation by transcriptional co-activator/acetyltransferases and this modification is capable of altering transcription factors function through multiple mechanisms, including effects on DNA binding, proteins stability, sub-cellular localization and protein-protein interactions (15). 2.8 Recognition Motifs for Acetyltransferase Proteins Examination of a large number of polypeptide sequences that are acetylated by various HATs has failed to uncover a strictly conserved consensus recognition motif. Therefore, it is not possible to identify potential acetylation sites by sequences analysis alone (94). Some evidence exists to suggest that lysine (K) preceded by glycine (G), alone (135) or as part of a larger GKXXP motif (where X equal any amino acid and P represents proline), represents a weak consensus for lysine acetylation for CBP/p300 and p/CAF (94). 29 Another potential consensus, KX1-2XKK, is apparent in several transcription factors. For example, both CBP/p300 and p/CAF have been found to acetylate MyoD at a group of closely spaced lysine residues (K99, K101, K102) adjacent to its bHLH domain, resulting in increased DNA binding and transcriptional activation. (136-138). Acetylation of p53 also occurs at a cluster of closely spaced, evolutionarily conserved lysines (129) as does acetylation of c-Myb (139,140). 2.9 CBP Interacts Directly with E2A Proteins 2.9.1 Identification of E2A-CBP Interactive Domains A particular requirement for AD1, the amino-terminal activation domain of E2A in E2Amediated transcriptional activation (10,12,141) as well as growth deregulation by oncogenic E2A-fusion proteins (84,141) has been noted. In the LeBrun lab, an unbiased affinity screen was performed to identify proteins that interact with this region of E2A. The transcriptional co-activator CBP was identified as an AD1-interactive protein in this screen (unpublished results). The E2A-CBP interaction was interesting since a functional interaction between E2A and CBP has been suggested in the literature. Several groups have shown enhanced E2Amediated transcriptional activation of target genes when E2A is co-transfected with CBP (11,142,143). Furthermore, co-expression of E2A and p300 in NIH 3T3 fibroblast cells enhances E2A-mediated transcription through the immunoglobulin heavy chain enhancer 30 (11). Subsequent to the identification of CBP as an AD1 interactive protein, the LeBrun lab has examined the domains required for both E2A and CBP to mediate the interaction. Activation domain 1 of E2A contains a phylogenetically conserved α-helical region (amino acids 11-28) (10) and is required for interaction with the yeast co-activator complex SAGA suggesting this region may mediate interactions with mammalian coactivators (12). An α-helical region within AD2 has also been identified but has no homology to the helical region of AD1 (27). The α-helical regions of AD1 and to a lesser extent, AD2 were found to contribute to the recruitment of CBP/p300 and p/CAF from crude nuclear extracts and full transcriptional activation of a reporter gene by E2A 1-483 indicating a role for the E2A-CBP interaction in regulating E2A transcriptional activation activity (13). Mapping of the E2A-interactive domain on CBP determined that the CREB-binding domain or KIX domain, is required and sufficient for the interaction with E2A (13). The KIX domain has been well characterized and is known to mediate interactions with the transcriptional activator domains of several proteins through interactions with LXXLL motifs (28,144-147). In E2A, an LXXLL motif is found as an amino-terminal extension of the LDFS motif (LXXLLDFS). The interaction between E2A and the KIX domain of CBP was subsequently shown to be direct and involve the LXXLLDFS motif (13,14). In fact, interaction between an E2A peptide containing this region of AD1 of E2A and the KIX domain promotes the adoption of an α-helical structure by the E2A peptide (14,148). Targeted amino acid substitution of the common leucine residue in the 31 LXXLLDFS motif, to alanine (L20A), impairs the interaction with CBP as well as the adoption of a helical conformation due to binding (14). Since AD1 of E2A is required for transcriptional activation (10,12,141) and growth deregulation by oncogenic E2A-fusion proteins (84,141) it is reasonable to suspect the E2A-CBP interaction might be involved in mediating the neoplastic functions of E2A-fusion proteins. 2.9.2 Functional Significance of the E2A-CBP Interaction for E2A-PBX1-mediated Leukemogenesis The E2A-PBX1 oncoprotein is capable of inducing the outgrowth of immortal, highly proliferative myeloid progenitors from retrovirally transduced primary mouse bone marrow (149). Abrogation of the E2A-CBP interaction by deletion of the AD1 α-helix alone, or in combination with, the AD2 α-helix reduced cellular proliferation of transduced cells (13). Furthermore, disruption of the E2A-CBP interaction by engineered mutations of the AD1 α-helical region (e.g. L20A) in E2A-PBX1 abrogated the ability of E2A-PBX1b to stimulate proliferation of primary hematopoietic cells retrovirally transduced to express E2A-PBX1b demonstrating a critical role for the LXXLLDFS motif in mediating growth deregulation by E2A-PBX1 (14). In a whole animal model sub-lethally irradiated mice reconstituted with bone marrow transduced with E2A-PBX1b-L20A did not develop an aggressive myeloproliferative disease seen in animals reconstituted with E2A-PBX1b transduced bone marrow (14). 32 This result suggests a role for the LXXLLDFS motif and interaction with the CBP KIX motif in mediating the oncogenic effects of E2A-PBX1 (13,14). In summary, the interaction between AD1 of E2A and the KIX domain of CBP is direct and required for E2A-PBX1-mediated oncogenesis. The E2A-PBX1-CBP interaction could potentially contribute to leukemogenesis in several ways: First, E2A-mediated recruitment of co-activators, such as CBP, to PBX1 defined target genes may promote aberrant transcriptional activation. Second, in a DNA binding independent manner, interaction between AD1 of E2A-PBX1 might sequester CBP, in a dominant-inhibitory fashion, from wild-type E2A or other factors. Finally, the association between E2APBX1 and CBP might alter the enzymatic activity of CBP itself. 33 Chapter 3 Hypothesis and Research Objectives The amino-terminal activation domain of E2A, AD1, is critical for transcriptional activation by wild-type or chimeric E2A proteins (10,26) as well as growth de-regulation and oncogenesis by E2A-PBX1 (13,83,84,141). This region mediates interactions with the histone acetyltransferase co-activator proteins CBP/p300 and p/CAF (13). The interaction between E2A-PBX1 and CBP/p300 is required for leukemia induction by E2A-PBX1 (13,14). Therefore, the direct physical interaction between E2A proteins and CBP/p300 has important functional consequences for both proteins. My research was undertaken with the general objective of elucidating these consequences. Hypothesis The direct physical interaction between E2A transcription factors and co-activator proteins CBP/p300 will result in post-translational modification of lysine residues within E2A. These modifications will affect the transcriptional activation activity of E2A. Furthermore, the interaction between E2A and CBP/p300 may affect the intrinsic acetyltransferase activity of CBP/p300 proteins. Research Objectives 1) Determine if the physical interaction between E2A and co-activator proteins results in acetylation of E2A itself and identify the acetyltransferase enzyme responsible. 34 2) Map the acetylated lysine residues within the E2A portion of E2A-PBX1 3) Determine if acetylation affects E2A-mediated protein-protein interactions 4) Determine the effect of E2A acetylation on the transcriptional regulatory function of E2A 5) Determine if the physical interaction between E2A and CBP affects the acetyltransferase activity of CBP and map the required domains on both proteins. 35 Chapter 4 Materials and Methods 4.1 Plasmids Plasmids conferring bacterial expression of the glutathione-S-transferase (GST)-E2A fusion proteins were generated by PCR amplification using a vector containing the full length E2A-PBX1 cDNA and ligated into BamHI and EcoRI sites of the GEX-2T vector (Amersham-Pharmacia Biotech) as described in (13). Site directed mutagenesis of lysine residues was performed using the Transformer Site-Directed Mutagenesis kit (Clontech). Amino acid substitutions were verified by sequencing. Mammalian plasmids conferring expression of GAL4-E2A proteins were generated by restriction digestion of the GST-E2A plasmids with EcoRI followed by creation of a blunt end using Klenow and digestion with BamHI. The E2A insert DNA was ligated into the CMV-GAL4 vector, digested with SmaI and BglII. Plasmids conferring bacterial expression of the glutathione-S-transferase (GST) fusion proteins GST-CBP-HAT, GST-PCAF-HAT, GST-p45 and GST-f(GATA-1) have been described previously (150-153) and were generous gifts of Dr. Gerd Blobel ( Children’s Hospital of Philadelphia, Philadelphia, Pennsylvania, USA). Mammalian expression vectors conferring expression of full-length wild-type and mutant CBP proteins (i.e. CMV5-CBP-WT-FLAG, CMV5-CBP-ΔKIX-FLAG, CMV5-CBP-ΔBROMO-FLAG and CMV5-CBP-ΔHAT) were also generous gifts of Dr. Gerd Blobel. 36 The mammalian expression vector pCMV-FLAG-SUMO-1 was a generous gift of Dr. G. Nucifora (154); pMT-35 c-JUN-HIS and pMT 123 HA-ubiquitin were generous gifts of Dr. Dirk Bohmann (University of Rochester) (155,156). 4.2 Preparation of GST-fusion Proteins Plasmid DNA was electroporated into competent BL-21 (DE3)pLysS or BL21CodonPlus-RP E.coli cells (Stratagene). Bacteria were grown in 2YT media supplemented with ampicillin (100μg/ml) (BL21) or ampicillin (100μg/ml) and chloramphenicol (50μg/ml) (BL21-RP). Protein was purified essentially as described in the GST Gene Fusion System handbook (Amersham-Pharmacia Biotech 3rd edition, Revison 2, 1997), dialyzed against Phosphate Buffered Saline (PBS) and stored at -20°C in 50% glycerol (v/v). Proteins used for in vitro HAT assays were dialyzed against BC100 Buffer (20mM TrisHCl pH 8.0, 100mM KCL, 0.5mM EDTA pH8.0, 20% glycerol, 0.5mM DTT, 0.5mM PMSF) after elution, concentrated with Centricon spin columns (Millipore) and stored at -80°C. Protein concentrations were determined by Coomassie brilliant blue staining of SDS-PAGE gels by comparison with known amounts of bovine serum albumin (BSA). 37 4.3 Preparation of nuclear extracts The pre-B acute lymphoblastic leukemia cell line RCH ACV associated with t(1;19) and expressing endogenous E2A-PBX1 protein, was grown to confluency in 100mm diameter tissue culture dishes in RPMI supplemented with 10% fetal bovine serum. Cells were harvested and nuclear extracts prepared using the Dignam and Roeder method (157). Briefly, the RCH ACV cells are pelleted and resuspended in a hypotonic buffer (Buffer A, 10mM Hepes-KOH pH7.9, 1.5mM MgCl2, 10mM KCl, 0.5mM DTT, protease inhibitors). The cell pellet is homogenized using a dounce homogenizer and checked for the presence of nuclei. The nuclei are pelleted by centrifugation and resuspended in a hypertonic buffer (Buffer B, 20mM Hepes-KOH pH7.9, 25% glycerol, 0.42M NaCl, 1.5mM MgCl2, 0.2mM EDTA pH8, protease inhibitors). The nuclei are homogenized and the solution stirred on ice for at least one hour. The nuclear solution is centrifuged and the supernatant containing nuclear proteins is dialysed for 5 hours at 4°C against Buffer D (20mM Hepes-KOH pH7.9, 20% glycerol, 0.1M KCl, 0.2mM EDTA pH8, 0.5mM DTT). Following dialysis the extract is centrifuged to remove proteins, including histones, which have precipitated. The supernatant is aliquoted and stored at –80°C. 4.4 Pull-downs using nuclear extracts 5 to 20μg of GST-E2A fusion protein bound to glutathione sepharose (AmershamPharmacia Biotech) beads were incubated overnight at 4°C with 0.5 to 1mg of RCH ACV nuclear extracts. Beads and retained proteins were washed in HEGN buffer (20mM 38 HEPES-KOH, pH 7.9, 0.1mM EDTA, pH 8.0, 10% glycerol, 0.1% NP4, 1mM DTT, 0.5mM PMSF) and analyzed by SDS-PAGE and immunoblotting. 4.5 Pull-downs using purified, recombinant CBP, p300 or p/CAF GST-E2A 1-483 (2.5μg) was bound to glutathione sepharose and blocked in HEGN buffer containing 1mg/ml Bovine serum albumin prior to incubation with 0.1pmol p300 or p/CAF. GST-E2A 1-483 (5pmol) was bound to glutathione sepharose and blocked in HEGN buffer containing 1mg/ml bovine serum albumin prior to incubation with 5pmol of CBP. In both experiments the proteins were incubated together overnight at 4°C. Beads were washed 3 times in HEGN buffer and retained proteins were analysed by SDS-PAGE and immunoblotting. 4.6 In Vitro acetylation site mapping 0.4μg of enzyme (GST-CBP-HAT or GST-PCAF-HAT) and 4.0μg of GST fusion protein are mixed in a final reaction volume of 30μl containing acetyltransferase (HAT) buffer (50mM Tris-HCl, pH 8.0, 50mM NaCl, 10% glycerol, 2.5mM MgCl2, 10mM sodium butyrate, 0.5mM dithiothretol (DTT), 0.5mM phenylmethylsulfonyl fluoride (PMSF)) (150) and 1μl 14C-acetyl-coEnzyme A (51.4 mCi/mmol, 0.02mCi/ml) (NEC313 Perkin Elmer) Reactions were incubated at 30°C for 1-2 hours. Acetylation products were 39 analyzed by SDS-PAGE and visualized following fluorography of the gel using En3Hance (NEF981 Perkin Elmer) and autoradiography. Acetylation levels were determined by scintillation counting of acetylated bands or quantified using a Phosphor screen and Phosphor Imager. In reactions using purified, recombinant p300 or p/CAF, 0.1pmol of enzyme was incubated with 5pmol of GST-E2A in HAT reaction buffer with and 1μl 14C-acetylcoEnzyme A for 2 hours at 30°C. Acetylation products were analyzed by SDS-PAGE and visualized following fluorography of the gel using En3Hance and autoradiography. Acetylation levels were quantified using a Phosphor screen and Phosphor Imager. 4.7 Pull-Down/Acetyltransferase reaction 5 to 20μg of GST-E2A fusion protein bound to glutathione sepharose (AmershamPharmacia Biotech) beads were incubated overnight at 4°C with 0.5 to 1mg of RCH ACV nuclear extracts. Beads and retained proteins were washed three times in HEGN buffer (20mM HEPES-KOH, pH 7.9, 0.1mM EDTA, pH 8.0, 10% glycerol, 0.1% NP40, 1mM DTT, 0.5mM PMSF). Beads were washed once in 1X HAT buffer, resuspended in 30μl HAT buffer plus 1μl 14C-acetyl-coEnzyme A and incubated at 30°C for 1-2 hours. Products were analyzed by SDS-PAGE and viewed following fluorography of the gel using En3Hance and autoradiography or are transferred to nitrocellulose and immunoblotted. 40 Immunoblotting was performed using antibodies specific for E2A (sc-416 Santa Cruz) or CBP (s-c7300 Santa Cruz). 4.8 Preparation of full-length CBP, p300 and p/CAF proteins Plasmid conferring expression of full-length CBP proteins were transiently transfected into SV293T cells, maintained in DMEM supplemented with 10% fetal bovine serum, using the calcium phosphate method. Two days after transfection cells were harvested and lysed in 50mM Tris pH 7.5, 150mM NaCl, 1mM EDTA, 1% Triton X-100 with protease inhibitors. Lysates were cleared by centrifugation and incubated with 20μl AntiFLAG M2 agarose (Sigma-Aldrich) per ml of lysates for 2 hours at 4°C. Beads were washed 3 times in TBS and the FLAG-tagged CBP eluted using excess 3X-FLAG peptide (200ng/μl). Proteins were aliquoted and stored at -80°C. The protein concentration was determined by Coomassie brilliant blue staining of SDS-PAGE gels by comparison with known amounts of bovine serum albumin. Full-length HIS-tagged p300 and FLAG-tagged p/CAF proteins were produced by baculovirus infection of the SF21 insect cell line (Generous gift of Dr. Roger Deeley, Cancer Research Institute, Queen’s University, Kingston, Ontario). Baculovirus supernatants for p/CAF and p300 and purification protocols were provided by Dr. Jeffrey Dilworth (Ottawa Health Research Institute, Ottawa, Ontario) with permission from Dr. Yoshihiro Nakatani (123) and Dr. W. Lee Kraus (158). 41 For HIS-tagged p300, cells were harvested 2 days after infection and resuspended in homogenization buffer (10mM Tris-HCl, pH 7.5, 0.5M NaCl, 10% glycerol (v/v), 0.1% NP40 (v/v), 15mM imidazole, 2mM β-mercaptoethanol, 2mM PMSF, 50μg/mL aprotinin) Cells were lysed by homogenization and cleared by centrifugation. The lysate was then incubated with nickel-NTA agarose (Qiagen) for 2 hours followed by four washes of the matrix in wash buffer (10mM Tris-HCl, pH 7.5, 0.2M NaCl, 10% glycerol (v/v), 0.2% NP40 (v/v), 15mM imidazole, 2mM β-mercaptoethanol, 2mM PMSF). The p300 protein was eluted in elution buffer (10mM Tris-HCl, pH 7.5, 0.2M NaCl, 10% glycerol (v/v), 0.1% NP40 (v/v), 250mM imidazole, 2mM β-mercaptoethanol, 2mM PMSF), aliquoted and stored at -80°C. The protein concentration was determined by Coomassie brilliant blue staining of SDS-PAGE gels by comparison with known amounts of bovine serum albumin. FLAG-tagged p/CAF, was purified from SF21 cells 2 days after infection. Cells were washed twice in PBS and resuspended in 1X FLAG Buffer (20mM HEPES- KOH pH 7.6, 150mM KCl, 2mM MgCl2, 0.2mM EDTA, 10mM β-glycerophospahte, 15% glycerol (v/v), 1mM DTT, 0.2mM PMSF). Cells were lysed by dounce homogenization and the cellular debris was pelleted by centrifugation. Cellular extract was incubated with AntiFLAG M2 agarose with 0.2% NP40 for 2 hours. The extract/agarose mix was added to a column, washed once with high salt FLAG buffer (0.6M KCl, 2mM DTT, 2mM PMSF, 0.2% NP-40) and once with low salt FLAG buffer (1X Flag buffer containing 1mM DTT, 0.3mM PMSF, 0.1% NP40). Proteins were eluted with 1X FLAG buffer containing 42 2mg/ml 3X-FLAG peptide and 1mM PMSF. Proteins were aliquotted and stored at 80°C. The protein concentration was determined by Coomassie brilliant blue staining of SDS-PAGE gels by comparison with known amounts of bovine serum albumin. 