Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
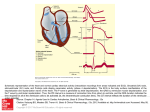
Sympathetic innervation modulates repolarizing K1 currents in rat epicardial myocytes QIN-YUE LIU,1 MICHAEL R. ROSEN,1 DAVID MCKINNON,2 AND RICHARD B. ROBINSON1 of Pharmacology and Pediatrics, College of Physicians and Surgeons of Columbia University, New York 10032; and 2Department of Neurobiology and Behavior, State University of New York at Stony Brook, Stony Brook, New York 11794-8661 1Departments transient outward potassium current; inwardly rectifying potassium current; nerve growth factor; potassium ion of sympathetic innervation occurs concurrently with a decrease in action potential duration (APD) in rat and dog ventricular muscle (32). We have previously demonstrated that daily injection of newborn rats for 10 days with nerve growth factor (NGF) or NGF antibody (Ab), respectively, accelerates or delays cardiac sympathetic innervation (29). In this earlier study, alteration in the time course of cardiac sympathetic innervation was confirmed using tissue catecholamine determinations, tyrosine hydroxylase assays, and the release of norepinephrine by tyramine infusion. With this method, we previously found that acceleration of the time course of innervation consistently shortens the electrocardiographic Q-T interval and ventricular APD (29, 36). K1 carries the major outward currents that determine repolarization, thereby controlling the duration of the action potential. The progressive shortening of APD POSTNATAL EVOLUTION in rat heart during postnatal development (26) has been ascribed to evolution of the transient outward K1 current (Ito ) and the inwardly rectifying K1 current (IK1; see Refs. 37 and 38). In addition, K1 currents are important targets for the actions of many neurohumors and intracellular second messengers that modulate repolarization (18). Therefore, we used protocols for sympathetic modulation similar to those employed previously (29) to determine the relationship between sympathetic innervation and the electrocardiogram and action potential and the K1 currents Ito and IK1 in rat epicardial myocytes. METHODS Treatment of rats. The method of injection with NGF, placebo, or Ab was described by Malfatto et al. (29). Littermate Wistar rats born in our animal care facility were randomly separated into three groups (3–4 animals/group per litter). Each rat was injected subcutaneously from days 0 through days 13–15 of life. One group was treated with 1.0 µg/day NGF on days 0–5, 1.5 µg/day on days 6–10, and 2 µg/day on days 11–15. A second group was administered 10 µl/day NGF Ab on days 0–5, 15 µl/day on days 6–10, and 20 µl/day on days 11–15. The third group received normal saline (the solvent for NGF and Ab) as a placebo. All solutions were administered in a final volume of 100 µl. NGF and Ab were purchased from Collaborative Research (Bedford, MA). Electrocardiograms of the animals in the three groups were recorded on days 0, 3, 6, 9, and 11, as reported in previous studies, and manifested comparable results (data not shown; see Refs. 29 and 36). Preparation of myocytes. Epicardial myocytes were isolated on days 13–15 using a method modified after Liu et al. (28) . On the day of study, two to three rats were narcotized with CO2 and decapitated, and the hearts were rapidly excised. Only the apical two-thirds of the ventricles were used to avoid contamination by atrial cells and the great vessels. The ventricles were cut open, and the endocardium and septum were removed. The remaining epicardium was washed three times in a solution (dissociation solution) containing the following (in mM): 110 NaCl, 5.4 KCl, 4 NaHCO3, 1.6 MgCl2 · 6H2O, 1.8 NaH2PO4, 20 N-2-hydroxyethylpiperazineN8-2-ethanesulfonic acid (HEPES), 5 glucose, 4 L-glutamine, and 10 taurine. The epicardium was then chopped into small pieces that were bathed in the dissociation solution, also containing 3 mg/ml trypsin (Sigma Chemical, St. Louis, MO) and 5 mg/ml bovine albumin (Sigma), and placed in a Gyrotory water bath at 37°C for 15 min. The first incubation solution was discarded to remove connective tissue and debris. There were four successive changes of enzyme solution under these conditions, and the tissue was triturated with a Pasteur pipette after each incubation. After the remaining tissue pieces were removed, the enzyme solution was collected and centrifuged. The cells were then resuspended using the dissociation solution containing 5 mg/ml of bovine albumin. All data are based on studies of at least three 0363-6135/98 $5.00 Copyright r 1998 the American Physiological Society H915 Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 Liu, Qin-Yue, Michael R. Rosen, David McKinnon, and Richard B. Robinson. Sympathetic innervation modulates repolarizing K1 currents in rat epicardial myocytes. Am. J. Physiol. 274 (Heart Circ. Physiol. 43): H915–H922, 1998.