Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
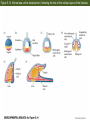
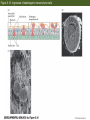
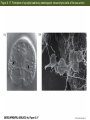
Figure 5.1 Cell cycles of somatic cells and early blastomeres (Part 1) Figure 5.1 Cell cycles of somatic cells and early blastomeres (Part 2) Figure 5.2 Role of microtubules and microfilaments in cell division Figure 5.3 Summary of the main patterns of cleavage (Part 1) Figure 5.3 Summary of the main patterns of cleavage (Part 2) Figure 5.4 Types of cell movements during gastrulation (Part 1) The Cells are given new position and new neighbors, and Multilayered body plan of organism are established Cell movement: ectoderm, endoderm and mesoderm formation Figure 5.4 Types of cell movements during gastrulation (Part 2) Figure 5.5 Axes of a bilaterally symmetrical animal Figure 5.6 Cleavage in the sea urchin (Part 1) Unequal equatorial cleavage Figure 5.7 Micrographs of cleavage in live embryos of the sea urchin Lytechinus variegatus, seen from the side Figure 5.14 Normal sea urchin development, following the fate of the cellular layers of the blastula Figure 5.8 Fate map and cell lineage of the sea urchin Strongylocentrotus purpuratus Figure 5.9 Ability of micromeres to induce presumptive ectodermal cells to acquire other fates Figure 5.10 Ability of micromeres to induce a secondary axis in sea urchin embryos Micromere: 1) autonomous specification 2) Paracrine production to specify the neighboring cells Figure 5.11 Role of Disheveled and -catenin proteins in specifying the vegetal cells of the sea urchin embryo (Part 1) The Wnt signal transduction pathways (Part 1) – Canonical Wnt pathway Figure 5.11 Role of Disheveled and -catenin proteins in specifying the vegetal cells of the sea urchin embryo (Part 2) LiCl treatment Animal cells become specified as Endoderm & mesoderm formation Inhibition of b-Cat transportation into nuclei Ciliated ectodermal cells Figure 5.12 Simplified, double-negative gated “circuit” for micromere specification double-negative gated “circuit” Figure 5.13 “Logic circuits” for gene expression double-negative gated “circuit” Feedforward circuit Figure 5.14 Normal sea urchin development, following the fate of the cellular layers of the blastula Figure 5.16 Ingression of skeletogenic mesenchyme cells Figure 5.17 Formation of syncytial cables by skeletogenic mesenchyme cells of the sea urchin Figure 5.18 Localization of skeletogenic mesenchyme cells Figure 5.19 Invagination of the vegetal plate Figure 5.20 Cell rearrangement during extension of the archenteron in sea urchin embryos Figure 5.21 Mid-gastrula stage of Lytechinus pictus, showing filopodial extensions of nonskeletogenic mesenchyme Figure 5.15 Entire sequence of gastrulation in Lytechinus variegatus Figure 5.42 The nematode Caenorhabditis elegans (Part 1) Figure 5.42 The nematode Caenorhabditis elegans (Part 2) Figure 5.42 The nematode Caenorhabditis elegans (Part 3) Figure 5.43 PAR proteins and the establishment of polarity Figure 5.44 Segregation of the P-granules into the germ line lineage of the C. elegans embryo the P-granules: riboneucleopotein complex, -RNA helicase, Poly A pol, translational initiation factors -move toward the posterior ends, P lineage blastomere, become germ cells Figure 5.46 Model for specification of the MS blastomere Figure 5.45 Deficiencies of intestine and pharynx in skn-1 mutants of C. elegans Figure 5.47 Cell-cell signaling in the 4-cell embryo of C. elegans If P2 is removed, EMS become two MS cells, no E cells If you reversed the position of Ap and Aba, their fates are similiary reversed Mom-2: Wnt Mom-5: Frizzled Apx-1: Delta Glp-1: Notch Figure 5.48 Gastrulation in C. elegans