4.9 Luciferase Assays HeLa cells were maintained in DMEM supplemented with 10% fetal bovine serum. The day before transfection 1.7 X105 cells/well were plated in 12 well plates. Cells were transiently transfected by the calcium phosphate method. Transfection cocktails were prepared using an 8:1 ratio of reporter to transactivator, total 6μg DNA per well (5.3μg 5X GAL4-luciferase reporter, 0.7μg GAL4 or GAL4-E2A 1-483 plasmids and 0.05μg pRL-CMV a renilla luciferase-expressing plasmid (Gift of Chris Mueller, Queen’s University, Ontario, Canada)). The cells were lysed 40 h later in passive lysis buffer (Promega), and a portion of the lysates was analysed using the dual luciferase reporter assay system (Promega) and an LB 96V Micro Lumat Plus Microplate Luminometer (EG and G Berthold Ltd., Bundoora, Australia). Each luciferase value was normalized using the corresponding renilla value as a measure of transfection efficiency. The mean value from each triplicate experiment was considered as one data point and the results shown are the average of at least three independent experiments. 4.10 Histone Acetyltransferase Gel Assays Reactions using purified, recombinant p300 (0.1 pmol) was incubated with 5 pmol of GST-E2A and 2.5μg unfractionated calf thymus histones in 1X HAT reaction buffer with 43 and 1μl 14C-acetyl-coEnzyme A for 2 hours at 30°C. Acetylation products were analyzed by SDS-PAGE and visualized following fluorography of the gel using En3Hance and autoradiography. Acetylation levels were quantified using a Phosphor screen and Phosphor Imager. 4.11 Histone Acetyltransferase Filter Assays Filter assays were performed as essentially as described (159-161). A typical reaction contained 0.5 pmol of CBP protein and 5.0 pmol of GST or GST-E2A 1-483 in 25 μl final volume of 1X reaction buffer (50mM Tris HCl, pH8, 10% Glycerol, 50mM NaCl, 12.5mM MgCl2, 10mM sodium butyrate, 0.5mM DTT, 0.5mM PMSF) (162). Enzyme and E2A were incubated for 10 minutes at 4°C to allow interaction between E2A and CBP and 5 minutes at 30°C to equilibrate the temperature. The HAT reaction was induced by addition of 1.0μl 14C- acetyl-coenzyme A (60mCi/mmole) and 2.5μg unfractionated calf thymus histones (Sigma-Aldrich, H9250) or nucleosomes. Reactions were incubation at 30°C for 5 minutes prior to stopping the reaction by spotting onto P81 phosphocellulose filter discs (Whatman). Dried filters were wash twice for 5 minutes each, in 50mM Na2CO3 pH 9.2, rinsed in acetone and thoroughly air-dried before liquid scintillation counting. 4.12 Chloramphenicol Acetyltransferase Assays 44 U2OS cells and pcDNA3-GAL4-CBPHAT plasmids were gifts from Gerd Blobel (Children’s Hospital of Philadelphia, Philadelphia, Pennsylvania, USA). Cells were maintained in DMEM supplemented with 10% fetal bovine serum. The day before transfection 1.5 X105 cells/well were plated in 12 well plates. Cells were transiently transfected by the calcium phosphate method with mammalian expression and reporter plasmids as described (163). Briefly 2μg of pcDNA3-GAL4-CBPHAT-CBP2 or pcDNA3-GAL4-CBPHATΔ-CBP2 and 2μg of CMV-E2A, CMV-E2A-PBX1 or CMVPBX1 plasmids along with 2μg of the CAT reporter plasmid (5XGAL4-AdML-CAT) and 0.1μg pRL-CMV were used. The cells were harvested, resuspended in 100ul 25mM Tris, pH 7.5 and lysed by three freeze-thaw cycles. A portion of the lysate (60μl) was analysed for CAT activity using the FAST CAT Green (deoxy) Chloramphenicol Acetyltransferase Assay Kit (Molecular Probes) and thin layer chromatography (TLC) on silica gel plates following the manufacturers instructions. The chloramphenicol substrate in this kit has been modified to incorporate a green fluorophor and contains only one potential acetylation site resulting in only one acetylated species, thus the rate of acetylation is a direct function of enzymatic activity. Using a Lichtox Research Imaging Platform the TLC plates were excited with 470nm +/- 20nm light and emission at 520nm +/- 20nm imaged with a Hamamatsu Orca Camera. Imaging was performed with the assistance of Mr. Jeff Mewburn. The acetylated products were then quantified using Image Pro Plus (Media Cybernetics) software and expressed relative to GAL4-CBPHAT-CBP2 alone. A portion of the lysates (20μl) was also analysed using the dual luciferase reporter assay system (Promega) and an LB 96V Micro Lumat Plus Microplate Luminometer (EG and G Berthold Ltd., Bundoora, Australia). Each CAT value was normalized using the 45 corresponding renilla result as a measure of transfection efficiency. Each experiment was performed in duplicate. Results shown represent a typical experiment. 4.13 Immunoblotting Antibodies Following SDS-PAGE and electorphoretic transfer to a nitrocellulose membrane (PROTRAN nitrocellulose membrane NBA085C, Perkin Elmer) membranes were blocked of at least 1 hour in PBS containing 5% non-fat dried milk and 0.1% Tween 20. Primary antibodies were incubated overnight at 4°C in PBS containing 0.1% Tween 20 and 3% dried milk. Membranes were washed in PBS containing 0.1% Tween 20 and incubated with a secondary antibody conjugated to horseradish peroxidase (Jackson ImmunoResearch Laboratories) for 1 hour. After washing, the secondary antibody was detected using chemiluminescence, according to the manufacturer’s instructions (Western Lightning Chemiluminsecent Reagent Plus, Perkin Elmer). The following primary antibodies were used in this thesis. Anti-FLAG M2 (Sigma-Aldrich F-3165), anti-GAL4 (DBD) sc-577, anti-E2A (Yae) sc-416, anti-CBP (A-22) sc-369, anti HA (Y-11) sc-805, anti-HIS (H-15) sc-803, anti-CBP (C-1) sc-7300, anti-p300 (C-20) sc-585 (Santa Cruz Biotechnology). 46 Chapter 5 Acetylation Of Lysine 34 in E2A Proteins Contributes to Transcriptional Activation 5.1 Contributions Mutagenesis of K34Q and luciferase experiments shown in Figure 12 were performed by Mr. Richard Bayly. GAL4-E2A constructs used in luciferase assays were sub-cloned from existing GST-E2A vectors by Mr. Richard Bayly and Mr. Richard Casselman. Mr. Bayly was responsible for preparation of RCH ACV nuclear extracts. Initial stocks of purified, recombinant, full-length p300 and p/CAF were a generous gift of Dr. Jeffrey Dilworth. Additional protein stocks were prepared by Mr. Bayly by infection of SF21 cells with p/CAF and p300 baculoviruses and subsequent purification of these recombinant proteins. 5.2 Rationale The LeBrun lab has demonstrated that the amino terminal two-thirds of E2A (amino acids 1-483) can recruit the co-activator proteins CBP, p300 and p/CAF from nuclear extract prepared from the t(1;19)-positive, acute lymphoblastic leukemia cell line RCH ACV (13). The interaction between E2A and CBP was determined to involve α-helical regions located within the transcriptional activation domains AD1 and AD2 of E2A and the KIX domain of CBP. The consequences of the interaction between E2A and coactivator proteins on E2A function have not been fully described. This chapter will 47 describe experiments examining the effects of the interaction with CBP, p300 and p/CAF on E2A. 5.3 Results 5.3.1 The Portion of E2A Included in E2A-PBX1 Interacts with Acetyltransferase Proteins Capable of Acetylating Both Histones and E2A A biochemical approach was employed to determine if the co-activator proteins recruited by E2A retain intrinsic acetyl-transferase activity. Amino acids 1-483 of E2A were expressed in bacteria as an in-frame fusion with glutathione-S-transferase (GST) and purified on glutathione sepharose beads. GST-E2A 1-483 protein bound to beads was incubated with crude nuclear extract to allow protein-protein interactions to occur. The beads and associated proteins were then washed and transferred to acetyl-transferase reaction buffer in the presence of unfractionated calf thymus histones and 14C-acetyl coenzyme A. The reactions were analyzed by SDS-PAGE and fluorography. The GST-E2A 1-483 protein, but not GST, was capable of recruiting interacting proteins that acetylated histones (Figure 4a). When the experiment was repeated in the absence of histones and exposed longer the predominant band visualized after autoradiography corresponded in size (~80 kDa) to GST-E2A 1-483 (Figure 4b). A large acetylated species specific to the GST-E2A pull-down lane was also apparent well above the 175 kDa marker, indicating the presence of an unidentified E2A interactive protein of nuclear extract origin. This protein was subsequently identified as CBP by an anti-CBP 48 immunoblot (Figure 4c). These results indicate that the portion of E2A contained within the leukemogenic fusion protein E2A-PBX1 (i.e. E2A 1-483) can recruit proteins capable of acetylating histones and E2A itself. 49 Figure 4: (a) E2A recruits histone acetyl-transferase activity from nuclear extracts. GST or GST-E2A 1-483 fusion protein bound to glutathione-Sepharose beads were incubated overnight with lymphoid nuclear extract derived from RCH ACV cells. Retained proteins were subjected to an in vitro acetyltransferase (AT) assay in the presence of purified histones and 14C-acetyl Co-enzyme A and then analyzed by SDSPAGE and autoradiography. (b) E2A is acetylated by the recruited histone acetyltransferase activity. A similar experiment with omission of purified histones from the reaction and a longer exposure of the gel shows acetylation of GST-E2A 1-483. Acetylation of a large, unidentified protein of nuclear extract origin that migrates well above 175 kDa is also evident, in additional to several faint bands that migrate faster than GST-E2A 1-483. A least some of the latter could be truncation products related to GSTE2A 1-483. (c) A large acetylated E2A-interactive protein corresponds to CBP. AntiCBP immunoblotting identifies the large E2A-interactive protein as CBP. 50 51 5.3.2 Acetylation of E2A Correlates with Recruitment of CBP Our lab has reported that the interaction between E2A and CBP is direct and requires a conserved α-helical region located within the amino terminal 28 residues of AD1 and to a lesser extent involves an α-helical region located within AD2. Recruitment of p/CAF from crude nuclear extracts by E2A also requires these domains (13). Experiments were performed to determine if the same regions are required for recruitment of acetyltransferase (AT) activity and for acetylation of the E2A protein itself. A series of GST-E2A fusions with amino- and carboxy-terminal deletions (Figure 5a) were used as ligands in pull-down experiments performed with crude nuclear extracts. GST-E2A, or engineered variants, and retained interactive proteins were subjected to SDS-PAGE and immunoblotted with a monoclonal anti-CBP antibody (Figure 5b) or transferred to acetyltransferase buffer in the presence of 14C acetyl-Co-enzyme A before analysis by SDS-PAGE and fluorography (Figure 5c). Constructs able to pull-down CBP (i.e. 1-483, 1-273, 1-220, 1-160) were also acetylated while those that could not recruit CBP (i.e. 1-99, 29-273, 60-273, 100-273, 274-483) were not acetylated. Amino acids 1-28 of E2A are required for the CBP interaction and consequent acetylation of E2A even in the context of a relatively large portion of E2A (compare 1273 and 29-273, Figure 5 and (13)). This region of E2A comprises a phylogenetically conserved α-helical region containing an LDFS (amino acids 20-23) motif required for interaction with the SAGA co-activator complex and maximal E2A transcriptional 52 activity (10,12) and an LXXLL motif (amino acids 16-20; where X equals any amino acid) (28,29), thought to be involved in mediating interaction with co-activators. These results demonstrate that the conserved α-helical region of AD1, is required to recruit acetyl-transferase activity capable of acetylating E2A. E2A amino acids 1-160 are sufficient for CBP recruitment and contain four putative substrate lysine residues (Figure 5d). The correlation between CBP recruitment and E2A acetylation suggests that CBP may be the responsible HAT enzyme. 53 Figure 5: Acetylation of E2A correlates with recruitment of CBP. (a) Schematic indicating the domains of E2A-PBX1a and the segments of E2A expressed as aminoterminal fusions with GST. AD, activation domain; HD, homeodomain. (b) Mapping of CBP-interactive elements within E2A. GST fusion proteins were bound to glutathione sepharose and incubated with nuclear extracts in pull-down assays. Retained proteins were washed and analysed by anti-CBP immunoblots (c) E2A fragments that retain CBP from nuclear extracts also serve as substrates for acetylation. Pulldown assays were followed by incubation of the retained proteins in acetyltransferase buffer in the presence of 14C acetyl-CoA. The small arrow indicates an acetylated protein whose presence correlates with E2A acetylation and whose large size is consistent with CBP/p300. * indicates intact GST-E2A proteins.At least some smaller bands visible in the autoradiograph and coomassie stained gels are truncation products related. (d) An Nterminal α-helical region of E2A is required for interaction with CBP (anti-CBP immunoblot, upper image) and consequent acetylation of E2A 1-160 (autoradiograph, lower image). The amino acid substitutions (L19R/F22R) were designed to disrupt the E2A-CBP interaction. 54 55 56 5.3.3 In Vitro Acetylation of E2A The co-activator p/CAF co-purifies with CBP and possesses intrinsic AT activity (123) that could also potentially acetylate E2A proteins. Experiments were performed to determine whether recombinant CBP or p/CAF were capable of acetylating E2A. GST fusion proteins expressing only the catalytic acetyl-transferase domains of CBP (GSTCBP-HAT, amino acids 1196-1718) or p/CAF (GST-p/CAF-HAT, amino acids 352-832) were incubated with GST-E2A 1-483 in HAT reaction buffer in the presence of 14Cacetyl Co-enzyme A. The reactions were analysed by SDS-PAGE and fluorography. The catalytic domain of p/CAF was capable of acetylating GST-E2A 1-483; GST-p45 served as a positive control (Figure 6a). Although it readily acetylated GST-f(GATA-1), which served as a positive control, the catalytic domain of CBP was unable to acetylate GSTE2A 1-483 (Figure 6b). Neither catalytic domain could acetylate GST alone, indicating that both enzymes retained substrate specificity. In order to investigate the possibility that acetylation of E2A by CBP/p300 requires a full-length acetyltransferase, the HAT assays were repeated using recombinant, fulllength p/CAF and p300. Full-length p/CAF was capable of acetylating E2A, (Figure 6c) consistent with the results using the isolated catalytic domain. Interestingly, full-length p300 was capable of acetylating E2A, unlike the isolated CBP catalytic domain. Quantification of acetylation levels by Phosphor Imager revealed that p300 acetylated E2A at a level 2.54 times greater than p/CAF. When p/CAF and p300 were incubated simultaneously with GST-E2A 1-483, acetylation of E2A appeared greater than with either p300 or p/CAF alone. Quantification revealed greater than additive acetylation 57 Figure 6: Acetylation of E2A by purified CBP/p300 or p/CAF. (a) The isolated catalytic domain of p/CAF acetylates amino acids 1-483 of E2A (lane 1) and the positive controls p45 and GATA-1 (lanes 2 and 3, respectively). The auto-acetylated p/CAF catalytic domain migrates as two bands, indicated by the arrows. In lane 1, full-length GST-E2A 1-483 overlaps or is juxtaposed with the upper p/CAF-related band. However, multiple GST-E2A-related truncation products are readily visible. * indicates p45, @ indicates GATA-1, arrows indicate auto-acetylated p/CAF. (b) The isolated catalytic domain of CBP cannot acetylate recombinant E2A 1-483. The positive control GATA-1 is acetylated (lane 2). @ indicates GATA-1 (c) Both full-length recombinant p300 and p/CAF acetylate E2A 1-483. Co-incubation of E2A with p300 and p/CAF results in greater than additive acetylation of E2A (1.7 fold higher). 58 59 60 relative to that resulting from the individual AT enzymes, suggesting p300 and p/CAF may co-operate to achieve maximal acetylation of E2A. 5.3.4 Both p300 and p/CAF Interact Directly with E2A We have previously demonstrated a direct interaction between E2A and CBP/p300 (13). p/CAF was identified due to its ability to interact with either CBP or p300 (123). The p/CAF protein also interacts with E2A (13,143), however, the possibility exists that the E2A-p/CAF interaction could be indirect and mediated by CBP/p300. To investigate this possibility purified, recombinant, full-length FLAG-tagged p/CAF and HIS-tagged p300 were incubated alone or in combination with GST-E2A 1-483 or GST bound to glutathione sepharose beads. Following pull-downs, proteins associated with the beads were analyzed by SDS-PAGE and immunoblotting with antibodies directed against the affinity tags present on p/CAF (FLAG) or p300 (HIS). GST-E2A 1-483, but not GST, was able to pull-down p300 and p/CAF individually (Figure 7, lanes 6 and 7) or in combination (lane 4). These results indicate that p/CAF, like CBP/p300, interacts specifically and directly with E2A. Interestingly, co-incubation of p300 and p/CAF seems to enhance the retention of p300 by E2A suggesting p/CAF may stabilize the interaction between E2A and p300. 