Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
1994 84: 3422-3428
B-cell non-Hodgkin's lymphoma cell line (Karpas 1106) with complex
translocation involving 18q21.3 but lacking BCL2 rearrangement and
expression
E Nacheva, MJ Dyer, C Metivier, D Jadayel, G Stranks, R Morilla, JM Heward, T Holloway, S
O'Connor and PC Bevan
Updated information and services can be found at:
http://www.bloodjournal.org/content/84/10/3422.full.html
Articles on similar topics can be found in the following Blood collections
Information about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requests
Information about ordering reprints may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprints
Information about subscriptions and ASH membership may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtml
Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.
Copyright 2011 by The American Society of Hematology; all rights reserved.
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
B-Cell Non-Hodgkin's Lymphoma Cell Line (Karpas 1106) With Complex
Translocation Involving 18q21.3 but Lacking BCL2 Rearrangement
and Expression
By E. Nacheva, M.J.S. Dyer, C. Metivier, D. Jadayel, G. Stranks, R. Morilla, J.M. Heward, T. Holloway, S. O'Connor,
P.C. Bevan, C.-J. Larsen, and A. Karpas
A B-cell non-Hodgkin's lymphoma (B-NHL) cell line (Karpas
1106) with anunusual three-way translocationinvolving
18q21.3 has been derived from a patient with mediastinal
lymphoblastic B-NHL. Although conventional cytogenetics
showed a derivative 18q- identical
to that seen in cases with
t(l4;18)(q32.3;q21.3), no translocationsofeitherchromosome 14 could be detected. Instead fluorescent in situ hybridization analysis using a chromosome-l8 paint showed
that the segment 18q21.3-18qter had become sandwiched
on a derivative chromosome Xbetween segments Xqter-cXq28 and 13q12-qter, with the centromeric site of 18q21.3
subband juxtaposed to the X sequences. Pulsed-field DNA
blots failed to detect rearrangement of the BCL2 gene. Conventional DNA blots using a variety of restriction digests
and both 5' and 3' BCL2 and FVT l probes also failed to
detectrearrangement in Karpas1106.Arearrangedfragment seen only in Hindlll digests with 5' BCLZ probes may
represent a local microalteration, which is eithermutation
a
or small deletion involving the Hindlll site as seen in other
cases of B-NHL.Neither BCLZ RNA nor BCLZ protein expression were detected. These and other data suggest
that genes
at 18q21.3, other than BCLZ and M 1 , may be targets for
translocation in certain subgroups of B-NHL.
0 1994 by The American Society of Hematology.
C
rncr or vcr, and other mechanisms such as the
ability to form
Z-DNAhavebeen
proposed toexplain the clustering of
breakpointstothe vcr." Therefore,the different types of
BCL2 translocations seen in the different subtypes of B-cell
malignancy may represent the action of different pathogenic
mechanisms.
Recently, translocationsinvolving18q21.3havebeen
identified that appear to deregulate the expression of genes
other
than
BCL2. A case of follicular
B-NHL
with
t(2;8)(pl l;q21.3) was shown to involve the IgK locus
with
a gene termed FVTI on 18q21.3 that lies immediately telomeric of BCL2.I3 Another translocation, which again does
not involve BCL2 directly, is the t( 11; 18)(q21 ;q21.3), seen
in lymphocytic and mucosa-associated lymphoid tissue BNHL.14"h The involvement
of the MLL gene at chromosome
1lq23 in a similar translocation (t( 1 1; 18)(q23;q2 1.3)) has
also beendescribed, although whetherthis case involved
BCLZ was not reported." These data indicate that in certain
specific subgroups of B-cell disease, genes at 18q21.3 other
than BCL2 may be activated by translocations.
We report here the preliminary characterization of an unusual B-NHL cell line derived froma patient with mediastinal lymphoblastic B-NHL with chromosomal translocation
involving 18q21.3, but with no rearrangement of the BCL2
gene and noBCL2 expression. Fluorescent in situ hybridization (FISH) analysis showed that the segment 18q21.3-qter
had become sandwiched between fractions of chromosome
X and 13. This cell line mayallow the isolationof other
genes at 18q21.3 of importance in the pathogenesis of some
B-NHL.
HROMOSOMAL translocation t( 14; 1Q(q32.3; q21.3)
was first identified in follicular B-cell non-Hodgkin's
lymphoma (B-NHL) in 1976 by Fukuhara and colleagues.'
