Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Synaptic gating wikipedia , lookup
Synaptogenesis wikipedia , lookup
Hypothalamus wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
Circumventricular organs wikipedia , lookup
Microneurography wikipedia , lookup
J. Comp. Neurol. 119:77-95 (1962)
Some Fiber Projections to the Superior Colliculus
in the Cat'
JOSEPH ALTMAN
Psychophysiological Laboratory, Massachusetts Institfrte of Technology,
Cambridge, Massachusetts
Our knowledge of fiber projections to
the optic tectum or superior colliculus, an
important way-station in the visual system
of vertebrates including mammals, is still
inadequate. Studies of these fiber projections, utilizing different techniques, have
been made in various mammals (Tsai,
'25; Bodian, '37 in opossum; Johnson, '54,
in mole; Cajal, '11, in mouse; Lashley,
'34; Tsang, '37; Le Gros Clark, '42; Krieg,
'47; Nauta and Bucher, '54, in rat; Castaldi, '23-'26; Hess, '58, in guinea pig;
Cajal, '11; Loepp, '12; Minkowski, '20;
Brouwer, Zeeman and Houwer, '23; Overbosch, '27, in rabbit; Jefferson, '40, in
ferret; Monakow, 1889; Probst, '02; Cajal,
'11; Poljak, '26; Barris, Ingram and Ranson, '35; Bucher and Biirgi, '50; Biirgi,
'57, in cat; Brouwer and Zeeman, '26; Mettler, '35; Hirasawa, '38; Crosby and Henderson, '48; Whitlock and Nauta, '56;
Polyak, '57, in monkey; and Juba, '39, in
man). The majority of these investigations has been concerned with retinal projection and with projections from the
spinal cord and visual cortex. In the present study, which is a continuation of a
previous one (Altman and Carpenter, '61)
dealing with the efferent connections of
the superior colliculus in the cat, we investigated by means of the Nauta-Gygax
('54) staining technique projections to
the superior colliculus in the cat from ( a )
the retina, (b) the lateral geniculate nucleus, (c) the pulvinar, and (d) the visual
cortex. In addition, attention was paid to
projection from these structures to other
mesencephalic and also to the various
diencephalic nuclei.
MATERIALS AND METHODS
Eleven adult cats were used in this experiment. The animals were anesthetized
with Nembutal, and surgery was per-
formed under aseptic conditions. In two
animals the right orbit was unroofed and
the optic nerve was cut. Stereotaxic lesions were produced in the right lateral
geniculate nucleus in five animals, and
in the right pulvinar in one animal. In
three animals large portions of the right
visual cortex (lateral and postlateral gyri)
were removed by suction. Most of the
animals were permitted to survive for 11
days; two of them were sacrificed after a
shorter period. All animals were sacrificed by cardiac perfusion with 10% neutral formalin. The brains were sectioned
in 2-3 mm blocks perpendicular to the
axis of the brain stem and further fixed in
10% neutral formalin. Four or five selected blocks of the brain were cut serially
at 25 LI on a freezing microtome, and the
sections were stained individually according to the Laidlaw modification of the
Nauta-Gygax method ('54). In animals
with stereotaxic lesions, several frozen sections were stained with cresyl violet to
permit more precise evaluation of the location and extent of the lesions produced.
OBSERVATIONS
Fiber projections from the retina
Complete sectioning of the right optic
nerve, verified histologically, was accomplished in two animals. All stained fibers
in the right optic nerve appeared degenerated, but no degenerated fibers were
seen in the left optic nerve. The chiasma
1 This investigation was caxried put at the Department of Anatomy, Colle e of Physicians and Surgeons,
Columbia University, w%ile the author was a postdoctoral trainee in neuroanatomy (supported by grant
2B-5242 from the Institute of Neurological Diseases
and Blindness). The continuous guidance and help
provided by Dr. Malcolm B. Carpenter is gratefully
acknowledged. Aid in the reparation of this report
was received from the Nazonal Institute of Mental
Healfh, upder its program grant M-5673 to the Psychohysiologlcal Laboratory, Massachusetts Institute of
echnology.
F
77
78
JOSEPH ALTMAN
contained degenerated fibers in abundance, while in the optic tract there was a
greater preponderance of such fibers in
the contralateral optic tract (see fig. 4 ) .
A relatively small proportion of the degenerated fibers of the optic tract reached
the superior colliculus. These fibers
passed mainly through the lateral branch
of the brachium of the superior colliculus,
after a short course over the dorsolateral
edge of the medial geniculate body. Although degenerated fibers reached the superior colliculus on both sides, these were
more numerous in the contralateral than
ipsilateral brachium. A few degenerated
fibers, particularly on the contralateral
side, were also seen reaching the dorsomedial surface of the superior colliculus
by way of the medial branch of the
brachium. The fibers entering the superior colliculus were concentrated mainly
in the upper part of the stratum opticum
(the third layer from the dorsal surface).
Some of these fibers appeared to terminate
around cells situated in this layer. A small
proportion of the degenerated fibers turned
dorsally and ventrally. The dorsally oriented fibers penetrated the stratum griseum superficiale, but did not reach the
most superficial layer, the stratum zonale.
The ventrally oriented fibers moved toward
the deeper layers but did not reach the
deepest layer, the stratum album profundum. The number of degenerated fibers in the contralateral colliculus greatly
exceeded those in the homolateral colliculus. No degenerated fibers were seen
to cross in the commissure of the superior
colliculus.
At the level of the superior colliculus
degenerated fibers were absent from the
following structures : the mesencephalic
reticular formation, periaqueductal gray,
nuclear complex of the third nerve, and the
various fibrous and cellular structures of
the tegmentum. Outside the superior colliculus degenerated retinal fibers were observed in the mesencephalon only in one
area, in a small region situated dorsolateral to the cerebral peduncle. This contralateral degenerated tract, very small in
the cat, may represent the posterior accessory optic tract. More anteriorly, at the
level of the pretectum, a few degenerated
fibers were observed on the contralateral
side medial to the cerebral peduncle. This
small retinal tract may be identical with
the anterior accessory optic tract.
Retinal fibers also reach the rostral
diencephalic extension of the superior colliculus, the pretectal region. Degenerated
fibers were seen to reach this structure laterally by way of the lateral branch of the
brachium of superior colliculus, and via
the optic tract, in the ventromedial aspects
of the lateral geniculate body (the latter
fibers were to some extent coursing over
the surface of the pulvinar). A considerable proportion of fibers, particularly in
the rostral parts of the pretectal region,
passed by way of the medial branch of
the brachium of the superior colliculus.