61 Figure 7: p/CAF interacts directly with E2A. GST-E2A-1-483 or GST were used in pull-down experiments with purified recombinant p/CAF and p300, alone (lanes 6 and 7) or in combination (lanes 4 and 5). Lanes 1,2, and 3 represent one-tenth input. Following pull-downs, protein complexes bound to glutathione-sepharose were washed and interactive proteins eluted from glutathione-sepharose with excess reduced glutathione. All samples were analysed by SDS-PAGE and immunoblotting with antibodies directed against the epitope tag present on p/CAF (FLAG) and p300 (HIS). 62 63 5.3.5 Acetylation of E2A Promotes its Interaction with CBP Acetylation of another member of the bHLH family, MyoD, results in increased affinity for CBP/p300 (131). Acetylation of the c-MYB transcription factor, which like E2A interacts with the KIX domain of CBP/p300, also increases the strength of its interaction with CBP (139). Therefore the effect of E2A acetylation on its interaction with CBP was tested. GST-E2A 1-483 was acetylated using recombinant GST-p/CAF-HAT prior to its use as a ligand in a pull-down experiment with RCH ACV nuclear extracts. The amount of CBP retained by acetylated versus non-acetylated E2A was determined by SDS-PAGE and an anti-CBP immunoblot. Pre-acetylated E2A was able to pull down more CBP than un-acetylated E2A. An anti-E2A immunoblot confirmed equivalent loading of GST-E2A fusion protein (Figure 8a). This result indicates that acetylation of E2A 1-483 enhances its ability to interact with CBP from nuclear extracts, at least under the assay conditions used here. In order to investigate this result further we examined the ability of GST-E2A 1-483, or an engineered version of GST-E2A in which all 10 lysines within the E2A portion were converted to arginine (lysine-less), to pull-down full-length recombinant p300. The GST proteins were bound to glutathione-sepharose beads prior to the addition of p300 and 14C acetyl-CoA to the reactions to allow acetylation. The amount of p300 retained was analysed by SDS-PAGE and immunoblotting. The results indicate that both wild-type and lysine-less E2A pull-down similar amounts of p300 (Figure 8b) under the conditions of this experiment. 64 Figure 8: Acetylation of E2A promotes its interaction with CBP/p300 from nuclear extracts. (a) Pre-acetylation of E2A by p/CAF enhances its recruitment of CBP/p300 from nuclear extracts (upper image). Acetyl-coenzyme A was omitted from parallel reactions as a negative control. Re-probing the same blot with anti-E2A indicates equivalent loading of the E2A-fusion protein (lower image). (b) E2A acetylation does not enhance its interaction with full-length, recombinant p300. Bacterially expressed and purified wild-type (WT) or lysine-less (LL) GST-E2A 1-483 immobilized on glutathione sepharose beads were incubated with baculovirus-expressed wild-type HIS-tagged p300 under acetyl-transferase assay reaction conditions. Bead-protein complexes were washed and analyzed by SDS-PAGE and anti-p300 immunoblotting. Lane 1 represents one-tenth input. 65 66 This result, using purified p300, is inconsistent with our previous finding showing enhanced ability of acetylated E2A to recruit CBP from nuclear extracts. The discordance between the results in Figures 8a and 8b could reflect the involvement of additional, unidentified nuclear proteins in stabilizing the E2A-CBP/p300 interaction. p/CAF represents a potential candidate as it enhanced retention of p300 by E2A using purified proteins (Figure 7). Alternatively, the ability of lysine-less E2A to recruit equivalent amounts of p300, compared to wildtype E2A, may reflect the supra-physiological amounts of p300 used in the assay. In addition, the lack of any apparent difference between the amount of p300 recruited by acetylatable (i.e. wildtype) verses nonacetylatable (i.e. lysine-less) E2A may also reflect a situation on which prior acetylation of E2A by p/CAF is required to mediate the enhanced interaction between E2A and p300. 5.3.6 Mapping of Acetylated Lysines E2A residues 1-483 contain ten lysines residues. Each of these is phylogenetically conserved and represents a potential substrate for acetylation (Figure 9). The aminoterminal 160 amino acid of E2A includes AD1, contains four lysines (K14, K34, K101, K144), is capable of recruiting CBP and HAT activity from nuclear extracts and is itself, acetylated (Figure 5). A weak consensus site for acetylation of non-histone proteins by CBP/p300 and p/CAF, GKXXP (X denotes any amino acid), has been identified (129,164). Lysine 34 is located within such a consensus (GKGRP) just carboxy-terminal of the putative α-helical region (amino acids 11-28) that is required for interaction with CBP (24). Therefore, K34 represents an attractive candidate for acetylation. Furthermore, 67 Figure 9: Schematic indicating the location and phylogentic conservation of lysine residues in the E2A portion of E2A-PBX1. The sequence alignment of E-proteins shows tight phylogenetic conservation lysine residues and of the GKXXP consensus. Numbers represent amino acid position of lysines in human E2A. 68 69 the location of this residue within AD1 suggests that its acetylation status may affect the transcriptional function of this domain. Site directed mutagenesis was used to replace lysine (K) residues with arginine (R) as arginine mimics an unacetylated, positivelycharged lysine. In vitro HAT reactions using the catalytic domain of p/CAF as the HAT protein and GST-E2A 1-160 fusion proteins as substrates were performed. The resulting acetylation of E2A was quantified by scintillation counting of excised E2A bands. A K34R substitution decreased (24.8% relative to wild-type), but did not abrogate, acetylation relative to wild-type 1-160, indicating that GST-p/CAF-HAT acetylates lysine 34 and at least one other residue within this portion of E2A (Figure 10, lane 2). The three remaining lysine residues (K14, K101, K144) were substituted to arginine in combination with K34R and tested in the acetylation assay. As expected, substitution of all four lysines essentially abrogated detectable acetylation (lane 6). Substitution of lysine 101 (partial consensus, GK) (lane 4) or lysine 144 (lane 5) in combination with K34R also essentially abrogated acetylation, indicating that these residues are also acetylated. In contrast, conversion of lysine 14 to arginine in combination with K34R did not result in decreased acetylation relative to K34R in isolation (33.4% versus 24.8%, lanes 3 and 4). These results suggest that, at least under the conditions of this assay, the catalytic domain of p/CAF acetylates lysines 34, 101, and 144, but not lysine 14, of E2A. 70 Figure 10: The catalytic domain of recombinant p/CAF acetylates lysines 34, 101 and 144 of E2A. In vitro acetylation of E2A 1-160 or lysine to arginine substitution mutants by the recombinant p/CAF catalytic domain. Closed arrows, p/CAF catalytic domain; open arrows, GST-E2A 1-160 proteins. * indicates the positive control p45. 71 72 Additional lysine to arginine substitution mutants were generated to examine acetylation of the carboxy-terminal lysines in E2A 1-483 in the context of K to R substitution of the 4 amino-terminal lysines (i.e., K14R, K34R, K101R and K144R). The isolated catalytic domain of p/CAF could not be used to examine acetylation of the six carboxy-terminal lysines in GST-E2A 1-483 as it co-migrates with GST-E2A and is auto-acetylated. Therefore, the in vitro acetylation assays were performed using full-length, recombinant FLAG-tagged p300 and the reactions subjected to SDS-PAGE and fluorography. The gel was then exposed to a Phosphor screen in order to quantify the level of acetylation of the various mutants relative to that of the wild-type E2A, using a Phosphor Imager. Substitution of the 4 amino-terminal lysine residues reduced acetylation of E2A 1-483 by p300 to 54% of wild-type level, indicating that p300 also acetylates at least a subset of these lysine residues as well as additional, more carboxy-terminal residues. Additional substitution of a cluster of three lysines residues (K171, K172, K175) further reduced acetylation (25% of wild-type E2A), indicating that full-length p300 acetylates at least one of these (Figure 11a, lane 6). A construct containing only K171, K172 and K175 (lane 6) as potential substrates for acetylation was acetylated at levels similar to the construct with substitution of the amino-terminal four lysines only (lane 3) (40% versus 54%, respectively), suggesting that acetylation by p300 of at least one of these residues accounts for most of the acetylation beyond that related to K34, K101 and K144. Lysine 334 falls within a partial GKXXP consensus, GK, and also appears to be acetylated in this assay as mutation of this residue further decreased acetylation of the E2A protein 73 (compare lanes 4 and 5) to levels barely above a lysine-less E2A construct, which served as the negative control (lane 7). The HAT activity recruited by E2A from crude nuclear extracts was examined to determine if it acetylates similar lysine residues as purified, full-length p300. GST-E2A 1-483 proteins were used as ligands in a pull-down experiment using nuclear extracts prepared from RCH ACV cells. The pull-downs were followed by incubation of the complex under acetylation reactions conditions and the resulting acetylation evaluated by SDS-PAGE and fluorography. This experiment (Figure 11b) revealed an identical pattern of acetylated residues compared to the previous experiment using purified recombinant p300 (Figure 11a). Of the six carboxy-terminal lysines examined, the E2Ainteractive HAT complex appears to acetylate at least one of the lysines in the cluster (i.e. K171, K172, K175; lane 6) as well as K334 (compare lanes 4 and 5) but not K204. Due to low amount of lysine-less protein (lane 7) we were unable to determine a relative acetylation value. However, this protein is not acetylated in all previous experiments. A large acetylated band corresponding in size to CBP/p300 consistently appears in all GSTE2A pull downs suggesting CBP/p300 may be the HAT protein responsible for acetylating E2A in this assay. Importantly, CBP/p300 is retained in equivalent quantities by the various lysine substitution mutants, indicating that differential acetylation of the mutant E2A proteins could not be accounted for by effects on the affinity of the interaction with CBP/p300 74 Figure 11: Recombinant p300 and the co-activator complex recruited from nuclear extracts acetylate identical lysine residues within E2A amino acids 161-483. (a) Fulllength p300 was incubated with GST-E2A 1-483 containing various lysine to arginine substitutions in acetyl-transferase reaction buffer in the presence of 14C acetyl-Co A. Reactions were analysed by SDS-PAGE and fluorography. (b) GST-E2A 1-483 substitution mutants were used as ligands in pull-down experiments with nuclear extracts. Immobilized GST-E2A 1-483 and interactive proteins were incubated with 14C acetyl-Co A in acetyl-transferase reaction buffer and analysed by SDS-PAGE and fluorography. 75 76 77 5.3.7 Transcriptional Activation by AD1 Involves Acetylation of K34 Acetylation of transcription factors is known to have a variety of effects, including alteration of transcriptional activation. Several studies have indicated enhanced transcriptional activation activity by E2A on co-transfection with co-activators, including CBP, p300 and p/CAF (11,142,143). In order to investigate whether acetylation of E2A amino acids 1-483 may alter its transcriptional function, plasmids conferring mammalian expression of the lysine substitution mutants, as fusions with a heterologous DNA binding domain derived from the yeast transcription factor GAL4, were constructed. These plasmids were co-transfected into HeLa cells with a reporter plasmid that confers expression of firefly luciferase regulated by five tandem GAL4 DNA-binding sites. All results were normalized for transfection efficiency using a co-transfected plasmid that confers constitutive expression of renilla luciferase. In the graph, the results are expressed relative to that obtained using wild-type GAL4-E2A 1-483 and represent an average of three independent experiments. Substitution of lysine 34 (K34R) significantly reduced transcriptional activation relative to that observed using wild-type E2A (74.7%) (Figure 12). Substitution of K14, K101, K144 or K144 to arginine in combination with K34R had little additional effect on transcriptional activation by E2A compared to K34R alone, (69.9%, 68.1% and 72% respectively versus 74.7%) Substitution of the amino-terminal three (K14R, K34R, K101R) lysines only slightly decreased E2A transcriptional activation compared to the double mutants. However, additional substitution of K144 (i.e. K14R, K34R, K101R, K144R) further decreased transactivation to 54.7% of wild-type suggesting post-translational modification of K34, K101 and K144 may be involved in regulating E2A activity. 78 The role of the more carboxy-terminal 6 lysine residues was investigated in the context of lysine to arginine mutation of the four amino-terminal lysines (K14R, K34R, K101R, K144R). An E2A protein with additional substitution of lysines 171, 172 and 175 further reduced the transcriptional activation compared to the quadruple mutant (43.2% versus 54.7%, respectively,) suggesting that at least one of these lysines plays a role in E2A function. Additional substitution of lysine 334 in combination with K171R, K172R and K175R did not significantly effect E2A transcriptional activation compared to K171R, K172R and K175R (44.7% versus 43.2%, respectively) suggesting that modifications at K334 do not contribute to transcriptional activation by E2A. An E2A protein containing only lysines 171,172 and 175 as candidate substrates for modification was not significantly different from the quadruple mutant (52% versus 54.7%) providing additional evidence that lysine modification of at least one of these residues, and not K204, K334 or K392, may play a role in modulating E2A function. Several observations implicate acetylation, as opposed to other lysine modification, of K34 in the transcriptional function of AD1. Arginine substitution at this site substantially impaired transcriptional activation. In addition, this residue is found within a GKXXP consensus motif for acetylation of non-histone proteins and is located adjacent to a helical region of E2A that we have shown to interact directly with the KIX domain CBP (13). We now demonstrate that this residue is a substrate for acetylation. Additional experiments were undertaken to show that acetylation, and not another lysine modification of K34 is the relevant modification. 79 Figure 12: Post-translational modification of lysine 34 is involved in transcriptional activation by E2A. (a) HeLa cells were co-transfected with wild-type or lysine to arginine substitution mutants of E2A fused to the DNA-binding domain of GAL4 along with a reporter plasmid that confers expression of firefly luciferase regulated by tandem GAL4 binding sites. The role of lysine residues in E2A-mediated transcriptional activation was examined using lysine substitution mutants. The plasmid pRL-CMV, which confers constitutive expression of renilla luciferase was also co-transfected and was used as a measure of transfection efficiency. Transfections were performed in replicates of four and the results represent the average from at least three independent experiments. (b) Anti-E2A immunoblot of whole cell lysates from a typical experiment shows expression of all E2A mutants. 