Identical translocations were subsequently detected in about
30% of diffuse B-NHL' and rarely in acute leukemias of
mature B cells3 and B-cell chronic lymphoid leukemias (BCLL).4 Molecularcloningstudieshaveshown
that the
t(14;18)(q32.3;q21.3)translocation
juxtaposesthe BCL2
gene with the Ig JH segments.'" In B-NHL, more than 90%
of breakpoints in the BCL2 gene cluster to two regions
of the
3' portion of the gene, which are termed the major
breakpoint
region (mbr) and the minor cluster
region (mcr).' A different
breakpoint cluster region lying between the mbr and
rncr
may be preferentially involved in rare cases of B-CLL with
t(14; 18)(932.3;q21.3).' In addition, BCL2 translocations involving the 5' region of the BCL2 within the variant cluster
region (vcr) with the Iglight-chain genes have been observed
preferentially in B-CLL, but also infollicular B-"L.'"'
Translocations involving the mbr may be promoted by the
presence of recombinogenic chi-like sequences withinthe
rnbr." However, no such sequences have been shown in the
From the Department of Haematology, University of Cambridge,
MRC Centre, Cambridge, UK;the Academic Departmentof Haematology and Cytogenetics, Institute of Cancer Research, Royal Marsden Hospital, Sutton, UK; and Unite'de gknitique cellulaireet mole"
culairedes IeucLrnies, lnstirut deGinitiqueMoliculaire,Paris,
France.
Submitted February 16, 1994; accepted July 6, 1994.
Supported by the Kay Kendall Leukaemia Trust (fo E.N. and
DJ.),
the Leukaemia Research Fund (to M.J.S.D.)and the Cancer
Research Campaign (to G.S.).
The cell line Karpas 1106 may be obtained by written request to
A. Karpas.
Address reprint requests to A Karpas, DSc, Department of Haematology, University of Cambridge, MRC Centre, Hills Rd, Cambridge CB2 2QH, UK.
The QubfiCatiOncosts of this article were defrayedin part by page
chargepayment. This article must therefore be hereby marked
"advertisement" in accordance with 18 U.S.C. section I734 solely to
indicate this fact.
0 1994 by The American Society of Hematology.
0006-4971/94/8410-0108$3.00/0
3422
MATERIALS AND METHODS
Patient Material, Cell Culture, and Cytogenetics
The cell line was derived from a 23-year-old lady with no significant previous medical history who presented in September 1983 with
mediastinal lymphoblastic B-NHL. She was treated with intravenous
combination chemotherapy (cyclophosphamide, hydroxydaunorubicin, vincristine, and prednisolone) and initially remitted, but relapsed in May 1984 with widespread disease. Despite further therapeutic attempts, she died 4 months later with progressive
chemotherapy-resistant B-NHL including massive ascites, bilateral
pleural effusions, and CNS disease. Cell lines were established from
Blood, Vol 84, No 10 (November 15).
1994:
pp 3422-3428
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
B-CELL LINE WITH 18q21.3 LACKING BCL2
3423
1kb
A) FVT-l
c-
3MOkb
CI
I
E
L
E
l
I
B
E
I
35R6
Fig l. Schema of both 5’ and
3’ regions of the B C U and M 1
gene involved in chromosomal
translocations in B-cell malignancies and positionof
DNA
probes used in this study. The 5‘
and 3‘ B C U regions are separated by an intron of a250 kb.
E denotes EcoRl sites; H, Hindlll
sites; B, BernHl sites. H+ represents Hindlll siteloston
one
B C U allele in Karpas 1106. The
map linking mbr and mcr
has
been taken with permission
from Kiem et
l
E
7161
B) 5’BCU
140CA
VCR
BCLZ
5‘
I
1kb
J
”
p H 6 0 4 pxX1-9
pB16
2 kb
c] 3 ’ B C U
c
(
pFLl
PMW-1
both pleural andasciticeffusions as previouslydescribed’*andhavewasperformedassisted
by computer-basedimage-analysissystem
(Smartcapture, Digital Scientific, Cambridge, UK). The ISCN 1991
beenmaintained in intermittentculturesinceOctober 1984. No patient material exists to comparedirectlywiththiscellline,butthenomenclaturewasusedtodescribetheabnormalchromosome
cytogeneticpatternobserved48hoursafterculturinghasremainedmarkers.
as described.” Briefly,
constant. Cytogenetic analysis was performed
cells were synchronized with ethidium bromide and harvested
24
Fluorescent In Situ Hybridization (FISH)
4 weeks
and 28 hours after the initiation of culture and at intervals
of
FISH wasperformedwithchromosomepaints(Cambio,Camthereafter,usingroutinemethods.Chromosomepreparationswere
UK) following the manufacturer’s protocol. In brief, chromoG banded with Wright’s stain, and high-resolution banding analysis bridge,
c
-
4
der(3)
9
8
7
l!‘
i(9p)
10
c
16
1 5 del(l5)
~
P P
m @ * ,a
0
-
0
20 del(20)
11
12 ins(l2)
i(:e
$8-
14 del(14)
5
4
3
2 del(2)
*
-
Fig 2.
c
*
1106P with the following karyotype is shown: 49,X.del (2)
p25.1).