As in the case of the superior colliculus, a
larger proportion of degenerated fibers
reached the contralateral than the ipsilateral pretectal regions. While numerical
estimates are difficult to obtain with the
technique employed, the impression was
gained that more direct retinal fibers reach
the pretectum than the superior colliculus.
The majority of retinal fibers in the pretectal region appeared to terminate laterally in the large-celled nucleus of the
optic tract, but some also penetrated the
whole extent of the pretectum by way of
the stratum opticum. Degenerated fibers
at this level were absent from the posterior
commissure and its nucleus.
The lateral geniculate nucleus was surrounded by a dense layer of degenerated
fibers; the amount of degeneration was
greater on the contralateral than ipsilateral side. The dorsal portion of the nucleus showed a laminar pattern of degeneration essentially similar to that described
recently by Hayhow ('58). The amount of
degenerated fibers seen in the dorsal nucleus of the lateral geniculate body greatly
exceeded that seen in the superior colliculus and pretectum. Also in the ventral
nucleus limited areas infiltrated by some
degenerated fibers were observed both contra- and ipsilaterally, but there was no evidence of preterminal arborization in this
area. Degenerated fibers were observed to
pass over the surface of the pulvinar; most
of these fibers appeared to terminate in
the pretectum. In limited regions of the
pulvinar, adjacent to the lateral geniculate
body, some indications of preterminal
PROJECTIONS TO SUPERIOR COLLICULUS
arborization were observed. However, extensive areas of the pulvinar were entirely
free of degenerated preterminal fibers. Degenerated fibers were also absent from all
other thalamic nuclei, including the posterolateral nucleus. The hypothalamus,
likewise, was free of degenerated fibers
with the exception of one region, above
the chiasma and in its lateral wall immediately adjacent to the optic tract,
where a small number of degenerated fibers were observed to pass. These latter
appeared to be aberrant fibers of passage
which showed no evidence of termination
in this region.
Fiber projections from the lateral
geniculate nucleus
In five animals extensive lesions were
produced in the right lateral geniculate
body. In three of these the lesions affected the ventral aspect of the dorsal nucleus, while in two the destruction was
limited mainly to the dorsal aspect of the
dorsal nucleus. As these two types of lesions produced somewhat differing patterns of degeneration, they will, in part,
be treated separately in the following description. Most of the lesions were restricted to the pars dorsalis of the lateral
geniculate body. In one animal the pars
ventralis of the lateral geniculate body
was to a large extent also destroyed; in
another the pulvinar was slightly affected,
and in several there was some involvement
of the adjacent forebrain structures (see
figs. 1, 2).
In all animals with lateral geniculate
lesions, degenerated fibers were seen to
enter the superior colliculus by way of its
brachium. Most of the degenerated fibers
penetrated by way of the lateral division
of the brachium of the superior colliculus
(situated dorsally to the medial geniculate body), while fewer fibers passed
through the medial branch of the brachium
(situated laterodorsally to the commissure
of the superior colliculus). In animals
with ventrally placed geniculate lesions
the number of degenerated fibers that entered the colliculus was considerable,
while in animals with dorsally placed
geniculate lesions the number of degenerated fibers in the colliculus was much
smaller. The involvement, in the former
79
case, of some direct retino-collicular fibers,
is possible.
Within the superior colliculus degenerated fibers were most numerous in the
stratum opticum, where these fibers were
mostly oriented horizontally. From the
stratum opticum some fibers turned dorsally to terminate in the lower parts of the
overlying stratum griseum superficiale,
while others turned ventrally and could
be followed into the deeper layers, the
stratum griseum intermediale and stratum
album intermediale. Dorsally, no fibers of
the stratum opticum reached the upper
part of the stratum griseum superficiale,
or the uppermost layer of the colliculus,
the stratum zonale. However, in two animals with ventrally placed geniculate lesions a thin band of degenerated fibers
was observed in the stratum zonale. This
degenerated band was separated from the
underlying degenerated layer (the lower
part of the stratum griseum superficiale)
by a zone devoid of degenerated fibers,
and the origin of this band could be traced
to the brachium, coursing medially over
the dorsal surface of the colliculus. In
animals with dorsally placed geniculate lesions no fibers could be traced to the lowest layer of the colliculus, the stratum
profundum, though they were present in
small numbers in animals with ventral
lesions. The large cells in the lower layers
of the colliculus were devoid of degenerated preterminal fibers in the dorsal lesion group, while in those with ventral
lesions a few degenerated fibers were observed around and in the neighborhood of
these cells. Likewise, no degenerated fibers were seen crossing in the commissure
of the superior colliculus in the dorsal lesion group, while in the two animals of
the ventral lesion group a few degenerated
fibers were seen crossing in the commissure and some were present in the contralateral colliculus, mainly in the stratum
opticum.
No degenerated fibers were found in
either group of animals in the following
mesencephalic structures : the reticular
formation, the oculomotor nucleus, the
nucleus of Darkschewitsch, or the interstitial nucleus of Cajal. In animals with
lesions involving parts of the cortex some
80
JOSEPH ALTMAN
degenerated fibers were observed in the
cerebral peduncle.
Degenerated fibers were observed in all
animals in the pretectum, though the number of degenerated fibers was considerably greater in those animals which had
ventrally placed geniculate lesions. These
degenerated fibers reached the pretectum
by way of the medial aspect of the optic
tract and the lateral and medial branches
of the brachium of the superior colliculus.
As in the superior colliculus, the majority
of degenerated fibers in the pretectum
were located in the stratum opticum,
whence they were distributed both dorsally and ventrally. Arborization of degenerated fibers around cells of the pretectum was observed. In the majority of
animals degeneration in the pretectum
was restricted to the lesion side; in one
animal some degenerated fibers were also
present in the posterior commissure and
in the contralateral pretectum.
In the lateral geniculate body degeneration was conspicuous around the lesion,
in the fibrous sheath surrounding the nucleus (fibers of the optic tract and optic
radiation), and in selected parts of the
lateral geniculate nucleus some distance
away from the lesion. In animals with dorsally placed lesions, in which the pars
ventralis of the lateral geniculate body
was spared, numerous degenerated fibers
could be observed in the latter structure.
This evidence suggests that fibers from
the dorsal or principal nucleus of the lateral geniculate body project to the ventral
or pregeniculate nucleus. No degenerated
fibers were observed in the contralateral
lateral geniculate body in any of these
animals.