80 81 5.3.8 Role for Acetylation of Lysine 34 in the Transcriptional Function of Activation Domain 1 Lysine residues are also substrates for several post-translational modifications besides acetylation, including ubiquitination and sumoylation. Whereas the side chain of arginine is positively charged at neutral pH, mimicking an acetylated lysine, the side chain of glutamine is uncharged, mimicking acetylated lysine. Therefore lysine 34 was substituted by site directed mutagenesis, to glutamine (K34Q) in the context of GAL4-E2A 1-483 and tested for transcriptional activation activity. The K34Q mutant reconstituted E2Amediated transcriptional activation to at least levels associated with the wild-type protein (Figure 12) further supporting a role for acetylation of K34 in the transcriptional function of AD1. The E2A transcription factors E12 and E47 possess a relatively short half-life in vivo and have been shown to be poly-ubiquitinated downstream of Notch-1 signaling (64,66). This results in accelerated degradation through the 26S proteasome. E12 and E47 are also known to interact with the SUMO conjugating enzyme UBC-9 through a domain that spans the fusion point with PBX1 in E2A fusion proteins (63-65). Ubiquitination and sumoylation of the oncogenic portion of E2A (i.e. amino acids 1-483) has not been examined. Additional experiments were performed to examine whether E2A 1-483 may serve as a substrate for ubiquitination or sumoylation. Plasmids conferring constitutive expression of HIS-tagged E2A 1-483 or c-JUN (positive control) were transiently co-transfected into 82 SV293T cells with a plasmid conferring expression of hemaglutinin (HA) tagged ubiquitin. The HIS-tagged proteins were affinity purified from whole cell lysates on nickel NTA agarose and eluted proteins were evaluated by SDS-PAGE and immunoblotting with an anti-HA antibody to detect ubiquitinated E2A or c-JUN. Since substrates may be poly- or mono-ubiquitinated at multiple lysine residues, a positive reaction will result in ubiquitinated proteins of increasing size, producing a smear on the anti-HA immunoblot. Although c-JUN was ubiquitinated as expected, E2A 1-483 protein was not ubiquitinated in this assay, suggesting that this portion of E2A is not a substrate for ubiquitination (Figure 13a). Similar experiments were performed to examine sumoylation of E2A 1-483. HIS-tagged JUN, wild-type and lysine-less E2A 1-483 were co-transfected with an expression vector for FLAG tagged-SUMO. Whole cell lysates were prepared and HIS-tagged proteins purified and analysed as described for ubiquitin experiments. An anti-FLAG immunoblot shows only the c-JUN positive control to be sumoylated; neither the wild-type E2A 1-483 nor the lysine-less (LL) negative control were sumoylated (Figure 13b). Together, these results suggest that acetylation and not ubiquitination or sumoylation of lysine residues within E2A 1-483 is the key post-translational modification involved in regulating E2A-mediated transcriptional activation activity. 83 Figure 13: The oncogenic portion of E2A is not a substrate for ubiquitination or sumoylation. (a) SV293T cell were transiently transfected with wild-type, HIS-tagged E2A 1-483, a lysine-less (LL) E2A mutant or HIS-tagged c-JUN (positive control), with or without HA-tagged ubiquitin. HIS-tagged proteins were affinity purified from whole cell lysates using nickel NTA agarose and subjected to SDS-PAGE and immunoblotting. Top panel, anti-HA, detects ubiquitinated proteins; bottom panel, anti-E2A. (b) Transfections and protein purification were performed as described for (a) except FLAGtagged SUMO was co-transfected in place of HA-ubiquitin. Top panel, anti-FLAG, detects sumoylated proteins; bottom panel, anti-E2A. Large white circle in the c-Jun + FLAG-SUMO lane is a result of excessive signal consuming the luminol reagent. The lysine-less E2A mutant serves as a useful negative control in this experiment. 84 85 86 5.4 Discussion Over the past several years the list of acetylated transcription factors has grown considerably. Initially thought to be restricted to histones and involved in modulation of chromatin conformation, protein acetylation is now know to also modulate a variety of functions associated with transcription factors, including: DNA binding, sub-cellular localization, protein-protein interactions and transcriptional activation (15). Recently, the LeBrun lab has described a direct interaction between AD1 of E2A and the KIX domain of CBP (13,148). The E2A-CBP interaction is required for E2A-PBX mediated growth deregulation in tissue culture models (13) and leukemogenesis in vivo (14). This chapter describes an investigation of one of the downstream consequences of the interaction between E2A and the co-activators CBP, p300 and p/CAF, namely, acetylation of E2A itself. In the work described here, I have used pull-downs to demonstrate the portion of E2A found in the oncogenic fusion protein E2A-PBX1, i.e. E2A amino acids 1-483, recruits histone acetyltransferase activity able to acetylate histones and E2A itself from the human ALL cell line RCH ACV. Furthermore, recruitment of HAT activity correlates with recruitment of the co-activators CBP/p300 and p/CAF, suggesting that one of these proteins may be the HAT protein responsible for acetylation of E2A. In fact, all three coactivators efficiently acetylate E2A 1-483. 87 The result showing a direct interaction between E2A and p/CAF is novel and raises the possibility that p/CAF may modulate the function of E2A activity. Interestingly, using purified proteins, p/CAF was able to enhance the amount of p300 retained by E2A. In addition, p300 and p/CAF seem to co-operate to achieve maximal E2A acetylation. Together, these results suggest p/CAF may be functioning to stabilize the interaction between E2A and p300 resulting in greater acetylation of E2A. These results also point to p/CAF as a potential candidate for the unknown nuclear factor that enhances the ability of acetylated E2A to retain CBP from nuclear extracts. Enhanced recruitment of CBP/p300 by acetylated E2A could have several functional implications. For instance, the enhanced localization of CBP/p300 to promoters regulated by acetylated E2A or E2A-PBX1 genes would likely result in increased acetylation of nucleosomal histones and other chromatin-associated proteins. Acetylation also creates docking sites for bromodomain-containing proteins resulting in recruitment of additional factors required for transcription. In the case of E2A-PBX1, this would result in aberrant expression of PBX1 defined target genes involved in leukemogenesis. In a DNA-binding independent manner, the enhanced recruitment of CBP/p300 could result in a dominantnegative effect on other transcription factors that require CBP/p300 for proper function. This study maps, for the first time, several acetylated lysines within the oncogenic portion of E2A, (i.e. amino acids 1-483) using recombinant CBP/p300 and p/CAF as well as the HAT complex recruited from nuclear extracts. Three amino-terminal lysines (K34, K101 and K144), in addition to the more carboxy-terminal K334 and at least one of the 88 lysines in a cluster (i.e. K171, K172 and K175) are substrates for acetylation. Interestingly, acetylation of E2A by CBP/p300 required more that an isolated catalytic domain. Discrepancies have been noted between the ability of the isolated catalytic domain of CBP/p300, relative to the full-length protein, to mediate acetylation of substrates. MyoD is a bHLH family member that is involved in promoting muscle cell terminal differentiation and is acetylated by p/CAF (137). Polesskaya et al demonstrated that full-length CBP/p300 was capable of acetylating MyoD both in vivo and in vitro (136). A later study, however, showed that the isolated catalytic domain of CBP/p300 was unable to acetylate MyoD (165). MyoD has been shown to interact with the C/H3 domain of CBP/p300 (166) and, like E2A, interacts with the KIX domain of CBP/p300 (167), these domains are truncated or absent when using an isolated catalytic domain. Polesskaya et al demonstrated that acetylation of MyoD, by p300, requires a direct interaction between the proteins (138). Therefore, the lack of E2A acetylation by the isolated catalytic domain of CBP is likely due to the loss of key protein-protein interaction domains. Bradney et al have described acetylation of the E2A splice variant E2-5 in vitro and in vivo by CBP/p300 and p/CAF (143). E2-5 lacks amino acids 1-99 (i.e. AD1) but contains the E47 bHLH domains. In this study, Bradney et al showed that E2-5 co-elutes from nuclear extracts with CBP/p300 and p/CAF in high molecular weight complexes. However, since E2-5 lacks AD1 and the LXXLLDFS motif required for interaction with the KIX domain of CBP (13,14,148) it is reasonable to speculate that the E2A-CBP interactions seen in this study must be mediated by other E2A domains or additional 89 proteins. The bHLH domain, for example has been shown to interact with CBP/p300 (11) or perhaps the α-helical region of AD2 (13). Consistent with previous studies (11,142) Bradney et al demonstrate that transcriptional activation activity by E2-5 is increased by co-transfection of CBP/p300 or p/CAF(143). However, this study did not examine the mechanism of enhanced E2A-mediated transcriptional activation. Since E2-5 lacks amino acids 1-99 of E2A it does not contain lysine 34. The location of K34 within AD1 suggested to us that post-translational modifications at this site may play a role in regulating transcriptional activation by E2A proteins. Mimicking an unacetylated state, by arginine substitution, substantially decreased E2A mediated transcription of a reporter gene while substitution to glutamine, mimicking the uncharged, acetylated state restores and perhaps enhances E2A activity supporting the role of acetylation in mediating E2A transcriptional activation. Furthermore, Mr. Bayly, in the LeBrun lab, has demonstrated in vivo acetylation of K34 using co-immunoprecipitations. An anti-acetyl-lysine antibody immunoprecipitated far less E2A 1-483 with K34R or K34Q substitutions than wild-type E2A 1-483, from cells co-transfected with CBP and E2A 1-483 expression vectors (unpublished results). This result indicates that lysine 34 is a substrate for acetylation by CBP/p300 or p/CAF in cells and confirms the in vitro result showing acetylation at this residue by the catalytic domain of p/CAF. In an attempt to rule out the involvement of other lysine post-translation modifications in mediating E2A transcriptional activation I examined potential ubiquitination and 90 sumoylation of E2A 1-483. However, I was unable to demonstrate ubiquitination or sumoylation of E2A 1-483 in spite of the fact that E12 and E47 proteins have been demonstrated to be ubiquitinated (66). My inability to demonstrate ubiquitination of E2A 1-483 could reflect the fact that most of the bipartite binding site for UBC9, required for proteasomal degradation of E2A, is missing (63,64). Interestingly, the two chimeric E2A oncoproteins, E2A-PBX1 and E2A-HLF, exclude the exon that encodes this degradation domain (65). Alternatively, Notch-1 signals required for E2A phosphorylation and subsequent interaction with the E3 ubiquitin ligase SCFSkp2 (66,69) were not induced in the cell culture system used here. Acetylation at the lysine cluster (K171, K172, K175) accounts for the majority of acetylation amongst the carboxy-terminal six lysine residues. Interestingly, lysine to arginine substitution of these residues also significantly decreased E2A-mediated transcriptional activation, suggesting a role for acetylation at K171, 172 and K175 in E2A driven transcription. The amino acid sequence at this lysine cluster (KKVRK) represents a potential nuclear localization site (NLS) (loose consensus, K(K/R)X(K/R), where X equals any amino acid) (168), suggesting that acetylation of these residues could alter the sub-cellular localization of E2A proteins. Recently, acetylation of the Max transcription factor at three lysine residues within its nuclear localization signal (NLS) has been demonstrated and seems to result in localization of the protein to the cytoplasm. Compartmentalization of Max to the cytoplasm inhibits its ability to suppress c-Myc activated transcription (169). Therefore, 91 the increased nuclear retention of E2A seen by Bradney et al, when co-transfected with CBP/p300 or p/CAF (143), might be explained by acetylation of lysines within this NLS. In the same manner, enhanced concentration of E2A in the nucleus might be partially responsible for increased transcriptional activation by E2A when co-transfected with CBP/p300 or p/CAF (143). Several examples exist in the literature of enhanced transcriptional activation by a transcription factor following acetylation. However, the mechanism of enhancement seems to involve increased affinity of DNA (129,131,139,140). In the reporter assay used in the current study, DNA-binding of E2A 1-483 is mediated by the GAL4 DNA-binding domain and not endogenous bHLH or the PBX1-derived homeodomain, suggesting another mechanism for regulation E2A transcriptional activation. In the case of the muscle specific bHLH protein MyoD, the increased transcriptional activation by acetylated MyoD has been suggested to be a result of the enhanced affinity of CBP/p300, due to an acetylation-dependent interaction with the bromodomain of CBP/p300 (131). It would be interesting to investigate the possibility of acetylated E2A interacting with the bromodomains of CBP/p300 and p/CAF. The carboxy-terminus of E2A (i.e. the portion not included in E2A-PBX1, containing the bHLH domain) contains 12 (E47) or 13 (E12) additional lysine residues. The portion of PBX1 included in E2A-PBX1 contains 15 lysines. In both cases several of the lysines are located within domains that mediate dimerization with other proteins or the respective DNA-binding domains. Post-translational modification of these lysine residues could 92 potentially have affects on DNA binding of normal or oncogenic E2A proteins. In addition, protein-protein interactions mediated by these regions could be affected by acetylation. Therefore, investigation of the acetylation status of these lysines should be pursued in the future to examine the functional consequences of post-translational modification, specifically acetylation, on E2A function. In summary, the discovery that the oncogenic portion of E2A (i.e. amino acids 1-483) can mediate direct interactions with the transcriptional co-activators CBP/p300 (13) and p/CAF (Figure 7, this thesis) resulting in acetylation of E2A itself is novel. E2A is acetylated at multiple sites and acetylation appears to play a role in regulating E2Amediated transcriptional activation activity. Acetylation of E2A results in enhanced retention of CBP/p300, in a manner that requires additional proteins, possibly p/CAF. One might envision a model in which E2A recruitment of CBP/p300 and p/CAF results in acetylation of E2A itself, stabilization of the complex followed by recruitment of additional co-activators, ATP-dependent remodeling complexes and the transcriptional machinery culminating in enhanced transcriptional activation of genes required for tissue specific differentiation (normal E2A proteins) or oncogenesis (E2A-PBX1). 93 Chapter 6 E2A Enhances the Acetyl-transferase Activity of CBP 6.1 Contributions Preparation of small oligonucleosomes (SONs) from HeLa cells was performed by Mr. Richard Bayly. Initial stocks of purified, recombinant, full-length p300 were a generous gift of Dr. Jeffrey Dilworth. Additional protein stocks were prepared by Mr. Bayly by infection of SF21 cells with p300 baculovirus and subsequent purification of the recombinant protein. 6.2 Rationale The E2A-CBP interaction is critical for E2A-PBX1-mediated leukemogenesis (14). In addition, acetylation of E2A at several conserved lysines, including one located within AD1, plays a role in transcriptional activation activity by E2A. An important consequence of transcription factor-mediated recruitment of co-activators, such as CBP/p300 and p/CAF, to gene promoters is acetylation of lysine residues located within the amino-terminal tails of nucleosomal histones, affecting the interaction between histones and DNA. Acetylated histones are generally found at transcriptionally active loci. Twist is a bHLH protein implicated in inhibition of differentiation of multiple cell lineages (170,170-176). Twist interacts directly with the HAT domains of p300 and p/CAF in vitro and in vivo (173,177) and completely abrogates acetylation of histones as well as auto-acetylation of the co-activators (177). Inhibition of HAT activity by Twist 94 does not require its bHLH domain suggesting dimerization with other bHLH proteins is not required for inhibition transactivation (177). Since E2A is also a bHLH transcription factor that interacts with co-activators we decided to investigate the possibility that the interaction between E2A and CBP may affect the catalytic activity of CBP. 6.3 Results 6.3.1 E2A Proteins Enhance in vitro Acetylation of Histones by CBP/p300 In order to investigate the possibility that interaction with E2A proteins are capable of modulating the catalytic activity of CBP/p300, amino acids 1-483 of E2A were expressed as an in-frame fusion with glutathione-S-transferase (GST) and purified on glutathione sepharose beads. GST E2A 1-483, or GST alone, was incubated with full-length p300 in the presence of free histones and 14C-acetyl co-enzyme A. Full-length, recombinant p300 was expressed by baculovirus infection of SF21 insect cells and affinity purification using nickel NTA agarose. Samples were subjected to SDS-PAGE and the resulting gel was Coomassie-stained and photographed prior to fluorography. After several weeks on film the gel was transferred to a Phosphor screen and acetylation levels of histones quantified using a Phosphor Imager. Values were normalized according to the quantity of histone protein present in each lane, as determined by densitometry of the Coomassie stained gel. GST-E2A 1-483, but not unfused GST, was capable of enhancing the acetylation of histones by approximately two-fold (Figure 14a) by p300. The enhancement of HAT 95 activity did not require acetylation of E2A, as a lysine-less E2A mutant was also capable of enhancing p300 HAT activity (2.7 fold increase), note the absence of a band corresponding to acetylated E2A in this lane. A faster method of examining the effects of E2A proteins on HAT activity that does not require lengthy exposure to film, would allow multiple replicates to be performed and that would more readily allow kinetic parameters to be investigated was sought out. A filter-binding HAT assay has been described. This entails incubating a HAT enzyme, histones as a substrate, and 14C-acetyl-CoA in reaction buffer for a period of time prior to stopping the reaction by spotting onto phosphocellulose filters (159-161). These are then washed to remove unincorporated label and the labeled (i.e., acetylated) proteins are quantified by liquid scintillation counting of the filter. In order to examine the effect of E2A on the rate of CBP-mediated acetylation of histones, a time course experiment was performed. Full-length, FLAG-tagged CBP (CBP-WT) was immuno-affinity purified from whole cell lysates of transiently transfected SV293T cells on M2 anti-FLAG agarose beads. GST-E2A 1-483 was incubated with CBP-WT in a 10:1 molar ratio (0.2 pmol/μl E2A, 0.02 pmol/μl CBP) for 10 minutes prior to addition of histones and 14C-acetyl-CoA. The results presented in Figure 15 represents the mean of two independent experiments. The linear portion of the graph was determined using linear regression analysis and the slope of the line represents reaction rate. Incubation of CBP with histones yielded a reaction rate of 2.3X10-3μM/s. Addition of E2A to the reaction mixture increased the rate to 5.3X10-3μM/s. Therefore, 96 the addition of E2A results in a 2.3-fold increase in the reaction rate, a value that is similar to that found using full-length p300 in the gel-based assay (Figure 14a). Extension of the lines to the axis reveals that they do not intersect zero on the graph. This is likely due to the fast rate of histone acetylation by CBP and technical limitations of the filter assay itself, at very short time points. A reaction time of 5 minutes was used in all subsequent filter assays. Together these results demonstrate for the first time that E2A is capable of enhancing the intrinsic HAT activity of CBP/p300. 