(p11.2p13.3).
+i(9p),
der ins(l2;))
(3)t(2;3) (p13.3;
(q13.1
a21 i(Xp) 22
der(l8)
A metaphase c e l l p r e
X
L
der(X)
q13.3). del (14) (q11.2q13.1), del
t
(15) (q11.2q15.31,
1181
der
IX;13;18)
(q28;q12.l;q21.3),
-20,
del (20)(q13.lq13.31 x2, der (X)
(X;13;18) (q28;q12.lq21.3), +i(Xp).
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
NACHEVA ET AL
3424
Fig 3.
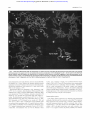
Dual-color FISH analysis with (A) chromosome 18 (in green) and X (in red) paints, (B) chromosome 18 (in green) and 13 (in red) and
(c)chromosome 13 (in green) and X (in red) confimed the three-way translocationt(X;13;18)(q28;q21.3;q12.1). Arrows point to the der(X) and
der(l81 markers. Note the presence of material from X chromosome at the long arm of der(l8). suggesting a two-step mechanism for the
marker formation : firstly, t (X;lSl followed by rearrangementwith chromosome 13. Therefore,the 18q segment (arrow) is sandwichedbetween
chromosome X and 13 segments with the 18q21.3 subband juxtaposed to Xq28 and 18q23 to 13q12.1, repectively (D).
some preparations were made after the cells were treated with BrdU
(20 mg/mL) for 17 hours followed by ethidium bromidelcolcemid
incubation for the last 90 minutes before harvest. Before denaturing,
chromosome preparations were treated in 2X saline sodium citrate
(SSC) for 60 minutes at 37°C.
Chromosomal DNA was denatured in 70% formamidel2xSSC
at 75°C for 2 to 4 minutes, then dehydrated. Probc (paint) DNA plus
unlabeled Cot-l DNA (S mglmL; GlBCO RRL. Gaithersburg. MD)
were denatured at 75°C for 10 minutes. allowed to prcanncal for 90
minutes at 37°C and laid onto prewarmed slides under sealed coverslips. The hybridization was performed at 42°C for 12 to IS hours.
After hybridization. the slides were washed three times on 2X SSC.
SO% formamide at42°C followed hy two washes in 2X SSC and
once on 0. I X SSC at 42°C. Visualization of the hybridized probe
was achieved after incubation with ( I ) S mglmL fluorescein isothiocyanate (FITC)-avidinDCS. (Vector Laboratories. Burlingame. CA).
(2) S mglmL biotinylated goat antiavidin (Vector) and (3) FITCavidin DCS. All incubations were at37°C for 30 minutes. Three
washes. each 3 minutes in length, were performed between layers
using 4 x SSC, 0.05% Tween 20. Chromosomes were counterstained
with DAPl dissolved in antifading solution (Citifluor, Canterbury,
U K ) at a final concentration of 80 mglmL. Images were captured
with CCD Camera (Photometrics. Tuscon. AZ) aided by dedicated
software (Smartcapture. Digital Scientific. Cambridge. UK). The Gbanded appearance of the counterstained chromosomes was obtained
by image processing using the same program.
Inlmrtrtr~phenor?pirlR
Cell surface antigen expression was detected using monoclonal
antibodies (MoAhs) and flow cytometry as described.’” Cytoplasmic
RCL2 expression was assessed using a cocktail of RCL2 MoAbs”
andboth immunocytochemistry and flow cytometry after cell permeahilization.”.” Cell lines Karpas 422 and DoHH2, whichboth
exhihit a RCL2 mbr-IgJ,, fusion gene were used as positive controls.’.‘.’’As a negative control, the primary antibody was omitted.
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
3425
B-CELL LINE WITH 18~121.3LACKING 6CL2
Table 1. lmmunophenotype of B-NHL Cell Line, Karpas 1106
Antigen
Expression
CD5
CD10
CD19
CD22
CD23
CD37
FMC7
-
+
++
-
++
+
-
IgM
-
I9D
IgG
++
-
IgA
++
-
BCL2
lmmunophenotypic analysis was performed by flow cytometry for
both cell surface and cytoplasmic BCL2 antigen.
Abbreviations: +,weak positive expression; ++, strong expression;
-, no detectable expression.