In all animals degenerated fibers from
the lateral geniculate body could be followed laterally in the optic radiation to the
striate cortex, and medially in the optic
tract to various thalamic nuclei. The latter included the pulvinar, the posterior
nucleus, the suprageniculate nucleus, the
posterolateral nucleus and the dorsolateral nucleus. All of these contained degenerated fibers in large numbers on the
lesion side, with indications of terminal
arborization around cells. No degenerated
fibers were present in the intralaminar nuclei, except in one animal. This lesion in-
volving the pars ventralis, produced degeneration in the centromedian nucleus.
In another animal a few degenerated fibers were observed in the habenular
nuclei.
Ventral to the lateral geniculate body,
and in its immediate vicinity, the optic
tract was full of black granules; these appeared to be debiis from the lateral geniculate nucleus. These granules, which could
be differentiated from degenerated fibers,
were present in decreasing numbers at
increasing distances from the lateral geniculate body and were not present at the
level of the chiasma or in the optic nerves.
In two animals with ventral lesions some
degenerated fibers were present in the
optic tract. These fibers could be traced
around the lateral wall of the medial geniculate body to the reticular nucleus of the
thalamus, where apparently some of them
terminated, and from there to Forel's field
HI and the zona incerta. This ventral band
was particularly conspicuous in the animal in which the pars ventralis of the
lateral geniculate body was destroyed. In
none of the animals were degenerated fibers observed in the hypothalamus.
Fiber projections from the pulvinar
In a single animal a large lesion was
produced in the right pulvinar. The lesion destroyed a large part of the dorsal
and rostra1 portions of the pulvinar; its
more caudal and ventral parts were less
affected. The lateral geniculate body and
other adjacent diencephalic structures
were spared (see fig. 3). In this animal
degenerated fibers, many of them of large
diameter, were present in the ipsilateral
superior colliculus. These fibers appeared
to be concentrated mainly in two separate
bands, the stratum opticum and stratum
griseum intermediale - with a relatively
greater concentration in the latter layer.
From the stratum opticum some fibers
could be followed dorsally into the stratum
griseum superficiale; a very few horizontally oriented degenerated fibers were
also present in the stratum zonale. From
the stratum griseum intermediale degenerated fibers (mostly coarse ones) could
be followed ventrally as far down as the
upper edge of the stratum profundum. In
general, the number of degenerated fibers
PROJECTIONS TO SUPERIOR COLLICULUS
in the colliculus following the lesion in
the pulvinar was smaller than following
severance of the optic nerve, lesion of the
lateral geniculate nucleus or ablation of
the striate cortex. Outside the superior
colliculus degenerated fibers were not
present in any portion of the mesencephalon, including the reticular formation and
the oculomotor nuclear complex.
Degenerated preterminal fibers were
also present in the homolateral pretectum,
mostly in its medial parts. The number
of degenerated fibers in the pretectum appeared to be much smaller than in the
superior colliculus. A few degenerated fibers were also present in the homolateral
posterior and suprageniculate nuclei, and
also in some parts oE the posterolateral nucleus. No degenerated fibers were present
in the lateral geniculate nucleus, either in
the pars dorsalis or pars ventralis. Some
degenerated fibers could be observed in
the outer fibrous sheath of the lateral
geniculate body; presumably these fibers
were destined to the superior colliculus,
pretectum and the few diencephalic structures previously mentioned. Degenerated
fibers were not present in the optic tract
below the level of the lateral geniculate
body. Degenerated fibers were also present
in the neighborhood of the pulvinar in the
internal capsule, and a few degenerated
fibers could be seen in the striate cortex.
Fiber projections from the
visual cortex
The right lateral and postlateral gyri
(visual cortex) were removed by suction
in three animals. In two animals the removal was extensive, involving the greater
part of the primary visual area (see fig.
5), while in the third, only a portion of
the visual cortex was ablated (see fig. 6).
In all three animals numerous degenerated
fibers were observed to leave the cortex
by way of the internal capsule, but the
degeneration was less extensive in the
animal with small striate cortex lesion.
In animals with unilateral removal of
the striate cortex (lateral and postlateral
gyri), degenerated fibers were observed to
reach the superior colliculus in large number on the lesion side. These fibers reach
the superior colliculus mainly by way of
the lateral division of the brachium of
81
superior colliculus and, to a lesser extent
(observed only in two animals), by way
of the medial branch of the brachium.
These corticofugal fibers were mostly concentrated in the stratum griseum intermediale, but were also numerous in the
stratum opticum. A few degenerated fibers were also observed to pass on the dorsal surface of the colliculus in the stratum
zonale. In these three layers the corticofugal fibers moved mainly in a horizontal
direction, with some fibers moving dorsally and ventrally. The dorsally turning
fibers of the stratum opticum reached the
lower part of the stratum griseum superficiale but did not appear to fuse with the
degenerated fibers of the stratum zonale,
from which they remained separated by a
thin band containing no degenerated fibers. The ventrally turning fibers leaving
the stratum opticum were found intermingled with fibers that moved dorsally
from the underlying stratum griseum intermediale. In all these layers degenerated
fibers were seen surrounding cells. In particular, the large cells situated laterally in
the region of the brachium and in the
stratum album intermediale (nucleus of
the optic tract) were richly supplied with
degenerated fibers.
N o degenerated fibers were seen to cross
the commissure of the superior colliculus
(except for a n occasional fiber noted in
all animals), and degenerated fibers were
absent from the contralateral superior colliculus. No degenerated fibers were seen
in the mesencephalic reticular formation
or in any of the other mesencephalic nuclei implicated in visual functions, e. g.,
the oculomotor nucleus, the nucleus of
Darkschewitsch and the interstitial nucleus of Cajal. Some degenerated fibers
were present in the lateroventral part of
the cerebral peduncle.
Degenerated fibers reached the pretectum on the lesion side in large numbers by way of the brachium of the
superior colliculus. These fibers were concentrated mainly on the lateral aspect of
the pretectum. While lamination in the
pretectum is less distinct than in the superior colliculus, the impression was gained
that the corticofugal fibers terminated here
largely in a layer ventral to the region of
termination of the optic tract fibers
82
JOSEPH ALTMAN
(stratum opticum). Degenerated fibers
were seen arborizing around large cells
situated in the lateral part of the pretectum (nucleus of optic tract). No degenerated fibers were seen crossing in the
posterior commissure, and the opposite
pretectal region was free of degeneration.
Numerous degenerated fibers were present in the fibrous sheath of the lateral
geniculate body. However, in the dorsal
nucleus of the lateral geniculate body degenerated fibers were present only in small
number in limited regions, and they were
altogether absent in others. In general,
degenerated fibers were more abundant
in the rostral and dorsal parts of the dorsal nucleus than in its more caudally and
ventrally situated parts. A limited number of fine degenerated fibers were also
seen in some parts of the ventral nucleus
of the lateral geniculate body. There was
no degeneration in the contralateral lateral geniculate body.
