Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
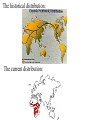
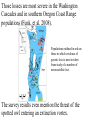
Case Studies I Black-footed ferret (Mustela nigripes): This species is a member of the weasel family that formerly occupied plains and prairie habitat from Saskatchewan to Texas. It was listed as threatened in the US in 1967 and endangered in 1973. An initial recovery plan was devised by the US Fish and Wildlife Service in 1978. By then, however, there were no known wild black-footed ferrets. Decline of the species coincided with, and may have been caused by, the tremendous decline (90-95%) in prairie dog abundance earlier this century. Prairie dogs are the primary (90%) food of black footed ferrets. Prairie dogs were targeted as pests because their burrows damaged farm implements and tractors. Prairie dogs also appear to have suffered from introduction of Sylvatic plague and from canine distemper. These diseases then affected ferrets. Prairie Dogs are Considered Keystone Species • are the primary (90%) food of black footed ferrets, swift fox, the golden eagle, the badger, and the ferruginous hawk. • species, such as the mountain plover and the burrowing owl rely on prairie dog burrows for nesting areas. • grazing species such as bison, pronghorn and mule deer may prefer the vegetative conditions after prairie dogs have foraged through the area. In 1964, small population discovered in S. Dakota. It was studied and brought into captivity in 1971 for captive rearing attempts. Captive breeding was a dismal failure. 1978 was the end of known ferret existence in wild. The species was re-discovered at a single site in Wyoming in 1981. A Species Survival Plan (SSP) was developed based on captive propagation to eventually re-release ferrets into the wild. A severe decline in prairie dog abundance was evident by 1985, when ferrets were collected to begin the captive propagation program. By 1985, black footed ferrets were limited to ~130 individuals in one population at Meteetsie, Wyoming. Another re-discovered population (from South Dakota) had been placed in a captive breeding program without success. (this population went extinct; good conservation biology?) The Wyoming population was surveyed but not captured. This population suffered after plague was discovered among its prairie dog prey. Six ferrets were captured for a captive breeding program; all died of canine distemper. Additional collections of 6 and 12 animals were made for captive breeding programs. The last known free-ranging individuals died from distemper, resting the fate of the species in the last 12 (eventually only 7) individuals in the captive breeding program. Goals were quickly set to maintain as much (90%) genetic diversity as possible for a minimum of 50 years. Two litters of kits were born in 1987. Since then 4800 kits have been born in captivity (Grenier 2007). In 1988 the captive population was subdivided into two isolated groups (to minimize chances of catastrophic extinction of the species). By the early 1990s the captive breeding program was was producing >100 kits annually at 6 captive breeding sites (including the Toronto zoo). Ferrets were reintroduced to southern Wyoming in 1991 (228 ferrets over the period 1991-4). By 1992 some individuals (12%) survived the winter and reproduced successfully in the wild. Coyotes appeared to be the primary predator and source of mortality, and survival was only moderate (20-25% for 30 days). The population is growing (slowly) and other introductions of ferrets have also occurred (see below). In the Shirley Basin (Wyoming) the release was thought to be failing. By 1996 there were <25 ferrets remaining from the release, and monitoring became sporadic. But then in 2003 there were 52, and continued increase found an estimated 223 in 2007. In 2005 the estimated ferret population in South Dakota was 400. Verification of this estimate has not proved easy. Since reintroductions in Wyoming and South Dakota, a number of successful reintroductions have occurred: 186 released beginning in 2001 in northwest Colorado; in January 2006 wild reproduction found. Continuing releases in Aubrey Valley, Arizona must have been successful. In 2005 14 ferrets were counted that must have been born in the wild (no microchip marker). There are four other active re-release sites (8, aiming for 10 in all) being monitored for success in the U.S. and Mexico. The SSP calls for 10 populations with 30 breeding adults in each. The species will then be downgraded to threatened. Location / Year introduced and population as of 2010 1. Shirley Basin (1991; > 30) 2. Conata Basin/Badlands (1994; > 30) 3. UL Bend Refuge (1996; < 30) 4. Aubrey Valley (1996; < 30) 6. Coyote Basin (2000; < 30) 7. Cheyenne River Reservation (2000; > 30) 8. Wolf Creek (2000; < 30) 9. 