97 Figure 14: E2A enhances the histone acetyltransferase activity of p300. (a) HAT assays were performed using full-length, HIS-tagged p300 produced by baculovirus infection of SF21 cells and affinity purification. Acetylation levels were quantified and normalized against the quantity of histone protein in each lane. WT, wild-type; lysineless, an engineered E2A mutant with arginine substitutions at all 10 E2A lysine residues; L20A, leucine to alanine substitution at E2A residue 20; Δ397-405, deletion of an αhelical region within E2A activation domain 2. (b) Increasing the amount of E2A from 5pmol to 15pmol does not further enhance p300 HAT activity. 98 99 100 Figure 15: Time course experiment showing that E2A enhances the catalytic activity of CBP approximately two-fold. CBP was incubated in reaction buffer in the presence of 14C-acetyl-co enzyme A and histones for different time periods. At the completion of the reaction, the solution was spotted onto a phosphocellulose filter which were then washed to remove unincorportated label. Acetylated proteins bound to the filter were quantified by scintillation counting. The graph shows the concentration of acetylated histones (μM) versus time (seconds) by full-length, recombinant CBP in the absence (dark circles) or presence (light circles) of a 10-fold molar excess of E2A 1-483. The addition of E2A to the reaction increases the reaction rate from 2.3X10-3 μM/sec to 5.3X10-3 μM/sec, a 2.3 fold increase. The experiment was performed twice and error bars indicate standard deviation. 101 102 6.3.2 Mapping of E2A Domains Required to Increase the HAT Activity of CBP We next set out to identify the domains required, in both proteins, for E2A-mediated enhancement of CBP/p300 histone acetylation. The LeBrun lab has previously identified a direct interaction between an α-helical region within AD1 of E2A and the KIX domain of CBP (13,148). Deletion or mutation of key residues within AD1 dramatically impairs the CBP (i.e. KIX) interaction. An α-helical region within AD2 also contributes to the interaction and is required for maximal transcriptional activation by E2A (13). The role of these α-helical domains in enhancement of CBP HAT activity was investigated. The filter-based HAT assay was performed using full-length GST-E2A 1-483 or GSTE2A 1-483 proteins with targeted deletions of the α-helical regions within AD1 and AD2. The results shown in Figure 16 are displayed relative to those obtained using fulllength CBP alone. Targeted deletion of the AD1 α-helix (Δ16-23) did not abrogate the ability of E2A to enhance CBP HAT activity (1.4 fold versus 1.9 fold). Similarly, deletion of the AD2 α-helical region (Δ 397-405) had little effect (1.5 fold versus 1.9 fold). Deletion of both α-helical regions did not significantly alter E2A-induced enhancement of HAT activity (1.7 fold versus 1.9 fold). These results suggest that protein-protein interactions mediated by the α-helical domains are not required for the effect. Further supporting the idea that the E2A-KIX interaction is not required for enhancement of CBP/p300 HAT activity a gel based assay shows similar results. Deletion or mutation 103 of the α-helical regions of E2A had no effect on the ability of E2A to enhance p300 HAT activity (Figure 14). Together, these results suggest that the α-helical regions and by implication the KIX interaction, is dispensable for the E2A-medaited stimulation of CBP HAT activity. 104 Figure 16: The α-helical domains within AD1 and AD2 of E2A are dispensable for enhancement of CBP histone acetylation. HAT assays were performed using fulllength, recombinant CBP and GST-E2A 1-483 proteins. Δ16-23 and Δ397-405, deletions of an α-helical regions within E2A activation domains 1 and 2. Error bars indicate standard deviation calculated from the results of 5 filters counted for each reaction. 105 106 6.3.3 The KIX- and Bromo-domains are Dispensable for E2A-induced Increase of CBP HAT activity We next wanted to investigate the requirement of the well-characterized KIX interaction in mediating the E2A-dependent enhancement of CBP catalytic activity. Full-length, FLAG-tagged CBP proteins or mutant versions with deletion of the entire KIX domain (amino acids 588-667, CBP-ΔKIX), most of the BROMO domain (Δ1107-1171, CBPΔBROMO) or a small region within the HAT domain (Δ1458-1475, CBP-ΔHAT) were expressed in SV293T cells by transient transfection and immuno-affinity purified using M2 anti-FLAG agarose beads for use in the HAT assay. The results shown in Figure 17a are expressed relative to that obtained using CBP-WT without E2A. As previously demonstrated (Figures 14, 15 and 16), addition of GST-E2A 1-483 increases the HAT activity of CBP (1.9 fold). The CBP-ΔBROMO protein has somewhat lower HAT activity compared to CBP-WT. However, the addition of E2A to the reaction results in increased HAT activity compared to CBP-ΔBROMO alone (1.6 fold), a response similar to that obtained using wild-type CBP. Deletion of the KIX domain has no effect on either the ability of CBP to acetylate histones in the absence of E2A, or the ability of E2A to enhance HAT activity (1.7 fold versus CBP-ΔKIX alone). As expected, the CBP-ΔHAT mutant has markedly reduced HAT in the absence of E2A that is not augmented on addition of E2A to the reaction. Together, these results indicate that the E2A-KIX interaction and the bromodomain are dispensable in the enhancement of CBP HAT activity. 107 The isolated catalytic domain of CBP (amino acids 1196-1718) fused in frame to GST was used to further investigate the possibility that an interaction between E2A and the HAT domain may be required to mediated enhanced CBP acetylation of histones. This region contains the C/H2, HAT and most of the C/H3 domains of CBP. Co-incubation of GST-CBP-HAT with E2A in the filter assay resulted in only a very small increase in histone acetylation compared to GST-CBP-HAT alone (1.2 +/- 0.08 versus 1.0 +/- 0.05) (Figure 17b). This result suggests that E2A interacts with a domain outside the isolated HAT domain to mediate enhancement of HAT. Previous mapping studies that identified the direct interaction between AD1 of E2A and the KIX domain of CBP used small CBP domains as individual GST-fusion proteins (13). Therefore, it remains possible that E2A could interact with additional CBP domains in the context of full-length CBP. Pull-down experiments using GST-E2A 1-483 and fulllength, FLAG-tagged CBP proteins, expressed by transient transfection and immunoaffinity purification were performed to investigate this possibility. Retained proteins were analysed by SDS-PAGE and anti-FLAG immunoblots. As expected, E2A was able to recruit significant amounts of wild-type, full length CBP (Figure 17c). Deletion of the entire KIX domain (amino acids 588-667) abrogated the interaction with E2A. Deletion of most of the BROMO domain (amino acids 1107-1171) or a small region of the HAT domain (amino acids 1458-1475) reduced but did not abrogate the interaction with E2A 1-483. These experiments confirm that the KIX domain is required to mediate a direct interaction with E2A in the context of full-length CBP. The results also suggest that the 108 BROMO and HAT domains of CBP may play a role in mediating the interaction with E2A. It is interesting that despite the inability of CBP-ΔKIX to interact with E2A in pull-down assays it is still able to respond to E2A in the filter assay. It is possible that a lower affinity interaction between E2A and CBP-ΔKIX is responsible for mediating the interaction (and the enhanced HAT activity) in the filter assay in the absence of the KIX domain. Reciprocally, the residual recruitment of CBP-ΔBROMO and CBP-ΔHAT in the pull-downs is likely mediated by the KIX domain. The ability of CBP-ΔBROMO to respond to E2A in the HAT filter assay reflects the ability of this protein to interact with E2A through the KIX or other CBP domains. 109 Figure 17: The KIX and BROMO domains of CBP are dispensable for enhancement of HAT activity by E2A. (a) HAT filter assays were performed using full-length, recombinant CBP or CBP mutants containing deletion of the entire KIX domain (ΔKIX), most of the bromodomain (ΔBROMO) or a small segment of the HAT domain (ΔHAT). Error bars indicate standard deviation calculated from the results of 5 filters counted for each reaction. (b) E2A only slightly enhances the HAT activity of an isolated CBP catalytic domain. HAT assays were performed using the isolated catalytic domain of CBP (GST-CBP-HAT) and GST-E2A 1-483. Error bars indicate standard deviation calculated from the results of 5 filters counted for each reaction. (c) The BROMO and HAT domains contribute to the interaction with E2A. GST-E2A-1-483 was used in pulldown experiments with purified recombinant full-length wild-type or deletion mutants of CBP. Following the pull downs, protein complexes bound to glutathione-sepharose beads were analysed by SDS-PAGE and immunoblotting with antibodies directed against the affinity tag present on CBP (FLAG). An anti-E2A immunoblot was performed as a loading control. 110 111 112 6.3.4 E2A does not enhance CBP Acetylation of Nucleosomes in vitro The results demonstrating enhanced histone acetylation by CBP/p300 when incubated with E2A were obtained using free histones purified from calf thymus. These are not associated with DNA, therefore the lysine substrates are not in their normal cellular configuration. Small oligonucleosomes (SONs) purified from HeLa cell nuclear pellets represent a more biologically relevant substrate. Histone H1 depleted SONs were prepared by Mr. Richard Bayly following the method described by Côté et al (178). Briefly, large amounts of HeLa cells are grown and the nuclei are purified. Nuclear pellets are lysed and the chromatin digested with micrococcal nuclease. Following digestion, the sample is applied to a sepharose column and fractions collected. The fractions containing short oligonucleosomes (four core histones only, determined by absorbance at 260nM and SDS-PAGE) are pooled and quantified against known amounts of free calf thymus histones on a Coomassie stained gel. HAT assays were performed using full-length CBP, GST-E2A 1-483 and SONs. An equivalent amount of SON derived histones, compared to free histones used in previous experiments (0.2μg/μl) were used for the HAT assays. Acetylation of the SONs by CBP was approximately 3 to 4 fold lower compared to acetylation of free histone by the same amount of enzyme. Increasing the reaction time from 5 minutes to 30 minutes had no effect on acetylation rates (not shown). Addition of E2A to the reactions did not enhance the low level of CBP SON acetylation and the increased reaction time resulted in greater E2A acetylation that accounts for any apparent increase in SON acetylation (Figure 18). 113 Figure 18: E2A is unable to enhance CBP-mediated acetylation of nucleosomes in vitro. HAT assays were performed using full-length, recombinant CBP and small oligonucleosomes purified from HeLa cell nuclei. An equivalent amount of nucleosomal histones was used as compared to free histones used in previous experiments. The HAT reaction time was increased form 5 minutes to 30 minutes. A control reaction containing CBP and E2A but lacking nucleosomes was performed and resulted in significant E2A acetylation. This value was subtracted from the reaction containing CBP, E2A and nucleosomes, to account for the contribution from acetylated E2A. Error bars indicate standard deviation calculated from the results of 5 filters counted for each reaction. 114 115 The low SON acetylation rate could be accounted for by the fact that the histones are structured in chromatin and may therefore be less accessible than free histone for acetylation. Since E2A proteins must bind to chromatinized DNA in order to promote gene expression, enhancement of CBP HAT activity by E2A may require a stable interaction with chromatin or DNA. We sought out an in vivo assay to address this possibility. 6.3.5 E2A Enhances CBP Catalytic Activity in vivo Martinez-Balbas et al have established that the intrinsic acetyl-transferase activity of CBP can stimulate transcription of a reporter gene when tethered to a promoter (163). In this system, a segment of CBP including the HAT domain (amino acids 1099-1877) is expressed as an in frame fusion with the DNA binding domain of GAL4 (Figure 19a) in cells co-transfected with a reporter plasmid consisting of five GAL4 binding sites upstream of the adenoviral major-late (AdML) promoter and the chloramphenicol acetyltransferase (CAT) coding region (Figure 19a). A GAL4-CBP fusion (GAL4CBPHAT-CBP2) but not a catalytically dead mutant is able to activate expression of the CAT reporter gene in the osteosarcoma cell line, U2-OS (163). Evidence exists to show that transfected plasmid DNA does get rapidly chromatinized (179-181). This system will allow us to investigate if E2A is able to enhance CBP-mediated histone acetylation on a chromatinized template in vivo. Plasmids conferring expression of GAL4-CBP and various E2A proteins were cotransfected into U2-OS cells with the AdML-CAT reporter. All values were corrected for 116 transfection efficiency using a co-transfected plasmid that confers constitutive expression of renilla luciferase. The level of CAT expression was assayed using the FAST-CAT deoxy-Green chloramphenicol kit (Molecular Probes). The chloramphenicol substrate in this kit has been modified to incorporate a green fluorophor and contains only one potential acetylation site. Therefore the rate of acetylation is a direct function of enzymatic activity. Whole cell lysates were incubated with the labeled chloramphenicol and the reactions products resolved by thin layer chromatography (TLC) to separate acetylated from unmodified substrates (Figure 19b). The resulting plates were excited with 470 nm +/20 nm light, emission at 520 nm +/- 20nm was imaged and fluorescent spots quantified using Image Pro Plus software. The results presented in Figure 19 are typical of those obtained in two independent experiments and are expressed relative to GAL4-CBPHATCBP2 alone. Co-transfection of GAL4-CBPHAT-CBP2 with E2A-PBX1 dramatically increased CAT expression (12.6 fold) (Figure 19c). Full length E2A (E47) was also able to enhance CBP-HAT dependent transcriptional activation (8.9 fold) as was E2A 1-483 (10.1 fold, data not shown) while neither PBX1a nor PBX1b was able to stimulate CBPHAT dependent transcriptional activation. Therefore, these results indicate the E2A portion of the oncogenic E2A-PBX1 protein is responsible for mediating enhanced CBP activity in vivo. In addition, the results demonstrate that E2A is able to enhance CBP acetylation of chromatin in vivo. The results also confirm that the E2A-KIX interaction is not required for E2A-mediated enhancement of CBP catalytic activity as this domain is not present in the GAL4-CBP fusions. 117 Figure 19: The E2A portion of E2A-PBX1 mediates enhanced acetyltransferasedependent transcriptional activation by CBP in vivo. (a) Schematic representation of CBP and the GAL4-CBP Fusions used in the CAT reporter Assay. Modified from reference 162. (b) E2A enhances CBP HAT activity in vivo. E2A and PBX1 proteins were co-transfected with the HAT domain of CBP fused the GAL4 DNA, in U2OS cells, along with a CAT reporter plasmid. The plasmid pRL-CMV, which confers constitutive expression of renilla luciferase was also co-transfected and was used as a measure of transfection efficiency. CAT assays were performed on whole cell lysates as described for the Fast CAT Green deoxy-Chloramphenicol Acetyltransferase kit (Molecular Probes). Fluorescent TLC plates were imaged and (c) quantified using Image ProPlus software. All TLC values were corrected for transfection efficiency. Transfections were performed in duplicate and error bars represent standard deviations. The values obtained using GAL4-CBPHATΔ-CBP2 without or with co-transfected E2A, E2A-PBX1 or PBX1 are not significantly above zero. Error bars for all GAL4-CBPHATΔ-CBP2 transfections overlap and cross zero suggesting that these are not significantly different from each other. 