DNA and RNA Studies
mosomal constitution (Fig 2): 49,X,de1(2)(pl1.2~13.3),der
(3) t (2;3) (p13.3;p25.1), +i9(p), ins(l2;?) (q13.lq13.3), del
(14)(q11.2q13.1), del (15)q11.2q15.3), der(l8) t (X;13;
18)(q28;q21.3;qlZ.I), -20, del(2O)(q13.1q13.3) x 2, der
(X)t(X; 13; 18) (q28;q21.3;q12.1), +iX(p).
To confirm this, dual-color FISH analysis with a chromosome 18 (in green) and X (in red) paints was performed (Fig
3). This showed that the 18q21.3-qter segment was retained
in thegenome as part of the der(X) marker,sandwiched
between segments Xqter-c-Xq28 and 13q12.1-qter (Fig 3D,
inset). The presence of material from chromosome X at the
telomeric endof der(l8) marker (see Fig3A) is suggestive of
a two-step mechanism of marker formation. Firstly t(X; 18)
(q28;q21.3) followed by rearrangement between the der(X)
and 13. This interpretation is in agreement with the results
of the dual-painting experiments with chromosome 18 and
13 (Fig 3B) as well as X and 13 (Fig 3C).
Chromosomes 14, 15, and 20 were found to have a partially deleted long arm.FISH analysis with relevant chromosome paints showed the presence of only two signals, thus
suggesting that these deletions were interstitial. Painting with
chromosome 12 probe showed uniform signal on
the der( 12)
chromosome, and this finding suggests that the inserted rnaterial belongs to chromosome 12. The t(2;3) was also confirmed with dual-color FISH chromosome painting.
Conventional DNA blotting. Conventional DNA blotting was
performed as described?6 High molecular-weight DNA from Karpas
1106 and normal peripheral blood mononuclear cells was digested
to completion with the following restriction endonucleases: EcoRI,
HindIII, BomHI, Psr I, BgEII, EcoRV, and Kpn I (Promega, Madison
Irnmunophenotypic Analysis
W .
Pulsed-jeld DNA blots (PFGE). Pulsed-field DNA blots were
Both Karpas 1106A and 1106P had the phenotype of maperformed as described.*’ DNAblocks were digested with restriction
ture, class-switched IgG expressing B-cells as shown in Taendonuclease Nor I which yields a 650-kb germline BCL2 fragble l. There wereno differences betweenthe1106Aand
ment.”
P sublines. BCLZ expression was sought on three separate
RNA studies. Nothern blots were peformed using poly (Af)
occasions
in two laboratories by both flow cytometry and
mRNA as described.” Quantitation of the amount of mRNA applied
immunocytochemistry; no BCL2 expressionwasdetected
and assessment of mRNA integrity were assessed by probing with
under conditions when both Karpas 422 and DoHH2 exhiba glyceraldehyde-3-phosphate dehydrogenase cDNA probe.
ited strong staining (Fig 4).
DNA probes. All DNA probes were used as gel-purified inserts
and were labeled with 3ZP-deoxycytidinetriphosphate (dCTP) to a
specific activity of 2 2 X lo9 disintegrations per minute (dpm)/pg
DNA and RNA Studies
DNA by the method of oligopriming. Derivation of the probes used
Rearrangements of both BCL2 and FVTl were sought in
in this study may be found in the following references: FVTI: probes
K q a s 1106. Results are summarized in Table 2. In pulsedfor the FVTl locus were 1 4 0 CA, 7/61, and 35R6. Probe 140 CA
is a 0.4-kb Kpn I FVTl 3’ cDNA fragment, whereas 7/61 and 35R6
field blocks digested with Not I, no BCL2 rearrangements
are genomic clones lying immediately 5’ and 30 to 40 kb 5’ of the
couldbedetectedusing
3‘ BCLZ probe pMWl (data not
F V T l gene, respectively. 7/61 is a 0.8-kb EcoRI-Xbu I fragment
shown). To confirm these data, and to extend the area anaand 35R6 a 0.8-kb HindIII fragmentI3 (and R. Rimokh. personal
lyzed, rearrangementsinKarpas1106weresoughtusing
communication, November 1993); 5’ BCLZ: two genomic clones
both BCL2 and FVTI probes in a wide variety of restriction
lying 5’ ofthe BCL2 coding sequences were used:pHB 0.6, a 0.6-kb
digests
by conventional DNA blot. No rearrangements were
HindIII-BamHI fragment” and pXX1.9, a 1.9-kb Xho I f~agment.’~
detected
using 3’ BCL2 and FVTI probes. Additionally, no
Additionally, a 1.6-kb cDNA clone, pB16, encoding part of the first
rearrangements were detected using the 5’ BCL2 probes exand second exons of BCLZ, was also used”; 3’ BCLZ (mbr): clone
cept in HindIII digests (Fig 5). The size of the rearranged
pm-l, a genomic fragment spanning the mb3; 3‘ BCL2 (mcr): clone
pm-2, a genomic fragment spanning the minor cluster region”; 3‘
HindIII fragment is consistent with a local microalteration
BCL2 (mbr-mcr): clone pMW-I, a 1.3-kb EcoRI-HindlII genomic
at the 5’ end of the BCL2 gene of one allele in the vicinity
fragment lying between the mbr and met?* The relationship of these
of the 5’ HindIII site {Fig 1) resulting in the loss of that
F W I and BCL.2 probes is shown in Fig 1. References for the derivaHindnl site. Furthermore,these data indicate that the cytogetion of immunoglobulin (Ig) probes may be found in reference 26.
