Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
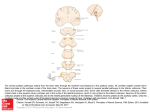
Histologic Analysis of Chronos in an Optogenetics-Based Auditory Brainstem Implant Model 1,2 MD, Massachusetts Eye and Ear 1,2 MD, 2,3 MS, Ashton E. Lehmann, Elliott D. Kozin, A. Ed Hight, Keith Darrow, 5 1,2 1,2 Ed Boyden, PhD, M. Chris Brown, PhD, Daniel J. Lee, MD 2,4 PhD, 1 Department of Otology and Laryngology, Harvard Medical School, Boston, MA. 2 Eaton-Peabody Laboratories, Massachusetts Eye and Ear Infirmary, Boston, MA; 3Program in Speech and Hearing Bioscience and Technology, Harvard Medical School, Boston, MA. 4 Department of Communication Sciences and Disorders, Worcester State University, Worcester, MA. 5 Departments of Brain and Cognitive Sciences and Biological Engineering, MIT Media Lab and McGovern Institute, MIT, Cambridge, MA. ABSTRACT INTRODUCTION RESULTS DISCUSSION Objectives: Optogenetics affords the potential for improved spatial resolution compared to electric stimulation in future auditory neuroprostheses. No histologic studies have yet examined the virally-mediated gene transfer of Chronos, a new opsin, to the murine cochlear nucleus (CN). Herein, we aim to 1) identify CN regions and neurons receptive to gene transfer of Chronos and 2) describe the morphology of putative cell types that express Chronos. • Current auditory brainstem implants rely on electrical stimulation to provide sound awareness to patients who are not candidates for cochlear implants. • Electric stimulation of neurons results in current spread, compromising signal specificity.1 • As optical stimulation may be able to excite discrete sets of neurons more selectively, optogenetics affords neuronal systems the potential for improved spatial and temporal resolution.2,3 • To date, no histologic studies have characterized the CN-targeted gene transfer of Chronos, a new generation auditory opsin. • We aim to 1) identify CN regions receptive to gene transfer of Chronos and 2) describe the morphology of putative cell types that express Chronos. 1. Opsin-linked GFP fluorescence demonstrates Chronos expression throughout the injection-targeted dorsal CN with labeling in all 3 CN subdivisions. Chronos expression contiguously extends routinely into the ventral CN and variably on into the auditory nerve. Extension medially into the inferior cerebellar peduncle is also occasionally noted. • The present results demonstrate expression of Chronos in somata and terminals within all layers of the CN, paralleling previous findings of Channelrhodopsin-2 fused to GFP.4 Methods: CBA/CaJ mice underwent CN-targeted injection of Chronos. The Chronos construct consisted of an adeno-associated viral vector (AA2/8), CAG promoter, and a fluorescent marker. Following a 4-week incubation period, mice were sacrificed and intravascularly fixed with paraformaldehyde, and brains were extracted, sucrose cryoprotected, and cryostatically sectioned. Sections of 35-micron thickness were co-labeled with neuron-specific markers microtubule associated protein-2 and antitubulin, beta III isoform and DAPI-fluoromounted. Sections of 60-micron thickness were DAPIfluoromounted, and confocal microscopy revealed cellular morphologies. Results: Opsin-linked fluorescence demonstrates Chronos expression throughout the dorsal CN with contiguous extension routinely into the ventral CN and variably into the auditory nerve and inferior cerebellar peduncle. Chronos localizes to neuronal-specific and non-neuronal populations. Confocal microscopy suggests involvement of a wide array of CN cell types, including morphologies consistent with pyramidal/fusiform, giant, and cartwheel cells. Conclusion: Our histologic analyses confirm widespread infection of multiple neuronal populations throughout the CN. This work sets the stage for correlation with ongoing neurophysiology experiments. Future work with CN-specific promoters to target neuronal subpopulations may further improve clinical potential for an optogenetics-based auditory neuroprosthesis. CONTACT Ashton Lehmann, MD Email: [email protected] Daniel J. Lee, MD, FACS Department of Otolaryngology Massachusetts Eye and Ear Infirmary Email: [email protected] • Chronos is also poised to influence activity in cells crucial to modulating auditory input as inhibitory cartwheel cells shape the output of the dorsal CN in auditory processing.7,9 METHODS & MATERIALS GENE TRANSFER & OPTICAL STIMULATION CBA/CaJ mice (n=4) underwent CN-targeted injection of Chronos via an adeno-associated viral vector (AA2/8) with a CAG