40-Complex (2000; < 30) 10. Janos, Chihuahua (2000; < 30) 11. Rosebud Sioux Reservation (2000; < 30) 12. Lower Brule Reservation (2006; < 30) 13. Wind Cave National Park (2007; < 30) 14. Logan County (2007; < 30) 15. Northern Cheyenne Reservation (2008; < 30) 16. Espee Ranch (2008; < 30) Dynamics of the Shirley Basin reintroduction: Grenier et al. used demographic modeling to determine keys to observed growth. They expected, as a mammal, adult survivorship and long-term fertility were important. However, first year survival and early fertility were the keys. Success is indicated by an estimated = 1.35. Note the implication of this rapid growth for bottleneck effects. For conservation, that means long-term monitoring may not be necessary. None have been observed in Canada since 1937. The re-introduction sites are a first example of the problem imposed by political opposition. The reintroduced ferrets are (in the language of the U.S. Endangered Species legislation) a nonessential, experimental population. Under this designation, the animals are protected at the re-introduction site, but are left unprotected should they move into a farmer's field or a rancher's pastureland. What are the genetic implications of reintroduction? Among the possible effects on re-introduced animals are: disease exposure and predation. In addition to the natural bottleneck that led to reintroduction, the ‘new’ population can face a bottleneck, as well. The genetic result of a bottleneck/re-introduction can be: founder effects genetic drift loss of genetic variation inbreeding depression allele fixation decreased fitness increased probability of local extinction What tools/procedures can alleviate the genetic danger to black-footed ferrets? 1.Translocation among isolated reintroduced populations. 2.Augmentation (annually) of reintroduced populations that are not growing from captive breeding stocks. This is what was done. What evidence points to the need for one or both of these steps? Among the 3 reintroduced populations studied (South Dakota, Wyoming and Arizona), dynamics suggest that the Wyoming population was most in need (Wisely, et al. 2008). Descriptive genetic parameters for three reintroduced populations of black-footed ferrets Population n He ± SE Ho ± SE A South Dakota 44 0.41 ± 0.01 0.40 ± 0.01 2.00 Wyoming 32 0.21 ± 0.04 0.21 ± 0.01 1.43 Arizona 31 0.34 ± 0.02 0.28 ± 0.01 2.14 Captive 78 0.37 ± 0.01 0.38 ± 0.01 2.00 There was no evidence that either Arizona or South Dakota populations had lost genetic diversity. All alleles in the captive source population were found present in the South Dakota reintroduced population. The genetic diversity was retained even after augmentation ended. Exponential growth has apparently obviated any bottleneck effects. Genetic diversity has been retained in Arizona, as well. Apparently this is due to continued augmentation, since growth has been slow (minimal). A new mutant allele not present in the source population has even increased the mean number of alleles to be larger than in the source. Wyoming has apparently been subjected to an extended (multiple generation) bottleneck. 4 of 7 loci studied have become fixed there. There was a 28% loss of allelic diversity. An extended period at low numbers apparently led to drift, decreased genetic diversity and increased inbreeding. There was also morphological evidence of inbreeding depression. Ferrets from Wyoming were significantly smaller than those from either Arizona or South Dakota, even though all originated from the same captive source population. African cheetah (Acinonyx jubatus): The cheetah was once found on 5 continents. At the turn of the 20th century it occurred in both Africa and Asia). Today it is limited to Africa and a small population in Iran. The historical distribution: The current distribution: Recent estimates (Selebatso, et al. 2008) place the total population of cheetahs at <10,000 mature individuals, with <1000 in each of the extant subpopulations. The main prey for cheetahs are: impala, springbok, the young of large antelopes, and only to a lesser extent farm animals like goats and sheep. That has led to a generally supportive atmosphere for cheetah conservation, at least for the largest natural population in Namibia. However, … The cheetah has the lowest frequency of polymorphic loci (0.0) and lowest average heterozygosity (0.0). Overall, the cheetah had between 10 and 100 times less genetic variability than other mammals. O'Brien attributed the patterns in cheetah to a severe population bottleneck followed by inbreeding. The bottleneck would reduce genetic diversity as a result of selection pressures and genetic drift. They attribute the bottleneck to decimation of the population by legal and illegal hunting by African cattle farmers about 100 cheetah generations ago. A low sperm count and abnormal sperm is another evidence of a bottleneck and inbreeding. In zoos noninbred cheetah mating had among the highest infant mortality rates of all mammals surveyed. As well, infant mortality rates for inbred and noninbred cheetah mating did not differ significantly, suggesting that inbreeding has no pronounced effect today (largely because strong effects were evident earlier). Another evidence of the limited genetic diversity among cheetahs comes from a study of skin grafts. 