RESULTS
Cytogenetic and FISH Studies
A mainline cell clone was seen in all analyzed cells of
Karpas 1106P, which was found to have the following chro-
netically observed breakpoint at 18q21.3
fell outside both
BCL2 and FVTI loci.
Studies with probes from the IG loci showed that both
Karpas1106Aand
1106P sublines had identical biallelic
IgJHrearrangements, indicative of a common clonal origin.
Both alleles of IgClc were deleted, with biallelic IgCh re-
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
3426
NACHEVA
7
K1106
AL
CELL LINE
1l
8
f
]PEG CON
DOHH2
CELL LINE
Fig 4. Flow cytometric profiles of B-NHL cell lines Karpas 1106 and DoHH2 stained with a cocktail of B C U MoAbs after permeabilization
ofthe cell
Abscissa, fluorescent intensity; Ordinate, logcell number. DoHH2 showslevelofstaining
comparable withthat seen
in other B-cell lines with t(14;18)(q32.3;21.3), whereas only background staining was seen in Karpas 1106. Identical results were obtained
reproducibly in two of our laboratories and were confirmed by immunocytochemistry.
arrangement, consistent with the observed Ig light-chain expression.
No BCL2 RNA expression could be detected in Northern
blots using 2 p,g of poly(A') RNA (Fig 6).
DISCUSSION
The cell line Karpas I106 is of interest for two reasons.
Firstly, although it was thought initially that BCL2 expression was a specific marker of the t(14; 18)(q32.3;q21.3)
translocation, it has become clear that expression of this gene
is, in fact, widespread, particularly in tissues characterized
by programmed cell death or apopto~is.~
Most follicular and
diffuse B-NHL and B-CLL express abundant BCLZ irrespective of whether they carry the t( 14; 18)(q32.3;q21.3) or
not.?2.33-37
Also, levels of BCLZ comparable with those seen
in B-NHL with t( 14; 18)(q32.3;q21.3) maybe seen in the
breast carcinoma cell line MCF-7 with no detectable genomic alteration of the BCL2 gene.3x,3q
Therefore, it is surprising that the Karpas I106 cell line should lack expression of
BCLZ. To our knowledge, the only other B-NHL cell line
with an 18q21.3 translocation that fails to express BCL2 is
Hind 111 BamHI
C T C T C T
Table 2. GenotvDe of B-NHL Cell Line, Karoas 1106
Probe
Configuration
IgJH
19CP
lgCK
lgCX
5' BCL2
mbr
mcr
oMWl
RIR
DID
DID
RIR
GIG'
GIG
GIG
GIG
K~~~~~ 1 1 0 6 ~and 1 1 0 6 ~showed identicalbiallelic
- 1.9kb
Probe pB 1G
JH re-
arrangements. Otherwise, both Cp and CK were biallelically deleted
with the
IgG expression, No rearrangement of
BCL2 was Seen by PFGE in a Not I digest when probed with pMW1.
Abbreviations: G. germline configuration; R. rearranged configuration; D, deleted DNA sequences.
Rearrangement of 5' BCL2seen in Hindlll digests only, which was
considered to represent a local microalteration (see text).
Fig 5. Southern blotof Karpas 1106 DNA. The blotwashybridized
with 5'
probe pB1'.% c denotes control
and denotes
tumor DNA. Germline configuration of Karpas 1106 DNA was observed in all digests with the exception of Hidlll, in which clear
rearrangement was Seen with probes pB16, and pxxl.s. The rethanthe germline,
arrangedfragment inK~~~~~1106 is-2.5 kb larger
and therefore, from the restridion map
of this region it seems likely
that the rearrangement represents a local microalteration resulting
in the loss of the Himllll sitedenotedbyan
asterisk in Fig 1.