— During postnatal development, sympathetic innervation of the heart evolves, and repolarization accelerates. Our goal in this study was to test whether sympathetic innervation modulates the ion channels that regulate repolarization. We studied action potentials and repolarizing K1 currents in epicardial myocytes from rats in which sympathetic innervation was accelerated or delayed, respectively, by subcutaneous injection of nerve growth factor (NGF) or NGF antibody (Ab) for the first 15 days of life. A placebo group was included as well. Action potential duration (APD) to 90% repolarization was greater in the Ab (158 6 18 ms)-treated than the NGF (106 6 10 ms)-treated animals (P , 0.05); the APD at 90% repolarization for the placebo group was intermediate (125 6 30 ms). The transient outward (Ito ) and inward rectifier (IK1 ) K1 currents were recorded in freshly dissociated cells using the whole cell patch-clamp technique. Ito was decreased in density at potentials positive to 140 mV in Ab-treated rats when compared with rats treated with NGF (P , 0.05). In addition, the inactivation curve of Ito in Ab-treated rats was shifted 13 mV positive to that of NGFtreated rats. IK1 also decreased in the Ab-treated group compared with the NGF group in the potential ranges of 2100 to 290 mV (P , 0.05). However, the channel transcript abundance (RNA) in NGF-, Ab-, or placebo-treated rat hearts did not differ. Our results suggest that sympathetic innervation contributes to the developmental differences in K1 currents and APD postnatally in the rat. H916 SYMPATHETIC MODULATION OF REPOLARIZING K1 CURRENTS of the prepulse voltage V, V1/2 is the voltage at which 50% of the current is inactivated, and S is the slope factor. This allowed calculation and comparison of the mean midpoint of the inactivation curve (V1/2 ) and the steepness of the curve (S) between the treatment groups. Preparation of RNA. The lower two-thirds of the left and right ventricle after 15 days of treatment with NGF, Ab, or placebo was rapidly dissected out and quick-frozen in liquid N2. Tissue samples were then homogenized in guanidinium thiocyanate. Total RNA was prepared by pelleting the homogenate over a CsCl step gradient. All RNA samples were quantified by spectrophotometric analysis. Ribonculease protection assay. DNA templates for the preparation of RNA probes were prepared as described previously (10, 11). In all cases, a significant amount of nonhybridizing sequence (<50 bp) was included in the probe to allow easy distinction between the probe and the specific protected band. The specificity of the assay was such that there was no evidence for unwanted cross-reaction between any probe and another nonspecific K1 channel transcript. Ribonuclease (RNase) protection assays were performed as described previously (10). For each sample point, 10 µg of total RNA were used in the assay. A probe for the rat cyclophilin gene (9) was included in the hybridization as an internal control to confirm that the sample was not lost or degraded during the assay. Five micrograms of yeast tRNA were used as a negative control to test for the presence of probe self-protection bands. RNA expression was quantified directly from dried RNase protection gels using a PhosphorImager (Molecular Dynamics, Sunnyvale, CA). RESULTS APD and membrane potential measurements. Current-clamp experiments were carried out using dissociated ventricular epicardial myocytes from rats treated with NGF, Ab, or placebo. The cells were stimulated via the patch electrode using suprathreshold constantcurrent pulses at 0.2 Hz. Figure 1 shows action potential records from three individual cells treated, respectively, with Ab, placebo, or NGF. The table in Fig. 1 shows the data from the three sets of cells. This limited number of action potentials was sufficient to confirm consistency with our previously reported results from intact myocardium (29, 36), i.e., hearts from rats treated with Ab have significantly longer APD at 90% repolarization (APD90 ) compared with NGF-treated animals, and results from placebo-treated rats are intermediate. Although the decrease that we report in maximum diastolic potential in Ab-treated rats is not significant with this small n value (Fig. 1, P . 0.05), it is of the same magnitude as that described by us previously in a larger series of animals, in which the change was significant (36). Ito. To test the role of Ito in determining APD in young rats, we studied the effect of the Ito blocker 4-AP (0.5 mM) on APD of myocytes from rats treated with placebo. We limited the concentration used in these action potential studies because we had previously demonstrated the importance of nonspecific effects of high 4-AP doses on APD in a prior study (27). Because 0.5 mM 4-AP may not fully block Ito, these experiments provide a lower limit on the contribution of Ito to APD. Figure 2 demonstrates that 0.5 mM 4-AP reversibly pro- Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 to four separate litters of animals. Only rod-shaped, quiescent cells were used for electrophysiological experiments. The cells from the three treatment groups were morphologically similar at the light microscopic level, although the Ab-treated cells had a smaller mean membrane capacitance (see RESULTS). Recording techniques. Epicardial myocytes were transferred to a 0.5-ml Lucite bath on the stage of an inverted microscope (Nikon Diaphot; Nikon Instrument, Tokyo, Japan). The cells were allowed to settle onto a glass coverslip coated with poly-L-lysine to facilitate adhesion of the cells to the glass. The cells were superfused at a rate of 2 ml/min with Tyrode solution (see below) for action potential recording. The standard gigaohm seal, whole cell recording method (17) was used to measure the action potentials and the ionic currents. The voltage-current clamp circuit was provided by an Axopatch 1C patch-clamp amplifier (Axon Instruments, Foster City, CA). Patch electrodes were made using borosilicate glass capillary tubes and a micropipette puller (model P 80/PC; Sutter Instrument, Novato, CA). The pipettes had a resistance of 3–4 MV. Data were