118 119 120 6.4 Discussion Transcriptional co-activators are intimately involved in regulation of chromatin structure, assembly and gene transcription. The co-activators CBP, p300 and p/CAF possess intrinsic acetyltransferase activity responsible for acetylation of lysine residues in core histone proteins (114,123,125) and acetylation of histones is highly correlated with gene expression (92,93). Since co-activators lack DNA binding elements, sequence specific DNA binding transcription factors must first recruit them to gene promoters prior to acetylation of chromatin (87,89). We demonstrate here that E47 and the portion of E2A found within the oncogenic fusion protein E2A-PBX1 (i.e. E2A 1-483) are able to enhance the histone acetyltransferase activity of CBP/p300 using both in vitro and in vivo assays. This finding is novel and represents a potential mechanism for regulation of target gene expression for normal and oncogenic E2A proteins. While the ability of E2A to enhance CBP/p300 catalytic activity is novel it is not unprecedented for a transcription factor to modulate HAT activity of coactivators. Several other transcription factors that interact with CBP/p300 and p/CAF have also been shown to inhibit or enhance catalytic activity. For example, The Early Bcell Factor (EBF) collaborates with E47 in regulating early B-cell development (182184). Unlike MyoD and E2A proteins, EBF does not require the HAT activity of CBP/p300 to stimulate transcription of its target genes (185). However, EBF does bind to CBP/p300 within the HAT domain and inhibit acetylation (185) suggesting it may function to sequester CBP/p300 from other transcription factors. 121 The Epstein Barr Virus (EBV) immediate early protein Zta is a member of the basic zipper (b-zip) family that interacts with CBP at its C/H3 domain. The interaction with CBP enhances transcriptional activation by Zta and promotes reactivation of latent EBV (186). Reciprocally, Zta stimulates HAT activity of CBP in a manner that requires the transactivation and DNA binding domains of Zta. Two additional b-zip family members, NF-E2 and C/EBPα, also stimulate CBP HAT activity of nucleosomal histones suggesting, these b-zip proteins stimulate HAT activity by directing CBP to nucleosomes (187,188). The examples above suggest that transcription factor mediated enhancement or inhibition of HAT activity requires a direct interaction with the HAT protein. A direct interaction between AD1 of E2A and the KIX domain of CBP has been described (13,148). However, I demonstrate here that the E2A-KIX interaction is dispensable for E2Ainduced stimulation of HAT activity both in vitro and in vivo. I provide evidence that the BROMO and HAT domains of CBP contribute to the interaction with E2A. However, the ability of E2A to enhance the catalytic HAT activity of CBP in vitro requires a domain outside the region containing intrinsic HAT activity (Figure 17b). The results from the in vivo CAT assays point to the BROMO, CH3 or TAZ domains as candidates for further investigation. Although the mechanism by which E2A and other transcription factors influence the HAT activity has not been fully elucidated, it is unlikely that E2A enhances the catalytic activity of CBP/p300 by an E2A-histone effect since E2A-mediated stimulation of 122 histone acetylation could not be further augmented by a three-fold increase in the amount of E2A added to the HAT reaction (Figure 14b). The E2A-mediated enhancement of CBP HAT activity may be due to induction of conformational changes that alter enzymatic activity or possibly affect interaction and/or accessibility of histones or other proteins. I also demonstrate that acetylation of E2A itself is not required for enhancement of p300 HAT activity as a lysine-less E2A was equally capable of mediating the effect. This result suggests that acetylation of E2A is important for mediating other effects, perhaps recruitment of additional factors through acetyl-lysine/bromodomain interactions. In summary, the results presented in this chapter suggest a model of E2A-mediated transcriptional regulation in which wild-type and oncogenic E2A-fusion proteins enhance expression of target genes by altering the catalytic HAT activity of the co-activators CBP/p300 to induce gene expression. 123 Chapter 7 General Discussion and Future Directions The co-activator proteins CBP and p300 are ubiquitously expressed and are intimately involved in regulating gene expression from many promoters (189,190). While the majority of the experiments described in this thesis used CBP, full-length p300 was used as the HAT enzyme in a few experiments. These proteins are highly homologous and are often used interchangeably in in vitro assays (113,115) however, in vivo they likely act differently. In mouse models, homozygous knockout of either CBP or p300 is embryonic lethal. The homozygous knockouts display similar phenotypes and defects suggesting nonoverlapping functions of the two proteins (191-193). Alternatively, the total level of CBP and p300 may be critical for proper development. In support of this model, mice expressing only one copy of each of the CBP and p300 genes (double heterozygotes) are also embryonic lethal (191). In humans, haplo-insufficiency of CBP and less frequently p300, results in Rubenstein-Taybi syndrome (RTS), a congenital disease characterized by mental retardation and congenital malformations in addition to a propensity for malignancies (194). In fact loss of HAT activity may be sufficient to cause RTS (195). Both p300 and CBP gene loci are involved in non-random chromosomal translocations associated with acute myeloid leukemia (AML) resulting in expression of chimeric oncoproteins. CBP, for example, is fused to MOZ (monocyte leukemia zinc finger protein). 124 MOZ is, itself, a HAT protein. MLL (mixed lineage leukemia protein) is a methyltransferase (MT) protein and is involved in translocations with CBP and less frequently, p300. In all cases, the portion of CBP/p300 retained in the fusion proteins includes the bromodomain, HAT domain and carboxy terminal residues (reviewed in (189)). The nature of the proteins involved in these translocations suggests mis-targeting of the HAT and MT activities may play a role in leukemogenesis (196). The E2A locus is involved in at least two non-random chromosomal translocations associated with acute lymphoblastic leukemia. In both cases the translocation results in expression of chimeric oncoproteins that contain the transcriptional activation domains for E2A fused in frame to DNA binding elements derived from the translocation partner. Importantly, deletion of transcriptional activation domain one (AD1) from E2A-PBX1 abrogates growth deregulation and neoplastic transformation (83,84,141) suggesting protein-protein interactions mediated by AD1 are important for E2A-PBX1-mediated leukemogenesis (84). Recently Dr. LeBrun’s lab demonstrated a direct interaction between a conserved motif in an α-helical region of AD1 of E2A and the KIX domain of the transcriptional coactivator CBP. Mutations that disrupt the E2A-CBP interaction (13,148) also abrogate E2A-PBX1 mediated induction of proliferation in primary hematopoietic cells (13) and leukemogenesis in lethally irradiated mice transplanted with primary bone marrow transduced to express E2A-PBX1 (14). These results suggest the E2A-CBP interaction is important for the function of normal and oncogenic E2A proteins. However, the 125 mechanism by which the interaction functions to regulate gene expression remains unclear. The experiments described in this thesis were undertaken to investigate the functional consequences, for both proteins, of the physical interaction between E2A and CBP. The experimental findings discussed in this thesis provide significant insight into the mechanisms involved in regulating the transcriptional activity of wild-type and oncogenic E2A proteins. Primarily, several acetylated lysine residues within E2A have been identified and determined to play a role in regulation the transcriptional activation of gene expression by E2A. Acetylation of E2A also appears to enhance the interaction between E2A and the co-activator CBP. The demonstration of a direct interaction between E2A and a second co-activator, p/CAF, is novel and provides evidence that more than one co-activator is involved in regulating E2A function. In particular, p/CAF appears to be the protein responsible for mediating the enhanced interaction between CBP/p300 and E2A, in addition to co-operating with p300 to maximally acetylate E2A. Together these results underscore the need to further characterize the interaction between E2A and p/CAF, at structural and functional levels. Perhaps the most interesting finding of this thesis is the identification of a novel mechanism of E2A-mediated regulation of transcription, namely enhancement of the intrinsic HAT activity of CBP/p300 both in vitro and in vivo. Surprisingly, the effect is independent of the E2A-KIX interaction previously described in our lab (13,148) and 126 suggests E2A interacts with CBP at additional domains. My results suggest several candidates including the BROMO, C/H3 or TAZ domains of CBP as potential interactive partners for E2A in mediating increased HAT activity. The finding that E2A-mediated enhancement of CBP HAT activity is independent of E2A acetylation is interesting. We have recently demonstrated that retroviral transduction on primary bone marrow cells with E2A-PBX1 containing arginine substitutions in all 10 of the E2A-derived lysine residues had no effect on in vitro immortalization, proliferation or extended clonogenic potential, compared to normal E2A-PBX1. The lethal myeloproliferative disease developed by irradiated mice transplantation with the transduced cells was only slightly less aggressive with lysine-substituted E2A-PBX1 (LeBrun Lab, unpublished results). Since acetylation of E2A-derived lysines is not required for E2A-PBX1-mediated oncogenesis, and E2A-PBX1 is capable of enhancing HAT activity in the in vivo system, this newly identified activity of E2A proteins, i.e. stimulation of CBP/p300 HAT activity, potentially plays a role in oncogenesis, by misdirecting CBP HAT activity to promoters of genes required for leukemogenesis. Alternatively, E2A-PBX1 can induce proliferation of cells in vitro (83,84,141) and induce lymphoid tumours in mice (141) independent of DNA binding. Both these functions require AD1, and by implication, interaction with CBP, suggesting E2A might exert gain of function effects on CBP that could be responsible for some leukemogenic properties. 127 Several lines of investigation remain to fully elucidate the downstream consequences of the interaction between E2A and co-activators. These include but are not limited to: 1) Are lysines located within the PBX1 portion of E2A-PBX1 or the carboxyterminus of E12 or E47 acetylated? 2) What role does acetylation of these residues play in mediating the function of oncogenic E2A proteins? Acetylation of lysine residues within domains required for DNA binding or dimerization domains could potentially affect protein-protein interactions and DNA binding affinity. 3) Does the direct interaction between E2A and p/CAF also result in enhanced HAT activity? While I demonstrated a direct interaction between E2A and p/CAF, the domains required to mediate the direct interaction have not been identified and the affect of the interaction of p/CAF activity was not examined. 4) What is the order of events at endogenous promoters? MyoD interacts with and is acetylated by both p/CAF and p300 (11,131,136-138). Using an in vitro transcription system it has recently been described that transactivation at a muscle specific promoter requires a specific sequence of events. MyoD/E47 heterodimers recruit p300 to the promoter resulting in acetylation of histones 3 and 4. Histone acetylation recruits p/CAF to the promoter where it acetylates MyoD (197). A similar system could be used to 128 investigate the sequence of events and roles of CBP/p300 and p/CAF in E2A-dependent transcriptional activation at endogenous naked or nucleosomal promoters. 5) Is an ATP-dependent chromatin remodeling complex recruited as a result of the E2A-CBP interaction? Acetylation of lysine residues within histone tails and nonhistone proteins generates recognition sites for bromodomains found in remodeling complexes. Identification of additional proteins required for mediating E2A transcriptional activation would provide additional insight into the mechanism oncogenesis by E2A-PBX1. In summary, the results presented in this thesis suggest a model of E2A-mediated transcriptional regulation in which wild-type and oncogenic E2A-fusion proteins enhance the local concentration of CBP to promoter elements, resulting in acetylation of E2A itself and stabilization of the E2A-CBP interaction. This interaction results in enhancement of CBP-HAT activity, likely due to an allosteric change, acetylation of histones and recruitment of the transcriptional machinery to induce target gene expression. Enhancement of CBP/p300 HAT activity by oncogenic E2A-PBX1 proteins in vivo, suggests that some of its leukemia-promoting effects may be due to E2A-induced gain of function effects on CBP/p300 and might play a role in the DNA-bindingindependent induction of proliferation. 129 References References 1. Canadian Cancer Society/ National Cancer Institute of Canada. Canadian Cancer Statistics 2007. 2007. Toronto, Canada. Ref Type: Report 2. Look, A. T. 1997. Oncogenic transcription factors in the human acute leukemias. Science 278:1059-1064. 3. Nourse, J., J. D. Mellentin, N. Galili, J. Wilkinson, E. Stanbridge, S. D. Smith, and M. L. Cleary. 1990. Chromosomal translocation t(1;19) results in synthesis of a homeobox fusion mRNA that codes for a potential chimeric transcription factor. Cell 60:535-545. 4. Kamps, M. P., C. Murre, X. H. Sun, and D. Baltimore. 1990. A new homeobox gene contributes the DNA binding domain of the t(1;19) translocation protein in pre-B ALL. Cell 60:547-555. 5. Mellentin, J. D., C. Murre, T. A. Donlon, P. S. McCaw, S. D. Smith, A. J. Carroll, M. E. McDonald, D. Baltimore, and M. L. Cleary. 1989. The gene for enhancer binding proteins E12/E47 lies at the t(1;19) breakpoint in acute leukemias. Science 246:379-382. 6. Hunger, S. P. 1996. Chromosomal translocations involving the E2A gene in acute lymphoblastic leukemia: clinical features and molecular pathogenesis. Blood 87:1211-1224. 7. Hunger, S. P., K. Ohyashiki, K. Toyama, and M. L. Cleary. 1992. Hlf, a novel hepatic bZIP protein, shows altered DNA-binding properties following fusion to E2A in t(17;19) acute lymphoblastic leukemia. Genes Dev. 6:16081620. 8. Inaba, T., W. M. Roberts, L. H. Shapiro, K. W. Jolly, S. C. Raimondi, S. D. Smith, and A. T. Look. 1992. Fusion of the leucine zipper gene HLF to the E2A gene in human acute B-lineage leukemia. Science 257:531-534. 9. Brambillasca, F., G. Mosna, M. Colombo, A. Rivolta, C. Caslini, M. Minuzzo, G. Giudici, L. Mizzi, A. Biondi, and E. Privitera. 1999. Identification of a novel molecular partner of the E2A gene in childhood leukemia. Leukemia 13:369-375. 10. Massari, M. E., P. A. Jennings, and C. Murre. 1996. The AD1 transactivation domain of E2A contains a highly conserved helix which is required for its activity in both Saccharomyces cerevisiae and mammalian cells. Mol.Cell Biol. 16:121-129. 130 11. Eckner, R., T. P. Yao, E. Oldread, and D. M. Livingston. 1996. Interaction and functional collaboration of p300/CBP and bHLH proteins in muscle and Bcell differentiation. Genes Dev. 10:2478-2490. 12. Massari, M. E., P. A. Grant, M. G. Pray-Grant, S. L. Berger, J. L. Workman, and C. Murre. 1999. A conserved motif present in a class of helixloop-helix proteins activates transcription by direct recruitment of the SAGA complex. Mol.Cell 4:63-73. 13. Bayly, R., L. Chuen, R. A. Currie, B. D. Hyndman, R. Casselman, G. A. Blobel, and D. P. LeBrun. 2004. E2A-PBX1 interacts directly with the KIX domain of CBP/p300 in the induction of proliferation in primary hematopoietic cells. J.Biol.Chem. 279:55362-55371. 14. Bayly, R., T. Murase, B. D. Hyndman, R. Savage, S. Nurmohamed, K. Munro, R. Casselman, S. P. Smith, and D. P. LeBrun. 2006. Critical role for a single leucine residue in leukemia induction by E2A-PBX1. Mol.Cell Biol. 26:6442-6452. 15. Glozak, M. A., N. Sengupta, X. Zhang, and E. Seto. 2005. Acetylation and deacetylation of non-histone proteins. Gene 363:15-23. 16. Murre, C., P. S. McCaw, and D. Baltimore. 1989. A new DNA binding and dimerization motif in immunoglobulin enhancer binding, daughterless, MyoD, and myc proteins. Cell 56:777-783. 17. Voronova, A. and D. Baltimore. 1990. Mutations that disrupt DNA binding and dimer formation in the E47 helix-loop-helix protein map to distinct domains. Proc.Natl.Acad.Sci.U.S A 87:4722-4726. 