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
B-CELLLINE WITH 18q21.3 LACKING BCLZ
PROBE BCL2 IDFLI)
Track:
3421
A
ACKNOWLEDGMENT
B
-W
< 9.5kb
< 4.4
< 1.3
PROBE: GAPDH
Fig 6. Northernblot of Karpas 1106 RNA. Poly (A') RNA from
Karpas 422 (track AI and Karpas 1106 (track B) cell lines was blotted
and probed with BCL2 probe pFL-1 (upper panel] and a glyceraldehyde-3-phosphate dehydrogenase (GAPDM probe (lower panel]. No
BCLZ mRNA expression was detected in Karpas 1106.
SU-DULS, in which one allele of BCL2 has been disrupted
in a three-way translocation with MYC and lgJl+3' Other BNHL lines that do not express BCL2 (eg, SU-DHL9) do not
exhibit I8q2 I .3 translocations."" Our examination of other
B-NHL lines that have 18q21.3 translocations but lack BCL2
rearrangement on DNA blot has shown abundant BCL2 expression in all three cases (M.J.S. Dyer, unpublished observations, December 1993). In Karpas 1106, it is possible that
microalterations (either mutations or small deletions) at the
S' end of the BCL2 gene (as detected by the rearrangement
seen in Hind111 digests alone) may have inactivated the promoter of one allele, whereas the other promoter maybe
physiologically repressed. Such microalterations in the 5'
region of the BCL2 gene have been previously reported both
in B-NHL
and
in fresh patient
In one
case of follicular B-NHL, it was shown that these alterations
reflect small deletions of the S' region of BCLZ.".' Similar
deletions havebeen observed in the B-NHL cell line SUDHL6; these deletions involved the major sites of initiation
of BCL2 tran~cription.~'
Secondly, despite lacking BCL2 expression, Karpas 1106
nevertheless has a translocation involving 18q2 I .3, although
this does not appear to involve the BCL2 gene directly. These
and other data""' suggest that genes at I8q21.3, other than
BCL2, may have a role in the pathogenesis of certain subsets
of B-NHL. Therefore, the Karpas 1106 cell line may allow
the study of mechanisms of B-cell neoplasia independent of
BCL2 expression.
We thank Drs M.L. Cleary(Stanford University. CA) and Y.
Tsujimoto(OsakaUniversity,Japan)for
kindly providing RCLZ
probes, Dr R. Rimokh (Lyon. France) forkindly providing the FVTl
probes.and Dr T.H.Rabbitts(Laboratory
o f Molecular Biology.
Cambridge. UK) and the Human Genome MappingProject Resource
Centre (Harrow, UK) for kindly providing If probes.
REFERENCES
I . Fukuhara S, Rowley JD. Variakojis D. Golomb HM: Chromosome abnormalities in poorly differentiated lymphocytic lymphoma.
Cancer Res 3 9 3 I 19. 1979
2. Lee MS, Blick MB, Pathak S, Trujillo JM. Butler JJ. Katc RL.
McLaughlin P, Hagermeister FB. Velasqucz WS. Goodacre A. Cork
A, Gutterman JU. Cabanillas F The gene located at chromosome
I8 band q21 is rearranged in uncultured diffuse lymphomas as well
as follicular lymphomas. Blood 70:90. 1987
3. Mufti GJ, Hamblin TJ, Oscicr DC, Johnson S: Common ALL
with pre-B-cell features showing (8: 14) and (14: 18) chromosome
translocations. Blood 62:l 146. 1983
4. Jonveaux P. Hillion J. Bennaccur AI. D'Agay MF. Brice P.
Daniel M-T, Sigaux F. Berger R: t( 14: 18) and HCL2 rearrangement
in a B-chronic lymphocytic leukaemia. Br J Haematol 8 I :620. 1992
5. Cleary ML. Sklar J: Nucleotide sequenceof a t( 14: 18) chromoa
somebreakpoint in follicularlymphomaanddemonstrationof
breakpoint clustering region near a transcriptionally active locus on
chromosome 18. Proc Natl Acad Sci USA 82:7439. 1985
6 . Tsujimoto Y. Croce CM: Analysisof the structure oftranscripts
and protein products ofRCLZ. the gene involvedin human follicular
lymphoma. Proc Natl Acad Sci USA 835714. 1986
7. Korsmeyer SJ: BCLZ initiates a new category of oncogenes:
Regulators of cell death. Blood 80:879. 1992
8. Dyer MJS. Zani VJ. Lu WZ. O'Byrne A. Mould A. Chapman
R. Heward JM. Kayano H, Jadayel D. Matutes E. Catovsky D. Oscier
DG:BCL2translocations
in leukemias of mature B-cells. Blood
833682, 1994
9.Adachi M. Tefferi A. Greipp PR. Kipps TJ.Tsujimoto Y:
Prcferential linkage of RCL2 to immunoglobulin light chain genes
in chronic lymphocytic leukaemia. J Exp Med l7 1:SSY. 1990
10. Bertheas M-F, Bachy M, Magaud J-P. Rimokh R. Vasselon
C. Berger F, Oriol PC. Jaubert J. Raynaud J. Brizard CP. Guyotat
D: t(2; 18) and t(18:22) variant chromosomal translocations inBcell malignancies. Leuk Lymphoma 8:197. 1992
I I . Wyatt RT. Rudders RA, Zelenetz A. Delellis RA. Krontiris
TG: RCL2 oncogene translocation is mediated by a c-like consensus.