promoter and a green fluorescent protein (GFP) marker. Following a 4-week incubation period, blue light (473nm) was delivered to the CN and responses were recorded in upstream regions of the auditory pathway. TISSUE PREPARATION Mice were sacrificed, intravascularly fixed with paraformaldehyde; brains were extracted, sucrose cryoprotected, and cryostatically sectioned. Sections of 35-micron thickness were co-labeled with neuronspecific markers microtubule associated protein-2 (MAP2) and anti-tubulin, beta III isoform (TUJ1) and DAPI-fluoromounted. Sections of 60-micron thickness were DAPI-fluoromounted, and confocal microscopy revealed cellular morphologies. IMMUNOHISTOCHEMISTRY Day1: • Thaw & rehydrate with rinse (PBS x5min x3) • Block/Permeabilize: 1% Triton-X in 5% normal horse serum (NHS) x1hr; then rinse • Primary antibody incubation: 1:500 mouse monoclonal MAP2, TUJ1 with 0.3% Triton-X in 1% NHS, overnight (omit primary for negative controls) Day 2: • Rinse then secondary antibody incubation: 1:1000 goat-anti-mouse Alexafluor 568 with 0.3% Triton-X in 1% NHS x1hr30min; then rinse • Tertiary antibody incubation: 1:1000 donkey-antigoat Alexafluor 568 with 0.3% Triton-X in 1% NHS x1hr30min; then rinse • DAPI-fluoromount, coverslip Inferior Cerebellar Peduncle • Following virally-mediated opsin gene transfer, the locations and numbers of infected cells differ between cases with likely implications for signal quality achieved optogenetically.10,11 1. Colabeling with neuron-specific markers MAP2 and TUJ1 demonstrates localization of Chronos to both neuronal and non-neuronal populations. • Future experimentation with different promoters may enable targeting of specific tissues and neuronal subpopulations to refine the expression of opsins in the auditory system. DAPI Chronos-GFP MAP2-TUJ1 (Neuron) 1. Confocal microscopy enables visualization of complex cellular morphology. Chronos expression involves a wide array of CN cell types, including morphologies consistent with pyramidal/fusiform cells, giant cells, and cartwheel cells. VIII VII nerve root 300 μm CONCLUSION • Histologic characterization of Chronos gene transfer reveals extensive involvement of multiple CN subregions and cell types, laying the groundwork for a new generation of improved auditory neuroprostheses based on optogenetics. REFERENCES IV Ventricle Neutral Red Nissl Stain FUTURE STUDIES • This work sets the stage for correlation with ongoing neurophysiology experiments. Dorsal CN Ventral CN • Chronos localizes to both principal excitatory cell types of the CN: the numerous pyramidal/fusiform cells and the sparse multipolar giant cells receive multimodal input from auditory nerve afferents and parallel fibers and both give off axons which join to form the dorsal acoustic stria, projecting directly upstream to the inferior colliculus.5,6,7,8 Arrow indicates pyramidal/fusiform cell 1. Fu QJ, Nogaki G. Noise susceptibility of cochlear implant users: the role of spectral resolution and smearing. J Assoc Res Otolaryngol. 2005 Mar;6(1):19-27. Epub 2005 Apr 22. 2. Boyden, ES, et al. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci., 2005. 8: p.1263-1268. 3. Chow BY, et al. High-performance genetically targetable optical neural silencing by lightdriven proton pumps. Nature. 2010. 4. Shimano T, et al. Assessment of the AAV-mediated expression of channelrhodopsin-2 and halorhodopsin in brainstem neurons mediating auditory signaling. Brain Research (2013). 1511: 138-152. 5. Oliver DL, Morest DK. The central nucleus of the inferior colliculus in the cat. J. Comp. Neurol. 1984. 222, 237-264. 6. Oertel D, Young ED. What’s a cerebellar circuit doing in the auditory system? Trends Neurosci.2004. 27:104-110. 7. Roberts MT, Trussell LO. Molecular layer inhibitory interneurons provide feedforward and lateral inhibition in the dorsal cochlear nucleus. J Neurophysiol. 2010. 104, 2462-2473. 8. Zhang S, Oertel D. Giant cells of the dorsal cochlear nucleus of mice: intracellular recordings in slices. J Neurophys. 1993. May, 69(5). 9. Golding NL, Oertel D. Context-Dependent Synaptic Action of Glycinergic and GABAergic Inputs in the Dorsal Cochlear Nucleus. J Neurosci. 1996. Apr 1, 16(7):2208-2219. 10. Darrow K, et al. Optogenetic stimulation of the cochlear nucleus using channelrhodopsin-2 evokes activity in the central auditory pathway Brain Research, Submitted. 2014. 11. Young ED. Response characteristics of neurons of the cochlear nuclei. In: Berlin, C.I., (Ed.), Hearing Science: Recent Advances. College-Hill Press, San Diego. 1984. pp. 423-460.