7 different skin grafts were performed on non-inbred pairs of cheetahs (14 individuals). Successful grafts depend on acceptance of 'donor' tissue by the 'recipient' individual, which is governed by a group of genes called the 'major histocompatability complex' (MHC). In all vertebrates, the MHC is the most polymorphic region of the genome, thus it should be most useful in differentiating genetically different individuals. All of the grafts succeeded through the typical stage of rejection, though control grafts of house cat tissue were rejected. A comparison of frequency of enzyme polymorphisms and heterozygosity levels in subspecies of the cheetah, the south African form A. jubatus jubatus and its east African relative A. jubatus raineyi found the genetic distance between subspecies was minimal (0.004), indicating that the cheetah became genetically impoverished before the subspecies diverged. These genetic patterns are most consistent with 2 bottlenecks (one 10,000 years ago and another during the past century) followed by inbreeding. Conservation genetics has moved past enzyme polymorphism into DNA analysis, using microsatellites and MtDNA. Menotti-Raymond and O'Brien (1993) used hypervariable minisatellite loci and mitochondrial loci to time the bottleneck in the cheetah population. Based on expected mutation rates and current levels of diversity, they back calculated the bottlenecks to between 3500 - 12,700 years and 28,000 - 36,000 years, respectively, for mitochondria and minisatellite techniques. These techniques also identified only 1 to 10% of DNA diversity found among other out-crossed cat species. O’Brien and his collaborators have become involved in controversy over their conclusions about cheetahs. Merola (1994) compared the cheetah's genetic variability with that of other carnivorous vertebrates. Of 24 terrestrial carnivores surveyed, 8 had no heterozygosity (H = 0), while the remaining ones averaged H = 0.042 (vs. H = 0.014 for the cheetah). Merola concluded stated that the lack of breeding success and high infant mortality rates were due to poor captive breeding program procedures, and that the feline virus that decimated the Oregon cheetahs was effective because the cheetahs were held at very high density. She argued that as long as recessive alleles (deleterious) were slowly purged from the population, the resulting population could be relatively homozygous but without inbreeding effects. The inbreeding effects observed in cheetahs would thus be an artifact of the artificial captive breeding environment. More recent molecular genetics (Marker et al. 2008) indicate that there is limited genetic variability and differentiation among cheetah populations from Namibia, but that there is panmixis across large areas. The distance between captures of close relatives indicates how far cheetahs may move: Relationship Mean Distance between captures (km) Dam & daughter 13 Dam & son 116.38 Sire & daughter 93.50 Sire & son 99.06 Sibs 121.00 Overall 90.66 For Namibian cheetahs, habitat conservation and promotion of natural dispersal and gene flow is critical to species conservation. Merola also acknowledged that the cheetah is suffering. From her perspective, similarly, it is from a loss of habitat and other adverse human effects. For example, habitat destruction has resulted in population densities of one cheetah per 6 km2 rather than the old rate of 1 per 100 km2. High densities facilitate transmission and spread of disease and 'focusing' of cheetah predators in the small reserves. A recent addition to the debate was demographic modeling contributed by Crooks et al. (1998) using published data from the Serengeti. The importance of elevated cub mortality was relatively minor relative to the large effects from variation in adult survivorship. Demographically, the adults have high reproductive value and cubs low value. Any change in the adult survivorship schedule has a much larger impact than an equal change in cub survivorship (for cheetahs). They argue that focusing on cub mortality could obscure the importance of factors producing even a small increase in adult mortality. This conclusion, based on modeling, is also controversial. How does this conclusion compare with the result of demographic modeling for black-footed ferrets? Recent matrix models of cheetah population dynamics (Lubben et al. 2008) suggest instead that small changes in the survivorship of cubs could greatly enhance the likelihood of population and species survival. High infant mortality • Kelley et al. (1998) radio-collared female cheetahs in the Serengeti and followed them as they traveled throughout their 800-km2 home ranges. Identified birthing sites (lairs). • Entered lairs when adults were away and counted young. Regular monitoring showed