In contrast to the scarcity of degenerated
corticofugal fibers in the lateral geniculate nuclei, numerous degenerated fibers
were present on the lesion side in the posterior nucleus and the pulvinar, and, to a
lesser extent, in the suprageniculate nucleus and the posterolateral nucleus. In all
these structures degenerated fibers were
seen arborizing around cells, in particular
around the larger cells of the posterior nucleus and pulvinar. In addition to these
thalamic nuclei, degenerated fibers were
also present in the dorsal parts of the
reticular nucleus in the neighborhood of
the pulvinar and lateral geniculate body.
Other parts of the thalamus, including
the intralaminar nuclei, were devoid of
degenerated fibers. No degenerated fibers
were seen in the optic tract below the
level of the lateral geniculate body or in
any part of the hypothalamus.
DISCUSSION
Fiber projection to the
superior colliculus
( a ) Retinal projection. Our observation that direct retinal fibers reach the
superior colliculus in the cat is in agreement with the findings of earlier investigators (Probst, ’02; Barris, Ingram and Ranson, ’ 3 5 ) . The description by Barris, et d.
that the crossed retinal fibers in the col-
liculus of the cat greatly exceed in number the uncrossed ones was also confirmed.
This asymmetry of projection is supported
by an electrophysiological investigation
made earlier (Altman, ’59) which showed
that, of the simultaneously recorded photically evoked potentials, those obtained
from the contralateral colliculus were consistently of larger amplitude than those
obtained from the ipsilateral colliculus.
The present investigation also confirms
the claim (Polyak, ’57) that the number
of retinocollicular fibers in the cat is quite
small in comparison with the retinogeniculate fibers. This would reflect a greater
“corticalization” in the feline visual system, as compared with lower mammalian
forms (marsupials and rodents) in which
the proportion of retinocollicular fibers is
much higher (cf. LeGros Clark, ’42; Polyak,
’57). The observation (Altman, ’59) that,
on simultaneous recording, evoked potentials to photic or optic nerve stimulation
are generally of greater amplitude in the
cat’s visual cortex than in its superior colliculus may reflect the same phenomenon.
There is general agreement in the literature that the optic fibers reach the superior colliculus through its brachium, and
that within the colliculus they are concentrated in the stratum opticum. There
is, on the other hand, a controversey of
long standing with regard to the termination of these fibers. The problem was first
investigated intensively by Cajal (‘1 1) with
the Golgi technique. He described two
kinds of fiber endings, “arborisations infkrieures,” which, entering the stratum
opticum, turn ventrally and end in the
same layer, and “arborisations supkrieures,”
which turn dorsally and end in the stratum
griseum superficiale. Some investigators
employing the Weigert and Marchi techniques (e.g., Brouwer, Zeeman and Houwer, ’ 2 3 ) similarly failed to observe terminal fibers reaching the uppermost
collicular layer, the stratum zonale. Other
histologists, however, such as Loepp (’12)
and Bodian ( ’ 3 7 ) ,claimed that some fibers
also reach the stratum zonale. In re-investigating this problem in the ferret,
Jefferson (’40) found that with the Marchi
technique osmic acid granulation is limited
to the stratum opticum and stratum griseum superficiale, with no signs of degen-
PROJECTIONS
T O SUPERIOR COLLICULUS
eration in the stratum zonale, whereas a
silver impregnation technique yielded evidence for the termination of unmyelinated
fiber branches or collaterals in the stratum
zonale. The results of the present study
do not lend support to the claim that preterminal optic tract fibers reach the stratum zonale.
( b ) Geniczilate projection. While the
existence of direct retinotectal projection
is well established, the present investigation suggests that the colliculus also receives indirect or relayed optic fibers by
way of the lateral geniculate nucleus.
This conclusion is supported by the observation that lesions of the dorsal nucleus
of the lateral geniculate body, including
those located rostrally and dorsally (and
thus presumably sparing the retinotectal
fiber system) produced preterminal degeneration in the colliculus. It should also be
noted that degeneration in the colliculus
following lesions in caudal and ventral
parts of the lateral geniculate nucleus
greatly exceeded that found after severance of the optic nerve. Such a lesion apparently affects both the direct retinotectal
fiber system situated in this region and
the indirect retino-geniculotectal projection. While the technique employed here
is not suitable for exact determination of
fiber diameters, the impression was gained
that the fibers of the retinotectal system
are generally of smaller diameter than the
fibers of the retinogeniculate system. This
has been claimed by several investigators
using histological material (e.g., Gudden,
1886, in rabbit; Bishop and Clare, ’55, in
cat; Polyak, ’57, in monkey) and is also
in agreement with the electrophysiological
evidence that the conduction velocity of
impulses is much lower in the retinotectal
than the retinogeniculo-striate projection
system, (e.g., Bishop and Clare, ’55; Altman, ’59).
( c ) Projection from the pulvinar. The
superior colliculus also receives fibers from
the pulvinar. As the pulvinar itself receives but few fibers from the optic tract,
this connection may not serve a function
similar to that of the retino-geniculo-tectal
fiber system.
Of the structures investigated in this
study, the pulvinar alone appears to have
reciprocal connections with the superior
83
colliculus; the lateral geniculate and striate cortex send fibers to the colliculus but
do not receive any from it.
( d ) Cortical projection. In addition to
fibers from optic tract, lateral geniculate
nucleus and pulvinar, the superior colliculus also receives a large number of
fibers from the ipsilateral striate cortex.
Evidence for corticotectal projection in
the cat was first obtained by Monakow
(1889), and was later confirmed by Probst
(’02); Beevor and Horsley (’02); Cajal
(’11);Poljak (’26,’28) and Barns, Ingram
and Ranson (’35) in the cat, and by several investigators in other mammalian species (Johnson, ’54, in mole; Krieg, ’47;
Nauta and Bucher, ’54, in rat; Leblanc,
’28, in rabbit; Probst, ’02, in dog; Jefferson,
’40; in ferret; Ferrier and Turner, 1897;
Mettler, ’35; Crosby and Henderson, ’48;
in monkey). With regard to the laminar
distribution of corticofugal fibers in the
colliculus there is considerable disagreement. Some investigators localized the penetrating corticotectal fibers in a single layer,
either in the stratum opticum (e.g., Probst,
’02 [described as “lower zonal layer”],
Barris, Ingram and Ranson, ’35; Jefferson,
’40), or ventral to the stratum opticum
(Cajal, ’ l l ) , with arborizations in the
layers situated dorsally and ventrally.