J Exp Med 175:1575, 1992
12. Adachi M. Tsujimoto Y: Potential Z-DNA elements surround
the breakpoints of chromosome translocation within the S' flanking
region of the BCL2 gene. Oncogene 5:1653, 1990
13. Rimokh R. Gadoux M. Bertheas M-F. Berger F. Garoscio
M. Deleage G, Germain D. MagaudJ-P: FVT-I. a novel human
transcription unit affected by variant translocation t(2: 18)(pl I:q21)
of follicular lymphoma. Blood 8 1:136. I 993
14. GriffinCA,ZehnbauerBA,Beschorner
WE. Amhinder R.
Mann R: t( 1 I ; 18) (q21 ;q21) is a recurrent chromosome abnormality
in small lymphocytic lymphoma. Genes Chromosom Cancer 4: 153.
1992
15. Horsman D, Gascoyne R. Klasa R. Coupland R: t ( I I : 18)
(q2 l:q21): A recurring translocationin lymphomas of mucosa-associated lymphoid tissue (MALT). Genes Chromosom Cancer 4: 183.
1992
16. Leroux D. S i i t i P. Hillion J, Le Marc-Hadour F. PCgouriiBandelier B. Jacob M-C. Larsen C-J. Sotto J-J: t( I I : 18)(q21:qZI )
From www.bloodjournal.org by guest on November 7, 2014. For personal use only.
3428
may delineate a spectnm of diffuse small B-cell lymphoma with
extranodal involvement. Genes Chromosom Cancer 734, 1993
17. Thirman MJ, Gill HJ, Burnett RC, Mbangkollo D, McCabe
NR, Kobayashi H, Zemin-van der Poel S, Kaneko Y, Morgan R,
Sandberg AA, Chaganti RSK, Larsen RA, Le Beau MM, Diaz MO,
Rowley JD: Rearrangement of the MLL gene in acute lymphoblastic
and acute myeloid leukaemias with 1lq23 chromosomal translocations. N Engl J Med 329:909, 1993
18. Karpas A, Hayhoe FGJ, Greenberger J, Barker CR, Cawley
JC, Lowenthal RH, Mahoney WC: The establishment, cytology,
cytochemistry and immunological characterisation of human haemic
cell lines: Evidence for heterogeneity. Leuk Res 1:35, 1977
19. Nacheva E, Dyer MJS, Fischer P, Stranks G, HewardJM,
Marcus RE, Grace C, Karpas A: C-MYC translocations in de novo
B-cell lineage acute leukaemias with t(14; 18) (cell lines Karpas 231
and 353). Blood 82:231, 1993
20. Buccheri V, Matutes E, Dyer MJS, Catovsky D: Lineage
commitment in biphenotypic acute leukemia. Leukemia 7:919, 1993
21.Liu Y-G, Mason DY, Johnson GD,Abbot S, Gregory CD,
Hardie DL, Gordon J, MacLennan IH: Germinal centre cells express
bcl-2 protein after activation by signals which prevent their entry
into apoptosis. Eur J Immunol 21:1905, 1991
22. Gaulard P, d’Agay M-F, Peuchmaur M, Brousse N. Gisselbrecht C, Solal-Celigny P, Diebold J, Mason DY: Expression of the
BCLZ product in follicular lymphoma. Am J Pathol 140:1, 1992
23. Sysala MT, Thrikainen M, Jansson S-E, Krusis T: Flow cytometric analysis of terminal deoxynucleotidyl transferase: A simplified method. Am J Clin Pathol 99:298, 1993
24. Dyer MJS, Fischer P, Nacheva E, Labastide W, Karpas A: A
new human B-cell non-Hodgkin’s lymphoma cell line (Karpas 422)
exhibiting both t(14; 18) and t(4; 11) translocations. Blood 75:709,
1990
25. Kluin-Nelemans HC, Limpens J, Meerabux J, Beverstock GC,
Jansen JH, de Jong D, Kluin PM: A new non-Hodgkin’s B-cell
line (DoHH2) with a chromosomal translocation t(14; 18)(q32;q21).