that young suffered from high mortality rates (80 %). • Most mortality was predation related – not genetic defects. Kelley et al. 1998. Journal of Zoology 244:473-488 Durant et al. (2007) reviewed information that suggests cheetah behaviour and interaction with lions explains demography in Serengeti populations and is critical in their conservation status. Cheetah males live in small social groups (coalitions) that move over small defended territories, while females move with cubs over larger areas. They hunt away from the cubs for short periods. The relative movements create ‘hotspots’ of cheetah population density. Cheetah cub (and adult) survival is negatively correlated with local lion density. Cheetah populations undergo ‘dangerous’ declines when they are in high local density and in the vicinity of lions. Lowered fecundity • Reproduction in captivity is low – as of 1986, only 17 of 108 females and 12 of 85 males had bred in zoos (~ 84 % of captive cheetah do not breed) Does this mirror natural conditions? Lowered fecundity • Wild cheetahs are polyestrus, cycling ~ every 12 days with a gestation period of ~ 93 days. • For wild cheetahs, high numbers of females breeding and rapid rates of litter production suggest that the reproductive physiology of neither sex is compromised. Survival in an area (even the entire Serengeti, an IUCN category II park including 14,000 km2, is too small to maintain a minimum viable population of cheetahs) is dependent on movement among population groups (and so that populations may grow in areas with low lion numbers) and access to the antelopes they hunt outside the Serengeti. Humans were also indicated as important in cheetah declines. However, Durant et al. report survey results that suggest the farmers consider cheetahs far less important than other large predators. It seems they are not likely important to the declines in cheetahs. Northern Spotted Owl (S. occidentalis caurina): Northern Spotted Owls occur in the southwest region of British Columbia and in Oregon and Washington. In all instances, the owl is rare (low abundance) even in the best of habitats. In southwestern B.C., the owl was found at 14 sites, with a total population of as few as 100 individuals (Dunbar et al. 1991). They attributed its rarity to habitat destruction (logging, fires, development) and to Barred Owls which live in the same old-growth habitat and which respond aggressively to spotted owl calls (thus potentially limiting its habitat availability). In the USA, the northern spotted owl has pitted environmentalists against loggers. The case was resolved during summer 1995 by the conservativeleaning Supreme Court in favour of preservation of essential lands for owl habitat. Habitat loss in the U.S. has been extensive: The result of the conflict was the development of the Northwest Forest Plan Here is the history of the conflict: • Historical practice of clearcut logging in Pacific Northwest. • U.S.F.W.S. reviewed the status of the Northern Spotted Owl in 1982 and 1987 - concluded it did not warrant listing as threatened or endangered. • Reviews in 1989 and 1990 proposed listing as a threatened species under the ESA. Loss of old-growth habitat was cited as the primary threat. • Listing was implemented on June 23, 1990. • Logging in national forests was stopped by court order in 1991. Bart and Forsman (1992) and Bart (1995) looked at spotted owl density and breeding success in habitats of differing quality in Washington and Oregon. In sum, the higher the percentage of old growth forest (good habitat), the higher the owls/km2, breeding pairs/km2, young fledged/km2, and adult survival. Figures showing you that: To provide clear answers to key questions about the spotted owl populations, Murphy and Noon (1992) formulated a number of important, testable hypotheses regarding the owl: 1.Is the owl population growing (is [finite rate of growth] >1? Answer: No 2.Do owls differentiate among forests of different ages or structures. Answer: Yes The owls prefer habitats based with old-growth forest disproportionate to the abundance of this habitat type. 3.Habitat type selected by the owls has not changed in abundance. Answer: No, it has decreased. 4. The probability of persistence is not related to the extent of its geographic distribution. Answer: It is. 5. There is a relationship between HCA (habitat conservation area) size and its owl carrying capacity. Answer: Yes. 6. A relationship exists between habitat fragmentation and persistence likelihood of species using that habitat landscape. Answer: Yes 7. Distance between habitat patches has a bearing on dispersal success of juvenile owls. Answer: Yes, there is a very strong relationship. Probability of successful owl dispersal versus distance between suitable habitat patches Based on these answers, a map of suitable habitat patches for spotted owl conservation was constructed for Oregon and Washington: What additional problems remain? Habitat destruction is clearly