Others described two zones of distribution
(e.g., Poljak, ’26, ’28 [“upper” and “middle” layers], Krieg, ’47 [“superficial fibrous
lamina” and ‘“deep lamina”] ) . The present investigation suggests that the corticotectal fibers are distributed in the cat
through three separate zones in the colliculus. First, a very small but distinct
band of fibers terminates in the stratum
zonale. These small fibers do not come by
way of the stratum opticum, as some in
vestigators suggested (e.g., Barris, Ingram
and Ranson, ’35), but may be followed
directly from the brachium as a thin band
of horizontal fibers moving over the dorsal surface of the colliculus. Second, some
fibers are present in the stratum opticum,
forming a dark band in this region. Some
of these penetrate dorsally into the stratum
griseum superficiale, but do not reach the
stratum zonale, while others turn ventrally and end in the stratum griseum intermediale. The largest share of corticofugal fibers is concentrated in this latter
84
JOSEPH ALTMAN
layer, and the majority of them appears to
come directly from the brachium rather
than via the stratum opticum. Thus, the
bulk of corticotectal fibers is concentrated
in a layer ventral to the stratum opticum
(the layer of termination of the direct retinal and indirect retinogeniculate fibers);
an observation that was made first by
Cajal ('11) and was noted later by other
investigators (e.g., Crosby and Henderson,
'48).
The corticotectal fibers from the striate
cortex and adjacent regions are strictly
homolateral in distribution, and no fibers
of this system cross in the commissure of
the superior colliculus. This observation
is in agreement with the reports of the
majority of investigators considered, with
the exception of Probst ('02),who claimed
that in the cat some corticotectal fibers go
to the contralateral colliculus by way of
the commissure. The observation made
by Mettler ('35) and Crosby and Henderson ('48) that, in the monkey, the occipital
cortex projects to the aculomotor nucleus,
the nucleus of Darkschewitsch, and interstitial nucleus of Cajal could not be substantiated in the cat. Apart from the
superficial and middle layers of the superior colliculus, which receive a massive
projection from the visual cortex, no other
mesencephalic structure receives fibers
from this region. This applies also to the
mesencephalic reticular formation which
was found to be devoid of degenerated
fibers.
With regard to the reticular formation,
the present investigation suggests that
this "activating" or "arousal" mechanism
does not receive direct input from the
optic tract, the lateral geniculate nucleus,
the pulvinar or striate cortex. On the
other hand, evidence was obtained in a
previous study (Altman and Carpenter,
'61) that a major efferent outflow of the
superior colliculus is into the mesencephalic and bulbar reticular formation.
Thus, it is conceivable that the EEG activation and behavioral arousal effects of
peripheral (retinal) and central (cortical)
visual stimulation are, at least partially,
mediated by way of the superior colliculus.
For the superior colliculus receives input
from all the visual structures considered
and discharges into the reticular forma-
tion. That electrical stimulation of the
superior colliculus produces EEG activation and behavioral arousal was observed
by Pearce (cf., Jefferson, '58), and there
is an unconfirmed report (Takebayashi,
'57) that after bilateral ablation of the colliculus in rabbits EEG activation by light
flash (blocking of alpha activity) is
abolished.
Fiber projections to and within
the diencephalon
( a ) Retinal projection to the diencephalon. Some investigators (e.g., Polyak,
'57) reported that in the cat, as in primates, the number of crossed and uncrossed fibers in the optic tract are approximately equal. The observation made
in the present investigation was that the
crossed optic fibers greatly exceed in
number the uncrossed fibers in this
species. Similarly, the amount of degeneration i n the contralateral lateral geniculate nucleus greatly exceeded the degeneration found on the homolateral side. No
detailed investigation was made here of
the pattern of termination of the crossed
and uncrossed components in the laminae
of the lateral geniculate nucleus, but the
essential features of a recent description
by Hayhow ('58) could be confirmed.
There is considerable controversy in the
literature with regard to the problem of
retinal projection to the ventral nucleus of
the lateral geniculate body. Some investigators have claimed that a few retinal fibers terminate in this structure (eg.,
Castaldi, '23, in guinea pig; Pavlow, '00;
Cajal, '11; Loepp, '12; Brouwer, Zeeman
and Houwer, '23, in rabbit; Minkowski,
'20; O'Leary, '40; Polyak, '57; Hayhow,
'58, in cat; Polyak, '57, in monkey).
Others maintained that, while retinal fibers do pass through the ventral nucleus,
none terminate in it (Tsai, '25, in opossum; Lashley, '34; Tsang, '37, in rat; Jefferson, '40, in ferret; Barris, Ingram and
Ranson, '35; Glees, '41, in cat). The present investigation does not presume to resolve this controversy. It should be
pointed out, however, that while retinal
fibers were observed to pass through the
ventral nucleus in our material, arborization around cells was not observed.
PROJECTIONS T O SUPERIOR COLLICULUS
Apart from the pretectum and the dorsal nucleus of the lateral geniculate body,
no other diencephalic structures were observed to receive direct optic fibers in any
great number. This applies to the suprageniculate, posterior and posterolateral nuclei
of the thalamus, which receive fibers from
various central visual structures but do
not receive any direct fibers from the
retina. Some optic fibers were observed
to pass over the dorsolateral surface of
the pulvinar, but only a few of these appeared to terminate there. This agrees
with observations by a majority of investigators (Monakow, 1889, Probst, '02;
Minkowski, '13, '20; Polyak, '57, in various
mammalian species), although others denied that the pulvinar receives any direct
optic fibers (Brouwer, Zeeman and Houwer, '23; Rioch, '29; Barris, Ingram and
Ranson, '35). Fibers of retinal origin were
also observed to course through the hypothalamus, above the optic chiasma, but no
evidence was obtained that these fibers
actually terminated in this region. Finally,
we may mention the two small fiber bundles of retinal origin, the so-called anterior and posterior accessory optic tracts,
both of which contain only crossed fibers.
The first of these terminates medial to
the cerebral peduncle at the level of the
pretectum, while the latter terminates in
a nuclear region dorsolateral to the cerebral peduncle at the level of the superior
colliculus. The recent description by Hayhow ('59) of the origin and course of
these fiber tracts in the cat could not be
adequately evaluated in the material available.
(b) Projections t o and from the lateral
geniculate nucleus. The main fiber projection of the dorsal nucleus of the lateral
geniculate nucleus is to the striate cortex.
In addition, the lateral geniculate nucleus
sends fibers (in part possibly collaterals)
to the superior colliculus, to the pretectum,
to the pulvinar and to the posterior,
posterolateral and suprageniculate nuclei of the thalamus. In contrast to this
extensive projection from the lateral geniculate body, the projection to it is very
sparse from the striate cortex and nonexistent from the pulvinar and, as a previous study has shown (Altman and Carpenter, '61), from the superior colliculus.