18. Church, G. M., A. Ephrussi, W. Gilbert, and S. Tonegawa. 1985. Cell-typespecific contacts to immunoglobulin enhancers in nuclei. Nature 313:798-801. 19. Ephrussi, A., G. M. Church, S. Tonegawa, and W. Gilbert. 1985. B lineage-specific interactions of an immunoglobulin enhancer with cellular factors in vivo. Science 227:134-140. 20. Lenardo, M., J. W. Pierce, and D. Baltimore. 1987. Protein-binding sites in Ig gene enhancers determine transcriptional activity and inducibility. Science 236:1573-1577. 21. Davis, R. L., H. Weintraub, and A. B. Lassar. 1987. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell 51:987-1000. 22. Tapscott, S. J., R. L. Davis, M. J. Thayer, P. F. Cheng, H. Weintraub, and A. B. Lassar. 1988. MyoD1: a nuclear phosphoprotein requiring a Myc homology region to convert fibroblasts to myoblasts. Science 242:405-411. 131 23. Wright, W. E., D. A. Sassoon, and V. K. Lin. 1989. Myogenin, a factor regulating myogenesis, has a domain homologous to MyoD. Cell 56:607-617. 24. Murre, C., P. S. McCaw, H. Vaessin, M. Caudy, L. Y. Jan, Y. N. Jan, C. V. Cabrera, J. N. Buskin, S. D. Hauschka, A. B. Lassar, and . 1989. Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a common DNA sequence. Cell 58:537-544. 25. Henthorn, P., R. McCarrick-Walmsley, and T. Kadesch. 1990. Sequence of the cDNA encoding ITF-1, a positive-acting transcription factor. Nucleic Acids Res. 18:677. 26. Aronheim, A., R. Shiran, A. Rosen, and M. D. Walker. 1993. The E2A gene product contains two separable and functionally distinct transcription activation domains. Proc.Natl.Acad.Sci.U.S A 90:8063-8067. 27. Quong, M. W., M. E. Massari, R. Zwart, and C. Murre. 1993. A new transcriptional-activation motif restricted to a class of helix-loop-helix proteins is functionally conserved in both yeast and mammalian cells. Mol.Cell Biol. 13:792-800. 28. McInerney, E. M., D. W. Rose, S. E. Flynn, S. Westin, T. M. Mullen, A. Krones, J. Inostroza, J. Torchia, R. T. Nolte, N. Assa-Munt, M. V. Milburn, C. K. Glass, and M. G. Rosenfeld. 1998. Determinants of coactivator LXXLL motif specificity in nuclear receptor transcriptional activation. Genes Dev. 12:3357-3368. 29. Ko, L., G. R. Cardona, and W. W. Chin. 2000. Thyroid hormone receptorbinding protein, an LXXLL motif-containing protein, functions as a general coactivator. Proc.Natl.Acad.Sci.U.S A 97:6212-6217. 30. Kee, B. L. and C. Murre. 1998. Induction of early B cell factor (EBF) and multiple B lineage genes by the basic helix-loop-helix transcription factor E12. J.Exp.Med. 188:699-713. 31. Schlissel, M., A. Voronova, and D. Baltimore. 1991. Helix-loop-helix transcription factor E47 activates germ-line immunoglobulin heavy-chain gene transcription and rearrangement in a pre-T-cell line. Genes Dev. 5:1367-1376. 32. Zhuang, Y., P. Soriano, and H. Weintraub. 1994. The helix-loop-helix gene E2A is required for B cell formation. Cell 79:875-884. 33. Bain, G., E. C. Maandag, D. J. Izon, D. Amsen, A. M. Kruisbeek, B. C. Weintraub, I. Krop, M. S. Schlissel, A. J. Feeney, M. van Roon, and . 1994. E2A proteins are required for proper B cell development and initiation of immunoglobulin gene rearrangements. Cell 79:885-892. 132 34. Bain, G., E. C. Robanus Maandag, H. P. te Riele, A. J. Feeney, A. Sheehy, M. Schlissel, S. A. Shinton, R. R. Hardy, and C. Murre. 1997. Both E12 and E47 allow commitment to the B cell lineage. Immunity. 6:145-154. 35. Xin, X. Q., C. Nelson, L. Collins, and K. Dorshkind. 1993. Kinetics of E2A basic helix-loop-helix-protein expression during myelopoiesis and primary B cell differentiation. J.Immunol. 151:5398-5407. 36. Kee, B. L., M. W. Quong, and C. Murre. 2000. E2A proteins: essential regulators at multiple stages of B-cell development. Immunol.Rev. 175:138149. 37. Roberts, V. J., R. Steenbergen, and C. Murre. 1993. Localization of E2A mRNA expression in developing and adult rat tissues. Proc.Natl.Acad.Sci.U.S A 90:7583-7587. 38. Goldfarb, A. N., J. P. Flores, and K. Lewandowska. 1996. Involvement of the E2A basic helix-loop-helix protein in immunoglobulin heavy chain class switching. Mol.Immunol. 33:947-956. 39. Rutherford, M. N. and D. P. LeBrun. 1998. Restricted expression of E2A protein in primary human tissues correlates with proliferation and differentiation. Am.J.Pathol. 153:165-173. 40. Quong, M. W., D. P. Harris, S. L. Swain, and C. Murre. 1999. E2A activity is induced during B-cell activation to promote immunoglobulin class switch recombination. EMBO J. 18:6307-6318. 41. Aronheim, A., R. Shiran, A. Rosen, and M. D. Walker. 1993. Cell-specific expression of helix-loop-helix transcription factors encoded by the E2A gene. Nucleic Acids Res. 21:1601-1606. 42. Peverali, F. A., T. Ramqvist, R. Saffrich, R. Pepperkok, M. V. Barone, and L. Philipson. 1994. Regulation of G1 progression by E2A and Id helix-loophelix proteins. EMBO J. 13:4291-4301. 43. Loveys, D. A., M. B. Streiff, and G. J. Kato. 1996. E2A basic-helix-loop-helix transcription factors are negatively regulated by serum growth factors and by the Id3 protein. Nucleic Acids Res. 24:2813-2820. 44. Benezra, R. 1994. An intermolecular disulfide bond stabilizes E2A homodimers and is required for DNA binding at physiological temperatures. Cell 79:1057-1067. 45. Markus, M. and R. Benezra. 1999. Two isoforms of protein disulfide isomerase alter the dimerization status of E2A proteins by a redox mechanism. J.Biol.Chem. 274:1040-1049. 133 46. Sun, X. H. and D. Baltimore. 1991. An inhibitory domain of E12 transcription factor prevents DNA binding in E12 homodimers but not in E12 heterodimers. Cell 64:459-470. 47. Benezra, R., R. L. Davis, D. Lockshon, D. L. Turner, and H. Weintraub. 1990. The protein Id: a negative regulator of helix-loop-helix DNA binding proteins. Cell 61:49-59. 48. Sun, X. H., N. G. Copeland, N. A. Jenkins, and D. Baltimore. 1991. Id proteins Id1 and Id2 selectively inhibit DNA binding by one class of helix-loophelix proteins. Mol.Cell Biol. 11:5603-5611. 49. Wilson, R. B., M. Kiledjian, C. P. Shen, R. Benezra, P. Zwollo, S. M. Dymecki, S. V. Desiderio, and T. Kadesch. 1991. Repression of immunoglobulin enhancers by the helix-loop-helix protein Id: implications for B-lymphoid-cell development. Mol.Cell Biol. 11:6185-6191. 50. Whitmarsh, A. J. and R. J. Davis. 2000. Regulation of transcription factor function by phosphorylation. Cell Mol.Life Sci. 57:1172-1183. 51. Shen, C. P. and T. Kadesch. 1995. B-cell-specific DNA binding by an E47 homodimer. Mol.Cell Biol. 15:4518-4524. 52. Sloan, S. R., C. P. Shen, R. McCarrick-Walmsley, and T. Kadesch. 1996. Phosphorylation of E47 as a potential determinant of B-cell-specific activity. Mol.Cell Biol. 16:6900-6908. 53. Neufeld, B., A. Grosse-Wilde, A. Hoffmeyer, B. W. Jordan, P. Chen, D. Dinev, S. Ludwig, and U. R. Rapp. 2000. Serine/Threonine kinases 3pK and MAPK-activated protein kinase 2 interact with the basic helix-loop-helix transcription factor E47 and repress its transcriptional activity. J.Biol.Chem. 275:20239-20242. 54. Page, J. L., X. Wang, L. M. Sordillo, and S. E. Johnson. 2004. MEKK1 signaling through p38 leads to transcriptional inactivation of E47 and repression of skeletal myogenesis. J.Biol.Chem. 279:30966-30972. 55. Chu, C. and D. S. Kohtz. 2001. Identification of the E2A gene products as regulatory targets of the G1 cyclin-dependent kinases. J.Biol.Chem. 276:85248534. 56. Glickman, M. H. and A. Ciechanover. 2002. The ubiquitin-proteasome proteolytic pathway: destruction for the sake of construction. Physiol Rev. 82:373-428. 57. Rechsteiner, M. 1990. PEST sequences are signals for rapid intracellular proteolysis. Semin.Cell Biol. 1:433-440. 134 58. Johnson, E. S. and G. Blobel. 1997. Ubc9p is the conjugating enzyme for the ubiquitin-like protein Smt3p. J.Biol.Chem. 272:26799-26802. 59. Johnson, E. S. and G. Blobel. 1999. Cell cycle-regulated attachment of the ubiquitin-related protein SUMO to the yeast septins. J.Cell Biol. 147:981-994. 60. Gong, L., T. Kamitani, K. Fujise, L. S. Caskey, and E. T. Yeh. 1997. Preferential interaction of sentrin with a ubiquitin-conjugating enzyme, Ubc9. J.Biol.Chem. 272:28198-28201. 61. Desterro, J. M., J. Thomson, and R. T. Hay. 1997. Ubch9 conjugates SUMO but not ubiquitin. FEBS Lett. 417:297-300. 62. Schwarz, S. E., K. Matuschewski, D. Liakopoulos, M. Scheffner, and S. Jentsch. 1998. The ubiquitin-like proteins SMT3 and SUMO-1 are conjugated by the UBC9 E2 enzyme. Proc.Natl.Acad.Sci.U.S A 95:560-564. 63. Loveys, D. A., M. B. Streiff, T. S. Schaefer, and G. J. Kato. 1997. The mUBC9 murine ubiquitin conjugating enzyme interacts with the E2A transcription factors. Gene 201:169-177. 64. Kho, C. J., G. S. Huggins, W. O. Endege, C. M. Hsieh, M. E. Lee, and E. Haber. 1997. Degradation of E2A proteins through a ubiquitin-conjugating enzyme, UbcE2A. J.Biol.Chem. 272:3845-3851. 65. Huggins, G. S., M. T. Chin, N. E. Sibinga, S. L. Lee, E. Haber, and M. E. Lee. 1999. Characterization of the mUBC9-binding sites required for E2A protein degradation. J.Biol.Chem. 274:28690-28696. 66. Nie, L., M. Xu, A. Vladimirova, and X. H. Sun. 2003. Notch-induced E2A ubiquitination and degradation are controlled by MAP kinase activities. EMBO J. 22:5780-5792. 67. Ordentlich, P., A. Lin, C. P. Shen, C. Blaumueller, K. Matsuno, S. Artavanis-Tsakonas, and T. Kadesch. 1998. Notch inhibition of E47 supports the existence of a novel signaling pathway. Mol.Cell Biol. 18:2230-2239. 68. King, A. M., P. E. Van der, B. B. Blomberg, and R. L. Riley. 2007. Accelerated Notch-dependent degradation of E47 proteins in aged B cell precursors is associated with increased ERK MAPK activation. J.Immunol. 178:3521-3529. 69. Huang, Z., L. Nie, M. Xu, and X. H. Sun. 2004. Notch-induced E2A degradation requires CHIP and Hsc70 as novel facilitators of ubiquitination. Mol.Cell Biol. 24:8951-8962. 135 70. Carroll, A. J., W. M. Crist, R. T. Parmley, M. Roper, M. D. Cooper, and W. H. Finley. 1984. Pre-B cell leukemia associated with chromosome translocation 1;19. Blood 63:721-724. 71. Williams, D. L., A. T. Look, S. L. Melvin, P. K. Roberson, G. Dahl, T. Flake, and S. Stass. 1984. New chromosomal translocations correlate with specific immunophenotypes of childhood acute lymphoblastic leukemia. Cell 36:101-109. 72. Michael, P. M., M. D. Levin, and O. M. Garson. 1984. Translocation 1;19--a new cytogenetic abnormality in acute lymphocytic leukemia. Cancer Genet.Cytogenet. 12:333-341. 73. Burglin, T. R. 1997. Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domain conserved between plants and animals. Nucleic Acids Res. 25:4173-4180. 74. Chang, C. P., W. F. Shen, S. Rozenfeld, H. J. Lawrence, C. Largman, and M. L. Cleary. 1995. Pbx proteins display hexapeptide-dependent cooperative DNA binding with a subset of Hox proteins. Genes Dev. 9:663-674. 75. Knoepfler, P. S. and M. P. Kamps. 1995. The pentapeptide motif of Hox proteins is required for cooperative DNA binding with Pbx1, physically contacts Pbx1, and enhances DNA binding by Pbx1. Mol.Cell Biol. 15:5811-5819. 76. Neuteboom, S. T., L. T. Peltenburg, M. A. van Dijk, and C. Murre. 1995. The hexapeptide LFPWMR in Hoxb-8 is required for cooperative DNA binding with Pbx1 and Pbx2 proteins. Proc.Natl.Acad.Sci.U.S A 92:9166-9170. 77. Piper, D. E., A. H. Batchelor, C. P. Chang, M. L. Cleary, and C. Wolberger. 1999. Structure of a HoxB1-Pbx1 heterodimer bound to DNA: role of the hexapeptide and a fourth homeodomain helix in complex formation. Cell 96:587-597. 78. Selleri, L., M. J. Depew, Y. Jacobs, S. K. Chanda, K. Y. Tsang, K. S. Cheah, J. L. Rubenstein, S. O'Gorman, and M. L. Cleary. 2001. Requirement for Pbx1 in skeletal patterning and programming chondrocyte proliferation and differentiation. Development 128:3543-3557. 79. Monica, K., N. Galili, J. Nourse, D. Saltman, and M. L. Cleary. 1991. PBX2 and PBX3, new homeobox genes with extensive homology to the human protooncogene PBX1. Mol.Cell Biol. 11:6149-6157. 80. van Dijk, M. A., P. M. Voorhoeve, and C. Murre. 1993. Pbx1 is converted into a transcriptional activator upon acquiring the N-terminal region of E2A in pre-B-cell acute lymphoblastoid leukemia. Proc.Natl.Acad.Sci.U.S A 90:60616065. 136 81. Lu, Q., D. D. Wright, and M. P. Kamps. 1994. Fusion with E2A converts the Pbx1 homeodomain protein into a constitutive transcriptional activator in human leukemias carrying the t(1;19) translocation. Mol.Cell Biol. 14:39383948. 82. LeBrun, D. P. and M. L. Cleary. 1994. Fusion with E2A alters the transcriptional properties of the homeodomain protein PBX1 in t(1;19) leukemias. Oncogene 9:1641-1647. 83. Kamps, M. P., D. D. Wright, and Q. Lu. 1996. DNA-binding by oncoprotein E2a-Pbx1 is important for blocking differentiation but dispensable for fibroblast transformation. Oncogene 12:19-30. 84. Bayly, R. and D. P. LeBrun. 2000. Role for homodimerization in growth deregulation by E2a fusion proteins. Mol.Cell Biol. 20:5789-5796. 85. LeBrun, D. P. 2003. E2A basic helix-loop-helix transcription factors in human leukemia. Front Biosci. 8:s206-s222. 86. Luger, K., A. W. Mader, R. K. Richmond, D. F. Sargent, and T. J. Richmond. 1997. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389:251-260. 87. Lemon, B. and R. Tjian. 2000. Orchestrated response: a symphony of transcription factors for gene control. Genes Dev. 14:2551-2569. 88. Kadonaga, J. T. 2004. Regulation of RNA polymerase II transcription by sequence-specific DNA binding factors. Cell 116:247-257. 89. Heintzman, N. D. and B. Ren. 2007. The gateway to transcription: identifying, characterizing and understanding promoters in the eukaryotic genome. Cell Mol.Life Sci. 64:386-400. 90. Naar, A. M., B. D. Lemon, and R. Tjian. 2001. Transcriptional coactivator complexes. Annu.Rev.Biochem. 70:475-501. 91. Strahl, B. D. and C. D. Allis. 2000. The language of covalent histone modifications. Nature 403:41-45. 92. Schubeler, D., D. M. MacAlpine, D. Scalzo, C. Wirbelauer, C. Kooperberg, F. van Leeuwen, D. E. Gottschling, L. P. O'Neill, B. M. Turner, J. Delrow, S. P. Bell, and M. Groudine. 2004. The histone modification pattern of active genes revealed through genome-wide chromatin analysis of a higher eukaryote. Genes Dev. 18:1263-1271. 93. Roh, T. Y., S. Cuddapah, and K. Zhao. 2005. Active chromatin domains are defined by acetylation islands revealed by genome-wide mapping. Genes Dev. 19:542-552. 137 94. Roth, S. Y., J. M. Denu, and C. D. Allis. 2001. Histone acetyltransferases. Annu.Rev.Biochem. 70:81-120. 95. Herrera, J. E., K. Sakaguchi, M. Bergel, L. Trieschmann, Y. Nakatani, and M. Bustin. 1999. Specific acetylation of chromosomal protein HMG-17 by PCAF alters its interaction with nucleosomes. Mol.Cell Biol. 19:3466-3473. 96. Bergel, M., J. E. Herrera, B. J. Thatcher, M. Prymakowska-Bosak, A. Vassilev, Y. Nakatani, B. Martin, and M. Bustin. 2000. Acetylation of novel sites in the nucleosomal binding domain of chromosomal protein HMG-14 by p300 alters its interaction with nucleosomes. J.Biol.Chem. 275:11514-11520. 97. Mehnert, J. M. and W. K. Kelly. 2007. Histone deacetylase inhibitors: biology and mechanism of action. Cancer J. 13:23-29. 98. Spotswood, H. T. and B. M. Turner. 2002. An increasingly complex code. J.Clin.Invest 110:577-582. 99. Kouzarides, T. 2007. Chromatin modifications and their function. Cell 128:693-705. 100. Havas, K., I. Whitehouse, and T. Owen-Hughes. 2001. ATP-dependent chromatin remodeling activities. Cell Mol.Life Sci. 58:673-682. 101. Peterson, C. L. and I. Herskowitz. 1992. Characterization of the yeast SWI1, SWI2, and SWI3 genes, which encode a global activator of transcription. Cell 68:573-583. 102. Hogan, C. and P. Varga-Weisz. 2007. The regulation of ATP-dependent nucleosome remodelling factors. Mutat.Res. 618:41-51. 103. Zhang, W., S. Kadam, B. M. Emerson, and J. J. Bieker. 2001. Site-specific acetylation by p300 or CREB binding protein regulates erythroid Kruppel-like factor transcriptional activity via its interaction with the SWI-SNF complex. Mol.Cell Biol. 21:2413-2422. 104. Deuring, R., L. Fanti, J. A. Armstrong, M. Sarte, O. Papoulas, M. Prestel, G. Daubresse, M. Verardo, S. L. Moseley, M. Berloco, T. Tsukiyama, C. Wu, S. Pimpinelli, and J. W. Tamkun. 2000. The ISWI chromatin-remodeling protein is required for gene expression and the maintenance of higher order chromatin structure in vivo. Mol.Cell 5:355-365. 105. Wysocka, J., T. Swigut, H. Xiao, T. A. Milne, S. Y. Kwon, J. Landry, M. Kauer, A. J. Tackett, B. T. Chait, P. Badenhorst, C. Wu, and C. D. Allis. 2006. A PHD finger of NURF couples histone H3 lysine 4 trimethylation with chromatin remodelling. Nature 442:86-90. 138 106. Li, H., S. Ilin, W. Wang, E. M. Duncan, J. Wysocka, C. D. Allis, and D. J. Patel. 2006. Molecular basis for site-specific read-out of histone H3K4me3 by the BPTF PHD finger of NURF. Nature 442:91-95. 107. Schneider, R., A. J. Bannister, F. A. Myers, A. W. Thorne, C. CraneRobinson, and T. Kouzarides. 2004. Histone H3 lysine 4 methylation patterns in higher eukaryotic genes. Nat.Cell Biol. 6:73-77. 108. Chrivia, J. C., R. P. Kwok, N. Lamb, M. Hagiwara, M. R. Montminy, and R. H. Goodman. 1993. Phosphorylated CREB binds specifically to the nuclear protein CBP. Nature 365:855-859. 109. Kwok, R. P., J. R. Lundblad, J. C. Chrivia, J. P. Richards, H. P. Bachinger, R. G. Brennan, S. G. Roberts, M. R. Green, and R. H. Goodman. 