sampled using pCLAMP software (5.5) and a TL1 analog-to-digital interface (Axon Instrument) and were stored on disk using a Dell computer. Data were analyzed using pCLAMP 6.0. The experiments were performed at 31°C. Recording solutions. The action potentials were recorded in Tyrode solution containing (in mM) 131 NaCl, 1.8 NaH2PO4, 18 NaHCO3, 5.5 dextrose, 0.5 MgCl2 · 6H2O, 4 KCl, and 2.7 CaCl2 and were gassed with 95% O2-5% CO2. The recording pipette solution contained (in mM) 120 potassium aspartate, 30 KCl, 1 MgCl2 · 6H2O, 5 MgATP, 10 HEPES, and 5 glucose. Ito was measured in a bath solution (5) containing (in mM) 144 N-methyl-D-glucamine chloride, 5.4 KCl, 1 MgCl2 · 6H2O, 2.5 CaCl2, 10 HEPES, and 0.3 CdCl2. N-methyl-D-glucamine was used to replace Na1 in the bath solution, and CdCl2 was employed to block Ca21 current. The pipettes were filled with a solution containing (in mM) 140 KCl, 1 MgCl2 · 6H2O, 5 ethylene glycol-bis(b-aminoethyl ether)-N,N,N8,N8-tetraacetic acid (EGTA), 5 MgATP, 5 Na2-creatine phosphate, 0.2 GTP, and 10 HEPES. In voltage-clamp experiments, 4-aminopyridine (4-AP) at a concentration of 2 mM was used to define Ito as the 4-AP-sensitive current (5). In the action potential studies, a lower concentration of 0.5 mM was employed to avoid potential nonspecific actions of 4-AP (27). The IK1 measurements were carried out in a bath solution containing (in mM) 140 NaCl, 5.4 KCl, 2 MgCl2 · 6H2O, 10 glucose, 10 HEPES, 1.8 CaCl2, and 0.3 CdCl2. In some experiments, BaCl2 (2 mM) was dissolved in this solution to block IK1. The composition of pipette solution for IK1 was as follows (in mM): 120 potassium aspartate, 30 KCl, 1 MgCl2 · 6H2O, 5 MgATP, 11 EGTA, 1 CaCl2, and 10 HEPES. All solutions were filtered (0.22 µm) before use. pH was adjusted to 7.2 with KOH for pipette solutions and 7.4 for bath solutions with HCl. None of the data have been corrected for liquid junction potential, which was determined to be 8 6 0.5 mV (n 5 4) and 4 6 0.4 mV (n 5 4) for IK1 and Ito solutions, respectively. Data analysis and curve fitting. Ito and IK1 were expressed as current density. Curves were compared using nested analysis of variance unless otherwise indicated in the text, and individual data points were subsequently compared using a Bonferroni t-test for multiple comparisons. The significance level was determined at P , 0.05. In addition, the inactivation curves for Ito in the three treatment groups were fit by the Boltzmann equation, I 5 (Imax 2 Imin )/51 1 exp[(V 2 V1/2 )/S]6 1 Imin, where Imax and Imin are maximal and minimal currents, respectively, I is the current expressed as a function SYMPATHETIC MODULATION OF REPOLARIZING K1 CURRENTS H917 Fig. 1. Action potentials recorded from epicardial myocytes of rats treated with nerve growth factor (NGF) antibody, placebo, or NGF. Action potentials in single cells were evoked at 5-s intervals by a suprathreshold depolarizing current pulse delivered through the recording pipette. Action potential duration at 50% (APD50 ) and 90% (APD90 ) repolarization and maximum diastolic potential (MDP) are displayed in the table. APD90 in antibody-treated epicardial myocytes is significantly longer than the NGF group. The placebo group is intermediate. Data obtained from 4 litters. To study Ito density, myocytes were initially perfused with Tyrode solution to form a gigaohm seal and the membrane ruptured. After equilibrium was reached between the pipette and the cytoplasm (,5 min), cell capacitance was measured. In cells treated with placebo, capacitance was 36 6 1 pF (n 5 39). This is the same as the cell capacitance, 36 6 1 pF (n 5 54), measured from control cells (without any treatment) and the NGF-treated group (36 6 1 pF, n 5 67), suggesting that the size of the myocytes in the three groups is comparable. In contrast, the cell capacitance was 32 6 1 pF (n 5 58) in the Ab-treated group (P , 0.05), consistent with decreased myocyte size and suggesting that NGF Ab may delay cell development. Once the cell capacitance was measured, the myocytes were perfused with Ito bath solution. As soon as Na1 current disappeared, the full current-voltage (I-V) relationship for Ito was recorded as presented in Fig. 4. Data were normalized to the capacitance of each cell. The highest Ito density was observed in epicardial myocytes treated with NGF, whereas Ito density in the Ab-treated group was the smallest (despite the smaller capacitance). Using the current at 160 mV as the maximum, we generated normalized I-V curves for NGF and Ab (Fig. 4, inset). The curves superimposed, indicating that the threshold for activation of this current is not changed. The results suggest that an Ito Fig. 2. Effect of 4-aminopyridine (4-AP) on action potential duration in epicardial myocytes treated with placebo. Left: action potential recorded from a single epicardial myocyte before, during, and after administration of 0.5 mM 4-AP. Action potentials were evoked as described in Fig. 1. Right: table of APD90 values from a group of cells before, during, and after 4-AP. APD90 was significantly increased by 0.5 mM 4-AP, and this effect was readily reversed on washout. Data are expressed as means 6 SE. * P , 0.05 cf. control (paired Student’s t-test); n value is 5 for control and 4-AP and 3 for washout. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 longed the APD by 26% (P , 0.05), suggesting that Ito is critically involved in action potential repolarization. Because the action potential studies confirmed that variations in Ito could potentially account for the differences in APD90 observed between NGF- and Ab-treated animals, we next investigated the nature and the kinetics of ventricular epicardial Ito in NGF-, Ab-, and placebo-treated animals. Ito was measured as the peak current relative to steady state at the end of a 250-ms test pulse. To confirm that this was Ito, we first tested its sensitivity to 4-AP. In these experiments, we employed a higher concentration of 4-AP (2 mM) than in the action potential studies, because the possible nonspecificity of the higher concentration is not a concern given the control of external and internal solutions and voltage provided by the voltage-clamp procedure. That 4-AP reduces Ito comparably under all three treatment conditions is demonstrated in Fig. 3. Results from two individual cells are shown in Fig. 3, top, for a test voltage of 150 mV; one cell was from a rat treated with NGF, and the other was from a rat treated with Ab. In this and all subsequent experiments, the current was measured as the peak current minus the steady-state current. Mean data are provided in Fig. 3, bottom, and clearly indicate that the transient current component is indeed Ito in all three treatment groups, i.e., 4-AP block was equivalent in the three groups. H918 SYMPATHETIC MODULATION OF REPOLARIZING K1 CURRENTS Fig. 3. Sensitivity of transient outward current (Ito ) to 4-AP in the different treatment conditions. Top: records from 2 cells of animals treated with antibody or NGF. Cells were depolarized to a test potential of 150 mV from a holding potential of 280 mV. j, Control, i.e., before application of 4-AP; r, effect of 2 mM 4-AP. Ito was almost completely blocked by 4-AP. Bottom: data from groups of cells are expressed as means 6 SE at the 150 mV test potential. Percentage of inhibition was used to compare the extent of blockade by 4-AP in the 3 treatment groups. Fig. 4. Density change of Ito in epicardial myocytes from rats treated with antibody, placebo, and NGF. Cells were depolarized to various potentials from a holding potential of 280 mV at a rate of 0.1 Hz. Current (I)-voltage (V) relationship was plotted after the currents were normalized to their capacitance. Density was decreased in the myocytes treated with antibody compared with those treated with NGF at potentials positive to 140 mV. * P , 0.05, antibody group vs. NGF group. Placebo group did not differ from the treated groups. Normalized I-V relations for NGF and antibody (AB) are plotted in inset and superimpose, indicating that the voltage dependence for Ito activation was not shifted. Current at 160 mV was used as maximal current (Imax ). mV, and S was 8.6 6 1.0 mV (n 5 7). However, at the normal resting potential of these myocytes (261 to 267 mV; Fig. 1), Ito is fully available under all treatment conditions. Therefore, the data for voltage-dependent inactivation demonstrate that the reduced Ito density (Fig. 4) and the greater APD (Fig. 1) in the Ab group are not due to a voltage shift in steady-state inactivation. IK1. The method for determination of IK1 is depicted in Fig. 6A. The voltage was stepped from 2100 to 0 mV from a holding potential of 240 mV, evoking a timedependent inwardly rectifying current that decayed to a sustained, apparently steady-state level at the end of the 250-ms pulse. This current was completely blocked by 2 mM Ba21. It manifests significant inward current at negative potentials and marked inward rectification at potentials positive to 280 mV. A small negative slope was observed in some of the cells, which was also reported by Wahler (37). Characteristics such as Ba21 Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 density change may explain, at least in part, the longer APD in the Ab-treated animals. We then constructed steady-state inactivation curves (13) to determine whether or not the change in Ito density in the three groups was in part due to an effect of the treatment conditions on the voltage dependence of inactivation of the channel (Fig. 5). In the placebo group, the currents were almost completely inactivated at around 110 mV, V1/2 was 223.3 6 0.8 mV, and S was 6.7 6 0.4 mV (n 5 5). Similar results were obtained from the NGF-treated group as follows: V1/2 was 222.1 6 0.8 mV and S was 7.6 6 0.6 mV (n 5 5). In contrast, in the Ab treatment group, the curve was shifted toward more depolarized potentials and was not completely inactivated at 110 mV. The 50% inactivation potential was shifted to more positive potentials (13 mV, P , 0.05 cf. the NGF group). The calculated V1/2 was 29.7 6 1.5 SYMPATHETIC MODULATION OF REPOLARIZING K1 CURRENTS H919 sensitivity, inward rectification, inactivation of inward current, and negative slope indicate that the current is indeed IK1 (Fig. 6A). The current densities of both peak and steady-state IK1 (defined as the Ba21 difference current) were compared (Fig. 6, B and C). At potentials of 2100 to 290 mV, both peak and steady-state IK1 were significantly smaller in the Ab-treated group compared with that treated with NGF. The values for placebo were between the others. The slope conductance (fitted from the linear portion of the curve from 2100 to 280 mV) was significantly different. The slope conductance of peak current for NGF versus Ab was 0.52 6 0.03 nS/pF (n 5 11) and 0.38 6 0.03 nS/pF (n 5 9); for steady-state current, NGF versus Ab was 0.45 6 0.05 nS/pF (n 5 11) and 0.28 6 0.03 nS/pF (n 5 9; P , 0.05). The reversal Fig. 6. Determination of peak and steady-state inward rectifier K1 current (IK1 ) in an epicardial myocyte. A: current records before (control) and in the presence of 2 mM Ba21. Difference records, i.e., Ba21-sensitive current, are at right. Peak currents were measured at the beginning of the pulse, and the steady-state I-V relationship was measured at the end of the pulse Vh , holding potential. B: I-V relationship of peak IK1 in the 3 groups. C: I-V relationship of steady-state IK1 in the 3 groups. Currents were normalized to their capacitance. Myocytes in antibody-treated group had the smallest density of both peak and steady-state IK1 when compared with NGF (* P , 0.05). Placebo group did not differ from either treated group. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 Fig. 5. Effects of NGF and antibody on steady-state inactivation of Ito. Protocol used is shown in inset. A 500-ms prepulse to a range of potentials was followed by a 300-ms test pulse to a fixed potential to elicit Ito. These paired pulses were delivered at 0.1 Hz from a holding potential of 280 mV. Amplitude measurements were normalized to the value obtained at the most negative potential in each experimental condition. Data from groups of cells are expressed as means 6 SE, and the curves are fit by the Boltzmann equation. Voltage at which 50% of the current is inactivated (V1/2 ) and slope factor were calculated from the equation described in METHODS. V1/2 of the inactivation curve from the cells treated with antibody was shifted 13 mV when compared with NGF and placebo (* P , 0.05). H920 SYMPATHETIC MODULATION OF REPOLARIZING K1 CURRENTS cannot be ruled out at this stage, although IK is relatively small in rats (38). Studies of transcript abundance. It has been suggested previously that the voltage-gated K1 channels (Kv) 4.2 and Kv4.3 underlie the Ito in rat heart (11). To determine whether the changes in Ito levels that we have observed in the NGF-treated and Ab-treated animals could be produced by changes in gene expression, we examined the expression of Kv4.2 and Kv4.3 mRNA in rat heart (Table 1). No significant changes in either Kv4.2 or Kv4.3 mRNA were observed in either experimental group relative to the control. The inward rectifier in heart is a ‘‘strong’’ inward rectifier (20) that is thought to be encoded by members of the inward rectifying K1 channal (Kir) 2.0 subfamily of inward rectifier channels (12). There are three members of this subfamily of inward rectifier genes [Kir2.1, Kir2.2, and Kir2.3 (12)]. Both the Kir2.1 and Kir2.2 mRNA are abundantly expressed in rat heart, whereas Kir2.3 mRNA is not expressed (12). There was no significant change in the level of Kir2.1 and Kir2.2 mRNA in either of the experimental groups compared with control (Table 1). DISCUSSION In our previous work (29), we found that 1) norepinephrine level in heart is highest in NGF-treated rats and lowest in Ab-treated rats when compared with placebo; 2) tyrosine hydroxylase-positive material in the ventricles of NGF-treated animals is greater than in placebo-treated animals; and 3) positive chronotropy in response to tyramine is greatest in NGF-treated animals and smallest in Ab-treated animals. These data, together, support our contention that NGF treatment increases and Ab attenuates the functional sympathetic innervation of the heart. The similarity in APD measurements in our previous and present results confirm that treatment with Ab in these animals is sufficient to retard the decrease in APD that is seen with normal development and with NGF treatment. In our earlier work, we also noted that the membrane potential of Ab-treated rats was lower than that of placebo- or NGF-treated rats. Both the lower memTable 1. Effects of NGF antibody and NGF on K 1 channel transcript abundance in the rat heart Kv4.2 Kv4.3 Kir2.1 Kir2.2 Cyclophilin Placebo NGF Antibody NGF 100 6 2 100 6 7 100 6 2 100 6 2 99 6 2 92 6 11 100 6 23 100 6 2 90 6 5 100 6 4 103 6 15 97 6 4 101 6 5 99 6 8 101 6 2 Data are means 6 SE from 4 independent experiments with pooled results from 6 to 8 animals for each experimental group. NGF, nerve growth factor; Kv, voltage-gated K1 channel; Kir, inward rectifying K1 channel. Data were normalized to the amount of cyclophilin transcript, which was not significantly different between experimental groups. Values for the experimental groups are expressed as a percentage of the placebo values. There was no statistically significant difference between either of the experimental groups and the placebo group for any of the 4 mRNAs tested. Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 potential (calculated from the steady-state curve) for IK1 was 277.4 and 274.3 mV for Ab- and NGF-treated myocytes, respectively (P . 0.05). Predicted relation between repolarization of action potential and Ito and IK1 current density. The current density of Ito in Ab-treated rats is ,70% of that in NGF-treated rats, as shown in the Ito I-V relationship curves (Fig. 4). The difference between these two groups is greatest as potentials become more positive and only reaches statistical significance at 140 mV (Fig. 4), a voltage not within the physiological range. A similar consideration applies to the IK1 data. Nonetheless, there are significant differences in repolarization of action potentials between the two treatment groups (Fig. 1). These observations led us to measure the slope of action potential repolarization at two voltages, 0 and 240 mV, to represent ranges at which Ito and IK1, respectively, would be expected to contribute to repolarizing current. The location