85
The pathway through the lateral geniculate is, accordingly, essentially unidirectional, with an input from the retina and
an output to the structures mentioned. If
the lateral geniculate nucleus receives any
reciprocal fibers at all from the structures
to which it projects, such a connection
must be by way of a central nervous region that was not investigated here (such
as, perhaps, the pre- and parastriate regions of the cortex). Of some importance
also is the finding that the pars dorsalis
of the lateral geniculate body projects to
the pars ventralis. Accordingly, the pars
ventralis of the lateral geniculate body,
which does not appear to receive direct
retinal fibers, receives input from at least
two structures implicated in vision, the
dorsal nucleus and, as was previously
shown (Altman and Carpenter, '61), the
superior colliculus. The possibility that
the ventral nucleus of the lateral geniculate body represents a motor outflow in the
visual system via the subthalamus (cf.
Polyak, '57), deserves consideration.
( c ) Projections to and from the pulvinar. The pulvinar, which receives few
direct retinal fibers, receives indirect optical input from the lateral geniculate nucleus, the striate cortex, and also from the
superior colliculus (Altman and Carpenter,
'61). It sends reciprocal fibers to the superior colliculus and the striate cortex. In
the single animal investigated in this
study, no evidence could be obtained that
it projects to the lateral geniculate nucleus. The possibility, of course, cannot
be excluded that with a larger and more
favorably placed lesion in the pulvinar
such a connection could be demonstrated.
The present evidence is compatible with
the view that the pulvinar is an important
secondary link in the visual system, comparable to the neighboring structures of
the posterior, suprageniculate and posterolateral nuclei of the thalamus.
( d ) Cortical projection to the diencephd o n . In contrast to the extensive projection from the dorsal nucleus of the lateral
geniculate body to the striate cortex,
corticofugal projection from the striate
cortex to this structure is suprisingly small.
The fibrous sheath of the lateral geniculate
body was full of degenerated fibers following ablation of the striate cortex; the dor-
86
JOSEPH ALTMAN
alon were also investigated. The following
conclusions were drawn from this study:
1. The superior colliculus receives afferent fibers directly from the retina.
These afferents, which are more numerous
on the contralateral than on the ipsilateral
side, are distributed through the stratum
opticum and reach dorsally the stratum
griseum superficiale and ventrally the stratum griseum intermediale. Fibers of retinal origin do not cross in the commissure
of the superior colliculus.
2. The superior colliculus also receives
indirect optic afferents from the ipsilateral
lateral geniculate nucleus. The intracollicular distribution of these fibers is similar to that of the direct optic fibers.
3. The superior colliculus receives ipsilaterally a few coarse fibers from the pulvinar and a bulky projection from the
striate cortex. The corticofugal fibers are
distributed separately through three layers,
the stratum zonale, stratum opticum and
the stratum griseum intermediale.
4. Fiber projection could not be demonstrated from the retina, lateral geniculate,
pulvinar or striate cortex to the third nerve
nuclear complex or the mesencephalic
reticular formation, Projection from these
structures to the pretectum is essentially
similar to that of the superior colliculus.
5. The number of retinal fibers reaching the lateral geniculate nucleus in the
cat greatly exceeds the number of fibers
reaching the superior colliculus and pretectum. The lateral geniculate nucleus
projects, not only to the striate cortex, but
also to the superior colliculus, pretectum,
pulvinar and several other thalarnic nuclei. It receives only a few reciprocal fibers from the striate cortex and none from
SUMMARY AND CONCLUSIONS
the superior colliculus or pulvinar.
6. Extensive projection from the striate
Afferent projection to the superior colliculus was investigated by means of the cortex was observed to the following thalNauta-Gygax staining technique in 11cats. amic nuclei: pulvinar, the posterior, supraIn two animals the right optic nerve was geniculate and posterolateral nuclei. These
cut; in five animals stereotaxic lesions structures also receive fibers from the
were produced in the right lateral geni- superior collicuIus and the lateral genicuculate body; in a single animal, a re- late.
LITERATURE CITED
stricted stereotaxic lesion was produced
in the pulvinar; and in three animals the Altman, J. 1959 A neurophysiological study of
the superior colliculus. Ph.D. thesis, New York
right striate cortex (lateral and postlatUniversity.
era1 gyri) was removed by suction. Fiber Altman,
J., and M. B. Carpenter 1961 Fiber
projections from these structures to other
projections of the superior colliculus in the
mesencephalic nuclei and to the diencephcat. J. Comp. New., 116: 157-178.
sal nucleus itself, however, showed no degenerated fibers over large areas and only
a few degenerated preterminal fibers over
limited regions, in particular, its rostra1
and dorsal portions. A few fine degenerated fibers, with suggestions of termination around cells, were seen in parts of
the ventral nucleus of the lateral geniculate body. While the lateral geniculate nucleus, whose main outflow is into the
striate cortex, receives but a few reciprocal
fibers from this region, numerous degenerated preterminal fibers were observed
after ablation of the striate cortex on the
lesion side in several other thalamic nuclei - the posterior nucleus, the pulvinar,
the suprageniculate and posterolateral nu.
clei, and also in the pretectal region.
Especially striking was the rich corticofugal projection to the posterior nucleus
and the pulvinar. It is possible that the
postulated reciprocal cortico-thalamic connection within the visual system (cf.,
Bartley, '59, p. 718) is mediated, at least
partly, by way of these nuclei. In the same
way, reciprocal cortical connections of the
superior colliculus (which receives many
corticofugal fibers but does not send fibers
either to the cortex directly or to the lateral geniculate nucleus) may be by way of
these thalamic nuclei, since all of them receive fibers from the colliculus (Altman
and Carpenter, '61 ). Our present findings
contradict, incidentally, the claim of
Barris, Ingram and Ranson ('35) that the
striate cortex sends no fibers to the pretectum, the pulvinar, the posterior and
posterolateral nuclei, or the dorsal and
ventral nuclei of the lateral geniculate
body.
PROJECTIONS TO SUPERIOR COLLICULUS
87
Hess, A. 1958 Optic centers and pathways
Barris, R. W., W. R. Ingram and S. W. Ranson
after eye removal i n fetal guinea pigs. Ibid.,
1935 Optic connections of the diencephalon
109: 91-116.
and midbrain of the cat. Ibid., 62: 117-153.