key – not just logging, but forest fires. The late successional forest areas most important to spotted owls also have fuel conditions that make fires likely. Ager et al. (2007) showed via modeling that treatment of fuel conditions on relatively small proportions (20%) of old growth forest areas had a disproportionate effect in reducing forest fire likelihood (a 44% reduction). In the last two years the draft management plan that resulted has been criticized from all fronts (Stokstad 2008). The forestry industry wants to see less forest protected. They claim that the barred owl is a much more serious threat. Environmentalists claim not enough is being protected. In the U.S. the spotted owl population continues to decline by 3.7% per year. A recent genetic analysis by the U.S. Geological Survey suggests that there is now evidence of loss of genetic variability (decreased Ne) and inbreeding depression even beyond the numerical decline. Those losses are most severe in the Washington Cascades and in southern Oregon Coast Range populations (Funk, et al. 2008). Populations outlined in red are those in which evidence of genetic loss is most evident from study of a number of microsatellite loci. The survey results even mention the threat of the spotted owl entering an extinction vortex. Some suggest that the threat of forest fires is so dire that forests need to be thinned to reduce the risk of catastrophic damage. Politics continues to play a major role. Many important scientists refuse to take part in the Fish & Wildlife Service’s development of a final management plan due to interference from the Dept. of the Interior and Bureau of Land Management. They forced inclusion of an “option 2”. That option reduced land set aside as spotted owl habitat and increased flexibility in permitting logging (particularly in Oregon). It appears this conflict is headed back into the courts! References Ager, A.A. et al. (2007) Modeling wildfire risk to northern spotted owl (Strix occidentalis caurina) habitat in Central Oregon, U.S.A. Forest Ecol. Manag. 246:45-56. Bart, J. 1995. Amount of suitable habitat and viability of northern spotted owls. Conservation Biology 9:943-946. Bart, J. and E.D. Forsman. 1992. Dependence of northern spotted owls (Strix occidentalis caurina), on old-growth forests in the western USA. Biological Conservation 62:95-100. Crooks, K.R., M.A. Sanjayan, D.F. Doaks. 1998. New insights on cheetah conservation through demographic modeling. Conservation Biology 12: 889-895. Dunbar, D.L. et al. 1991. Status of Spotted Owl, Strix occidentalis, and Barred Owl, Strix varia, in southwestern British Columbia. Canadian Field Naturalist 105:464-468. Durant, S.M. et al. 2007. Relating long-term studies to conservation practice: the case of the Serengeti Cheetah project. Conservation Biology 21:602-611. Funk, W.C., E.D. Forsman, T.D. Mullins and S.M. Haig. 2008. Genetics show currect decline and Pleistocene expansion in Northern Spotted Owls. U.S. Geological Survey Open-File Report 2008-1239, 12 p. Grenier, M.B., D.B. McDonald and S.W. Buskirk. Rapid population growth of a critically endangered carnivore. Science 317:779. Laurenson, M.K., N. Wielebnowski and T.M. Caro. 1995. Extrinsic factors and juvenile mortality in cheetahs. Conservation Biology 9:1329-1331. Lubben, J. et al. 2008. Management recommendations based on matrix projection models: The importance of considering biological limits. Biological Conservation 141:517-523. Marker, L.L. et al. 2008. Molecular genetic insights on Cheetah (Acinonyx jubatus) ecology and conservation in Namibia. J. Heredity 99:2-13. May, R.M. 1995. The cheetah controversy. Nature 374:309-310. Menotti-Raymond, M. and S.J. O'Brien. 1993. Dating of the genetic bottleneck of the African cheetah. Proceedings of the National Academy of Science 90:3172-3176. Merola, M. 1994. A reassessment of homozygosity and the case for inbreeding depression in the cheetah, Acinonyx jubatus: implications for conservation. Conservation Biology 8:961-971. Murphy, D.D. and B.R. Noon. 1992. Integrating scientific methods with habitat conservation planning: reserve design for northern spotted owls. Ecological Applications 2:3-17. O'Brien, S.J., D. Wildt, D. Goldman, C. Merril and M. Bush. 1983. The cheetah is depauperate in genetic variation. Science 221:469-462. O'Brien, S.J. and 9 others. 1985. Genetic basis for species vulnerability in the cheetah. Science 227:1428-1434. O'Brien, S.J. and 7 others. 1987. East African cheetahs: evidence for two population bottlenecks? Proceedings of the National Academy of Science 84:508-511. O'Brien, S.J. 1994. The cheetah's conservation controversy. Conservation Biology 8:1153-1155. Stokstad, E. 2008. Spotted owl recovery plan flawed, Review panel finds. Science 320:594-5. Wisely, S.M., R.M. Santymire, T.M. Livieri, S.A. Mueting and J. Howard. Genetic and phenotypic consequences of reintroduction history in the black-footed ferret (Mustela nigripes). Conservation Genetics 9:389-399.