Leukemia 5:221, 1991
26. Dyer MJS, Heward JM, Zani VJ, Buccheri V, Catovsky D:
Unusual deletions within the immunoglobulin heavy-chain locus in
acute leukemias. Blood 82:865, 1993
27. Jadayel D, Matutes E, Dyer MJS, Brito-Babapulle V, Khokar
MT, Oscier DG, Catovsky D: Splenic lymphoma with villous lymphocytes: Rearrangments of the BCL-I locus and cyclin D1 expression. Blood 83:3664, 1994
28. Zelenetz AD, Chu G, Galili N, Bangs CD, Homing SJ, DonIon TA, Cleary ML, LevyR: Enhanced detection of the t(14;18)
translocation in malignant lymphoma using pulsed-field gel electrophoresis. Blood 78: 1552, 1991
29. Stitt P, Leroux D, Hillion J, Monteil M, Berger R, MathieuMahul D, Larsen C-J: Molecular analysis of a variant translocation
in a case of lymphocytic lymphoma. Genes Chromosom Cancer
6:39. 1993
NACHEVA ET AL
30. Tsujimoto Y, Bashir MM, Givol 1, Cossman J , Jaffe E, Croce
CM: DNA rearrangements in human follicular lymphomacan involve the 5’ and 3’ region ofthe BCLZ gene. Proc Natl Acad Sci
USA 84:1329, 1987
31. Cleary ML, Galili N, Sklar J: Detection of a second t( 14; 18)
breakpoint cluster region in humanfollicular lymphomas. J Exp Med
164:315, 1986
32. Kiem H-P, Nourse J, Saltman DL, Blume KG, Cleary ML:
Concurrent activation of MYC and inactivation of BCLZ by chromosomal translocation in a lymphoblastic lymphoma cell line. Oncogene 5:1815, 1990
33. Hanada M, Delia D, hello A, Stadthauer E, Reed JC: BCLZ
gene hypomethylation and high-level expression in B-cell chronic
lymphocytic leukemia. Blood 82:1820, 1993
34. Zutter M,HockenbemyD,
Silverman GA, Korsmeyer SJ:
Immunolocalization of the bcl-2 protein within hematopoietic neoplasms. Blood 78:1062, 1991
35. Stitt P, Hillion J, d’Abay M-F, Gaulard P, Cazals D, Badoux
F, Berger R, Larsen C-J: BCLZ gene activation and protein expression in follicular lymphoma: A report on 64 cases. Leukemia 7:410,
1993
36. Achena M, Larson L-G, Gottardi D, Gaidano G, Carlsson M,
NilssonK, Caligaris-Cappio F Growth and differentiation-associated expression of bcl-2 in B-chronic lymphocytic leukemia cells.
Blood 79:2981, 1992
37. Pezzella F, Tse AGD, Cordell JL, Pulford KAF, Gatter KC,
Mason DY: Expression of the bcl-2 oncogene protein is not specific
for the 14; 18 chromosomal translocation. Am J Pathol 137:225,
1990
38. Monaghan P, Robertson D, Amos TAS, Dyer MJS, Mason
DY, Greaves MF: Ultrastructural localization of the BCLZ protein.
J Histochem Cytochem 40:1819, 1992
39. Gusterson BA, Nathan B, Jadayel D, O’Hare M, Anbazhagan
R, Jayatilake H, Ebbs S, Micklem K, Price K, Gelber R, Goldhirsch
A, Dyer M: Expression of the proto oncogene BCL2 inprimary
breast cancer and its correlation with tumour phentoype. Ann Oncol
5:409, 1994
40. Young RL, Korsmeyer SJ: A negative regulatory element in
the BCL2 5‘ untranslated region inhibits expression from anupstream promoter. Mol Cell Biol 13:3686, 1993
41. Seto M, Jaeger U, Hockett RD, Graninger W, Bennett S,
Goldman P, Korsmeyer SJ: Alternative promoters and exons, somatic mutationand deregulation of the BCL-2-Ig fusion gene in
lymphoma. EMBO J 7:123, 1988
42. Mikraki V, Ladanyi M, Chaganti RSK. Structural alterations
in the 5’ region ofthe BCLZ gene in follicular lymphomas with
BCLZ-MER or BCLZ-MCR rearrangements. Genes Chromosom Cancer 3:117, 1991
43. Stitt P, Hillion J, d’Agay M-F, Berger R, Larsen C-J: BCL2
complex rearrangement in follicular lymphoma: Translocation mbr/
JH and deletion in the vcr region of the same BCL2 allele. Oncogene
8:3073, 1993