1994. Nuclear protein CBP is a coactivator for the transcription factor CREB. Nature 370:223-226. 110. Arias, J., A. S. Alberts, P. Brindle, F. X. Claret, T. Smeal, M. Karin, J. Feramisco, and M. Montminy. 1994. Activation of cAMP and mitogen responsive genes relies on a common nuclear factor. Nature 370:226-229. 111. Harlow, E., P. Whyte, B. R. Franza, Jr., and C. Schley. 1986. Association of adenovirus early-region 1A proteins with cellular polypeptides. Mol.Cell Biol. 6:1579-1589. 112. Eckner, R., M. E. Ewen, D. Newsome, M. Gerdes, J. A. DeCaprio, J. B. Lawrence, and D. M. Livingston. 1994. Molecular cloning and functional analysis of the adenovirus E1A-associated 300-kD protein (p300) reveals a protein with properties of a transcriptional adaptor. Genes Dev. 8:869-884. 113. Arany, Z., W. R. Sellers, D. M. Livingston, and R. Eckner. 1994. E1Aassociated p300 and CREB-associated CBP belong to a conserved family of coactivators. Cell 77:799-800. 114. Eckner, R., Z. Arany, M. Ewen, W. Sellers, and D. M. Livingston. 1994. The adenovirus E1A-associated 300-kD protein exhibits properties of a transcriptional coactivator and belongs to an evolutionarily conserved family. Cold Spring Harb.Symp.Quant.Biol. 59:85-95. 115. Lundblad, J. R., R. P. Kwok, M. E. Laurance, M. L. Harter, and R. H. Goodman. 1995. Adenoviral E1A-associated protein p300 as a functional homologue of the transcriptional co-activator CBP. Nature 374:85-88. 116. Arany, Z., D. Newsome, E. Oldread, D. M. Livingston, and R. Eckner. 1995. A family of transcriptional adaptor proteins targeted by the E1A oncoprotein. Nature 374:81-84. 139 117. Yuan, W., G. Condorelli, M. Caruso, A. Felsani, and A. Giordano. 1996. Human p300 protein is a coactivator for the transcription factor MyoD. J.Biol.Chem. 271:9009-9013. 118. Abraham, S. E., S. Lobo, P. Yaciuk, H. G. Wang, and E. Moran. 1993. p300, and p300-associated proteins, are components of TATA-binding protein (TBP) complexes. Oncogene 8:1639-1647. 119. Nakajima, T., C. Uchida, S. F. Anderson, J. D. Parvin, and M. Montminy. 1997. Analysis of a cAMP-responsive activator reveals a two-component mechanism for transcriptional induction via signal-dependent factors. Genes Dev. 11:738-747. 120. Nakajima, T., C. Uchida, S. F. Anderson, C. G. Lee, J. Hurwitz, J. D. Parvin, and M. Montminy. 1997. RNA helicase A mediates association of CBP with RNA polymerase II. Cell 90:1107-1112. 121. Neish, A. S., S. F. Anderson, B. P. Schlegel, W. Wei, and J. D. Parvin. 1998. Factors associated with the mammalian RNA polymerase II holoenzyme. Nucleic Acids Res. 26:847-853. 122. Cho, H., G. Orphanides, X. Sun, X. J. Yang, V. Ogryzko, E. Lees, Y. Nakatani, and D. Reinberg. 1998. A human RNA polymerase II complex containing factors that modify chromatin structure. Mol.Cell Biol. 18:53555363. 123. Yang, X. J., V. V. Ogryzko, J. Nishikawa, B. H. Howard, and Y. Nakatani. 1996. A p300/CBP-associated factor that competes with the adenoviral oncoprotein E1A. Nature 382:319-324. 124. Brownell, J. E., J. Zhou, T. Ranalli, R. Kobayashi, D. G. Edmondson, S. Y. Roth, and C. D. Allis. 1996. Tetrahymena histone acetyltransferase A: a homolog to yeast Gcn5p linking histone acetylation to gene activation. Cell 84:843-851. 125. Bannister, A. J. and T. Kouzarides. 1996. The CBP co-activator is a histone acetyltransferase. Nature 384:641-643. 126. Ogryzko, V. V., R. L. Schiltz, V. Russanova, B. H. Howard, and Y. Nakatani. 1996. The transcriptional coactivators p300 and CBP are histone acetyltransferases. Cell 87:953-959. 127. Huo, X. and J. Zhang. 2005. Important roles of reversible acetylation in the function of hematopoietic transcription factors. J.Cell Mol.Med. 9:103-112. 128. Kouzarides, T. 2000. Acetylation: a regulatory modification to rival phosphorylation? EMBO J. 19:1176-1179. 140 129. Gu, W. and R. G. Roeder. 1997. Activation of p53 sequence-specific DNA binding by acetylation of the p53 C-terminal domain. Cell 90:595-606. 130. Munshi, N., M. Merika, J. Yie, K. Senger, G. Chen, and D. Thanos. 1998. Acetylation of HMG I(Y) by CBP turns off IFN beta expression by disrupting the enhanceosome. Mol.Cell 2:457-467. 131. Polesskaya, A., I. Naguibneva, A. Duquet, E. Bengal, P. Robin, and A. Harel-Bellan. 2001. Interaction between acetylated MyoD and the bromodomain of CBP and/or p300. Mol.Cell Biol. 21:5312-5320. 132. Chen, H., R. J. Lin, W. Xie, D. Wilpitz, and R. M. Evans. 1999. Regulation of hormone-induced histone hyperacetylation and gene activation via acetylation of an acetylase. Cell 98:675-686. 133. Ito, A., C. H. Lai, X. Zhao, S. Saito, M. H. Hamilton, E. Appella, and T. P. Yao. 2001. p300/CBP-mediated p53 acetylation is commonly induced by p53activating agents and inhibited by MDM2. EMBO J. 20:1331-1340. 134. Ito, A., Y. Kawaguchi, C. H. Lai, J. J. Kovacs, Y. Higashimoto, E. Appella, and T. P. Yao. 2002. MDM2-HDAC1-mediated deacetylation of p53 is required for its degradation. EMBO J. 21:6236-6245. 135. Bannister, A. J. and E. A. Miska. 2000. Regulation of gene expression by transcription factor acetylation. Cell Mol.Life Sci. 57:1184-1192. 136. Polesskaya, A., A. Duquet, I. Naguibneva, C. Weise, A. Vervisch, E. Bengal, F. Hucho, P. Robin, and A. Harel-Bellan. 2000. CREB-binding protein/p300 activates MyoD by acetylation. J.Biol.Chem. 275:34359-34364. 137. Sartorelli, V., P. L. Puri, Y. Hamamori, V. Ogryzko, G. Chung, Y. Nakatani, J. Y. Wang, and L. Kedes. 1999. Acetylation of MyoD directed by PCAF is necessary for the execution of the muscle program. Mol.Cell 4:725734. 138. Polesskaya, A. and A. Harel-Bellan. 2001. Acetylation of MyoD by p300 requires more than its histone acetyltransferase domain. J.Biol.Chem. 276:44502-44503. 139. Tomita, A., M. Towatari, S. Tsuzuki, F. Hayakawa, H. Kosugi, K. Tamai, T. Miyazaki, T. Kinoshita, and H. Saito. 2000. c-Myb acetylation at the carboxyl-terminal conserved domain by transcriptional co-activator p300. Oncogene 19:444-451. 140. Sano, Y. and S. Ishii. 2001. Increased affinity of c-Myb for CREB-binding protein (CBP) after CBP-induced acetylation. J.Biol.Chem. 276:3674-3682. 141 141. Monica, K., D. P. LeBrun, D. A. Dedera, R. Brown, and M. L. Cleary. 1994. Transformation properties of the E2a-Pbx1 chimeric oncoprotein: fusion with E2a is essential, but the Pbx1 homeodomain is dispensable. Mol.Cell Biol. 14:8304-8314. 142. Qiu, Y., A. Sharma, and R. Stein. 1998. p300 mediates transcriptional stimulation by the basic helix-loop-helix activators of the insulin gene. Mol.Cell Biol. 18:2957-2964. 143. Bradney, C., M. Hjelmeland, Y. Komatsu, M. Yoshida, T. P. Yao, and Y. Zhuang. 2003. Regulation of E2A activities by histone acetyltransferases in B lymphocyte development. J.Biol.Chem. 278:2370-2376. 144. Zor, T., R. N. De Guzman, H. J. Dyson, and P. E. Wright. 2004. Solution structure of the KIX domain of CBP bound to the transactivation domain of cMyb. J.Mol.Biol. 337:521-534. 145. Le Douarin, B., A. L. Nielsen, J. M. Garnier, H. Ichinose, F. Jeanmougin, R. Losson, and P. Chambon. 1996. A possible involvement of TIF1 alpha and TIF1 beta in the epigenetic control of transcription by nuclear receptors. EMBO J. 15:6701-6715. 146. Heery, D. M., E. Kalkhoven, S. Hoare, and M. G. Parker. 1997. A signature motif in transcriptional co-activators mediates binding to nuclear receptors. Nature 387:733-736. 147. Torchia, J., D. W. Rose, J. Inostroza, Y. Kamei, S. Westin, C. K. Glass, and M. G. Rosenfeld. 1997. The transcriptional co-activator p/CIP binds CBP and mediates nuclear-receptor function. Nature 387:677-684. 148. Chitayat, S., C. M. Denis, M. J. Plevin, S. Lui, H. L. Spencer, M. Ikura, D. P. LeBrun, and S. P. Smith. Unpublished data. 2007. 149. Kamps, M. P. and D. D. Wright. 1994. Oncoprotein E2A-Pbx1 immortalizes a myeloid progenitor in primary marrow cultures without abrogating its factordependence. Oncogene 9:3159-3166. 150. Hung, H. L., J. Lau, A. Y. Kim, M. J. Weiss, and G. A. Blobel. 1999. CREBBinding protein acetylates hematopoietic transcription factor GATA-1 at functionally important sites. Mol.Cell Biol. 19:3496-3505. 151. Hung, H. L., A. Y. Kim, W. Hong, C. Rakowski, and G. A. Blobel. 2001. Stimulation of NF-E2 DNA binding by CREB-binding protein (CBP)-mediated acetylation. J.Biol.Chem. 276:10715-10721. 152. Weiss, M. J., C. Yu, and S. H. Orkin. 1997. Erythroid-cell-specific properties of transcription factor GATA-1 revealed by phenotypic rescue of a genetargeted cell line. Mol.Cell Biol. 17:1642-1651. 142 153. Crossley, M., M. Merika, and S. H. Orkin. 1995. Self-association of the erythroid transcription factor GATA-1 mediated by its zinc finger domains. Mol.Cell Biol. 15:2448-2456. 154. Chakrabarti, S. R., R. Sood, S. Nandi, and G. Nucifora. 2000. Posttranslational modification of TEL and TEL/AML1 by SUMO-1 and cellcycle-dependent assembly into nuclear bodies. Proc.Natl.Acad.Sci.U.S A 97:13281-13285. 155. Musti, A. M., M. Treier, and D. Bohmann. 1997. Reduced ubiquitindependent degradation of c-Jun after phosphorylation by MAP kinases. Science 275:400-402. 156. Treier, M., L. M. Staszewski, and D. Bohmann. 1994. Ubiquitin-dependent cJun degradation in vivo is mediated by the delta domain. Cell 78:787-798. 157. Dignam, J. D., P. L. Martin, B. S. Shastry, and R. G. Roeder. 1983. Eukaryotic gene transcription with purified components. Methods Enzymol. 101:582-598. 158. Kraus, W. L., E. T. Manning, and J. T. Kadonaga. 1999. Biochemical analysis of distinct activation functions in p300 that enhance transcription initiation with chromatin templates. Mol.Cell Biol. 19:8123-8135. 159. Horiuchi, K. and D. Fujimoto. 1975. Use of phosph-cellulose paper disks for the assay of histone acetyltransferase. Anal.Biochem. 69:491-496. 160. Mizzen, C. A., J. E. Brownell, R. G. Cook, and C. D. Allis. 1999. Histone acetyltransferases: preparation of substrates and assay procedures. Methods Enzymol. 304:675-696. 161. Berndsen, C. E. and J. M. Denu. 2005. Assays for mechanistic investigations of protein/histone acetyltransferases. Methods 36:321-331. 162. Zong, H., Z. Li, L. Liu, Y. Hong, X. Yun, J. Jiang, Y. Chi, H. Wang, X. Shen, Y. Hu, Z. Niu, and J. Gu. 2005. Cyclin-dependent kinase 11(p58) interacts with HBO1 and enhances its histone acetyltransferase activity. FEBS Lett. 579:3579-3588. 163. Martinez-Balbas, M. A., A. J. Bannister, K. Martin, P. Haus-Seuffert, M. Meisterernst, and T. Kouzarides. 1998. The acetyltransferase activity of CBP stimulates transcription. EMBO J. 17:2886-2893. 164. Zhang, W. and J. J. Bieker. 1998. Acetylation and modulation of erythroid Kruppel-like factor (EKLF) activity by interaction with histone acetyltransferases. Proc.Natl.Acad.Sci.U.S A 95:9855-9860. 143 165. Mal, A., M. Sturniolo, R. L. Schiltz, M. K. Ghosh, and M. L. Harter. 2001. A role for histone deacetylase HDAC1 in modulating the transcriptional activity of MyoD: inhibition of the myogenic program. EMBO J. 20:1739-1753. 166. Sartorelli, V., J. Huang, Y. Hamamori, and L. Kedes. 1997. Molecular mechanisms of myogenic coactivation by p300: direct interaction with the activation domain of MyoD and with the MADS box of MEF2C. Mol.Cell Biol. 17:1010-1026. 167. Riou, P., F. Bex, and L. Gazzolo. 2000. The human T cell leukemia/lymphotropic virus type 1 Tax protein represses MyoD-dependent transcription by inhibiting MyoD-binding to the KIX domain of p300. A potential mechanism for Tax-mediated repression of the transcriptional activity of basic helix-loop-helix factors. J.Biol.Chem. 275:10551-10560. 168. Lange, A., R. E. Mills, C. J. Lange, M. Stewart, S. E. Devine, and A. H. Corbett. 2007. Classical nuclear localization signals: definition, function, and interaction with importin alpha. J.Biol.Chem. 282:5101-5105. 169. Faiola, F., Y. T. Wu, S. Pan, K. Zhang, A. Farina, and E. Martinez. 2007. Max is acetylated by p300 at several nuclear localization residues. Biochem.J. 403:397-407. 170. Hebrok, M., K. Wertz, and E. M. Fuchtbauer. 1994. M-twist is an inhibitor of muscle differentiation. Dev.Biol. 165:537-544. 171. Spicer, D. B., J. Rhee, W. L. Cheung, and A. B. Lassar. 1996. Inhibition of myogenic bHLH and MEF2 transcription factors by the bHLH protein Twist. Science 272:1476-1480. 172. Hebrok, M., A. Fuchtbauer, and E. M. Fuchtbauer. 1997. Repression of muscle-specific gene activation by the murine Twist protein. Exp.Cell Res. 232:295-303. 173. Hamamori, Y., H. Y. Wu, V. Sartorelli, and L. Kedes. 1997. The basic domain of myogenic basic helix-loop-helix (bHLH) proteins is the novel target for direct inhibition by another bHLH protein, Twist. Mol.Cell Biol. 17:65636573. 174. Murray, S. S., C. A. Glackin, K. A. Winters, D. Gazit, A. J. Kahn, and E. J. Murray. 1992. Expression of helix-loop-helix regulatory genes during differentiation of mouse osteoblastic cells. J.Bone Miner.Res. 7:1131-1138. 175. Lee, M. S., G. N. Lowe, D. D. Strong, J. E. Wergedal, and C. A. Glackin. 1999. TWIST, a basic helix-loop-helix transcription factor, can regulate the human osteogenic lineage. J.Cell Biochem. 75:566-577. 144 176. Rice, D. P., T. Aberg, Y. Chan, Z. Tang, P. J. Kettunen, L. Pakarinen, R. E. Maxson, and I. Thesleff. 2000. Integration of FGF and TWIST in calvarial bone and suture development. Development 127:1845-1855. 177. Hamamori, Y., V. Sartorelli, V. Ogryzko, P. L. Puri, H. Y. Wu, J. Y. Wang, Y. Nakatani, and L. Kedes. 1999. Regulation of histone acetyltransferases p300 and PCAF by the bHLH protein twist and adenoviral oncoprotein E1A. Cell 96:405-413. 178. Cote, J., R. T. Utley, and J. Workman. 1995. Basic Analysis of Transcription Factor Binding to Nucleosomes. Methods in Molecular Genetics 6:108-128. 179. Reeves, R., C. M. Gorman, and B. Howard. 1985. Minichromosome assembly of non-integrated plasmid DNA transfected into mammalian cells. Nucleic Acids Res. 13:3599-3615. 180. Pullner, A., J. Mautner, T. Albert, and D. Eick. 1996. Nucleosomal structure of active and inactive c-myc genes. J.Biol.Chem. 271:31452-31457. 181. Stanfield-Oakley, S. A. and J. D. Griffith. 1996. Nucleosomal arrangement of HIV-1 DNA: maps generated from an integrated genome and an EBV-based episomal model. J.Mol.Biol. 256:503-516. 182. Sigvardsson, M., M. O'Riordan, and R. Grosschedl. 1997. EBF and E47 collaborate to induce expression of the endogenous immunoglobulin surrogate light chain genes. Immunity. 7:25-36. 183. Sigvardsson, M. 2000. Overlapping expression of early B-cell factor and basic helix-loop-helix proteins as a mechanism to dictate B-lineage-specific activity of the lambda5 promoter. Mol.Cell Biol. 20:3640-3654. 184. Gisler, R. and M. Sigvardsson. 2002. The human V-preB promoter is a target for coordinated activation by early B cell factor and E47. J.Immunol. 168:51305138. 185. Zhao, F., R. McCarrick-Walmsley, P. Akerblad, M. Sigvardsson, and T. Kadesch. 2003. Inhibition of p300/CBP by early B-cell factor. Mol.Cell Biol. 23:3837-3846. 186. Zerby, D., C. J. Chen, E. Poon, D. Lee, R. Shiekhattar, and P. M. Lieberman. 1999. The amino-terminal C/H1 domain of CREB binding protein mediates zta transcriptional activation of latent Epstein-Barr virus. Mol.Cell Biol. 19:1617-1626. 187. Chen, C. J., Z. Deng, A. Y. Kim, G. A. Blobel, and P. M. Lieberman. 2001. Stimulation of CREB binding protein nucleosomal histone acetyltransferase activity by a class of transcriptional activators. Mol.Cell Biol. 21:476-487. 145 188. Deng, Z., C. J. Chen, M. Chamberlin, F. Lu, G. A. Blobel, D. Speicher, L. A. Cirillo, K. S. Zaret, and P. M. Lieberman. 2003. The CBP bromodomain and nucleosome targeting are required for Zta-directed nucleosome acetylation and transcription activation. Mol.Cell Biol. 23:2633-2644. 189. Goodman, R. H. and S. Smolik. 2000. CBP/p300 in cell growth, transformation, and development. Genes Dev. 14:1553-1577. 190. Blobel, G. A. 2000. CREB-binding protein and p300: molecular integrators of hematopoietic transcription. Blood 95:745-755. 191. Yao, T. P., S. P. Oh, M. Fuchs, N. D. Zhou, L. E. Ch'ng, D. Newsome, R. T. Bronson, E. Li, D. M. Livingston, and R. Eckner. 1998. Gene dosagedependent embryonic development and proliferation defects in mice lacking the transcriptional integrator p300. Cell 93:361-372. 192. Oike, Y., N. Takakura, A. Hata, T. Kaname, M. Akizuki, Y. Yamaguchi, H. Yasue, K. Araki, K. Yamamura, and T. Suda. 1999. Mice homozygous for a truncated form of CREB-binding protein exhibit defects in hematopoiesis and vasculo-angiogenesis. Blood 93:2771-2779. 193. Kung, A. L., V. I. Rebel, R. T. Bronson, L. E. Ch'ng, C. A. Sieff, D. M. Livingston, and T. P. Yao. 2000. Gene dose-dependent control of hematopoiesis and hematologic tumor suppression by CBP. Genes Dev. 14:272277. 194. Hennekam, R. C. 2006. Rubinstein-Taybi syndrome. Eur.J.Hum.Genet. 14:981-985. 195. Kalkhoven, E., J. H. Roelfsema, H. Teunissen, A. den Boer, Y. Ariyurek, A. Zantema, M. H. Breuning, R. C. Hennekam, and D. J. Peters. 2003. Loss of CBP acetyltransferase activity by PHD finger mutations in Rubinstein-Taybi syndrome. Hum.Mol.Genet. 12:441-450. 196. Yang, X. J. 2004. The diverse superfamily of lysine acetyltransferases and their roles in leukemia and other diseases. Nucleic Acids Res. 32:959-976. 197. Dilworth, F. J., K. J. Seaver, A. L. Fishburn, S. L. Htet, and S. J. Tapscott. 2004. In vitro transcription system delineates the distinct roles of the coactivators pCAF and p300 during MyoD/E47-dependent transactivation. Proc.Natl.Acad.Sci.U.S A 101:11593-11598. 146