of these voltages during action potential repolarization can be appreciated by examining Fig. 2, which displays a voltage axis to the left of the action potentials. If the action potentials generated are membrane action potentials, then the current flowing across the cell membrane at any time can be calculated according to the equation I 5 CdV/dt, where C is capacitance; therefore, I/C 5 dV/dt. I/C is the current density and the unit of measurement is pA/pF, and dV/dt is the slope of action potential repolarization. The predicted current density (calculated from the action potential repolarization) around 0 mV is 3.15 pA/pF (n 5 5) in NGF-treated rats and 1.92 pA/pF (n 5 4) in Ab-treated rats. Therefore, the predicted current density around 0 mV in Ab-treated rats is 61% that of NGF-treated rats. The actual Ito current densities at 0 mV obtained from our studies are 2.01 (n 5 16) and 1.47 pA/pF (n 5 14) in NGF- and Ab-treated rats, respectively, indicating that the Ab-treated group has an Ito density that is 73% of the NGF group. It has been reported that Cd21 (100 µM) shifts the availability of Ito positive by 10 mV (1). Because Cd21 was present in our recording solution, we therefore also calculated Ito at 110 mV. The ratio of Ab and NGF is 77% (2.66 vs. 3.44 pA/pF). The good agreement between the current measurements and slope calculation suggests that the difference in actual Ito density in these two groups probably is a significant contributor to the observed difference in early action potential repolarization. A similar approach was taken to measure the slope of repolarization around 240 mV to determine whether or not IK1 is involved. The predicted current density for repolarization around 240 mV is 0.7 pA/pF (n 5 5) in NGF-treated rats and 0.26 pA/pF (n 5 4) in Ab-treated rats (i.e., Ab current 5 37% of NGF current). The actual measured IK1 densities at 240 mV are 1.24 (n 5 9) and 0.73 (n 5 11) pA/pF in NGF- and Ab-treated rats, respectively, (i.e., Ab IK1 5 58% of NGF IK1 ). Again, the reasonable agreement between the predicted and calculated changes in current density at 240 mV suggests that IK1 may play a role in the developmental regulation of the later phase of repolarization of the action potential. However, other K1 currents, such as IK, SYMPATHETIC MODULATION OF REPOLARIZING K1 CURRENTS and 10 and then remains constant through adulthood (30, 37). Such developmental changes may reflect a variety of influences on the current, including that of the sympathetic nervous system. The changes seen in IK1, in turn, could reflect differences in channel number, conductance, and/or open probability (30, 37). Changes in IK1 impact on the action potential by influencing the resting potential (35) and the final phase of repolarization (14, 19). Hence, an increase in IK1 can shorten APD and hyperpolarize the membrane. Although we did not observe a significant change in maximum diastolic potential in our three treatment groups, the membrane potential was lower in the Ab than in the placebo or NGF groups. In an earlier report, we also found a lower membrane potential in the Ab group, consistent with the present observation of reduced IK1 (36). In this respect, the magnitude of membrane potential difference was comparable to that in the present study, but the n was larger, and the result was statistically significant. Our data clearly suggest that the densities of Ito and IK1 are lower in rats treated with NGF Ab than those treated with NGF for the first 15 days of life and that the reduced densities of Ito and IK1 may contribute to the prolongation of APD. Our investigation of K1 channel transcript abundance in hearts from the three treatment groups showed no significant differences in message levels. Given the modest difference in current seen among the three groups, this result is not surprising. The change in transcript level may have been below our limit of resolution. Alternatively, it may be that innervation or NGF regulates current level by affecting the channel posttranscriptionally or otherwise modifying current density. Finally, although we have focused on Ito and IK1, this does not rule out additional changes in other currents during development and cardiac innervation, which thereby impact on action potential configuration. For example, it has been reported that L-type Ca21 current density increases developmentally in the rat ventricle (15), and nerve-muscle coculture experiments suggest that sympathetic innervation may play a role (31). However, due to additional developmental changes in channel kinetics (as measured in rabbit), the contribution of the current during normal activity may not increase and may even decrease with age (39). Another current worthy of future consideration is the Na1/Ca21 exchange current, which has been reported to contribute an inward current that would slow terminal repolarization in the rat ventricle (34). Furthermore, the message levels of the Na1/Ca21 exchanger decrease developmentally in both rat and rabbit as the sarcoplasmic reticulum develops (4), which would be consistent with a developmental reduction in the slowing of repolarization and thus a shorter APD in the adult ventricle. However, this developmental decrease in message level of the exchanger has been attributed to the postnatal surge in thyroid hormone level (3) rather than the ontogeny of sympathetic innervation. We express gratitude to Dr. Ira Cohen for thoughtful comments during the performance of these studies and for critique of the Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 brane potential and the long Q-T interval seen may be explained, at least in part, by the differences we have found in the density of IK1 and Ito. The current that we identified as Ito was recorded in Na1-free solution and in the presence of Ca21 channel blockade, with 0.3 mM CdCl2. It was an outward current with a time-dependent inactivation consistent with an earlier description of Ito (23). The external application of 2 mM 4-AP rapidly reduced the peak amplitude of the inactivating component of the outward current without significantly affecting the sustained current component, as observed in other studies (5, 6). The 4-AP sensitivity of this current further confirms that it is Ito. That Ito is the major repolarizing current responsible for the alteration of APD after injection of NGF or Ab to the neonatal rat is suggested by the following: 1) there is a lower Ito density in Ab-treated rats than NGF-treated animals; 2) the magnitude of reduction at 0 mV is consistent with the change in repolarization slope at this voltage; 3) APD of epicardial myocytes from rats treated with placebo is prolonged by low concentrations of 4-AP, which preferentially blocks Ito; and 4) Ito plays an important role in the shortening of APD in rat heart during development (38). That Ito density increases severalfold during the postnatal development of heart has been reported in freshly dissociated rat ventricular myocytes (38), cultured rat ventricular myocytes (25), and human atrial myocytes (8). Jeck and Boyden (22) reported that neonatal canine ventricular myocytes completely lack a definable Ito until ,2 mo of age. Hence, the majority of earlier studies are consistent with a developmental increase in Ito, the timing of which roughly parallels that of sympathetic innervation. This is consistent with our results with NGF and Ab, although we did not see the severalfold change in Ito density reported developmentally by others. This may be due to the fact that NGF and its Ab modulate sympathetic development only to a degree and/or that sympathetic innervation, in its own right, may be a modulator rather than the prime determinant of Ito changes during development. No shift of Ito activation was observed by us. However, the Ito inactivation curve shifted toward more positive potentials in the Ab-treated groups, which is in contrast to reports of no change in inactivation during development in human atrial (16) and rabbit ventricular myocytes (33). This could arise from species differences and/or regional variations in the ventricular wall (2, 7) and/or a nonspecific action of Ab. However, this shift is unlikely to impact on the contribution of Ito to action potential repolarization, since, in all treatment groups, the inactivation was fully relieved at the resting potential. That the density of peak and steady-state IK1 was smaller in the Ab-treated animals than in animals treated with NGF suggests a role for sympathetic innervation in the evolution of the current. This finding is consistent with reports that IK1 increases developmentally in embryonic chick ventricular myocytes (24), young rabbit ventricular myocytes (21), and freshly isolated rat ventricular cells between neonatal days 3 H921 H922 SYMPATHETIC MODULATION OF REPOLARIZING K1 CURRENTS manuscript. We also express gratitude to Drs. Irina Golyakhovsky and Natalia Egorova for assistance in the performance of the experiments and to Eileen Franey for careful attention to the preparation of the manuscript. Address for reprint requests: R. B. Robinson, Dept. of Pharmacology, College of Physicians and Surgeons of Columbia Univ., 630 West 168 St., PH 7W-318, New York, NY 10032. Received 3 April 1997; accepted in final form 1 December 1997. REFERENCES Downloaded from http://ajpheart.physiology.org/ by 10.220.33.4 on May 13, 2017 1. Agus, Z. S., I. D. Dukes, and M. Morad. Divalent cations modulate the transient outward current in rat ventricular myocytes. Am. J. Physiol. 261 (Cell Physiol. 30): C310–C318, 1991. 2. Antzelevitch, C., S. Sicouri, S. H. Litovsky, A. Lukas, S. C. Krishnan, J. M. Di Diego, G. A. Gintant, and D. W. Liu. Heterogeneity within the ventricular wall. Circ. Res. 69: 1427– 1449, 1991. 3. Boerth, S. R., and M. Artman. Thyroid hormone regulates Na1-Ca21 exchanger expression during postnatal maturation and in adult rabbit ventricular myocardium. Cardiovasc. Res. 31: E145–E152, 1996. 4. Boerth, S. R., D. B. Zimmer, and M. Artman. Steady-state mRNA levels of the sarcolemmal Na1-Ca21 exchanger peak near birth in developing rabbit and rat hearts. Circ. Res. 74: 354–359, 1994. 5. Campell, D. L., R. L. Rasmusson, Y. Qu, and H. C. Strauss. The calcium-independent transient outward potassium current in isolated ferret right ventricular myocytes. J. Gen. Physiol. 101: 571–601, 1993. 6. Castle, N. A., and M. T. Slawsky. Characterization of 4aminopyridine block of the transient outward K1 current in adult rat ventricular myocytes. J. Pharmacol. Exp. Ther. 264: 1450–1459, 1992. 7. Clark, R. B., R. A. Bouchard, E. Salinas-Stefanon, J. Sanchez-Chapula, and W. R. Giles. Heterogeneity of action potential waveforms and potassium currents in rat ventricle. Cardiovasc. Res. 27: 1795–1799, 1993. 8. Crumb, W. J., Jr., J. D. Pigott, and C. W. Clarkson. Comparison of Ito in young and adult human atrial myocytes: evidence for developmental changes. Am. J. Physiol. 268 (Heart Circ. Physiol. 37): H1335–H1342, 1995. 