Hirasawa, K., S. Okano and S. Kamio 1938
Bartley, S. H. 1959 Central mechanisms of
Beitrag zur Kenntnis uber die corticalen extravision. In: Handbook of Physiology, section
pyramidalen Fasern aus der Area temporalis
1, vol. 1, Neurophysiology. American Physiosuperior (Area 2 2 ) beim Affen. Z. mikr. anat.
logical Society, Washington, chap. 30, pp. 713Forsch., 44: 74-84.
740.
Beevor, C. E., and V. Horsley 1902 On the pal- Jefferson, G. 1958 Discussion. I n : Henry Ford
Hospital International Symposium, Reticular
lio-tectal or corticomeseocephalic system of
Formation of the Brain. Little, Brown, Boston,
fibers. Brain, 25: 436-443.
pp. 65-68.
Bishop, G. H., and M. H. Clare 1955 Organization and distribution of fibers in the optic Jefferson, J. M. 1940 A study of the subcortical connexions of the optic tract system of
tract of the cat. J. Comp. Neur., 103: 269-304.
the ferret with special reference to gonadal
Bodian, D. 1937 An experimental study of the
activation by retinal stimulation. J. Anat.,
optic tracts and retinal projection of the
London, 75: 106-134.
Virginia opossum. Ibid., 66: 113-144.
Johnson, T. N. 1954 The superior and inferior
Brouwer, B., and W. P. C. Zeeman 1926 The
colliculi of the mole (Scalopus aquaticus
projection of the retina i n the primary optic
machrinus). J. Comp. Neur., 101: 765-799.
neuron i n monkeys. Brain, 49: 1-35.
Juba, A. 1939 Die optischen Verbindungen der
Brouwer, B., W. P. C. Zeeman and A. W. M.
oberen Vierhiigelgegegnd. 2. ges. Neurol.
Houwer 1923 Experimentell-anatomischeUnPsychiat., 164: 273-285.
tersuchungen uber die Projektion der Retina
auf die primaren Opticuszentren. Schweiz. Krieg, W. J. S. 1947 Connections of the cerebral cortex. I. The albino rat. C. Extrinsic
Arch. Neurol. Psychiat., 23: 118-137.
connections. J. Comp. Neur., 86: 267-394.
Bucher, V. M., and S. M. Biirgi 1950 Some observations on the fiber connections of the di- Lashley, K. S. 1934 The mechanism of vision.
VII. The projection of the retina upon the
and mesencephalon i n the cat. I. Fiber conprimary optic centers in the rat. Ibid., 59:
nections of the optic tectum. J. Comp. Neur.,
341-3 73,
93: 139-172.
Burgi, S. 1957 Das Tectum opticum. Seine Leblanc, L. 1928 Recherches sur la systematisation des voies corticotectales. La Cellule,
Verbindungen bei der Katze und seine Bedeut38: 353-384.
ung beim Menschen. Dtsch. Z. Nervenheilk.,
Loepp, W. H. 1912 Ueber die zentralen Opti176: 701-729.
cusendigung beim Kaninchen. Anat. Anz., 40:
Cajal, S. Ram6n Y. 1952-1955 (1909-1911)
309-323.
Histologie du systBme nerveux de l'homme et
des vertkbrks, 2 vols. Consejo de Investiga- Mettler, F. A. 1935 Corticifugal fiber connections of the cortex of Macaca mulatta. A. The
ciones Cientificas, Madrid, vol. 2, chap. 9, pp.
occipital region. J. Comp. Neur., 61: 221-256.
175-193.
Castaldi, L. 1923, 1924, 1926 Studi sulla strut- Minkowski, M. 1913 Experimentelle Untersuchungen iiber die Beziehungen der Grosshirntura e sullo sviluppo del mesencefalo. Ricerche
rinde und der Netzhaut zu den primaren
in Cavia Cobaya. Parte 1, 2, 3. Arch. ital. anat.
optischen Zentren, besonders zum Corpus genembryol., 20: 23-225; 21: 172-263; 23: 481iculatum externum. Arb. hirnanat. Inst.,
609.
Ziirich, 7: 259-362.
Clark, W. E. Le Gros 1942 Visual centers of
1920 Ueber den Verlauf, die Endigung
the brain and their connections. Physiol. Rev.,
und die zentrale Reprasentation von gekreuzten
22: 205-232.
und ungekreuzten Sehnervenfasern bei einiCrosby, E. C., J. W. Henderson 1948 The mammalian midbrain and isthmus regions. 11. Figen Saugetieren und beim Menschen. Schweiz.
Arch. Neurol. Psychiat., 6: 201-252; 7: 268ber conections of the superior colliculus. B.
303.
Pathways concerned in automatic eye moveMonakow, C. von 1889 Experimentelle unct
ments. J. Comp. Neur., 88: 53-91.
pathologisch-anatomische
Untersuchungen iiber
Ferrier, D., and W. A. Turner 1897 An experidie optischen Centren und Bahnen. Arch. Psymental research upon cerebrocortical afferent
chiat., 20: 714-787.
and efferent tracts. Proc. Roy. SOC., London,
Nauta, W. J. H., and V. N. Bucher 1954 Effer62: 1-3.
ent connections of the striate cortex in the
Glees, P. 1941 The termination of optic fibres
albino rat. J. Comp. Neur., 100: 257-286.
i n the lateral geniculate body of the cat. J.
Nauta, W. J. H., and P. A. Gygax 1954 Silver
Anat., London, 76: 65-92.
impregnation of degenerating axons in central
Gudden, B . von 1886 Demonstration der Sehfanervous system. A modified technique. Stain
sern und Pupillarfasern des Nervus opticus.
Tech., 29: 91-93.
Sitzungsb. Gesellsch. Morphol. Physiol., I : 169.
O'Leary, J. L. 1940 A structural analysis of
Hayhow, W. R. 1958 The cytoarchitecture of
the lateral geniculate body in the cat in relathe lateral geniculate nucleus of the cat. J.
Comp. Neur., 73: 405-430.
tion to the distribution of crossed and unOverbosch, J. F. A. 1927 Experimenteel-anatcrossed optic fibers. J. Comp. Neur., 110: 1-63.
omische onderzoekingen over de projecfie der
1959 An experimental study of the accessory optic fiber system in the cat. Ibid.,
retina in het centrale zenuwstelsel. Academisch proefschrift, Amsterdam.
113: 281-313.
88
JOSEPH ALTMAN
Pavlow, M. 1900 Les voies descendantes des
tubercules quadrijunleaux supCrieurs, 1-11, Le
Nevraxe, I : 57-75, 129-136.
Poljak, S. 1926 Die Verbindungen der Area
striata (intrahemisphaerale, kommissurale, palliodienzephalische, palliotektale Fasern) bei
der Katze und deren funktionelle Bedeutung.