9. Danielson, P. E., S. Fross-Petter, M. A. Brown, L. Calavetta, J. Douglass, R. J. Milner, and J. G. Sutcliffe. p1B15: a cDNA clone of the rat mRNA encoding cyclophilin. DNA (NY) 7: 261–267, 1988. 10. Dixon, J. E., and D. McKinnon. Quantitative analysis of potassium channel mRNA expression in atrial and ventricular muscle of rats. Circ. Res. 75: 252–260, 1994. 11. Dixon, J. E., W. Shi, H. S. Wang, C. MacDonald, H. Yu, R. Wymore, I. S. Cohen, and D. McKinnon. The role of the Kv4.3 potassium channel in ventricular muscle. Circ Res. 79: 659–668, 1996. 12. Doupnik, C. A., N. Davidson, and H. A. Lester. The inward rectifier potassium channel family. Curr. Biol. 5: 268–277, 1995. 13. Fedida, D., Y. Shimoni, and W. R. Giles. Alpha-adrenergic modulation of the transient outward current in rabbit atrial myocytes. J. Physiol. Paris 423: 257–277, 1990. 14. Fermini, B., and D. F. Schanne. Determinants of action potential duration in neonatal rat ventricle cells. Cardiovasc. Res. 25: 225–243, 1991. 15. Gomez, J. P., D. Potreau, J. E. Branka, and G. Raymond. Developmental changes in Ca21 currents from newborn rat cardiomyocytes in primary culture. Pflügers Arch. 428: 241–249, 1994. 16. Gross, G. J., R. P. Burke, and N. A. Castle. Characterization of transient outward current in young human atrial myocytes. Cardiovasc. Res. 29: 112–117, 1994. 17. Hamill, O. P., A. Marty, E. Neher, B. Sakmann, and F. Sigworth. Improved patch-clamp technique for high resolution recording from cells and cell-free membrane patch. Pflügers Arch. 391: 85–100, 1981. 18. Hartzell, H. C. Regulation of cardiac ion channels by catecholamine, acetylcholine and second messengers. Prog. Biophys. Mol. Biol. 52: 165–247, 1988. 19. Hille, B. Ionic Channels of Excitable Membrane. Sunderland, MA: Sinaauer, 1984, p. 61–86. 20. Hille, B. Modulation of ion-channel function by G-proteincoupled receptors. Trends Neurosci. 17: 531–536, 1994. 21. Huynh, T. V., F. Chen, G. T. Wetzel, W. F. Friedman, and T. S. Klitzner. Developmental changes in membrane Ca21 and K1 currents in fetal, neonatal, and adult rabbit ventricular myocytes. Circ. Res. 70: 508–515, 1991. 22. Jeck, C. D., and P. A. Boyden. Age-related appearance of outward currents may contribute to developmental difference in ventricular repolarization. Circ. Res. 71: 1390–1403, 1992. 23. Josephson, I. R., J. Sanchez-Chapula, and A. M. Brown. Early outward current in rat single ventricular cells. Circ. Res. 54: 157–162, 1984. 24. Josephson, I. R., and N. Sperelakis. Developmental increase in the inwardly-rectifying K1 current of embryonic chick ventricular myocytes. Biochim. Biophys. Acta 1052: 123–127, 1990. 25. Kilborn, M. J., and D. Fedida. A study of the developmental changes in outward currents of rat ventricular myocytes. J. Physiol. Paris 430: 37–60, 1990. 26. Langer, G. A., A. J. Brady, S. N. Tan, and S. D. Serena. Correlation of the glycoside response, the force staircase and the action potential in the neonatal rat heart. Circ. Res. 36: 744–752, 1975. 27. Lee, J. H., and M. R. Rosen. Alpha1-adrenergic receptor modulation of repolarization in canine Purkinje fibers. J. Cardiovasc. Electrophysiol. 5: 232–240, 1994. 28. Liu, Q. Y., E. Karpinski, and P. K. T. Pang. Tetrandrine inhibits both T- and L-type calcium channel currents in neonatal rat ventricular cells. J. Cardiovasc. Pharmacol. 20: 513–519, 1992. 29. Malfatto, G., T. S. Rosen, S. F. Steinberg, C. U. Ursell, L. S. Sun, S. Daniel, P. Danilo, Jr., and M. R. Rosen. Sympathetic neural modulation of cardiac impulse initiation and repolarization in the newborn rat. Circ. Res. 66: 427–437, 1990. 30. Masuda, H., and N. Sperelakis. Inwardly rectifying potassium current in rat fetal and neonatal ventricular cardiomycytes. Am. J. Physiol. 265 (Heart Circ. Physiol. 34): H1107–H1111, 1993. 31. Ogawa, S., J. V. Barnett, L. Sen, J. B. Galper, T. W. Smith, and J. D. Marsh. Direct contact between sympathetic neurons and rat cardiac myocytes in vitro increases expression of functional calcium channels. J. Clin. Invest. 89: 1085–1093, 1992. 32. Rosen, M. R, P. Danilo, Jr., R. B. Robinson, A. Shah, and S. F. Steinberg. Sympathetic neural and a-adrenergic modulation of arrhythmias. Ann. NY Acad. Sci. 533: 200–209, 1988. 33. Sanchez-Chapula, J., A. Elizalde, R. Navarro-Polanco, and H. Barajas. Differences in outward currents between neonatal and adult rabbit ventricular cells. Am. J. Physiol. 266 (Heart Circ. Physiol. 35): H1184–H1194, 1994. 34. Schouten, V. J., and H. E. ter Keurs. The slow repolarization phase of the action potential in rat heart. J. Physiol. Paris 360: 13–25, 1985. 35. Shimoni, Y., R. B. Clark, and W. R. Giles. Role of an inwardly rectifying potassium current in rabbit ventricular action potential. J. Physiol. Paris 448: 709–727, 1992. 36. Sun, L. S., M. J. Legato, T. S. Rosen, S. F. Steinberg, and M. R. Rosen. Sympathetic innervation modulates ventricular impulse propagation and repolarization in the immature rat heart. Cardiovasc. Res. 27: 459–463, 1993. 37. Wahler, G. M. Developmental increase in the inwardly rectifying potassium current of rat ventricular myocytes. Am. J. Physiol. 262 (Cell Physiol. 31): C1266–C1272, 1992. 38. Wahler, G. M., S. J. Dollinger, J. M. Smith, and K. L. Flemal. Time course of postnatal changes in heart action potential and in transient outward current is different. Am. J. Physiol. 267 (Heart Circ. Physiol. 36): H1157–H1166, 1994. 39. Wetzel, G. T., F. Chen, and T. S. Klitzner. Ca21 channel kinetics in acutely isolated fetal, neonatal, and adult rabbit cardiac myocytes. Circ. Res. 72: 1065–1074, 1993.