2.ges. Neurol. Psychiat., 100: 545-563.
1928 An experimental study of the
association, callosal and projection fibers of
the cerebral cortex of the cat. J. Comp. Neur.,
44: 197-258.
Polyak, S. 1957 The vertebrate visual system.
H. Kliiver, ed. Univ. of Chicago Press, Chicago,
chap. 6, pp. 288-389.
Probst, M. 1902 Ueber den Verlauf der centralen Sehfasern (Rinde-Sehhiigelfasern) und
deren Endigung im Zwischen- und Mittelhirne
und uber die Associations- und Commissuren-
fasern der Sehsphare. Arch. Psychiat. Nervenkr., 35: 22-43.
Rioch, D.M. 1929 Studies on the diencephalon
of carnivora. I. Nuclear configuration of the
thalamus, epithalamus and hypothalamus of
dog and cat. J. Comp. Neur., 49: 1-120.
Takebayashi, H. 1957 Superior colliculus, playing roles in the optokinetic and neurovegetative mechanism. Wakayama Med. Rep., 4 :
1-12.
Tsai, C. 1925 The descending tracts of the
thalamus and midbrain of the opossum, Didelp h i s virginiana. J. Comp. Neur., 39: 317-348.
Tsang, Y.-(2. 1937 Visual centers in blinded
rats. Ibid., 66: 211-261.
Whitlock, D. G.,and W. J. H.Nauta 1956 Subcortical projections from the temporal neocortex
i n Macaca mulatta. Ibid., 106: 183-212.
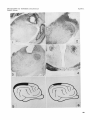
PLATE 1
EXPLANATION OF FIGURES
Low power microphotographs and drawings illustrating some of the
lesions produced.
Cat C-439. Lesion in the right lateral geniculate nucleus, pars dorsalis. Frozen section; Nissl. x 5.
Cat C-434. Less extensive lesion in right lateral geniculate nucleus,
pars dorsalis. Frozen section; Nissl. X 5.
Cat C-438. Lesion in right pulvinar. Frozen section; Nissl. X 5.
Cat C-429. Degenerated fibers in the left (severe) and right (milder)
optic tract, following severance of the right optic nerve. Nauta-Gygax.
x 10.
Cat C-426. Schematic drawing, illustrating extensive removal of the
striate cortex in this animal.
Cat C-411. Schematic drawing, illustrating partial removal of the striate cortex in this animal.
PROJECTIONS TO SUPERIOR COLLICULUS
PLATE 1
Joseph Altman
89
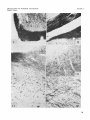
PLATE 2
EXPLANATION OF FIGURES
Low and high power microphotographs illustrating axonal and preterminal degcncration in mesencephalic and dicncephalic regions following
severance of the right optic nerve.
7
Cat C-429. Degcneratcd fibers in and surrounding the contralateral
lateral geniculate nucleus. Nauta-Gvgax. X 73.
8
Cat C-429. Degencrated fibers surrounding the contralateral lateral
geniculate nucleus (upper part) and i n the brachiurn of the superior
colliculus at the level of the pretectal region (lower part). NautaGygax. X 73.
9
Cat C-428. Degenerated fibers in the contralateral brachium of the
superior colliculus and in the lateral parts of the superior colllculus.
Nauta-Gygax. X 73.
10 Cat C-429. Degenerated fibcrs i n the stratum opticum i n the contralateral superior colliculus. Nauta-Gygax. X 73.
90
11
Cat C-428. Deqenerated fibers in the contralateral superior colliculus
at higher magnification. Nauta-Gygax. X 360.
12
Cat C-429. Degenerated fibers in the contralateral prctcctal region
at higher magnification. Nauta-Gygax. X 360.
PROJECTIONS TO SUPERIOR COLLICULUS
Joseph Altman
PLATE 2
91
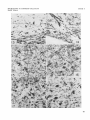
PLATE 3
EXPLANATION OF FIGURES
High power microphotographs illustrating axonal and preterminal degeneration. Figures 13-14: degeneration iollowing severance of the right
optic nerve. Figures 15-18 : degeneration following coagulation of the
lateral geniculate nucleus, pars dorsalis.
13
Cat C-429. Degenerated “aberrant” optic fibers above the chiasma
i n the hypothalamus. Nauta-Gygax. x 360.
14
Cat C-429. Degenerated optic fibers i n the contralateral medial accessory tract. Nauta-Gygax. X 360.
15
Cat C-439. Degenerated fibers in the ipsilateral superior colliculus
following lesion in the lateral geniculate nucleus, pars dorsalis.
Nauta-Gygax. x 360.
16
Cat (2-439. Degenerated fibers in the ipsilateral pretectal area following lesion i n the lateral geniculate nucleus, pars dorsalis. NautaGygax. x 360.
17
Cat C-439. Degenerated fibers in the pars ventralis of the lateral
geniculate nucleus following lesion in the pars dorsalis. Nauta-Gygax.
x 360.
18 Cat C-439. Degenerated fibers in the ipsilateral pulvinar following
lesion in the lateral geniculate nucleus, pars dorsalis. Nauta-Gygax.
X 360.
92
PROJECTIONS TO SUPERIOR COLLICIJLUS
Joseph Altman
PLATE 3
93
PLATE 4
EXPLANATION OF FIGURES
Low and high power microphotographs illustrating axonal and preterminal degeneration. Figures 19-22: Degeneration following partial
removal of the right striate cortex (lateral and postlateral gyri). Figures
23-24 : Degeneration following lesion in the right pulvinar.
19
Cat C-427. Degenerated fibers in ipsilateral superior colliculus (particularly abundant in stratum griseuin intermediale; lower half of
picture) following striate cortex lesion. Nauta-Gygax. x 73.
20
Cat C-426. Degenerated fibers in ipsilateral superior colliculus following striate cortex lesion, at higher magnification. Nauta-Gygax.
X 360.
21 Cat C-427. Degenerated fibers in ipsilateral lateral geniculate nucleus, pars dorsalis, following striate cortex lesion. Nauta-Gygax.
X 360.
94
22
Cat C-437. Degenerated fibers in ipsilateral posterior nucleus of the
thalamus following striate cortex lesion. Nauta-Gygax. x 360.
23
Cat C-438. Degenerated fibers in ipsilateral superior colliculus following lesion in pulvinar. Nauta-Gygax. X 360.
24
Cat C438. Degenerated fibers in the stratum griseum intermediale
of the ipsilateral superior colliculus following lesion in pulvinar.
Nauta-Gygax. x 360.
PROJECTIONS TO SUPERIOR COLLICULUS
Joseph Altman
PLATE 4
95